
Muscle Transcriptome Analysis Reveals Molecular Pathways Related to Oxidative Phosphorylation, Antioxidant Defense, Fatness and Growth in Mangalitsa and Moravka Pigs

 , , , ,
, , , ,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Sampling
2.3. Phenotypic Analysis
2.4. Transcriptome Analysis
2.4.1. RNA Extraction
2.4.2. RNA Library Construction and Sequencing
2.4.3. Mapping, Assembly and Differential Expression Analysis
2.4.4. Reverse Transcription Quantitative PCR (RT-qPCR)
2.4.5. Statistical Analysis of qPCR Data
2.4.6. Functional Analysis
3. Results and Discussion
3.1. Validation by RT-qPCR
3.2. Breed and Diet Effects on Studied Phenotype
3.3. Breed Effect on Transcriptome
3.4. Functional Analysis of Breed Effects
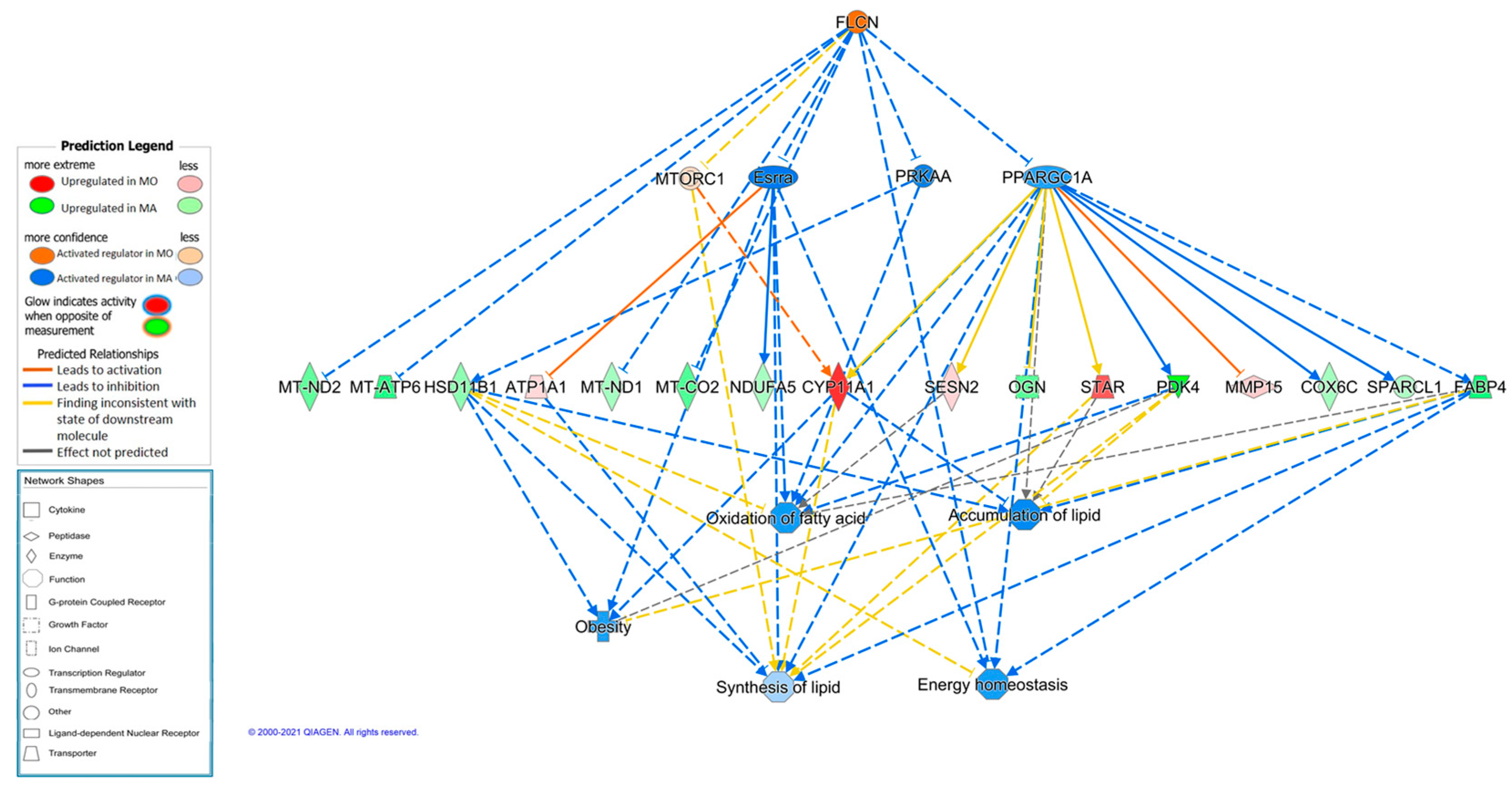
3.5. Prediction of Regulators for Breed Effects
3.6. Tannin Effect on Transcriptome
3.7. Functional Analysis of Tannin Effects
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Radović, Č.; Savić, R.; Petrović, M.; Gogić, M.; Lukić, M.; Radojković, D.; Batorek-Lukač, N. Mangalitsa (Swallow-Belly Mangalitsa) Pig, European Local Pig Breeds—Diversity and Performance: A Study of Project TREASURE; Marjeta, C.-P., Rosa, M., Nieto, L., Eds.; IntechOpen: London, UK, 2019; Available online: https://www.intechopen.com/books/european-local-pig-breeds-diversity-and-performance-a-study-of-project-treasure/mangalitsa-swallow-belly-mangalitsa-pig (accessed on 6 February 2019). [CrossRef] [Green Version]
- Savić, R.; Radović, Č.; Petrović, M.; Gogić, M.; Radojković, D.; Batorek-Lukač, N. Moravka Pig, European Local Pig Breeds—Diversity and Performance: A Study of Project TREASURE; Marjeta, C.-P., Rosa, M., Nieto, L., Eds.; IntechOpen: London, UK. Available online: https://www.intechopen.com/books/european-local-pig-breeds-diversity-and-performance-a-study-of-project-treasure/moravka-pig (accessed on 6 February 2019). [CrossRef] [Green Version]
- Egerszegi, I.; Rátky, J.; Solti, L.; Brüssow, K. Mangalica—An indigenous swine breed from Hungary (Review). Arch. Tierz 2003, 46, 245–256. [Google Scholar] [CrossRef]
- Petrović, M.; Radović, Č.; Parunović, N.; Mijatović, M.; Radojković, D.; Aleksić, S.; Stanišić, N.; Popovac, M. Quality traits of carcass sides and meat of moravka and mangalitsa pig breeds. Biotechnol. Anim. Hunsbandry 2010, 26, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, M.; Bozzi, R.; García, F.; Núñez, Y.; Geraci, C.; Crovetti, A.; García-Casco, J.; Alves, E.; Škrlep, M.; Charneca, R.; et al. Diversity across major and candidate genes in European local pig breeds. PLoS ONE 2018, 13, e0207475. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, M.; Bozzi, R.; García-Casco, J.; Núñez, Y.; Ribani, A.; Franci, O.; García, F.; Škrlep, M.; Schiavo, G.; Bovo, S.; et al. Genomic diversity, linkage disequilibrium and selection signatures in European local pig breeds assessed with a high density SNP chip. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Mueller-Harvey, I. Unravelling the conundrum of tannins in animal nutrition and health. J. Sci. Food Agric. 2006, 86, 2010–2037. [Google Scholar] [CrossRef]
- Frankič, T.; Salobir, J. In vivo oxidative potential of Sweet chestnut (Castanea sativa Mill.) wood extract in young growing pigs to n-3 PUFA-induced oxidative stress. J. Sci. Food Agric. 2011, 91, 1432–1439. [Google Scholar] [CrossRef] [PubMed]
- Voljč, M.; Levart, A.; Žgur, S.; Salobir, J. The effect of α-tocopherol, sweet chestnut wood extract and their combination on oxidative stress in vivo and the oxidative stability of meat in broilers. Br. Poult. Sci. 2013, 54, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Salobir, J.; Kostanjevec, B.; Štruklec, M.; Salobir, K. Tannins reduce protein but not phosphorus utilization of feed with added phytase in pigs. J. Anim. Feed Sci. 2005, 14, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Čandek-Potokar, M.; Škrlep, M.; Batorek Lukac, N.; Zamaratskaia, G.; Prevolnik Povše, M.; Velikonja Bolta, S.; Kubale, V.; Bee, G. Hydrolysable tannin fed to entire male pigs affects intestinal production, tissue deposition and hepatic clearance of skatole. Vet. J. 2015, 204, 162–167. [Google Scholar] [CrossRef]
- Bilić-Šobot, D.; Zamaratskaia, G.; Rasmussen, M.K.; Čandek-Potokar, M.; Škrlep, M.; Povše, M.P.; Škorjanc, D. Chestnut wood extract in boar diet reduces intestinal skatole production, a boar taint compound. Agron. Sustain. Dev. Springer Verl. Edp Sci. Inra 2016, 36, 62. [Google Scholar] [CrossRef] [Green Version]
- Bee, G.; Silacci, P.; Ampuero-Kragten, S.; Čandek-Potokar, M.; Wealleans, A.L.; Litten-Brown, J.; Salminen, J.P.; Mueller-Harvey, I. Hydrolysable tannin-based diet rich in gallotannins has a minimal impact on pig performance but significantly reduces salivary and bulbourethral gland size. Animal 2017, 11, 1617–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tretola, M.; Maghin, F.; Silacci, P.; Ampuero, S.; Bee, G. Effect of Supplementing Hydrolysable Tannins to a Grower–Finisher Diet containing Divergent PUFA Levels on Growth Performance, Boar Taint Levels in Back Fat and Intestinal Microbiota of Entire Males. Animals 2019, 9, 1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezar, V.; Salobir, J.; Levart, A.; Tomažin, U.; Škrlep, M.; Lukač, N.B.; Čandek-Potokar, M. Supplementing entire male pig diet with hydrolysable tannins: Effect on carcass traits, meat quality and oxidative stability. Meat Sci. 2017, 133, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Benítez, R.; Núñez, Y.; Óvilo, C. Nutrigenomics in Farm Animals. J. Investig. Genom. 2017, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Óvilo, C.; Benítez, R.; Fernández, A.; Núñez, Y.; Ayuso, M.; Fernández, A.I.; Rodríguez, C.; Isabel, B.; Rey, A.I.; López-Bote, C.; et al. Longissimus dorsi transcriptome analysis of purebred and crossbred Iberian pigs differing in muscle characteristics. BMC Genom. 2014, 15, 413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damon, M.; Wyszynska-Koko, J.; Vincent, A.; Herault, F.; Lebret, B. Comparison of muscle transcriptome between pigs with divergent meat quality phenotypes identifies genes related to muscle metabolism and structure. PLoS ONE 2012, 7, e33763. [Google Scholar] [CrossRef] [Green Version]
- Ayuso, M.; Fernández, A.; Núñez, Y.; Benítez, R.; Isabel, B.; Barragán, C.; Fernández, A.I.; Rey, A.I.; Medrano, J.F.; Cánovas, A.; et al. Comparative Analysis of Muscle Transcriptome between Pig Genotypes Identifies Genes and Regulatory Mechanisms Associated to Growth, Fatness and Metabolism. PLoS ONE 2015, 10, e0145162. [Google Scholar] [CrossRef] [Green Version]
- García-Contreras, C.; Madsen, O.; Groenen, M.A.M.; López-García, A.; Vázquez-Gómez, M.; Astiz, S.; Núñez, Y.; Benítez, R.; Fernández, A.; Isabel, B.; et al. Impact of genotype, body weight and sex on the prenatal muscle transcriptome of Iberian pigs. PLoS ONE 2020, 15, e0227861. [Google Scholar] [CrossRef] [Green Version]
- Spiric, A.; Trbović, D.; Vranić, D.; Djinović, J.; Petronijević, R.; Matekalo-Sverak, V. Statistical evaluation of fatty acid profile and cholesterol content in fish (common carp) lipids obtained by different sample preparation procedures. Anal. Chim. Acta 2010, 672, 66–71. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 0034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, C.L.; Ledet-Jensen, J.; Orntoft, T. Normalization of real-time quantitative RT-PCR data: A model based variance estimation approach to identify genes suited for normalization—applied to bladder- and colon-cancer data-sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Steibel, J.P.; Poletto, R.; Coussens, P.M.; Rosa, G.J.M. A powerful and flexible linear mixed model framework for the analysis of relative quantification RT-PCR data. Genomics 2009, 94, 146–152. [Google Scholar] [CrossRef] [Green Version]
- Miron, M.; Woody, O.Z.; Marcil, A.; Murie, C.; Sladek, R.; Nadon, R. A methodology for global validation of microarray experiments. BMC Bioinform. 2006, 7, 333. [Google Scholar] [CrossRef] [Green Version]
- Radović, Č.; Petrović, M.; Savić, R.; Gogić, M.; Lukić, M.; Stanišić, N.; Čandek-Potokar, M. Growth potential of Serbian local pig breeds mangalitsa and moravka. Agric. Conspec. Sci. 2017, 82, 217–220. [Google Scholar]
- Petrović, M.; Mijatovic, M.; Radović, C.; Radojkovic, D.; Josipovic, S. Genetic resources in pig breeding: Carcass quality traits of breeds Moravka and Mangalitsa. Biotechnol. Anim. Husb. 2007, 23, 421–428. [Google Scholar] [CrossRef]
- Radović, Č.; Petrović, M.; Parunovic, N.; Radojkovic, D.; Savić, R.; Stanišic, N.; Gogić, M. Carcass and pork quality traits of indigenous pure breeds (Mangalitsa, Moravka) and their crossbreads. Indian J. Anim. Res. 2017, 51, 371–376. [Google Scholar] [CrossRef]
- Petrović, M.; Wähner, M.; Radović, Č.; Radojković, D.; Parunović, N.; Savić, R.; Brkić, N. Fatty acid profile of m. longissimus dorsi of Mangalitsa and Moravka pig breeds. Arch. Tierz. 2014, 57, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Prevolnik, M.; Škrlep, M.; Brus, M.; Pugliese, C.; Čandek-Potokar, M.; Škorjanc, D. Supplementing pig diet with 0.2% sweet chestnut (Castanea sativa mill.) wood extract had no effect on growth, carcass or meat quality. Acta Argic. Slov. 2012, 3, 83–88. [Google Scholar]
- Lee, S.H.; Shinde, P.L.; Choi, J.Y.; Kwon, I.K.; Lee, J.K.; Pak, S.I.; Cho, W.T.; Chae, B.J. Effects of tannic acid supplementation on growth performance, blood hematology, iron status and faecal microflora in weanling pigs. Livest. Sci. 2010, 131, 281–286. [Google Scholar] [CrossRef]
- Biagi, G.; Cipollini, I.; Paulicks, B.R.; Roth, F.X. Effect of tannins on growth performance and intestinal ecosystem in weaned piglets. Arch. Anim. Nutr. 2010, 64, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Cadoudal, T.; Distel, E.; Durant, S.; Fouque, F.; Blouin, J.M.; Collinet, M.; Bortoli, S.; Forest, C.; Benelli, C. Pyruvate dehydrogenase kinase 4: Regulation by thiazolidinediones and implication in glyceroneogenesis in adipose tissue. Diabetes 2008, 57, 2272–2279. [Google Scholar] [CrossRef] [Green Version]
- Lan, J.; Lei, M.G.; Zhang, Y.; Wang, J.; Feng, X.; Xu, D.; Gui, J.; Xiong, Y. Characterization of the porcine differentially expressed PDK4 gene and association with meat quality. Mol. Biol. Rep. 2009, 36, 2003–2010. [Google Scholar] [CrossRef] [Green Version]
- Shi-Zheng, G.; Su-Mei, Z. Physiology, Affecting Factors and Strategies for Control of Pig Meat Intramuscular Fat. Recent Pat. Food Nutr. Agric. 2009, 1, 59–74. [Google Scholar] [CrossRef] [Green Version]
- Qiu, X.B.; Shao, Y.M.; Miao, S.; Wang, L. The diversity of the DnaJ/Hsp40 family, the crucial partners for Hsp70 chaperones. Cell Mol. Life Sci. 2006, 63, 2560–2570. [Google Scholar] [CrossRef]
- Ishibashi, J.; Perry, R.L.; Asakura, A.; Rudnicki, M.A. MyoD induces myogenic differentiation through cooperation of its NH2- and COOH-terminal regions. J. Cell Biol. 2005, 171, 471–482. [Google Scholar] [CrossRef] [Green Version]
- Shelton, M.; Ritso, M.; Liu, J.; O’Neil, D.; Kocharyan, A.; Rudnicki, M.A.; Stanford, W.L.; Skerjanc, I.S.; Blais, A. Gene expression profiling of skeletal myogenesis in human embryonic stem cells reveals a potential cascade of transcription factors regulating stages of myogenesis, including quiescent/activated satellite cell-like gene expression. PLoS ONE 2019, 14, e0222946. [Google Scholar] [CrossRef]
- Frank, D.A. STAT3 as a central mediator of neoplastic cellular transformation. Cancer Lett. 2007, 251, 199–210. [Google Scholar] [CrossRef]
- Listrat, A.; Lebret, B.; Louveau, I.; Astruc, T.; Bonnet, M.; Lefaucheur, L.; Picard, B.; Bugeon, J. How Muscle Structure and Composition Influence Meat and Flesh Quality. Sci. World J. 2016, 2016, 3182746. [Google Scholar] [CrossRef] [Green Version]
- Serra, X.; Gil, F.; Pérez-Enciso, M.; Oliver, M.A.; Vázquez, J.M.; Gispert, M.; Díaz, I.; Moreno, F.; Latorre, R.; Noguera, J.L. A comparison of carcass, meat quality and histochemical characteristics of Iberian (Guadyerbas line) and Landrace pigs. Livest. Prod. Sci. 1998, 56, 215–223. [Google Scholar] [CrossRef]
- Murgia, M.; Nagaraj, N.; Deshmukh, A.S.; Zeiler, M.; Cancellara, P.; Moretti, I.; Reggiani, C.; Schiaffino, S.; Mann, M. Single muscle fiber proteomics reveals unexpected mitochondrial specialization. EMBO Rep. 2015, 16, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Stamler, J.S.; Meissner, G. Physiology of nitric oxide in skeletal muscle. Physiol Rev. 2001, 81, 209–237. [Google Scholar] [CrossRef] [PubMed]
- Puig-Oliveras, A.; Ballester, M.; Corominas, J.; Revilla, M.; Estellé, J.; Fernández, A.I.; Ramayo-Caldas, Y.; Folch, J.M. A Co-Association Network Analysis of the Genetic Determination of Pig Conformation, Growth and Fatness. PLoS ONE 2014, 9, e114862. [Google Scholar] [CrossRef] [Green Version]
- Vomund, S.; Schäfer, A.; Parnham, M.J.; Brüne, B.; von Knethen, A. Nrf2, the Master Regulator of Anti-Oxidative Responses. Int. J. Mol. Sci. 2017, 18, 2772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillet, L.C.; Scharer, O.D. Molecular mechanisms of mammalian global genome nucleotide excision repair. Chem. Rev. 2006, 106, 253–276. [Google Scholar] [CrossRef]
- Marteijn, J.A.; Lans, H.; Vermeulen, W.; Hoeijmakers, J.H. Understanding nucleotide excision repair and its roles in cancer and ageing. Nat. Rev. Mol. Cell Biol. 2014, 15, 465–481. [Google Scholar] [CrossRef]
- Melis, J.P.; van Steeg, H.; Luijten, M. Oxidative DNA damage and nucleotide excision repair. Antioxid Redox Signal. 2013, 18, 2409–2419. [Google Scholar] [CrossRef] [Green Version]
- Pakos-Zebrucka, K.; Koryga, I.; Mnich, K.; Ljujic, M.; Samali, A.; Gorman, A.M. The integrated stress response. EMBO Rep. 2016, 17, 1374–1395. [Google Scholar] [CrossRef] [Green Version]
- Martínez, J.A. Mitochondrial oxidative stress and inflammation: An slalom to obesity and insulin resistance. J. Physiol. Biochem. 2006, 62, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Bote, C.J. Sustained utilization of the Iberian pig breed. Meat Sci 1998, 49, S17–S27. [Google Scholar] [CrossRef]
- Torres-Rovira, L.; Astiz, S.; Caro, A.; Lopez-Bote, C.; Ovilo, C.; Pallares, P.; Perez-Solana, M.L.; Sanchez-Sanchez, R.; Gonzalez-Bulnes, A. Diet-Induced Swine Model with Obesity/Leptin Resistance for the Study of Metabolic Syndrome and Type 2 Diabetes. Sci. World J. 2012, 2012, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Nieto, R.; García-Casco, J.; Lara, L.; Palma-Granados, P.; Izquierdo, M.; Hernandez, F.; Dieguez, E.; Duarte, J.L.; Batorek-Lukač, N. Ibérico (Iberian) Pig, European Local Pig Breeds—Diversity and Performance: A Study of Project TREASURE; Marjeta, C.-P., Rosa, M., Ni, L., Eds.; IntechOpen: London, UK, 2019; Available online: https://www.intechopen.com/books/european-local-pig-breeds-diversity-and-performance-a-study-of-project-treasure/ib-rico-iberian-pig (accessed on 13 March 2021). [CrossRef] [Green Version]
- Berdeaux, R.; Stewart, R. cAMP signaling in skeletal muscle adaptation: Hypertrophy, metabolism, and regeneration. Am J Physiol. Endocrinol. Metab. 2012, 303, E1–E17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watt, K.I.; Goodman, C.A.; Hornberger, T.A.; Gregorevic, P. The Hippo Signaling Pathway in the Regulation of Skeletal Muscle Mass and Function. Exerc. Sport Sci. Rev. 2018, 46, 92–96. [Google Scholar] [CrossRef]
- Shao, D.; Liu, Y.; Liu, X.; Zhu, L.; Cui, Y.; Cui, A.; Qiao, A.; Kong, X.; Liu, Y.; Chen, Q.; et al. PGC-1β-Regulated mitochondrial biogenesis and function in myotubes is mediated by NRF-1 and ERRα. Mitochondrion 2010, 10, 516–527. [Google Scholar] [CrossRef]
- Cho, Y.; Hazen, B.C.; Russell, A.P.; Kralli, A. Peroxisome proliferator-activated receptor γ coactivator 1 (PGC-1)- and estrogen-related receptor (ERR)-induced regulator in muscle 1 (Perm1) is a tissue-specific regulator of oxidative capacity in skeletal muscle cells. J. Biol Chem. 2013, 288, 25207–25218. [Google Scholar] [CrossRef] [Green Version]
- Lu, K.; Policar, T.; Song, X.; Rahimnejad, S. Molecular Characterization of PGC-1β (PPAR Gamma Coactivator 1β) and its Roles in Mitochondrial Biogenesis in Blunt Snout Bream (Megalobrama amblycephala). Int. J. Mol. Sci. 2020, 21, 1935. [Google Scholar] [CrossRef] [Green Version]
- Hasumi, H.; Baba, M.; Hasumi, Y.; Huang, Y.; Oh, H.; Hughes, R.M.; Klein, M.E.; Takikita, S.; Nagashima, K.; Schmidt, L.S.; et al. Regulation of Mitochondrial Oxidative Metabolism by Tumor Suppressor FLCN. J. Natl. Cancer Inst. 2012, 104, 1750–1764. [Google Scholar] [CrossRef] [Green Version]
- Beharry, A.W.; Sandesara, P.B.; Roberts, B.M.; Ferreira, L.F.; Senf, S.M.; Judge, A.R. HDAC1 activates FoxO and is both sufficient and required for skeletal muscle atrophy. J. Cell Sci. 2014, 127, 1441–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayuso, M.; Fernández, A.; Núñez, Y.; Benítez, R.; Isabel, B.; Fernández, A.I.; Rey, A.I.; González-Bulnes, A.; Medrano, J.F.; Cánovas, Á.; et al. Developmental Stage, Muscle and Genetic Type Modify Muscle Transcriptome in Pigs: Effects on Gene Expression and Regulatory Factors Involved in Growth and Metabolism. PLoS ONE 2016, 11, e0167858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, J.; Ghanem, L.; Bar-Joseph, Z.; McNamara, M.; Brown, J.; Steinman, R.A. IL-3 and Oncogenic Abl Regulate the Myeloblast Transcriptome by Altering mRNA Stability. PLoS ONE 2009, 4, e7469. [Google Scholar] [CrossRef]
- MacLachlan, T.K.; Giordano, A. TRAF2 expression in differentiated muscle. J. Cell Biochem. 1998, 71, 461–466. [Google Scholar] [CrossRef]
- Ouchi, N.; Shibata, R.; Walsh, K. AMP-Activated Protein Kinase Signaling Stimulates VEGF Expression and Angiogenesis in Skeletal Muscle. Am. Heart Assoc. 2005, 96, 838–846. [Google Scholar] [CrossRef] [Green Version]
- Kahn, B.B.; Alquier, T.; Carling, D.; Hardie, D.G. AMP-activated protein kinase: Ancient energy gauge provides clues to modern understanding of metabolism. Cell Metab. 2005, 1, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Hardie, D.G. AMPK: A key regulator of energy balance in the single cell and the whole organism. Int. J. Obes. 2008, 32, S7–S12. [Google Scholar] [CrossRef] [Green Version]
- Óvilo, C.; Benítez, R.; Fernández, A.; Isabel, B.; Núñez, Y.; Fernández, A.I.; Rodríguez, C.; Daza, A.; Silió, L.; López-Bote, C. Dietary energy source largely affects tissue fatty acid composition but has minor influence on gene transcription in Iberian pigs. J. Anim. Sci. 2014, 92, 939–954. [Google Scholar] [CrossRef]
- Benítez, R.; Trakooljul, N.; Núñez, Y.; Isabel, B.; Murani, E.; De Mercado, E.; Gómez-Izquierdo, E.; García-Casco, J.; López-Bote, C.; Wimmers, K.; et al. Breed, Diet, and Interaction Effects on Adipose Tissue Transcriptome in Iberian and Duroc Pigs Fed Different Energy Sources. Genes 2019, 10, 589. [Google Scholar] [CrossRef] [Green Version]
- Mennen, L.I.; Walker, R.; Bennetau-Pelissero, C.; Scalbert, A. Risks and safety of polyphenol consumption. Am. J. Clin. Nutr. 2005, 81, 326S–329S. [Google Scholar] [CrossRef] [Green Version]
- Barrett, A.H.; Farhadi, N.F.; Smith, T.J. Slowing starch digestion and inhibiting digestive enzyme activity using plant flavanols/tannins—A review of efficacy and mechanisms. LWT 2018, 87, 394–399. [Google Scholar] [CrossRef]
- Lizardo, R.; Peiniau, J.; Aumaitre, A. Effect of sorghum on performance, digestibility of dietary components and activities of pancreatic and intestinal enzymes in the weaned piglet. Anim. Feed Sci. Technol. 1995, 56, 67–82. [Google Scholar] [CrossRef]
- Smulikowska, S.; Pastuszewska, B.; Switch, E.; Ochtabinska, A.; Mieczkowska, A.; Nguyen, V.C.; Buraczewska, L. Tannin content affects negatively nutritive value of pea for monogastrics. J. Anim. Feed Sci. 2001, 10, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Jansman, A.J.M.; Verstegen, M.W.A.; Huisman, J.; van den Berg, J.W.O. Effects of hulls of faba beans (Vicia faba L.) with a low or high content of condensed tannins on the apparent ileal and fecal digestibility of nutrients and the excretion of endogenous protein in ileal digesta and feces of pigs. J. Anim. Sci. 1995, 73, 118–127. [Google Scholar] [CrossRef]
- Rodríguez-Estévez, V.; Sánchez-Rodríguez, M.; Garcia, A.; Gómez-Castro, G. Average daily weight gain of Iberian fattening pigs when grazing natural resources. Livest. Sci. 2011, 137, 292–295. [Google Scholar] [CrossRef]
- Mole, S.; Butler, L.G.; Iason, G. Defense against dietary tannin in herbivores: A survey for proline rich salivary proteins in mammals. Biochem. Syst. Ecol. 1990, 18, 287–293. [Google Scholar] [CrossRef]
- Brus, M.; Keller, M.; Lévy, F. Temporal features of adult neurogenesis: Differences and similarities across mammalian species. Front. Neurosci. 2013, 7, 135. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Choi, I.H.; Kim, D.H.; Amanullah, S.M.; Kim, S.C. Nutritional characterization of tannin rich chestnut (Castanea) and its meal for pig. J. Appl. Anim. Res. 2016, 44, 258–262. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Zhou, Y.; Jiang, S.; Huang, F.; Peng, J.; Jiang, S. Transcriptional response of porcine skeletal muscle to feeding a linseed-enriched diet to growing pigs. J. Anim. Sci. Biotechnol. 2016, 7, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Tena, S.; Reyes-Zurita, F.J.; Díaz-Moralli, S.; Vinardell, M.P.; Reed, M.; García-García, F.; Dopazo, J.; Lupiáñez, J.A.; Günther, U.; Cascante, M. Maslinic Acid-Enriched Diet Decreases Intestinal Tumorigenesis in ApcMin/+ Mice through Transcriptomic and Metabolomic Reprogramming. PLoS ONE 2013, 8, e59392. [Google Scholar] [CrossRef] [Green Version]
- Krasteva, S.; Heiss, E.; Krenn, L. Optimization and application of a fluorimetric assay for the identification of histone deacetylase inhibitors from plant origin. Pharm. Biol. 2011, 49, 658–668. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Xiao, H.; Zhu, Y.; Liu, S.; Yuan, Z.; Wu, J.; Wen, L. Tannic Acid Induces the Mitochondrial Pathway of Apoptosis and S Phase Arrest in Porcine Intestinal IPEC-J2 Cells. Toxins 2019, 11, 397. [Google Scholar] [CrossRef] [Green Version]
- Swan, L.E.; Wichmann, C.; Prange, U.; Schmid, A.; Schmidt, M.; Schwarz, T.; Ponimaskin, E.; Madeo, F.; Vorbrüggen, G.; Sigrist, S.J. A glutamate receptor-interacting protein homolog organizes muscle guidance in Drosophila. Genes Dev. 2004, 18, 223–237. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.B.; Park, H.B.; Yoo, C.K.; Kim, H.S.; Cho, I.C.; Lim, H.T. Association of a missense mutation in the positional candidate gene glutamate receptor-interacting protein 1 with backfat thickness traits in pigs. Asian-Australas J. Anim. Sci 2017, 30, 1081–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, S.; Ito, Y.; Ueno-Kudoh, H.; Shimizu, H.; Uchibe, K.; Albini, S.; Mitsuoka, K.; Miyaki, S.; Kiso, M.; Nagai, A.; et al. A systems approach reveals that the myogenesis genome network is regulated by the transcriptional repressor RP58. Dev. Cell. 2009, 17, 836–848. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g/kg) | Mixture I (25–60 kg) | Mixture II (60–120 kg) |
|---|---|---|
| Corn (dry) | 632.7 | 691.5 |
| Wheat flour | 150.0 | 150.0 |
| Soybean meal | 139.0 | 90.0 |
| Sunflower meal | 50.0 | 40.0 |
| Calcium carbonate | 10.0 | 9.0 |
| Dicalcium | 9.0 | 10.0 |
| Salt | 4.3 | 4.5 |
| Premix | 5.0 | 5.0 |
| Energy (MJ/kg) | 13.6 | 13.5 |
| Protein (g/kg) | 147 | 130 |
| Lysine (g/kg) | 6.6 | 5.5 |
| Gene | MA vs. MO | |||||
|---|---|---|---|---|---|---|
| RNA-Seq | qPCR | Correlation | ||||
| log2 FC | q | log2 FC | p | r | p Value | |
| ND2 | −1.406 | 0.019 | −1.260 | 0.0088 | 0.744 | 0.013 |
| ATP6 | −1.661 | 0.001 | −0.980 | 0.0301 | 0.626 | 0.053 |
| COX2 | −1.497 | 0.003 | −1.407 | 0.0001 | 0.724 | 0.018 |
| PDK4 | −2.869 | 0.101 | −3.563 | 0.0157 | 0.999 | 2.4 × 10−12 |
| JAZF1 | −1.185 | 0.065 | −0.711 | 0.0246 | 0.651 | 0.041 |
| NOS1 | 1.333 | 0.010 | 0.896 | 0.0091 | 0.824 | 0.003 |
| MYOD1 | 1.390 | 0.074 | 0.571 | 0.2939 | 0.974 | 1.9 × 10−6 |
| STAT3 | 0.642 | 0.070 | 0.432 | 0.1351 | 0.670 | 0.076 |
| FOS | 2.333 | 0.261 | 1.102 | 0.4670 | 0.877 | 0.0008 |
| MA vs. MAT | ||||||
| ARID5A | −1.488 | 0.087 | −1.108 | 0.0028 | 0.871 | 0.002 |
| DAPK3 | −1.392 | 0.000 | −1.447 | 0.0001 | 0.894 | 0.001 |
| TNFRSF12A | −1.563 | 0.000 | −1.099 | 0.0510 | 0.888 | 0.001 |
| RUNX1 | −1.117 | 0.560 | −0.919 | 0.1590 | 0.936 | 0.075 |
| PPARGC1B | 0.948 | 0.007 | 0.503 | 0.1660 | 0.620 | 0.0002 |
| ACLY | 1.266 | 0.581 | 1.088 | 0.1886 | 0.992 | 1.5 × 10−7 |
| Trait | Least Squares Means | MA vs. MO | MA vs. MAT | ||||
|---|---|---|---|---|---|---|---|
| MA n = 12 | MO n = 10 | MAT n = 12 | SEM | p | SEM | p | |
| Slaughter Age, days | 363.2 | 357.3 | 361.9 | 22.80 | 0.555 | 19.63 | 0.878 |
| Average Daily Gain, g | 318.9 | 366.6 | 306.4 | 13.40 | 0.002 | 10.22 | 0.397 |
| Slaughter Weight, kg | 115.5 | 130.2 | 111.1 | 5.70 | 0.018 | 3.98 | 0.441 |
| Carcass Weight, kg | 89.91 | 103.04 | 85.40 | 4.78 | 0.013 | 3.43 | 0.361 |
| Longissimus dorsi | |||||||
| Loin Thickness, mm | 57.5 | 63.0 | 54.1 | 3.0 | 0.084 | 1.75 | 0.182 |
| Intramuscular fat percentage, % | 8.55 | 8.15 | 11.66 | 1.56 | 0.801 | 0.99 | 0.037 |
| C14:0, % | 2.20 | 2.18 | 2.25 | 0.07 | 0.787 | 0.05 | 0.459 |
| C15:0, % | 0.03 | 0.03 | 0.03 | 0.01 | 0.826 | 0.00 | 0.509 |
| C16:0, % | 27.92 | 27.77 | 28.60 | 0.50 | 0.762 | 0.34 | 0.166 |
| C16:1, % | 3.95 | 3.22 | 3.80 | 0.21 | 0.002 | 0.12 | 0.332 |
| C17:0, % | 0.18 | 0.22 | 0.16 | 0.03 | 0.324 | 0.02 | 0.571 |
| C18:0, % | 11.09 | 12.25 | 11.64 | 0.33 | 0.003 | 0.18 | 0.044 |
| C18:1, % | 46.91 | 46.53 | 46.54 | 0.71 | 0.597 | 0.40 | 0.513 |
| C18:2, % | 4.96 | 4.89 | 4.40 | 0.48 | 0.894 | 0.31 | 0.221 |
| C20:0, % | 0.18 | 0.20 | 0.23 | 0.01 | 0.064 | 0.03 | 0.343 |
| C18:3n-3, % | 0.18 | 0.19 | 0.14 | 0.03 | 0.720 | 0.02 | 0.130 |
| C20:1, % | 0.71 | 0.85 | 0.71 | 0.05 | 0.007 | 0.02 | 0.958 |
| C20:2, % | 0.36 | 0.42 | 0.36 | 0.04 | 0.147 | 0.02 | 0.872 |
| C20:3n-6, % | 0.62 | 0.67 | 0.57 | 0.11 | 0.603 | 0.08 | 0.657 |
| C20:3n-3, % | 0.03 | 0.03 | 0.07 | 0.01 | 0.592 | 0.04 | 0.376 |
| C22:1+C20:4, % | 0.12 | 0.11 | 0.08 | 0.02 | 0.422 | 0.01 | 0.016 |
| Ingenuity Canonical Pathways | p Value * | Uregulated in MA ŧ | Upregulated in MO ŧ | DE Genes |
|---|---|---|---|---|
| Oxidative Phosphorylation | 0.17 × 10−7 | 11/109 (10%) | 0/109 (0%) | COX6C, MT-ATP6, MT-CO2, MT-ND1, MT-ND2, MT-ND3, MT-ND4, NDUFA1, NDUFA5, NDUFB3, UQCRB |
| Mitochondrial Dysfunction | 0.14 × 10−5 | 11/171 (6%) | 0/171 (0%) | COX6C, MT-ATP6, MT-CO2, MT-ND1, MT-ND2, MT-ND3, MT-ND4, NDUFA1, NDUFA5, NDUFB3, UQCRB |
| NER (Nucleotide Excision Repair) Pathway | 0.10 × 10−3 | 4/35 (11%) | 0/35 (0%) | CDK7, GTF2H5, POLR2K, XPA |
| Sirtuin Signaling Pathway | 0.14 × 10−3 | 9/291 (3%) | 2/291 (1%) | ARG2, MT-ATP6, MT-ND1, MT-ND2, MT-ND3, MT-ND4, NDUFA1, NDUFA5, NDUFB3, STAT3, XPA |
| Estrogen Receptor Signaling | 0.27 × 10−2 | 5/328 (2%) | 4/328 (1%) | ARG2, CAV1, MED12, MED6, MMP15, MT-ATP6, PAK1, PLCB4, PLCD1 |
| NRF2-mediated Oxidative Stress Response | 0.36 × 10−2 | 2/189 (1%) | 4/189 (2%) | FMO1, FOSL1, HERPUD1, JUNB, PMF1/PMF1-BGLAP, STIP1 |
| Wnt/Ca+ pathway | 0.45 × 10−2 | 2/62 (3%) | 1/62 (2%) | FZD7, PLCB4, PLCD1 |
| Protein Ubiquitination Pathway | 0.64 × 10−2 | 6/273 (2%) | 1/273 (0%) | DNAJC2, HSPA1L, PSMA3, PSMC6, UBE2V2, USP53, USP8 |
| EIF2 Signaling | 0.69 × 10−2 | 5/223 (2%) | 1/223 (0%) | EIF2S2, EIF3J, EIF5B, RPL22L1, RPL36AL, RPS13 |
| Leptin Signaling in Obesity | 0.70 × 10−2 | 2/74 (3%) | 1/74 (1%) | PLCB4, PLCD1, STAT3 |
| Glucocorticoid Receptor Signaling | 0.70 × 10−2 | 5/336 (1%) | 3/336 (1%) | ANXA1, CCL3, CDK7, GTF2H5, HMGB1, HSPA1L, POLR2K, STAT3 |
| cAMP-mediated signaling | 0.76 × 10−2 | 1/228 (0%) | 5/228 (2%) | AKAP1, CNGB1, GRM2, LAMTOR3, PDE4A, STAT3 |
| Adipogenesis pathway | 0.95 × 10−2 | 3/134 (2%) | 1/134 (1%) | CDK7, FABP4, FZD7, GTF2H5 |
| HIPPO signaling | 0.97 × 10−2 | 0/85 (0%) | 3/85 (4%) | PPP1R10, TEAD3, TP53BP2 |
| Upstream Regulator | Predicted Activation | Activation z-Score | p-Value | Target Molecules in Dataset |
|---|---|---|---|---|
| PDGF BB | Moravka | 2.318 | 2.05 × 10−4 | BHLHE40, CCN2, FOSB, JUNB, KLHL21, RYR3, SLC2A3, ZFP36L1 |
| NFkB | Moravka | 2.346 | 6.85 × 10−2 | CCL3, CCN2, DBP, ERAP1, GPR34, HSPA1L, JUNB, MT-CO2, NOS1 |
| SIRT3 | Moravka | 2.635 | 1.31 × 10−6 | CYP11A1, MT-ATP6, MT-CO2, MT-ND1, MT-ND2, MT-ND3, MT-ND4 |
| RICTOR | Moravka | 2.121 | 2.97 × 10−2 | NDUFA1, NDUFA5, NDUFB3, POMP, PSMA3, PSMC6, RPS13, UQCRB |
| IGF1 | Moravka | 2.179 | 1.00 × 10−1 | BHLHE40, CYP11A1, FABP4, MYOD1, SLC25A25, STAR |
| EGFR | Moravka | 2.191 | 9.58 × 10−2 | CAV1, CCN2, HERPUD1, JUNB, MT-CO2, STAT3 |
| FOXO1 | Moravka | 2.200 | 3.20 × 10−1 | CAV1, CCN2, FABP4, FOSB, JUNB, PDK4 |
| MYRF | Moravka | 2.000 | 1.38 × 10−2 | CCN2, JUNB, KLHL21, PDK4 |
| NR4A1 | Moravka | 2.372 | 7.78 × 10−2 | FABP4, MYOD1, NDUFA1, NDUFB3, PDK4, STAR |
| BMP6 | Moravka | 2.000 | 3.80 × 10−4 | ADAMTS1, CCN2, CYP11A1, FOSL1, STAR, TEF |
| FOXO3 | Moravka | 2.200 | 8.70 × 10−2 | CAV1, CCN2, FABP4, FOSB, JUNB |
| ERBB2 | Moravka | 2.000 | 3.66 × 10−1 | BHLHE40, CCN2, CHST10, JUNB, POLR3A, SMC2, SPARCL1, STAT3 |
| ALKBH1 | Mangalitsa | −2.000 | 2.12 × 10−6 | MT-ATP6, MT-CO2, MT-ND2, MT-ND4 |
| CAB39L | Mangalitsa | −2.000 | 1.63 × 10−3 | COX6C, NDUFA1, NDUFB3, UQCRB |
| NSUN3 | Mangalitsa | −2.000 | 2.12 × 10−6 | MT-ATP6, MT-CO2, MT-ND2, MT-ND4 |
| PPARGC1B | Mangalitsa | −2.236 | 3.65 × 10−4 | MT-ATP6, MT-CO2, MT-ND1, MT-ND2, PDK4 |
| LONP1 | Mangalitsa | −2.646 | 7.80 × 10−5 | GARS1, MT-ATP6, MT-CO2, MT-ND1, MT-ND2, MT-ND3, MT-ND4 |
| TAL1 | Mangalitsa | −2.000 | 9.89 × 10−3 | ARPP21, BCOR, CWC27, FRG1, MS4A2, NOS1, SLC22A16, SLC2A3 |
| SIRT1 | Mangalitsa | −2.201 | 2.96 × 10−1 | CAV1, DBP, EIF2S2, FABP4, PDK4, STAT3 |
| DAP3 | Mangalitsa | −2.449 | 8.73 × 10−9 | MT-ATP6, MT-CO2, MT-ND1, MT-ND2, MT-ND3, MT-ND4 |
| Categories | Functions Annotation | p-Value | Predicted Activation | Activation z-Score | Molecules |
|---|---|---|---|---|---|
| Cell Death and Survival | Apoptosis | 7.49 × 10−3 | MA | −1.296 | BCL6, BMPR1B, DAPK3, DIO3, HDAC5, PPARGC1B, TNFRSF12A, TNFRSF19, ZBTB16 |
| Organismal Survival | Organismal death | 1.91 × 10−2 | MA | −1.166 | ARID5A, BCL6, DIO3, HDAC5, KCNAB1, PPARGC1B, TNFRSF12A, ZNRF3 |
| Cell Death and Survival | Apoptosis of tumor cell lines | 8.54 × 10−3 | MA | −0.588 | BCL6, BMPR1B, DAPK3, HDAC5, PPARGC1B, ZBTB16 |
| Tissue Morphology | Quantity of cells | 4.57 × 10−2 | MA | −0.532 | ARID5A, BCL6, BMPR1B, DIO3, TNFRSF12A, ZBTB16 |
| Cellular Development, Connective Tissue Development and Function, Tissue Development | Differentiation of connective tissue cells | 1.15 × 10−4 | MA | −0.045 | ARID5A, BCL6, BMPR1B, HDAC5, PPARGC1B, ZBTB16 |
| Cellular Development, Cellular Growth and Proliferation | Proliferation of blood cells | 2.47 × 10−2 | MAT | 1.887 | BCL6, BMPR1B, TNFRSF12A, ZBTB16 |
| Cellular Development, Cellular Growth and Proliferation | Colony formation of tumor cell lines | 9.51 × 10−4 | MAT | 1.165 | BCL6, HDAC5, PPARGC1B, ZBTB16 |
| Cellular Development, Connective Tissue Development and Function, Tissue Development | Differentiation of bone cells | 8.47 × 10−4 | MAT | 0.988 | BCL6, BMPR1B, HDAC5, PPARGC1B |
| Gene Expression | Activation of DNA endogenous promoter | 6.12 × 10−3 | MAT | 0.981 | ARID5A, BCL6, BMPR1B, HDAC5, PPARGC1B, ZBTB16 |
| Lipid Metabolism, Molecular Transport, Small Molecule Biochemistry | Concentration of lipid | 4.23 × 10−3 | MAT | 0.638 | DIO3, HDAC5, PPARGC1B, TNFRSF12A, ZBTB16 |
| Gene Expression | Transcription of RNA | 1.05 × 10−2 | MAT | 0.403 | ARID5A, BCL6, BMPR1B, DAPK3, HDAC5, PARGC1B, ZBTB16 |
| Cellular Development, Cellular Growth and Proliferation | Cell proliferation of tumor cell lines | 3.96 × 10−2 | MAT | 0.352 | BCL6, BMPR1B, DAPK3, HDAC5, PPARGC1B, ZBTB16 |
| Lipid Metabolism, Molecular Transport, Small Molecule Biochemistry | Quantity of steroid | 2.87 × 10−3 | MAT | 0.152 | DIO3, PPARGC1B, TNFRSF12A, ZBTB16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Núñez, Y.; Radović, Č.; Savić, R.; García-Casco, J.M.; Čandek-Potokar, M.; Benítez, R.; Radojković, D.; Lukić, M.; Gogić, M.; Muñoz, M.; et al. Muscle Transcriptome Analysis Reveals Molecular Pathways Related to Oxidative Phosphorylation, Antioxidant Defense, Fatness and Growth in Mangalitsa and Moravka Pigs. Animals 2021, 11, 844. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11030844
Núñez Y, Radović Č, Savić R, García-Casco JM, Čandek-Potokar M, Benítez R, Radojković D, Lukić M, Gogić M, Muñoz M, et al. Muscle Transcriptome Analysis Reveals Molecular Pathways Related to Oxidative Phosphorylation, Antioxidant Defense, Fatness and Growth in Mangalitsa and Moravka Pigs. Animals. 2021; 11(3):844. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11030844
Chicago/Turabian StyleNúñez, Yolanda, Čedomir Radović, Radomir Savić, Juan M. García-Casco, Marjeta Čandek-Potokar, Rita Benítez, Dragan Radojković, Miloš Lukić, Marija Gogić, María Muñoz, and et al. 2021. "Muscle Transcriptome Analysis Reveals Molecular Pathways Related to Oxidative Phosphorylation, Antioxidant Defense, Fatness and Growth in Mangalitsa and Moravka Pigs" Animals 11, no. 3: 844. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11030844