
Peripheral Modulators of the Central Fatigue Development and Their Relationship with Athletic Performance in Jumper Horses

 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Rectal Temperature (RT) and Heart Rate (HR) Measurement
2.3. Blood Sampling Procedures and Laboratory Analysis
2.4. Statistical Analysis
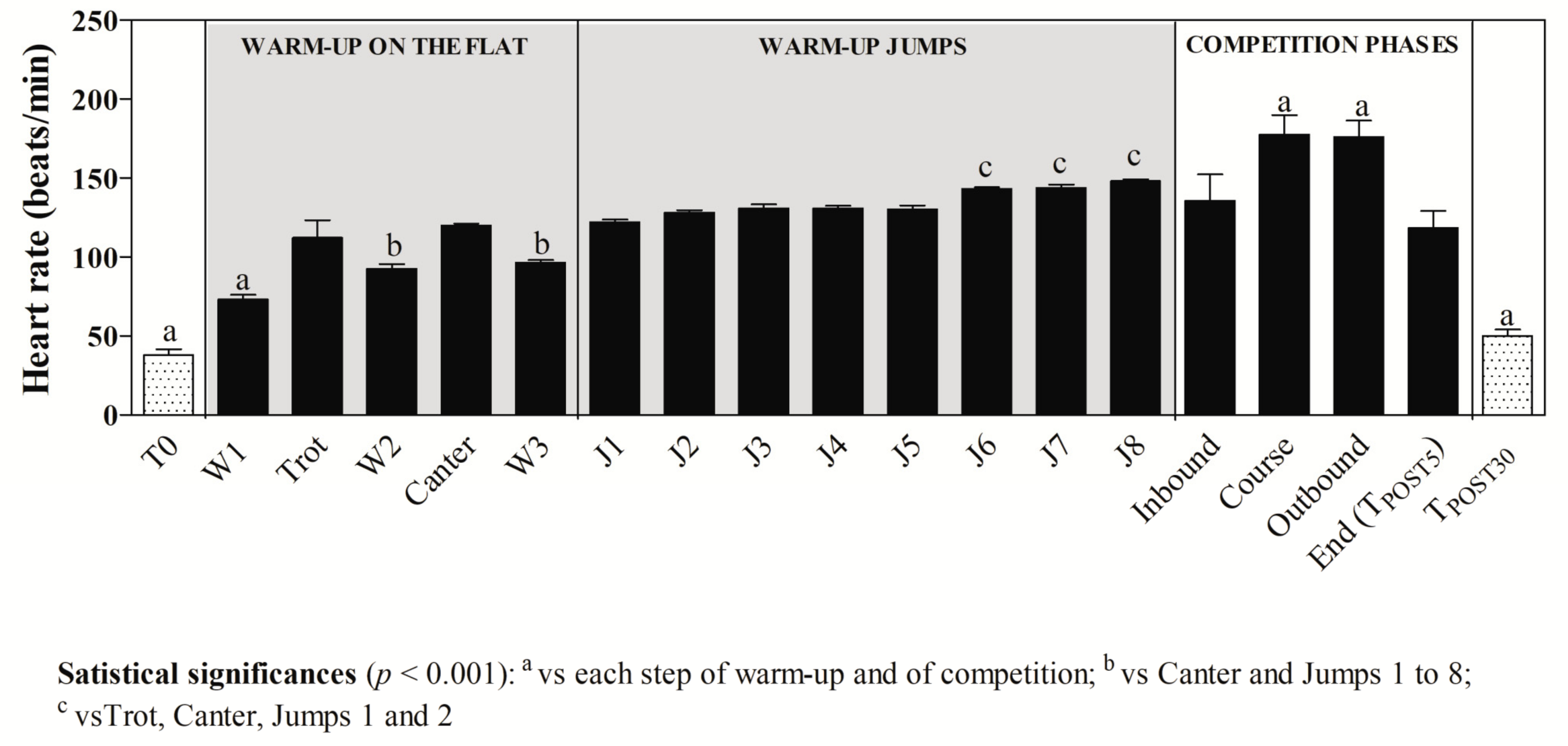
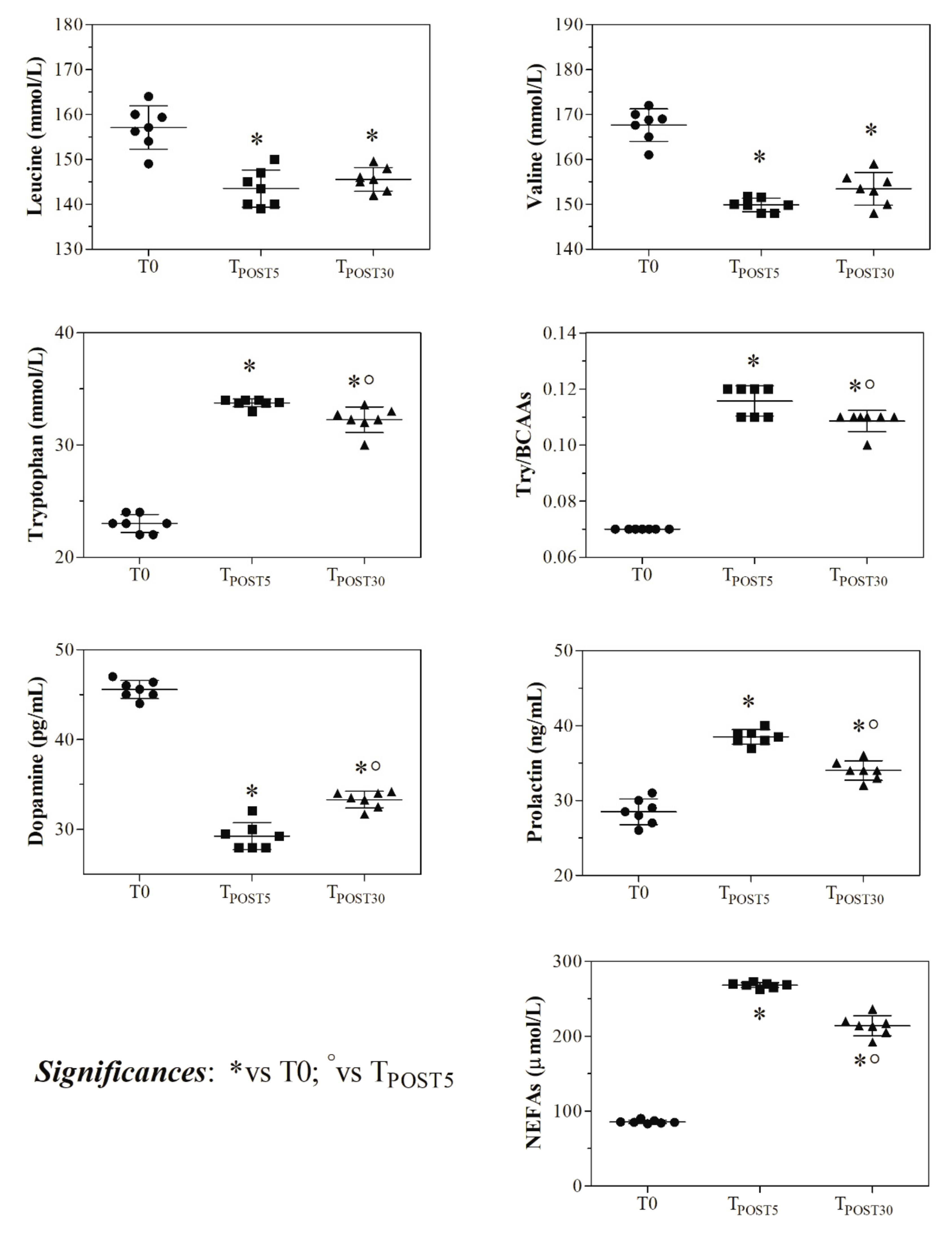
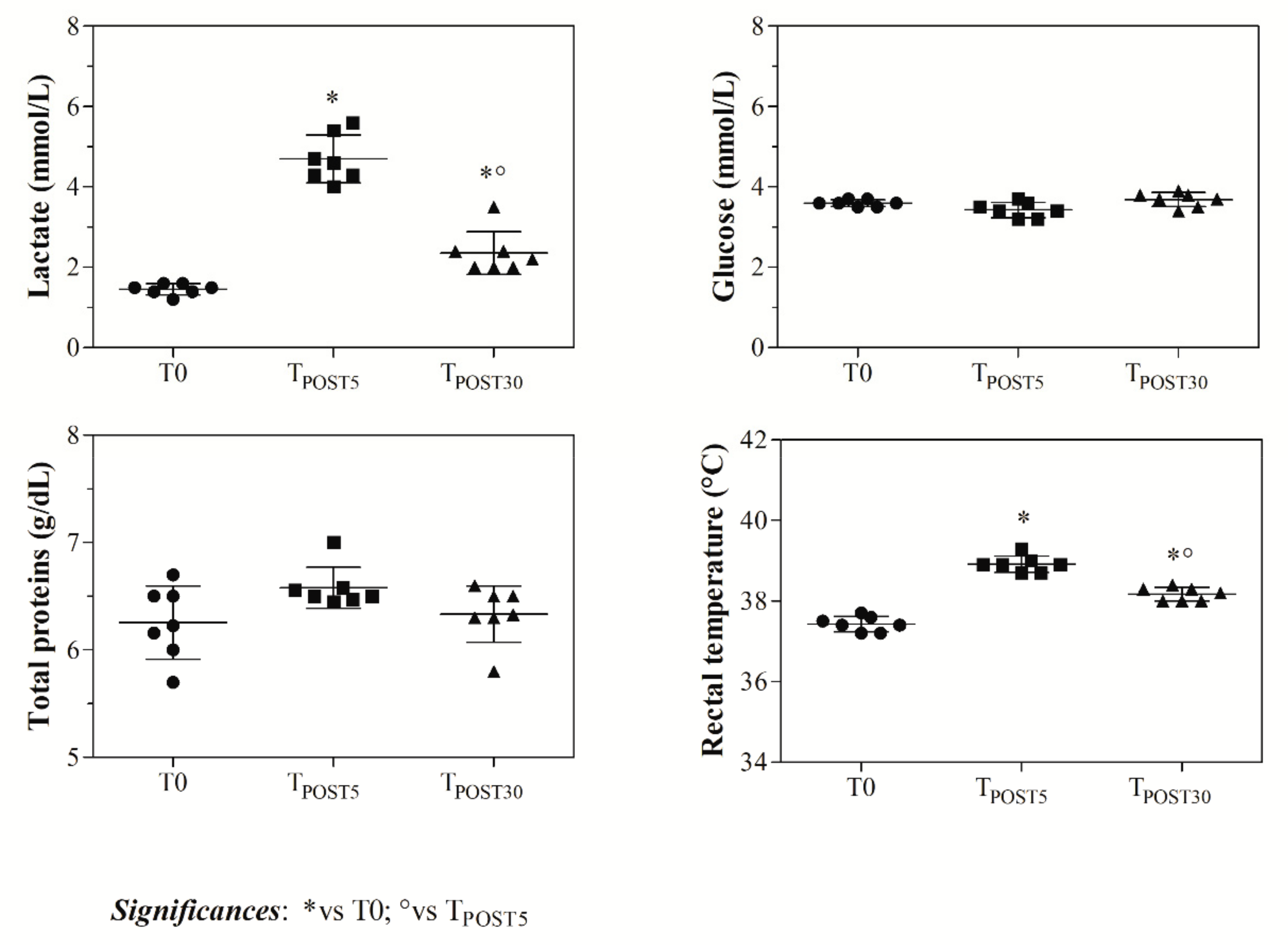
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hodgson, D.R.; McGowan, C.; McKeever, K. The Athletic Horse: Principles and Practice of Equine Sport Medicine, 2nd ed.; W.B. Saunders: St. Louis, MO, USA, 2014. [Google Scholar]
- Witkowska-Piłaszewicz, O.; Pingwara, R.; Winnicka, A. The effect of physical training on peripheral blood mononuclear cell ex vivo proliferation, differentiation, activity, and reactive oxygen species production in racehorses. Antioxidants (Basel) 2020, 9, 1155. [Google Scholar] [CrossRef]
- Witkowska-Piłaszewicz, O.; Bąska, P.; Czopowic, M.; Żmigrodzka, M.; Szarska, E.; Szczepaniak, J.; Nowak, Z.; Winnicka, A.; Cywińska, A. Anti-inflammatory state in Arabian horses introduced to the endurance training. Animals (Basel) 2019, 9, 616. [Google Scholar] [CrossRef] [Green Version]
- Munster, C.C.; van Iwaarden, A.; van Weeren, R.; Sloet van Oldruitenborgh-Oosterbaan, M.M. Exercise testing in Warmblood sport horses under field conditions. Vet. J. 2014, 202, 11–19. [Google Scholar] [CrossRef]
- Williams, R.J.; Chandler, R.E.; Marlin, D.J. Heart rate of horses during competitive dressage. Comp. Exerc. Physiol. 2009, 6, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Bazzano, M.; Giudice, E.; Rizzo, M.; Congiu, F.; Zumbo, A.; Arfuso, F.; Di Pietro, S.; Bruschetta, D.; Piccione, G. Application of a combined global positioning and heart rate monitoring system in jumper horses during an official competition—A preliminary study. Acta Vet. Hung. 2016, 64, 189–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newsholm, E.A. Application of principles of metabolic control to the problem of metabolic limitations in sprinting, middledistance, and marathon running. Int. J. Sports Med. 1986, 7, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Arfuso, F.; Assenza, A.; Fazio, F.; Rizzo, M.; Giannetto, C.; Piccione, G. Dynamic change of serum levels of some branched-chain amino acids and tryptophan in athletic horses after different physical exercises. J. Equine Vet. Sci. 2019, 77, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Edwards, R.H.T. Human muscle function and fatigue. In Human Muscle Fatigue: Physiological Mechanisms; Porter, R., Whelan, J., Eds.; Ciba Foundation Symposium 82; Pitman Medical: London, UK, 1981; pp. 1–18. [Google Scholar]
- Newsholme, E.A.; Acworth, I.N.; Blomstrad, E. Amino acids, brain neurotransmitters and a functional link between muscle and brain that is important in sustained exercise. In Advances in Myochemistry; Benzi, G., Ed.; John Libby Eurotext: London, UK, 1987; pp. 127–138. [Google Scholar]
- Davis, J.M.; Alderson, N.L.; Welsh, R.S. Serotonin and central nervous system fatigue: Nutritional considerations. Am. J. Clin Nutr. 2000, 72, 573S–578S. [Google Scholar] [CrossRef] [Green Version]
- Chaouloff, F. Physical exercise and brain monoamines: A review. Acta Physiol. Scand. 1989, 137, 1–13. [Google Scholar] [CrossRef]
- Fernstrom, J.D. Aromatic amino acids and monoamine synthesis in the central nervous system: Influence of the diet. J. Nutr. Biochem. 1990, 1, 508–517. [Google Scholar] [CrossRef]
- Dalsgaard, M.; Ide, K.; Cai, Y.; Quistorff, B.; Secher, N.H. The intent to exercise influences the cerebral O2/carbohydrate uptake ratio in humans. J. Physiol. 2002, 540, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Dalsgaard, M.; Nybo, L.; Cai, Y.; Secher, N.H. Cerebral metabolism is influenced by muscle ischaemia during exercise in humans. Exp. Physiol. 2003, 88, 297–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greer, B.K.; White, J.P.; Arguello, E.M.; Haymes, E.M. Branched-chain amino acid supplementation lowers perceived exertion but does not affect performance in untrained males. J. Strength. Cond. Res. 2011, 25, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.M.; Bailey, S.P. Possible mechanisms of central nervous system fatigue during exercise. Med. Sci. Sports. Exerc. 1997, 29, 45–57. [Google Scholar] [CrossRef]
- Rojas Vega, S.; Hollmann, W.; Strüder, H.K. Influences of exercise and training on the circulating concentration of prolactin in humans. J. Neuroendocrinol. 2011, 24, 395–402. [Google Scholar] [CrossRef]
- Hackney, A.C. Characterization of the prolactin response to prolonged endurance exercise. Acta Kinesiol. Univ. Tartu. 2008, 13, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Melis, F.; Crisafulli, A.; Rocchitta, A.; Tocco, F.; Concu, A. Does reduction of blood prolactin levels reveal the activation of central dopaminergic pathways conveying reward in top athletes? Med. Hypotheses 2003, 61, 133–135. [Google Scholar] [CrossRef]
- Sutoo, D.; Akiyama, K. Regulation of brain function by exercise. Neurobiol. Dis. 2003, 13, 1–14. [Google Scholar] [CrossRef]
- Chaouloff, F.; Laude, D.; Merino, D. Amphetamine and alfa-methyl-p-tyrosine affect the exercise induced imbalance between the availability of tryptophan and synthesis of serotonin in the brain of the rat. Neuropharmacology 1987, 26, 1099–1106. [Google Scholar] [CrossRef]
- Freed, C.; Yamamoto, B. Regional brain dopamine metabolism: A marker for the speed, direction and posture of moving animals. Science 1985, 229, 62–65. [Google Scholar] [CrossRef] [PubMed]
- Lekeux, P.; Art, T.; Linden, A.; Desmecht, D.; Amory, H. Heart rate, haematological and serum biochemical responses to show jumping. Equine Exerc. Physiol. 1991, 3, 385–390. [Google Scholar]
- Art, T.; Amory, H.; Desmecht, D.; Lekeux, P. Effect of show jumping on heart rate, blood lactate and other plasma biochemical values. Equine Vet. J. Suppl. 1990, 9, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Clayton, H.M. Show Jumping. In Conditioning Sport Horses; Clayton, H.M., Ed.; Sport horse publications: Saskatoon, Saskatchewan, 1991; pp. 173–180. [Google Scholar]
- Loving, N.S.; Johnston, A.M. Veterinary Manual for the Performance Horse; Blackwell Science Ltd.: London, UK, 1995. [Google Scholar]
- Rietmann, T.R.; Stuart, A.E.A.; Bernasconi, P.; Stauffacher, M.; Auer, J.A.; Weishaupt, M.A. Assessment of mental stress in warmblood horses: Heart rate variability in comparison to heart rate and selected behavioural parameters. Appl. Anim. Behav. Sci. 2004, 88, 121–136. [Google Scholar] [CrossRef]
- Jansen, F.; Van der Krogt, J.; Van Loon, K.; Avezzù, V.; Guarino, M.; Quanten, S.; Berckmans, D. Online detection of an emotional response of a horse during physical activity. Vet. J. 2009, 181, 38–42. [Google Scholar] [CrossRef]
- Hinchcliff, K.W.; Kaneps, A.J.; Geor, R.J. Equine Sports Medicine and Surgery; Saunders: London, UK, 2004. [Google Scholar]
- Rizzo, M.; Arfuso, F.; Giudice, E.; Abbate, F.; Longo, F.; Piccione, G. Core and surface temperature modification during road transport and physical exercise in horse after acupuncture needle stimulation. J. Equine Vet. Sci. 2017, 55, 84–89. [Google Scholar] [CrossRef]
- Feldberg, W.; Myers, R.D. Effects on temperature of amines injected into the cerebral ventricles. A new concept of temperature regulation. J. Physiol. 1964, 173, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Struder, H.K.; Weicker, H. Physiology and pathophysiology of the serotonergic system and its implications on mental and physical performance. Part II. Int. J. Sports Med. 2001, 22, 482–497. [Google Scholar] [CrossRef] [PubMed]
- Arfuso, F.; Giannetto, C.; Giudice, E.; Fazio, F.; Piccione, G. Dynamic modulation of platelet aggregation, albumin and nonesterified fatty acids during physical exercise in thoroughbred horses. Res. Vet. Sci. 2016, 104, 86–91. [Google Scholar] [CrossRef]
- Witkowska-Piłaszewicz, O.; Maśko, M.; Domino, M.; Winnicka, A. Infrared thermography correlates with lactate concentration in blood during race training in horses. Animals (Basel) 2020, 10, 2072. [Google Scholar] [CrossRef]
- Blomstrand, E. A role for branched-chain amino acids in reducing central fatigue. J. Nutr. 2006, 136, S544–S547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaouloff, F.; Laude, D.; Elghozi, J.L. Physical exercise: Evidence for differential consequences of tryptophan on 5-HT synthesis and metabolism in central serotonergic cell bodies and terminals. J. Neural. Transm. 1989, 78, 121–130. [Google Scholar] [CrossRef]
- Piccione, G.; Assenza, A.; Fazio, F.; Percipalle, M.; Caola, G. Central fatigue and nycthemeral change of serum tryptophan and serotonin in the athletic horse. J. Circadian Rhythm. 2005, 3, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Alberghina, D.; Giannetto, C.; Piccione, G. Peripheral serotoninergic response to physical exercise in athletic horses. J. Vet. Sci. 2010, 11, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Ben-Jonathan, N.; Hnasko, R. Dopamine as a prolactin (PRL) inhibitor. Endocr. Rev. 2001, 22, 724–763. [Google Scholar] [CrossRef]
- Hardebo, J.; Owman, C. Barrier mechanisms for neurotransmitter monoamines and their precursors at the blood-brain interface. Ann. Neurol. 1980, 8, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Freeman, M.; Kanyicska, B.; Lerant, A.; Nagy, G. Prolactin: Structure, function, and regulation of secretion. Physiol. Rev. 2000, 80, 1523–1631. [Google Scholar] [CrossRef] [PubMed]
- Ferrier, C.; Jennings, G.L.; Eisenhofer, G.; Lambert, G.; Cox, H.S. Evidence for increased noradrenalin release from subcortical brain regions in essential hypertension. J. Hypertens. 1993, 11, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Hardebo, J.; Owman, C. Characterization of the in vitro uptake of monoamines into brain microvessels. Acta Physiol. Scand. 1980, 108, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Podolak, M.; Kedzierski, W.; Bergero, D. Comparison of the blood plasma catecholamines level in thoroughbred and Arabian horses during the same-intensity exercise. Pol. J. Vet. Sci. 2006, 9, 71–73. [Google Scholar]
- Rothman, R.B.; Clark, R.D.; Partilla, J.S.; Baumann, M.H. (+)-Fenfluramine and its major metabolite, (þ)-norfenfluramine, are potent substrates for norepinephrine transporters. J. Pharmacol. Exp. Ther. 2003, 305, 1191–1199. [Google Scholar] [CrossRef] [Green Version]
- Gordon, J.; Barnes, N.M. Lymphocytes transport serotonin and dopamine: Agony or ecstasy? Trends Immunol. 2003, 24, 438–443. [Google Scholar] [CrossRef]
- Di Giovanni, L.C.; Thompson, D.L.; Valencia, N.A.; Oberhaus, E.L. Factor affecting basal and post-exercise prolactin secretion in horse. J. Vet. Sci. 2015, 35, 437–445. [Google Scholar]
- Strüder, H.K.; Hollmann, W.; Platen, P.; Wöstmann, R.; Ferrauti, A.; Weber, K. Effect of exercise intensity on free tryptophan to branched-chain amino acids ratio and plasma prolactin during endurance exercise. Can. J. Appl. Physiol. 1997, 22, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H.G.; Hollmann, W.; De Meirleir, K. Exercise changes in plasma tryptophan fractions and relationship with prolactin. Int. J. Sports Med. 1991, 12, 487–489. [Google Scholar] [CrossRef] [PubMed]
- Struder, H.; Hollmann, W.; Platen, P.; Duperly, J.; Fischer, H.; Weber, K. Alterations in plasma free tryptophan and large neutral amino acids do not affect perceived exertion and prolactin during 90 min of treadmill exercise. Int. J. Sports Med. 1996, 17, 73–79. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Parameters- | Try/BCAAs | Prolactin (ng/mL) | Dopamine (pg/mL) | NEFAs (μmol/L) | HR (beats/min) | RT (°C) | Lactate (mmol/L) | Glucose (mmol/L) |
|---|---|---|---|---|---|---|---|---|
| Tryptophan (mmol/L) | r = 0.99 p < 0.0001 | r = −0.26 p = 0.29 | r = −0.97 p < 0.0001 | r = 0.75 p < 0.0001 | r = 0.69 p = 0.002 | r = 0.88 p < 0.0001 | r = 0.75 p < 0.0001 | r = −0.26 p = 0.29 |
| Leucine (mmol/L) | r = −0.84 p < 0.0001 | r = −0.26 p = 0.29 | r = 0.79 p = 0.0001 | r = 0.75 p < 0.0001 | r = −0.57 p = 0.01 | r = −0.66 p = 0.002 | r = −0.69 p = 0.001 | r = 0.5 p = 0.84 |
| Valine (mmol/L) | r = −0.90 p < 0.0001 | r = −0.26 p = 0.29 | r = 0.89 p < 0.0001 | r = 0.75 p < 0.0001 | r = −0.69 p = 0.001 | r = −0.81 p < 0.0001 | r = −0.68 p = 0.002 | r = 0.09 p = 0.70 |
| Try/BCAAs | - | r = 0.89 p < 0.0001 | r = −0.97 p < 0.0001 | r = 0.97 p < 0.0001 | r = 0.69 p = 0.002 | r = 0.88 p < 0.0001 | r = 0.75 p < 0.0001 | r = −0.26 p = 0.29 |
| Prolactin (ng/mL) | r = 0.89 p < 0.0001 | - | r = −0.91 p < 0.0001 | r = 0.93 p < 0.0001 | r = 0.86 p < 0.0001 | r = 0.92 p < 0.0001 | r = 0.87 p < 0.0001 | r = −0.36 p = 0.014 |
| Dopamine (pg/mL) | r = −0.97 p < 0.0001 | r = −0.91 p < 0.0001 | - | r = −0.99 p < 0.0001 | r = −0.78 p = 0.0001 | r = −0.93 p < 0.0001 | r = −0.80 p < 0.0001 | r = 0.27 p = 0.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arfuso, F.; Giannetto, C.; Giudice, E.; Fazio, F.; Panzera, M.; Piccione, G. Peripheral Modulators of the Central Fatigue Development and Their Relationship with Athletic Performance in Jumper Horses. Animals 2021, 11, 743. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11030743
Arfuso F, Giannetto C, Giudice E, Fazio F, Panzera M, Piccione G. Peripheral Modulators of the Central Fatigue Development and Their Relationship with Athletic Performance in Jumper Horses. Animals. 2021; 11(3):743. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11030743
Chicago/Turabian StyleArfuso, Francesca, Claudia Giannetto, Elisabetta Giudice, Francesco Fazio, Michele Panzera, and Giuseppe Piccione. 2021. "Peripheral Modulators of the Central Fatigue Development and Their Relationship with Athletic Performance in Jumper Horses" Animals 11, no. 3: 743. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11030743