
Molecular Diagnosis of Koala Retrovirus (KoRV) in South Australian Koalas (Phascolarctos cinereus)

 , , ,
, , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. DNA Analysis for KoRV Provirus: DNA Extraction
2.3. DNA Analysis for KoRV Provirus: DNA Proviral PCR and qPCR
2.4. RNA Extraction and Sequencing
2.5. KoRV Gene Expression from RNA-seq Analysis
2.6. BLAST Analysis
2.7. Viral Two Step RT-qPCR Analysis
2.8. Relative qPCR Statistical Analysis
3. Results
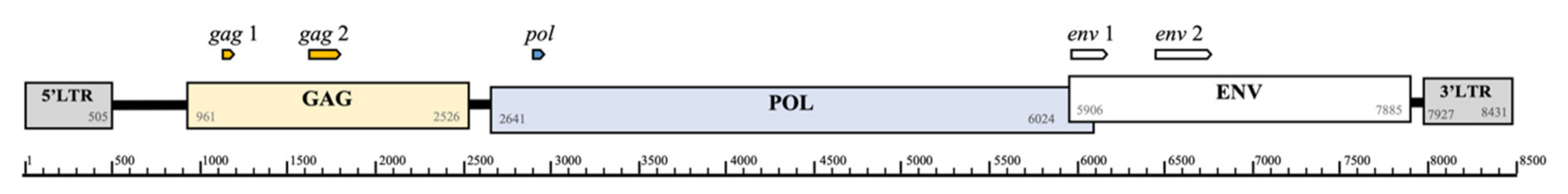
3.1. Proviral DNA Analyses
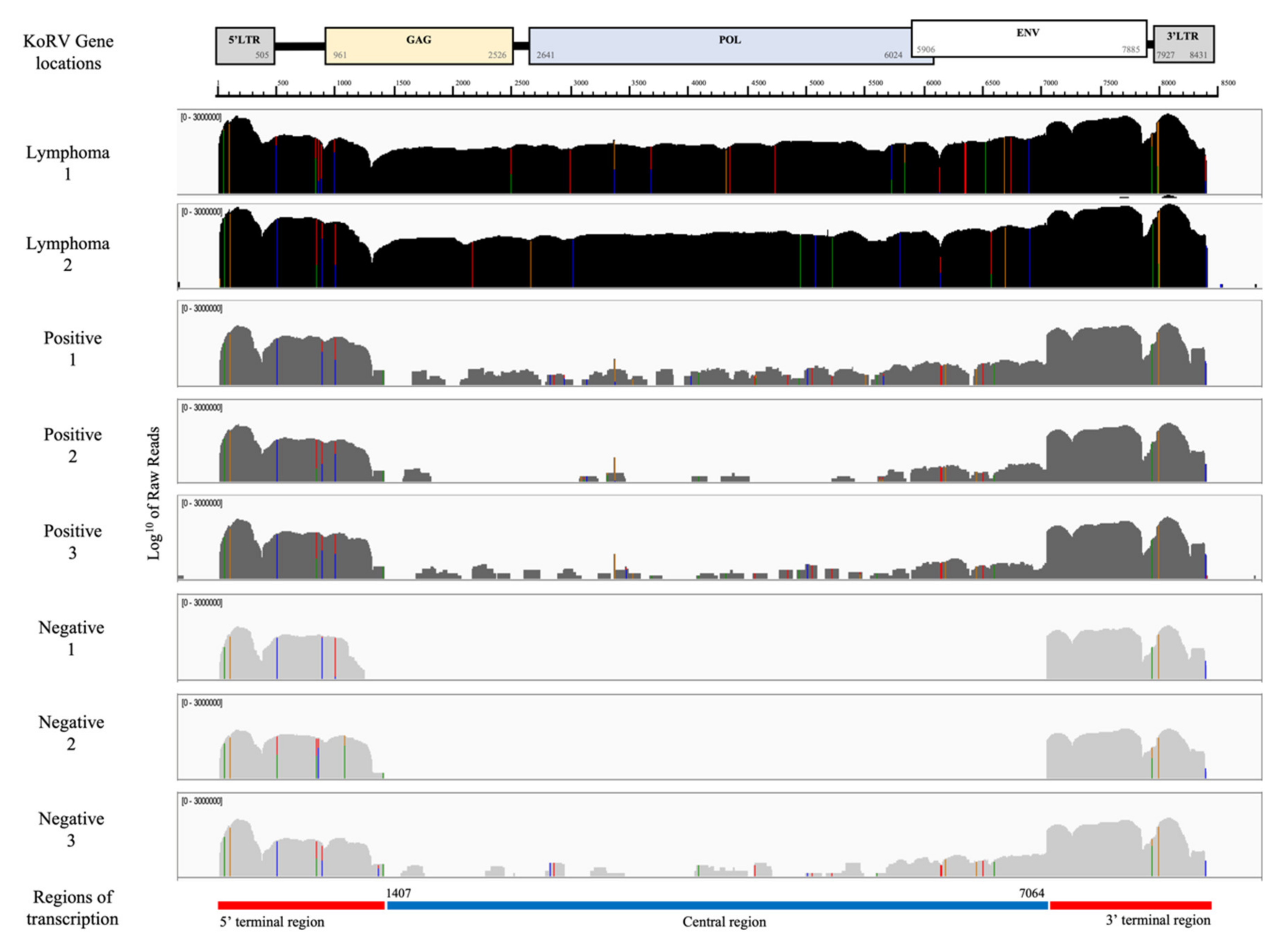
3.2. KoRV Gene Expression from RNA-seq Analysis
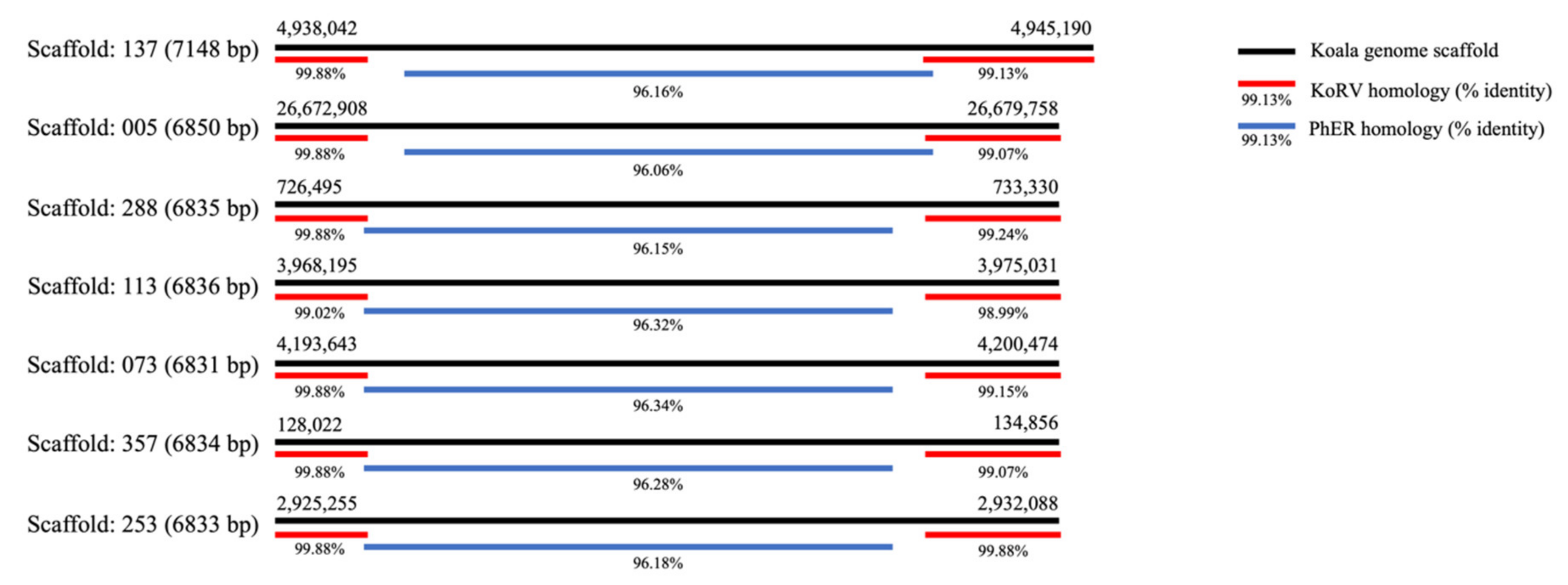
3.3. Terminal Regions of Homology to KoRV
3.4. KoRV Gene Expression from Viral Two Step RT-qPCR Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanger, J.J.; Bromham, L.D.; McKee, J.J.; O’Brien, T.M.; Robinson, W.F. The nucleotide sequence of koala (Phascolarctos cinereus) retrovirus: A novel type C endogenous virus related to gibbon ape leukemia virus. J. Virol. 2000, 74, 4264–4272. [Google Scholar] [CrossRef] [Green Version]
- Vogt, V.M. Retroviral Virions and Genomes. In Retroviruses; Coffin, J.M., Hughes, S.H., Varmus, H.E., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997. [Google Scholar]
- Shojima, T.; Hoshino, S.; Abe, M.; Yasuda, J.; Shogen, H.; Kobayashi, T.; Miyazawa, T. Construction and characterization of an infectious molecular clone of koala retrovirus. J. Virol. 2013, 87, 5081–5088. [Google Scholar] [CrossRef] [Green Version]
- Shojima, T.; Yoshikawa, R.; Hoshino, S.; Shimode, S.; Nakagawa, S.; Ohata, T.; Nakaoka, R.; Miyazawa, T. Identification of a Novel Subgroup of Koala Retrovirus from Koalas in Japanese Zoos. J. Virol. 2013, 87, 9943–9948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Stadler, C.K.; Gorman, K.; Jensen, N.; Kim, D.; Zheng, H.; Tang, S.; Switzer, W.M.; Pye, G.W.; Eiden, M.V. An exogenous retrovirus isolated from koalas with malignant neoplasias in a US zoo. Proc. Natl. Acad. Sci. USA 2013, 110, 11547–11552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimode, S.; Nakagawa, S.; Yoshikawa, R.; Shojima, T.; Miyazawa, T. Heterogeneity of koala retrovirus isolates. FEBS Lett. 2014, 588, 41–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.Q.; Gorman, K.; Santiago, J.C.; Kluska, K.; Eiden, M.V. Genetic diversity of koala retroviral envelopes. Viruses 2015, 7, 1258. [Google Scholar] [CrossRef] [Green Version]
- Chappell, K.J.; Brealey, J.C.; Amarilla, A.A.; Watterson, D.; Hulse, L.; Palmieri, C.; Johnston, S.D.; Holmes, E.C.; Meers, J.; Young, P.R. Phylogenetic Diversity of Koala Retrovirus within a Wild Koala Population. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Simmons, G.; Young, P.; McKee, J.; Meers, J.; Mizuno, T. The epidemiology of Koala retrovirus. J. Vet. Epidemiol. 2011, 15, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Simmons, G.S.; Young, P.R.; Hanger, J.J.; Jones, K.; Clarke, D.; McKee, J.J.; Meers, J. Prevalence of koala retrovirus in geographically diverse populations in Australia. Aust. Vet. J. 2012, 90, 404–409. [Google Scholar] [CrossRef]
- Stoye, J.P. Koala retrovirus: A genome invasion in real time. Genome Biol. 2006, 7, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, N.M.; Farrell, K.B.; Eiden, M.V. In vitro characterization of a koala retrovirus. J. Virol. 2006, 80, 3104–3107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabijan, J.; Miller, D.; Olagoke, O.; Woolford, L.; Boardman, W.; Timms, P.; Polkinghorne, A.; Simmons, G.; Hemmatzadeh, F.; Trott, D.J.; et al. Prevalence and clinical significance of koala retrovirus in two South Australian koala (Phascolarctos cinereus) populations. J. Med. Microbiol. 2019, 68, 1072–1080. [Google Scholar] [CrossRef] [PubMed]
- Fiebig, U.; Hartmann, M.G.; Bannert, N.; Kurth, R.; Denner, J. Transspecies transmission of the endogenous koala retrovirus. J. Virol. 2006, 80, 5651–5654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legione, A.R.; Patterson, J.L.; Whiteley, P.; Firestone, S.M.; Curnick, M.; Bodley, K.; Lynch, M.; Gilkerson, J.R.; Sansom, F.M.; Devlin, J.M. Koala retrovirus genotyping analyses reveal a low prevalence of KoRV-A in Victorian koalas and an association with clinical disease. J. Med. Microbiol. 2017, 66, 236–244. [Google Scholar] [CrossRef]
- Sarker, N.; Fabijan, J.; Seddon, J.; Tarlinton, R.; Owen, H.; Simmons, G.; Thia, J.; Blanchard, A.M.; Speight, N.; Kaler, J.; et al. Genetic diversity of Koala retrovirus env gene subtypes: Insights into northern and southern koala populations. J. Gen. Virol. 2019, 100, 1328–1339. [Google Scholar] [CrossRef] [PubMed]
- Sarker, N.; Fabijan, J.; Owen, H.; Seddon, J.; Simmons, G.; Speight, N.; Kaler, J.; Woolford, L.; Emes, R.D.; Hemmatzadeh, F.; et al. Koala retrovirus viral load and disease burden in distinct northern and southern koala populations. Sci. Rep. 2020, 10, 263. [Google Scholar] [CrossRef] [PubMed]
- Tarlinton, R.; Meers, J.; Young, P. Biology and evolution of the endogenous koala retrovirus. Cell. Mol. Life Sci. 2008, 65, 3413–3421. [Google Scholar] [CrossRef] [PubMed]
- Tarlinton, R.E.; Sarker, N.; Fabijan, J.; Dottorini, T.; Woolford, L.; Meers, J.; Simmons, G.; Owen, H.; Seddon, J.M.; Hemmatzedah, F.; et al. Differential and defective expression of Koala Retrovirus reveal complexity of host and virus evolution. bioRxiv 2017. [Google Scholar] [CrossRef] [Green Version]
- Lober, U.; Hobbs, M.; Dayaram, A.; Tsangaras, K.; Jones, K.; Alquezar-Planas, D.E.; Ishida, Y.; Meers, J.; Mayer, J.; Quedenau, C.; et al. Degradation and remobilization of endogenous retroviruses by recombination during the earliest stages of a germ-line invasion. Proc. Natl. Acad. Sci. USA 2018, 115, 8609–8614. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Koppetsch, B.S.; Pagliarani, S.; Johnston, S.; Silverstein, N.J.; Luban, J.; Chappell, K.; Weng, Z.; Theurkauf, W.E. The piRNA Response to Retroviral Invasion of the Koala Genome. Cell 2019, 179, 632–643. [Google Scholar] [CrossRef]
- Fabijan, J.; Woolford, L.; Lathe, S.; Simmons, G.; Hemmatzadeh, F.; Trott, D.J.; Speight, N. Lymphoma, Koala Retrovirus Infection and Reproductive Chlamydiosis in a Koala (Phascolarctos cinereus). J. Comp. Pathol. 2017, 157, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Tarlinton, R.; Meers, J.; Hanger, J.; Young, P. Real-time reverse transcriptase PCR for the endogenous koala retrovirus reveals an association between plasma viral load and neoplastic disease in koalas. J. Gen. Virol. 2005, 86, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Waugh, C.A.; Hanger, J.; Loader, J.; King, A.; Hobbs, M.; Johnson, R.; Timms, P. Infection with koala retrovirus subgroup B (KoRV-B), but not KoRV-A, is associated with chlamydial disease in free-ranging koalas (Phascolarctos cinereus). Sci. Rep. 2017, 7, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabijan, J.; Sarker, N.; Speight, N.; Owen, H.; Meers, J.; Simmons, G.; Seddon, J.; Emes, R.D.; Tarlinton, R.; Hemmatzadeh, F.; et al. Pathological Findings in Koala Retrovirus-positive Koalas (Phascolarctos cinereus) from Northern and Southern Australia. J. Comp. Pathol. 2020, 176, 50–66. [Google Scholar] [CrossRef]
- Krueger, F. Trim Galore: A Wrapper Tool Around Cutadapt and FastQC to consistently Apply Quality And Adapter Trimming to FastQ Files. Available online: https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/ (accessed on 8 October 2019).
- Schubert, M.; Lindgreen, S.; Orlando, L. AdapterRemoval v2: Rapid adapter trimming, identification, and read merging. BMC Res. Notes 2016, 9, 88. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 8 October 2019).
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Coordinators, N.R. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2018, 46, D8–D13. [Google Scholar] [CrossRef] [Green Version]
- Stanton, K.A.; Edger, P.P.; Puzey, J.R.; Kinser, T.; Cheng, P.; Vernon, D.M.; Forsthoefel, N.R.; Cooley, A.M. A Whole-Transcriptome Approach to Evaluating Reference Genes for Quantitative Gene Expression Studies: A Case Study in Mimulus. G3 Genes Genomes Genet. 2017, 7, 1085–1095. [Google Scholar] [CrossRef]
- Hobbs, M.; King, A.; Salinas, R.; Chen, Z.; Tsangaras, K.; Greenwood, A.D.; Johnson, R.N.; Belov, K.; Wilkins, M.R.; Timms, P. Long-read genome sequence assembly provides insight into ongoing retroviral invasion of the koala germline. Sci. Rep. 2017, 7, 15838. [Google Scholar] [CrossRef] [Green Version]
- Hanger, J. An Investigation of the Role of Retroviruses in Leukaemia and Related Diseases in Koalas; University of Queensland: Brisbane, Australia, 1999. [Google Scholar]
- Kinney, M.E.; Pye, G.W. Koala Retrovirus: A Review. J. Zoo Wildl. Med. 2016, 47, 387–396. [Google Scholar] [CrossRef] [PubMed]
- McEwen, G.K.; Alquezar-Planas, D.E.; Dayaram, A.; Gillett, A.; Tarlinton, R.; Mongan, N.; Chappell, K.J.; Henning, J.; Tan, M.; Timms, P.; et al. Retroviral integrations contribute to elevated host cancer rates during germline invasion. Nat. Commun. 2021, 12, 1316. [Google Scholar] [CrossRef] [PubMed]
- Quigley, B.L.; Ong, V.A.; Hanger, J.; Timms, P.; Simon, V. Molecular Dynamics and Mode of Transmission of Koala Retrovirus as It Invades and Spreads through a Wild Queensland Koala Population. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Ahlenstiel, C.; Mendez, C.; Lim, S.T.; Marks, K.; Turville, S.; Cooper, D.A.; Kelleher, A.D.; Suzuki, K. Novel RNA Duplex Locks HIV-1 in a Latent State via Chromatin-mediated Transcriptional Silencing. Mol. Ther. Nucleic Acids 2015, 4, e261. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Wang, P.J. Mammalian piRNAs: Biogenesis, function, and mysteries. Spermatogenesis 2014, 4, e27889. [Google Scholar] [CrossRef]
- Tarazona, S.; Garcia-Alcalde, F.; Dopazo, J.; Ferrer, A.; Conesa, A. Differential expression in RNA-seq: A matter of depth. Genome Res. 2011, 21, 2213–2223. [Google Scholar] [CrossRef] [Green Version]
- Goff, S.P. Cellular Factors That Regulate Retrovirus Uncoating and Reverse Transcription. In Retrovirus-Cell Interactions; Parent, L.J., Ed.; Academic Press: London, UK, 2018; pp. 51–112. [Google Scholar]
- Olagoke, O.; Miller, D.; Hemmatzadeh, F.; Stephenson, T.; Fabijan, J.; Hutt, P.; Finch, S.; Speight, N.; Timms, P. Induction of neutralizing antibody response against koala retrovirus (KoRV) and reduction in viral load in koalas following vaccination with recombinant KoRV envelope protein. NPJ Vaccines 2018, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Nitta, T.; Ha, D.; Galvez, F.; Miyazawa, T.; Fan, H. Human and murine APOBEC3s restrict replication of koala retrovirus by different mechanisms. Retrovirology 2015, 12, 68. [Google Scholar] [CrossRef] [Green Version]
- Balasuriya, U.B.R.; Barratt-Boyes, S.; Beer, M.; Bird, B.; Brownlie, J.; Coffey, L.L.; Cullen, J.M.; Delhon, G.A.; Donis, R.O.; Gardner, I.; et al. Retroviridae. In Fenner’s Veterinary Virology, 5th ed.; MacLachlan, N.J., Dubovi, E.J., Eds.; Academic Press: London, UK, 2017; pp. 269–297. [Google Scholar]
- Greenwood, A.D.; Ishida, Y.; O’Brien, S.P.; Roca, A.L.; Eiden, M.V. Transmission, Evolution, and Endogenization: Lessons Learned from Recent Retroviral Invasions. Microbiol. Mol. Biol. Rev. 2018, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magiorkinis, G.; Gifford, R.J.; Katzourakis, A.; De Ranter, J.; Belshaw, R. Env-less endogenous retroviruses are genomic superspreaders. Proc. Natl. Acad. Sci. USA 2012, 109, 7385–7390. [Google Scholar] [CrossRef] [Green Version]
- Mcdougall, A.S.; Terry, A.; Tzavaras, T.; Cheney, C.; Rojko, J.; Neil, J.C. Defective Endogenous Proviruses Are Expressed in Feline Lymphoid Cells: Evidence for a Role in Natural Resistance to Subgroup B Feline Leukemia Viruses. J. Virol. 1994, 68, 2151–2160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nethe, M.; Berkhout, B.; van der Kuyl, A.C. Retroviral superinfection resistance. Retrovirology 2005, 2, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.; Yan, Y.; Kozak, C.A. Rmcf2, a xenotropic provirus in the Asian mouse species Mus castaneus, blocks infection by polytropic mouse gammaretroviruses. J. Virol. 2005, 79, 9677–9684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoye, J.P. Studies of endogenous retroviruses reveal a continuing evolutionary saga. Nat. Rev. Microbiol. 2012, 10, 395–406. [Google Scholar] [CrossRef]
- Robinson, H.L.; Lamoreux, W.F. Expression of Endogenous ALV Antigens and Susceptibility to Subgroup E ALV in Three Strains of Chickens (Endogenous Avian C-Type Virus). Virology 1976, 69, 50–62. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Target | Primers | Product Size | Reference |
|---|---|---|---|
| Koala ß-actin | Fwd 5′-GAGACCTTCAACACCCCAGC-3′ Rev 5′-GTGGGTCACACCATCACCAG-3′ | 111 bp | Shojima et al. 2013 [4] |
| KoRV gag 1 | Fwd 5′-CGGACCAGGTTCCCTACATC-3′ Rev 5′- TCGCCCGTTATCTTGACCAG-3′ | 110 bp | This study |
| KoRV gag 2 | Fwd 5′-TTGGCCTTTCTCCTCAGCAG-3′ Rev 5′-CCGTGTTGTGATCCCACTGA-3′ | 290 bp | This study |
| KoRV pol | Fwd 5′-TTGGAGGAGGAATACCGATTACAC-3′ Rev 5′-GCCAGTCCCATACCTGCCTT-3′ | 111 bp | Tarlinton et al. 2005 [23] |
| KoRV-A env 1 | Fwd 5′-TCCTGGGAACTGGAAAAGAC-3′ Rev 5′-GGGTTCCCCAAGTGATCTG-3′ | 321 bp | Waugh et al. 2017 [24] |
| KoRV env 2 | Fwd 5′-GCCCTCGGCCCTCCTTATTA-3′ Rev 5′-GGCAATCTGGAGGCTAGTCAA-3′ | 522 bp | Sarker et al. 2020 [17] |
| KoRV Gene Target | Count (n = 216) | % | Median (Range) of Proviral Load KoRV Copies/ 103 ß-Actin Copies * | ||||
|---|---|---|---|---|---|---|---|
| gag 1 | gag 2 | pol | env 1 | env 2 | |||
| + | + | + | + | + | 89 | 41.2 | 41 (0.2–270) |
| + | - | - | - | - | 123 | 56.9 | 0 |
| + | - | - | - | + | 2 | 0.9 | 0 |
| + | - | - | + | - | 1 | 0.5 | 0 |
| - | - | - | - | - | 1 | 0.5 | 0 |
| KoRV Gene | 2−∆∆CT Lymphomic Koalas Relative to Positive Koalas | ||
|---|---|---|---|
| Mean FC * | FC Range | p Value | |
| gag 2 | 299 | 47.7–1880.5 | 0.060 |
| pol | 602 | 185.8–1950.2 | 0.012 |
| env 1 | 242 | 42.5–1375.6 | 0.012 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stephenson, T.; Speight, N.; Low, W.Y.; Woolford, L.; Tearle, R.; Hemmatzadeh, F. Molecular Diagnosis of Koala Retrovirus (KoRV) in South Australian Koalas (Phascolarctos cinereus). Animals 2021, 11, 1477. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11051477
Stephenson T, Speight N, Low WY, Woolford L, Tearle R, Hemmatzadeh F. Molecular Diagnosis of Koala Retrovirus (KoRV) in South Australian Koalas (Phascolarctos cinereus). Animals. 2021; 11(5):1477. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11051477
Chicago/Turabian StyleStephenson, Tamsyn, Natasha Speight, Wai Yee Low, Lucy Woolford, Rick Tearle, and Farhid Hemmatzadeh. 2021. "Molecular Diagnosis of Koala Retrovirus (KoRV) in South Australian Koalas (Phascolarctos cinereus)" Animals 11, no. 5: 1477. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11051477