
Reproductive Toxicity of 3,4-dichloroaniline (3,4-DCA) on Javanese Medaka (Oryziasjavanicus, Bleeker 1854)

 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Source of Chemicals
2.2. Javanese Medaka Maintenance
2.3. Selection of Reproductively Active Adults
2.4. Preparation of Test Chemicals and Exposure
2.5. Morphometric and Somatic Indices
2.6. Fecundity
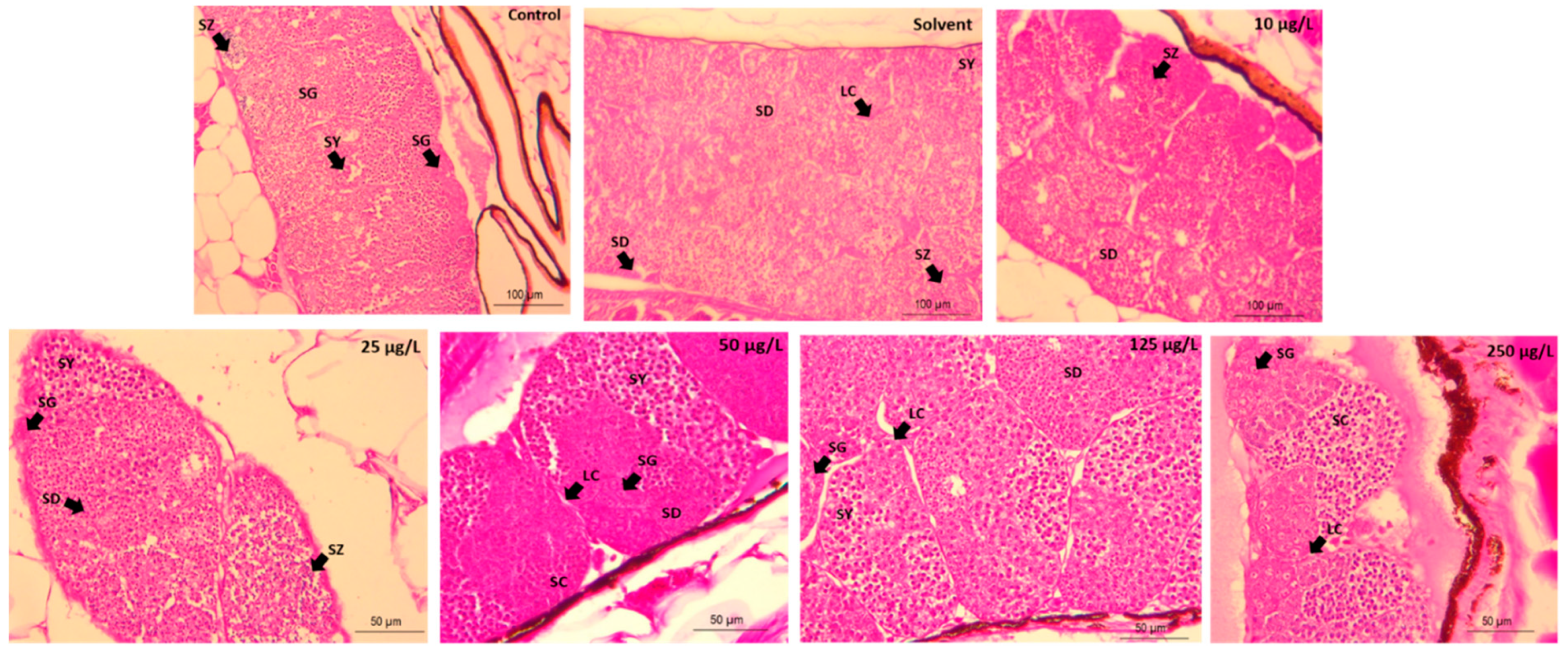
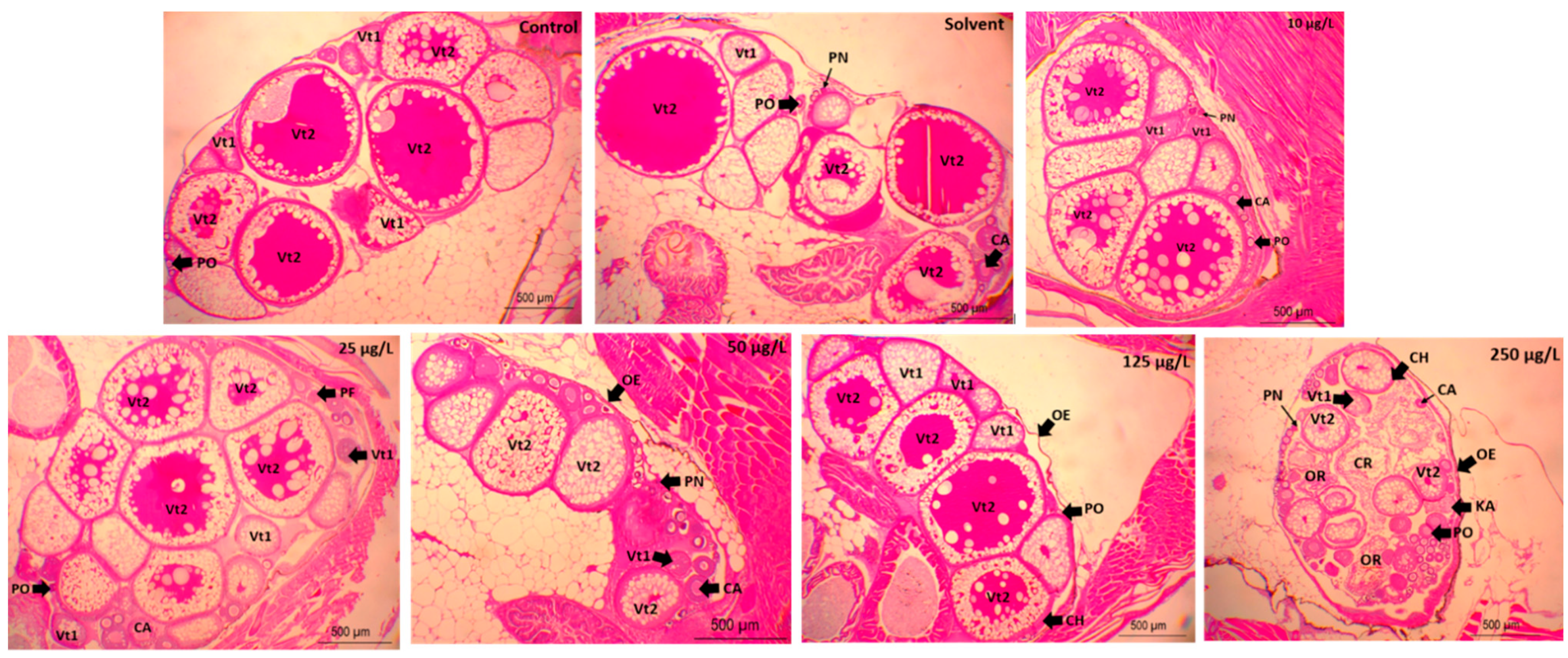
2.7. Gonad Histopathology
2.8. Statistical Analysis
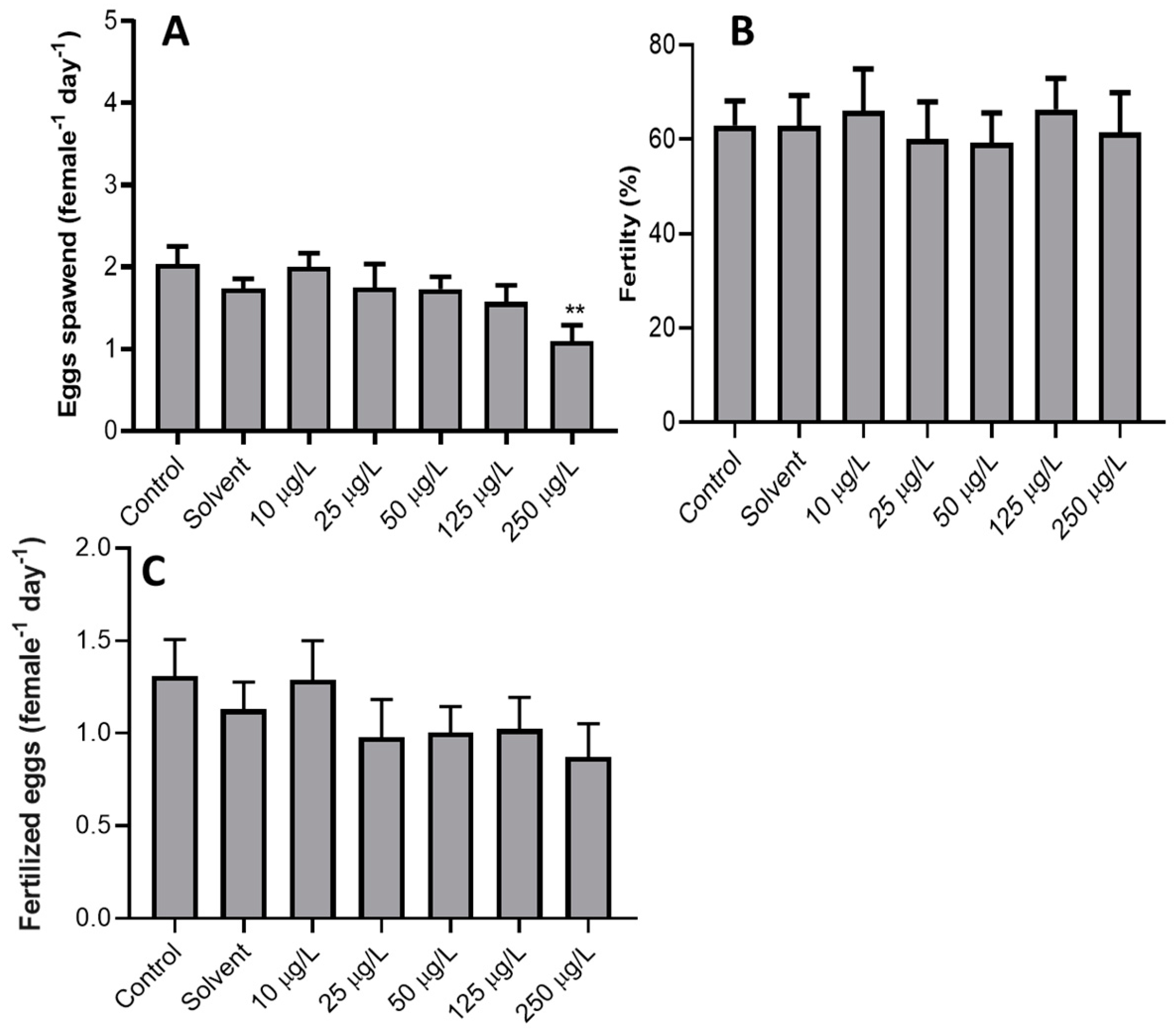
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Philippe, C.; Grégoir, A.F.; Thoré, E.S.J.; Brendonck, L.; De Boeck, G.; Pinceel, T. Acute sensitivity of the killifish Nothobranchius furzeri to a combination of temperature and reference toxicants (cadmium, chlorpyrifos and 3,4-dichloroaniline). Environ. Sci. Pollut. Res. 2018, 25, 10029–10038. [Google Scholar] [CrossRef]
- Eissa, F.I.; El Makawy, A.I.; Badr, M.I.; Elhamalawy, O.H. Assessment of 3,4-Dichloroaniline Toxicity as Environmental Pollutant in Male Mice. Eur. J. Biol. Sci. 2012, 4, 73–82. [Google Scholar] [CrossRef]
- Bhuiyan, M.N.H.; Kang, H.; Kim, J.H.; Kim, S.; Kho, Y.; Choi, K. Endocrine disruption by several aniline derivatives and related mechanisms in a human adrenal H295R cell line and adult male zebrafish. Ecotoxicol. Environ. Saf. 2019, 180, 326–332. [Google Scholar] [CrossRef]
- Carena, L.; Proto, M.; Minella, M.; Ghigo, G.; Giovannoli, C.; Brigante, M.; Mailhot, G.; Maurino, V.; Minero, C.; Vione, D. Evidence of an Important Role of Photochemistry in the Attenuation of the Secondary Contaminant 3,4-Dichloroaniline in Paddy Water. Environ. Sci. Technol. 2018, 52, 6334–6342. [Google Scholar] [CrossRef]
- Tasca, A.L.; Fletcher, A. State of the art of the environmental behaviour and removal techniques of the endocrine disruptor 3,4-dichloroaniline. J. Environ. Sci. Health Part A 2018, 53, 260–270. [Google Scholar] [CrossRef]
- Philippe, C.; Hautekiet, P.; Grégoir, A.F.; Thoré, E.S.; Brendonck, L.; De Boeck, G.; Pinceel, T. Interactive effects of 3,4-DCA and temperature on the annual killifish Nothobranchius furzeri. Aquat. Toxicol. 2019, 212, 146–153. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Zulkifli, S.Z.; Azmai, M.N.A.; Mohamat-Yusuff, F.; Ismail, A. Embryonic toxicity of 3,4-dichloroaniline (3,4-DCA) on Javanese medaka (Oryzias javanicus Bleeker, 1854). Toxicol. Rep. 2020, 7, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Vieira, L.R.; Hissa, D.C.; De Souza, T.M.; Sá, C.A.; Evaristo, J.A.M.; Nogueira, F.C.S.; Carvalho, A.F.U.; Farias, D.F. Proteomics analysis of zebrafish larvae exposed to 3,4-dichloroaniline using the fish embryo acute toxicity test. Environ. Toxicol. 2020, 35, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Zhan, P.; Gan, N.; Cao, Y.; Li, T.; Sang, W.; Wang, L. Highly Selective Molecular Recognition and Ultrasensitive Detection of 3,4-dichloroaniline Based on Molecularly Imprinted Sol-Gel Film Combined with Multi-Walled Carbon Nanotubes. J. Electrochem. Soc. 2013, 160, H742–H748. [Google Scholar] [CrossRef]
- Qu, X.; Yan, B. Zn(ii)/Cd(ii)-based metal–organic frameworks: Crystal structures, Ln(iii)-functionalized luminescence and chemical sensing of dichloroaniline as a pesticide biomarker. J. Mater. Chem. C 2020, 8, 9427–9439. [Google Scholar] [CrossRef]
- Xiao, H.; Kuckelkorn, J.; Nüßer, L.K.; Floehr, T.; Hennig, M.P.; Roß-Nickoll, M.; Schäffer, A.; Hollert, H. The metabolite 3,4,3′,4′-tetrachloroazobenzene (TCAB) exerts a higher ecotoxicity than the parent compounds 3,4-dichloroaniline (3,4-DCA) and propanil. Sci. Total Environ. 2016, 551–552, 304–316. [Google Scholar] [CrossRef]
- Pinto, P.I.S.; Estêvão, M.D.; Power, D.M. Effects of Estrogens and Estrogenic Disrupting Compounds on Fish Mineralized Tissues. Mar. Drugs 2014, 12, 4474–4494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellock, K.A.; Moore, A.P.; Bringolf, R.B. Chronic nitrate exposure alters reproductive physiology in fathead minnows. Environ. Pollut. 2018, 232, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Liang, M.; Chen, R.; Hong, X.; Zha, J. Reproductive toxicity and estrogen activity in Japanese medaka (Oryzias latipes) exposed to environmentally relevant concentrations of octocrylene. Environ. Pollut. 2020, 261, 114104. [Google Scholar] [CrossRef]
- Bai, J.; Han, H.; Wang, F.; Su, L.; Ding, H.; Hu, X.; Hu, B.; Li, H.; Zheng, W.; Li, Y. Maternal linuron exposure alters testicular development in male offspring rats at the whole genome level. Toxicology 2017, 389, 13–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, T.S.B.; Boscolo, C.N.P.; Felício, A.A.; Batlouni, S.R.; Schlenk, D.; De Almeida, E.A. Estrogenic activities of diuron metabolites in female Nile tilapia (Oreochromis niloticus). Chemosphere 2016, 146, 497–502. [Google Scholar] [CrossRef] [Green Version]
- Frankel, T.E.; Meyer, M.T.; Kolpin, D.W.; Gillis, A.B.; Alvarez, D.A.; Orlando, E.F. Exposure to the Contraceptive Progestin, Gestodene, Alters Reproductive Behavior, Arrests Egg Deposition, and Masculinizes Development in the Fathead Minnow (Pimephales promelas). Environ. Sci. Technol. 2016, 50, 5991–5999. [Google Scholar] [CrossRef] [PubMed]
- Brehm, E.; A Flaws, J. Transgenerational Effects of Endocrine-Disrupting Chemicals on Male and Female Reproduction. Endocrinology 2019, 160, 1421–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, T.S.B.; Boscolo, C.N.P.; Da Silva, D.G.H.; Batlouni, S.R.; Schlenk, D.; De Almeida, E.A. Anti-androgenic activities of diuron and its metabolites in male Nile tilapia (Oreochromis niloticus). Aquat. Toxicol. 2015, 164, 10–15. [Google Scholar] [CrossRef]
- Yusof, S.; Ismail, A.; Alias, M.S. Effect of glyphosate-based herbicide on early life stages of Java medaka (Oryzias javanicus): A potential tropical test fish. Mar. Pollut. Bull. 2014, 85, 494–498. [Google Scholar] [CrossRef] [PubMed]
- Aziz, Z.; Zulkifli, S.Z.; Mohamat-yusuff, F.; Azmai, M.N.A.; Ismail, A. An Histological Study on Mercury-Induced Gonadal Impairment in Javanese Medaka (Oryzias javanicus) An Histological Study on Mercury-Induced Gonadal Impairment. Turk. J. Fish. Aquat. Sci. 2017, 17, 621–627. [Google Scholar] [CrossRef]
- Salleh, A.F.M.; Amal, M.N.A.; Nasruddin, N.S.; Zulkifli, S.Z.; Yusuff, F.M.; Ibrahim, W.N.W.; Ismail, A. Water pH effects on survival, reproductive performances, and ultrastructure of gonads, gills, and skins of the Javanese medaka (Oryzias javanicus). Turk. J. Vet. Anim. Sci. 2017, 41, 471–481. [Google Scholar] [CrossRef]
- Amal, M.N.A.; Roseli, M.F.A.M.; Sutra, J.; Roslan, S.; Mazlan, S.N.K.; Basri, L.; Yie, L.J.; Mohamad, N.; Ismail, A.; Saad, M.Z.; et al. Javanese Medaka (Oryzias javanicus Bleeker, 1854) as Potential Model Organism for Aeromoniasis and Vibriosis Study in Fish. Pertanika J. Trop. Agric. Sci. 2019, 42, 1049–1065. [Google Scholar]
- Rusni, S.; Sassa, M.; Takehana, Y.; Kinoshita, M.; Inoue, K. Correlation between cytochrome P450 1A (cyp1a) mRNA expression and ambient phenanthrene and pyrene concentration in Javanese Medaka Oryzias javanicus. Fish. Sci. 2020, 86, 605–613. [Google Scholar] [CrossRef]
- Lee, B.-Y.; Park, J.C.; Kim, M.-S.; Choi, B.-S.; Kim, D.-H.; Lim, J.-S.; Yum, S.; Hwang, U.-K.; Nah, G.J.; Lee, J.-S. The genome of the Java medaka (Oryzias javanicus): Potential for its use in marine molecular ecotoxicology. Mar. Pollut. Bull. 2020, 154, 111118. [Google Scholar] [CrossRef]
- Imai, S.; Koyama, J.; Fujii, K. Effects of 17β-estradiol on the reproduction of Java-medaka (Oryzias javanicus), a new test fish species. Mar. Pollut. Bull. 2005, 51, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Sardo, M.; Azeiteiro, U.M.; Pereira, L.; Morgado, F.; Soares, M. Histological evaluation of the exposure to 3,4-dichloroaniline in the estuarine mysid mesopodopsis slabberi, under experimental conditions. Fresenius Environ. Bull. 2005, 14, 579–583. [Google Scholar]
- Bhuiyan, N.H.; Kang, H.; Choi, J.; Lim, S.; Kho, Y.; Choi, K. Effects of 3,4-dichloroaniline (3,4-DCA) and 4,4′-methylenedianiline (4,4′-MDA) on sex hormone regulation and reproduction of adult zebrafish (Danio rerio). Chemosphere 2021, 269, 128768. [Google Scholar] [CrossRef]
- Li, W.-M.; Yin, D.-Q.; Zhou, Y.; Hu, S.-Q.; Wang, L.-S. 3,4-Dichloroaniline-induced oxidative stress in liver of crucian carp (Carassius auratus). Ecotoxicol. Environ. Saf. 2003, 56, 251–255. [Google Scholar] [CrossRef]
- Boscolo, C.N.P.; Pereira, T.S.B.; Batalhão, I.G.; Dourado, P.L.R.; Schlenk, D.; De Almeida, E.A. Diuron metabolites act as endocrine disruptors and alter aggressive behavior in Nile tilapia (Oreochromis niloticus). Chemosphere 2018, 191, 832–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OECD Test No. 229: Fish short term reproduction assay. In OECD Guidelines for the Testing of Chemicals, Section 2; Organisation for Economic Development and Coorporation: Paris, France, 2012; Volume 40. [CrossRef]
- Naderi, M.; Wong, M.Y.; Gholami, F. Developmental exposure of zebrafish (Danio rerio) to bisphenol-S impairs subsequent reproduction potential and hormonal balance in adults. Aquat. Toxicol. 2014, 148, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Wu, D.; Dang, Y.; Yu, L.; Liu, C.; Wang, J. Reproduction impairment and endocrine disruption in adult zebrafish (Danio rerio) after waterborne exposure to TBOEP. Aquat. Toxicol. 2017, 182, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Ke, M.; Fan, X.; Zhang, M.; Zhu, Y.; Lu, T.; Sun, L.; Qian, H. Reproductive and endocrine-disrupting toxicity of Microcystis aeruginosa in female zebrafish. Chemosphere 2018, 192, 289–296. [Google Scholar] [CrossRef]
- Luzio, A.; Monteiro, S.M.; Rocha, E.; Fontaínhas-Fernandes, A.A.; Coimbra, A.M. Development and recovery of histopathological alterations in the gonads of zebrafish (Danio rerio) after single and combined exposure to endocrine disruptors (17α-ethinylestradiol and fadrozole). Aquat. Toxicol. 2016, 175, 90–105. [Google Scholar] [CrossRef] [PubMed]
- Carvan, M. Fish Models in Toxicology. Zebrafish 2007, 4, 9–20. [Google Scholar] [CrossRef]
- Martin, O.V.; Voulvoulis, N. Sustainable risk management of emerging contaminants in municipal wastewaters. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2009, 367, 3895–3922. [Google Scholar] [CrossRef] [Green Version]
- Glinka, C.O.; Frasca, S.; Provatas, A.A.; Lama, T.; DeGuise, S.; Bosker, T. The effects of model androgen 5α-dihydrotestosterone on mummichog (Fundulus heteroclitus) reproduction under different salinities. Aquat. Toxicol. 2015, 165, 266–276. [Google Scholar] [CrossRef]
- Watanabe, H.; Horie, Y.; Takanobu, H.; Koshio, M.; Flynn, K.; Iguchi, T.; Tatarazako, N. Medaka extended one-generation reproduction test evaluating 4-nonylphenol. Environ. Toxicol. Chem. 2017, 36, 3254–3266. [Google Scholar] [CrossRef]
- Crossland, N.O.; Hillaby, J.M. Fate and Effects of 3,4-Dichloroaniline in the Laboratory and in Outdoor Ponds: Ii. Chronic Toxicity to Daphnia Spp. and Other Invertebrates. Environ. Toxicol. Chem. 1985, 4, 489. [Google Scholar] [CrossRef]
- Carnevali, O.; Santangeli, S.; Forner-Piquer, I.; Basili, D.; Maradonna, F. Endocrine-disrupting chemicals in aquatic environment: What are the risks for fish gametes? Fish Physiol. Biochem. 2018, 44, 1561–1576. [Google Scholar] [CrossRef]
- Carbajal-Hernández, A.L.; Valerio-García, R.C.; Martínez-Ruíz, E.B.; Jarquín-Díaz, V.H.; Martínez-Jerónimo, F. Maternal-embryonic metabolic and antioxidant response of Chapalichthys pardalis (Teleostei: Goodeidae) induced by exposure to 3,4-dichloroaniline. Environ. Sci. Pollut. Res. 2017, 24, 17534–17546. [Google Scholar] [CrossRef]
- Imai, S.; Koyama, J.; Fujii, K. Effects of estrone on full life cycle of Java medaka (Oryzias javanicus), a new marine test fish. Environ. Toxicol. Chem. 2007, 26, 726–731. [Google Scholar] [CrossRef]
- Ankley, G.T.; Jensen, K.M. A novel framework for interpretation of data from the fish short-term reproduction assay (FSTRA) for the detection of endocrine-disrupting chemicals. Environ. Toxicol. Chem. 2014, 33, 2529–2540. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Takei, Y. Diverse Adaptability in Oryzias Species to High Environmental Salinity. Zool. Sci. 2002, 19, 727–734. [Google Scholar] [CrossRef]
- Cao, Q.; Gu, J.; Wang, D.; Liang, F.; Zhang, H.; Li, X.; Yin, S. Physiological mechanism of osmoregulatory adaptation in anguillid eels. Fish Physiol. Biochem. 2018, 44, 423–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilannejad, N.; Dorafshan, S.; Heyrati, F.P.; Soofiani, N.M.; Asadollah, S.; Martos-Sitcha, J.A.; Prat, F.; Yúfera, M.; Martínez-Rodríguez, G. Vitellogenin expression in wild cyprinid Petroleuciscus esfahani as a biomarker of endocrine disruption along the Zayandeh Roud River, Iran. Chemosphere 2016, 144, 1342–1350. [Google Scholar] [CrossRef]
- Celino-Brady, F.T.; Petro-Sakuma, C.K.; Breves, J.P.; Lerner, D.T.; Seale, A.P. Early-life exposure to 17β-estradiol and 4-nonylphenol impacts the growth hormone/insulin-like growth-factor system and estrogen receptors in Mozambique tilapia, Oreochromis mossambicus. Aquat. Toxicol. 2019, 217, 105336. [Google Scholar] [CrossRef]
- Araújo, F.G.; Morado, C.N.; Parente, T.T.E.; Paumgartten, F.J.R.; Gomes, I.D. Biomarkers and bioindicators of the environmental condition using a fish species (Pimelodus maculatus Lacepède, 1803) in a tropical reservoir in Southeastern Brazil. Braz. J. Biol. 2017, 78, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Pavione, P.D.M.; Da Costa, K.G.; Perônico, C.; McMaster, M.E.; Parrott, J.L.; Hewitt, L.M.; Munkittrick, K.R.; Barreto, F.C.C.; Basilo, T.H.; Gomes, M.P.; et al. Development of environmental effects monitoring protocol in Brazil: A fish guide study of three river estuaries. Environ. Monit. Assess. 2019, 191, 658. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, E.C.; Auache-Filho, A.A.; Damasio, D.; Ghisi, N.D.C.; Michels-Souza, M.A. Reproductive indicators of the endemic species Astyanax bifasciatus (Teleostei: Characidae) in a tributary of the Lower Iguaçu River Basin, Brazil. Acta Sci. Biol. Sci. 2019, 41, e47720. [Google Scholar] [CrossRef] [Green Version]
- Burkina, V.; Zamaratskaia, G.; Sakalli, S.; Giang, P.T.; Kodes, V.; Grabic, R.; Velisek, J.; Turek, J.; Kolarova, J.; Zlabek, V.; et al. Complex effects of pollution on fish in major rivers in the Czech Republic. Ecotoxicol. Environ. Saf. 2018, 164, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Mabansag, C.J.A.; Paraso, M.G.V.; Marcelino, R.T.; Clavecillas, A.A.; Lola, M.S.E.G. A Preliminary Survey of Estrogenic Effects in Cultured Adult Male Nile Tilapia (Oreochromis niloticus) in the Seven Lakes of San Pablo City, Philippines. Bull. Environ. Contam. Toxicol. 2019, 103, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Rodgers-Gray, T.P.; Jobling, S.; Kelly, C.; Morris, S.; Brighty, G.; Waldock, M.J.; Sumpter, J.P.; Tyler, C.R. Exposure of Juvenile Roach (Rutilus rutilus) to Treated Sewage Effluent Induces Dose-Dependent and Persistent Disruption in Gonadal Duct Development. Environ. Sci. Technol. 2000, 35, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Amiard-Triquet, C.; Amiard, J.C.; Rainbow, P.S. (Eds.) Ecological Biomarkers Indicators of Ecotoxicological Effects; CRS Press: Boca Raton, FL, USA, 2013; Volume 53, ISBN 9788578110796. Available online: https://www.routledge.com/ (accessed on 22 March 2013).
- Chen, T.-H.; Chou, S.-M.; Tang, C.-H.; Chen, C.-Y.; Meng, P.-J.; Ko, F.-C.; Cheng, J.-O. Endocrine disrupting effects of domestic wastewater on reproduction, sexual behavior, and gene expression in the brackish medaka Oryzias melastigma. Chemosphere 2016, 150, 566–575. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (µg/L) | Total Length (mm) | Body Weight (mg) | Condition Factor (K) |
|---|---|---|---|
| Control | 31± 3.7 | 213.6 ± 11.9 | 0.72 |
| Solvent | 28± 0.4 | 214.6 ± 18.8 | 1.01 |
| 10 | 28± 1.2 | 209.8 ± 29.9 | 0.94 |
| 25 | 27 ± 0.6 | 209.1 ± 11.2 | 1.04 |
| 50 | 25 ± 0.3 | 233.2 ± 9.6 | 1.49 |
| 125 | 27 ± 0.3 | 210.3 ± 3.5 | 1.07 |
| 250 | 29 ± 1.5 | 260.3 ± 33.3 | 1.07 |
| Concentration (µg/L) | HSI (%) | GSI (%) | ||
|---|---|---|---|---|
| Male | Female | Male | Female | |
| Control | 3.65 ± 0.9 | 4.14 ± 0.8 | 2.44 ± 0.1 | 4.19 ± 0.2 |
| Solvent | 3.89 ± 1.3 | 6.34 ± 0.4 | 3.55 ± 0.8 | 4.92 ± 0.4 |
| 10 | 4.12 ± 0.8 | 4.15 ± 0.8 | 2.78 ± 0.5 | 3.64± 0.5 |
| 25 | 4.39 ± 1.3 | 6.03 ± 0.4 | 3.94 ± 0.3 | 3.79 ± 0.6 |
| 50 | 4.62 ± 1.4 | 5.11 ± 0.4 | 2.87 ± 0.2 | 4.36 ± 0.4 |
| 125 | 3.87 ± 1.6 | 5.16 ± 0.7 | 2.12 ± 0.6 | 2.84 ± 0.4 |
| 250 | 4.55 ± 0.3 | 5.90 ± 0.5 | 1.99 ± 0.4 | 2.47 ± 0.3 * |
| Treatment (µg/L) | Staging (Median) | Lesions (%) | |||
|---|---|---|---|---|---|
| 1 Male | 2 Female | Atretic OOCYTE | Oocyte Retraction | Karyoptic Clumping | |
| Control | 4 | 4 | 0.0 | 0.0 | 0.0 |
| Solvent | 4 | 4 | 0.0 | 0.0 | 0.0 |
| 10 | 4 | 4 | 0.0 | 0.0 | 1.8 |
| 25 | 4 | 3 | 2.1 | 5.2 | 7.3 |
| 50 | 3 | 4 | 4.9 | 1.5 | 3.0 |
| 125 | 4 | 4 | 1.3 | 0.0 | 5.7 |
| 250 | 3 | 2 * | 11.2 * | 5.2 | 4.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, M.A.; Zulkifli, S.Z.; Azmai, M.N.A.; Mohamat-Yusuff, F.; Ismail, A. Reproductive Toxicity of 3,4-dichloroaniline (3,4-DCA) on Javanese Medaka (Oryziasjavanicus, Bleeker 1854). Animals 2021, 11, 798. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11030798
Ibrahim MA, Zulkifli SZ, Azmai MNA, Mohamat-Yusuff F, Ismail A. Reproductive Toxicity of 3,4-dichloroaniline (3,4-DCA) on Javanese Medaka (Oryziasjavanicus, Bleeker 1854). Animals. 2021; 11(3):798. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11030798
Chicago/Turabian StyleIbrahim, Musa Adamu, Syaizwan Zahmir Zulkifli, Mohammad Noor Amal Azmai, Ferdaus Mohamat-Yusuff, and Ahmad Ismail. 2021. "Reproductive Toxicity of 3,4-dichloroaniline (3,4-DCA) on Javanese Medaka (Oryziasjavanicus, Bleeker 1854)" Animals 11, no. 3: 798. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11030798