
Stable Isotopes Reveal the Dominant Species to Have the Widest Trophic Niche of Three Syntopic Microtus Voles

 ,
,  ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
- Interspecific differences of stable isotope distribution in common (Microtus arvalis), field (M. agrestis), and root (M. oeconomus) voles do exist, with the trophic niche of the dominant species, the common vole, being widest,
- The width of the trophic niche of the dominant species is related to the presence of syntopically co-occurring Microtus species,
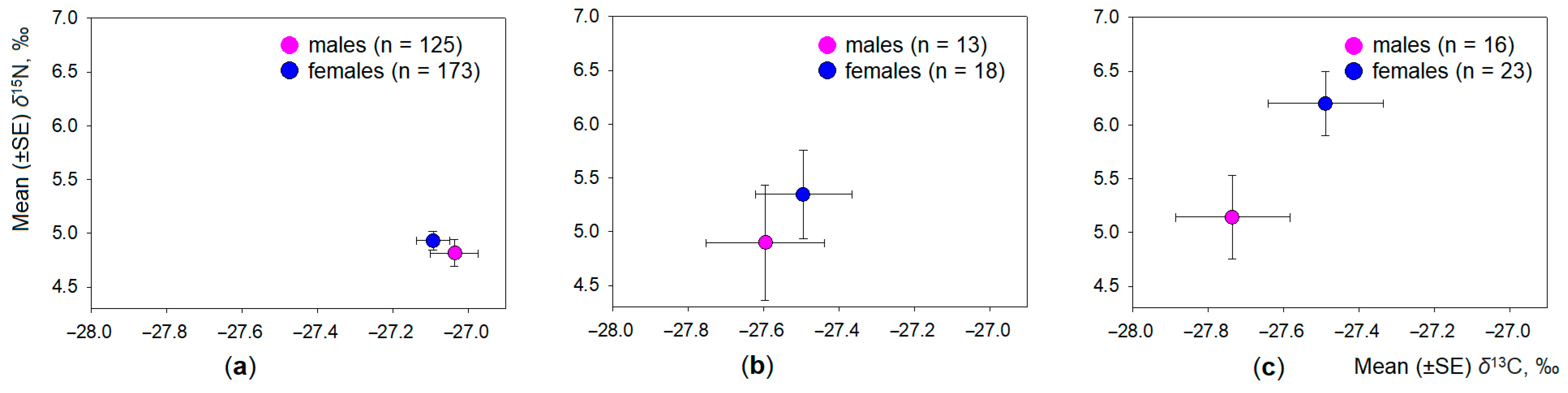
- Intraspecific differences of the stable isotope distribution are absent, with no differences in average carbon and nitrogen stable isotope values in males and females and between the age groups of the same species.
2. Material and Methods
2.1. Study Sites and Small Mammal Trapping
2.2. Stable Isotope Analysis
2.3. Statistical Analyses
3. Results
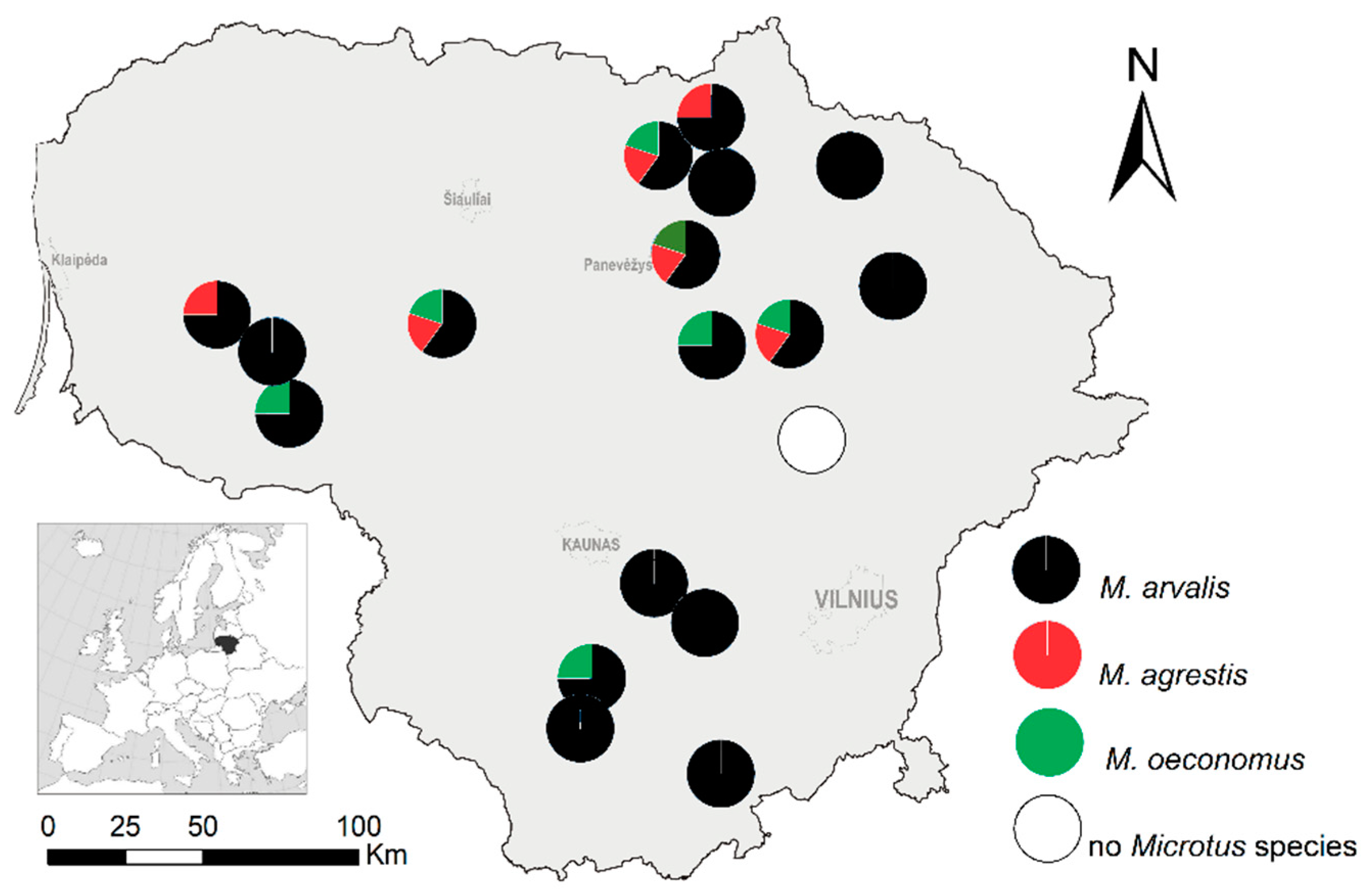
3.1. Distribution of Co-Occurring Microtus Populations
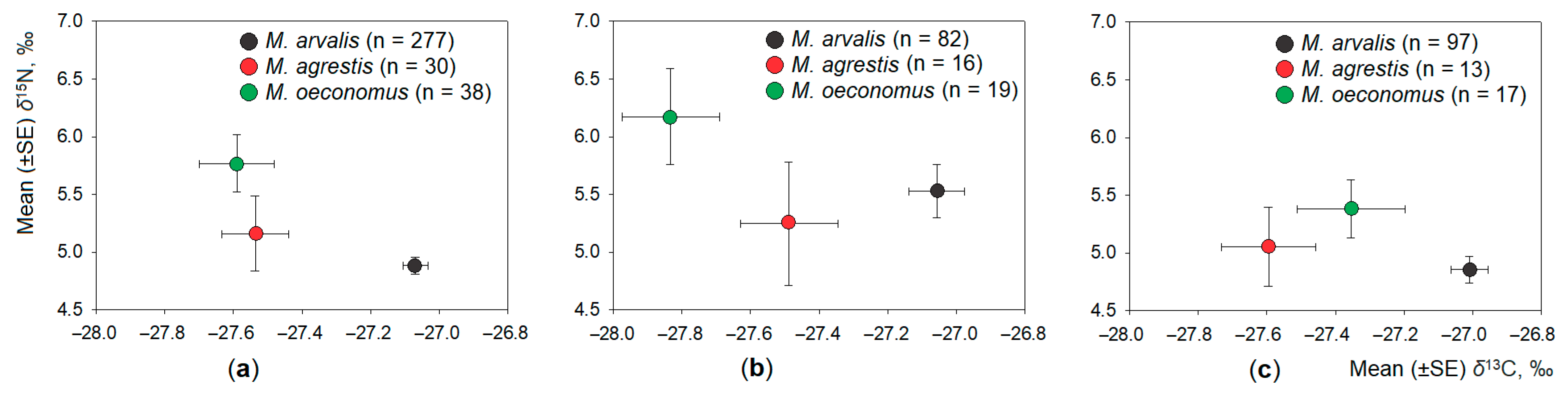
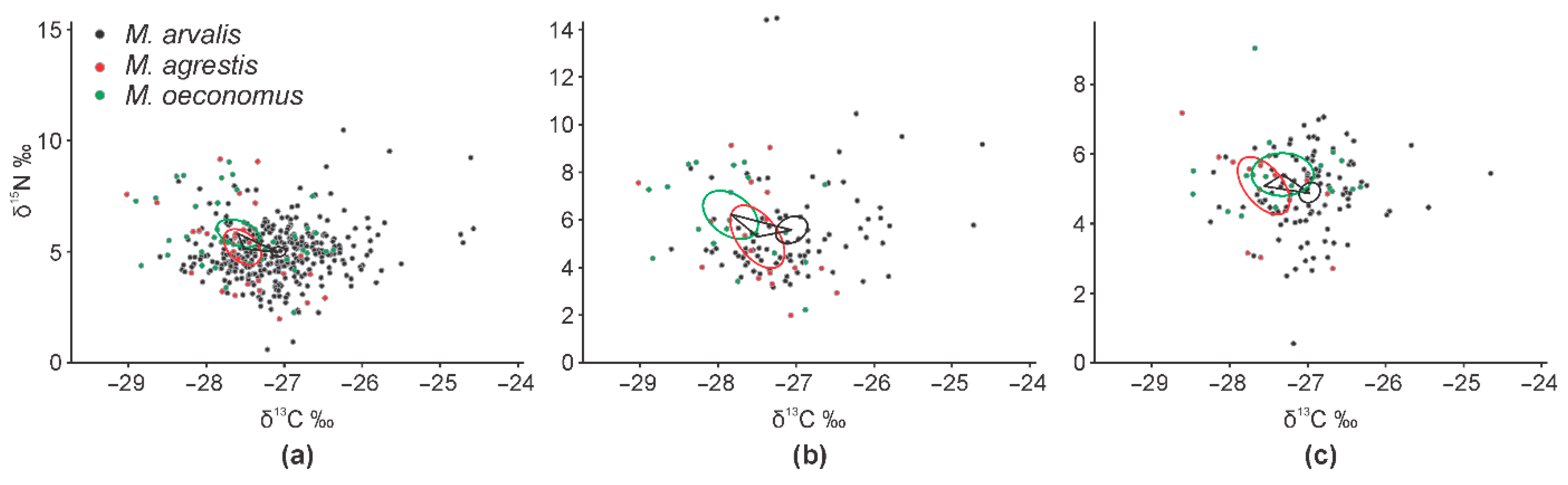
3.2. Interspecific Differences in Dietary Space between Three Microtus Vole Species
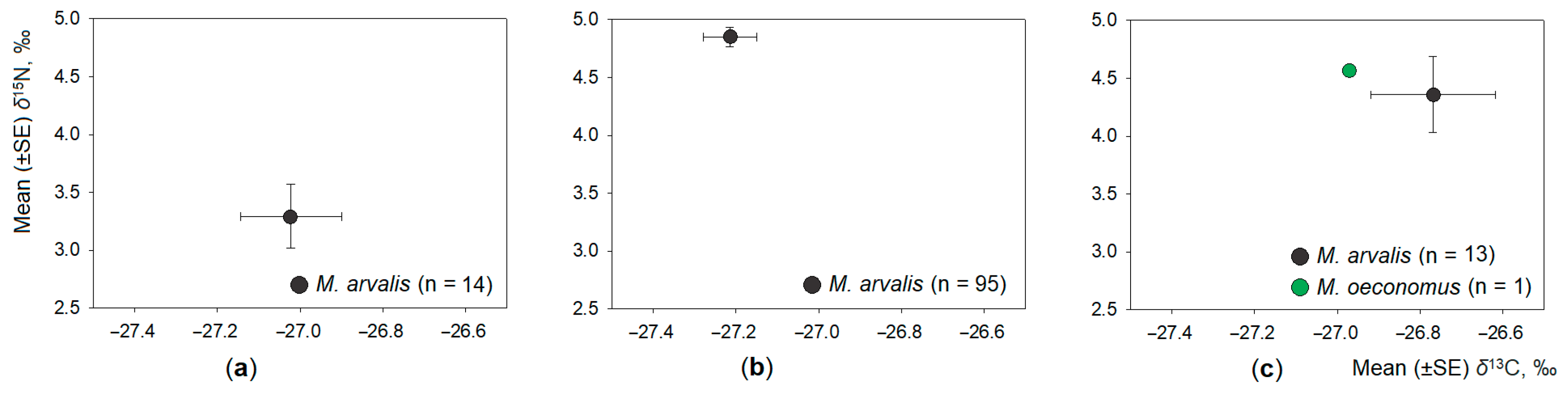
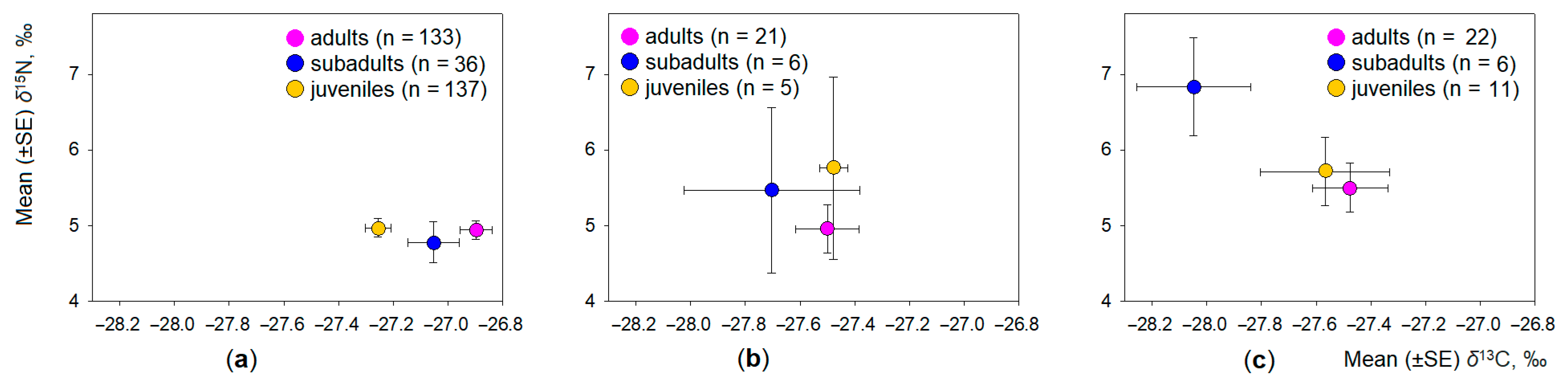
3.3. Intraspecific Differences in Dietary Space of Microtus Voles
4. Discussion
4.1. Trophic Niche of Microtus Voles in Different Habitats
4.2. Possible Factors Influencing Changes in Trophic Niche
4.3. Role of Community and Intraspecific Patterns in Defining Trophic Niche
4.4. Specificity of Agricultural Habitats to Microtus Voles
5. Conclusions
- Shown on sympatric and syntopic Microtus voles, our results highlight the importance of interspecific competition for interpreting patterns of habitat selection and resource sharing among coexisting herbivore species.
- The widest trophic niche was characteristic to the dominant species, common vole.
- In the case of co-occurrence with other Microtus species, the width of the trophic niche of the common vole increased, separating the species according to δ15N values.
- Intraspecific differences in the dietary space were best expressed in the dominant common vole (differences according to δ13C, but not δ15N values) and in the root vole (according δ15N values).
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ishikawa, N.F. Use of compound-specific nitrogen isotope analysis of amino acids in trophic ecology: Assumptions, applications, and implications. Ecol. Res. 2018, 33, 825–837. [Google Scholar] [CrossRef]
- Whitaker, M.R.; Baker, C.C.; Salzman, S.M.; Martins, D.J.; Pierce, N.E. Combining stable isotope analysis with DNA metabarcoding improves inferences of trophic ecology. PLoS ONE 2019, 14, e0219070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galetti, M.; Rodarte, R.R.; Neves, C.L.; Moreira, M.; Costa-Pereira, R. Trophic niche differentiation in rodents and marsupials revealed by stable isotopes. PLoS ONE 2016, 11, e0152494. [Google Scholar] [CrossRef]
- Gasperini, S.; Bonacchi, A.; Bartolommei, P.; Manzo, E.; Cozzolino, R. Seasonal cravings: Plant food preferences of syntopic small mammals. Ethol. Ecol. Evol. 2017, 30, 12–25. [Google Scholar] [CrossRef]
- Verde Arregoitia, L.D.; D’Elía, G. Classifying rodent diets for comparative research. Mammal. Rev. 2021, 51, 51–65. [Google Scholar] [CrossRef]
- Cramer, M.J.; Willig, M.R. Habitat heterogeneity, species diversity and null models. Oikos 2005, 108, 209–218. [Google Scholar] [CrossRef]
- Hart, K.M.; Iverson, A.R.; Fujisaki, I.; Lamont, M.M.; Bucklin, D.; Shaver, D.J. Sympatry or syntopy? Investigating drivers of distribution and co-occurrence for two imperiled sea turtle species in Gulf of Mexico neritic waters. Ecol. Evol. 2018, 8, 12656–12669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauffrey, J.F.; Catzeflis, F. Ecological and isotopic discrimination of syntopic rodents in a neotropical rain forest of French Guiana. J. Trop. Ecol. 2003, 19, 209–214. Available online: https://0-www-jstor-org.brum.beds.ac.uk/stable/4092159 (accessed on 12 February 2021). [CrossRef]
- Butet, A.; Delettre, Y.R. Diet differentiation between European arvicoline and murine rodents. Acta Theriol. 2011, 56, 297. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Skipitytė, R.; Balčiauskienė, L.; Jasiulionis, M. Resource partitioning confirmed by isotopic signatures allows small mammals to share seasonally flooded meadows. Ecol. Evol. 2019, 9, 5479–5489. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Balčiauskienė, L.; Stirkė, V. Mow the Grass at the Mouse’s Peril: Diversity of Small Mammals in Commercial Fruit Farms. Animals 2019, 9, 334. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.M. Seasonal variations in the body composition and nutrition of the vole Microtus agrestis. J. Anim. Ecol. 1973, 42, 1–18. [Google Scholar] [CrossRef]
- Yigit, N.; Hutterer, R.; Kryštufek, B.; Amori, G. Microtus arvalis. The IUCN Red List of Threatened Species. 2016. Available online: https://www.iucnredlist.org/species/13488/22351133 (accessed on 16 June 2021).
- Linzey, A.V.; Shar, S.; Lkhagvasuren, D.; Juškaitis, R.; Sheftel, B.; Meinig, H.; Amori, G.; Henttonen, H. Microtus oeconomus (errata version published in 2017). The IUCN Red List of Threatened Species. 2016. Available online: https://www.iucnredlist.org/species/13451/115113894 (accessed on 16 June 2021).
- Kryštufek, B.; Vohralík, V.; Zima, J.; Zagorodnyuk, I. Microtus agrestis (errata version published in 2017). The IUCN Red List of Threatened Species. 2016. Available online: https://www.iucnredlist.org/species/13426/115112050 (accessed on 16 June 2021).
- Heroldová, M. Food selection of Microtus agrestis in air-pollution affected clearings in the Beskydy Mts, Czech Republic. Folia Zool. 2002, 51, 83–91. [Google Scholar]
- Ostfeld, R.S. Limiting resources and territoriality in microtine rodents. Am. Nat. 1985, 126, 1–15. [Google Scholar] [CrossRef]
- Soininen, E.M.; Ravolainen, V.T.; Bräthen, K.A.; Yoccoz, N.G.; Gielly, L.; Ims, R.A. Arctic Small Rodents Have Diverse Diets and Flexible Food Selection. PLoS ONE 2013, 8, e68128. [Google Scholar] [CrossRef] [Green Version]
- Fortin, D.; Boyce, M.S.; Merrill, E.H. Multi-tasking by mammalian herbivores: Overlapping processes during foraging. Ecology 2004, 85, 2312–2322. [Google Scholar] [CrossRef]
- Romanowski, J.; Lesiński, G.; Bardzińska, M. Small mammals of the suburban areas of Warsaw in the diet of the tawny owl Strix Aluco. Studia Ecol. Bioethicae 2020, 18, 349–354. [Google Scholar] [CrossRef]
- Sîtnic, V.; Nistreanu, V.; Larion, A.; Savin, A. The demographic structure of Microtus arvalis and Microtus rossiaemeridionalis (Mammalia, Rodentia, Cricetidae) populations in agrocenoses from the Republic of Moldova. Olten. Stud. Si Comun. Stiintele Nat. 2020, 36, 129–136. [Google Scholar]
- Magioli, M.; Moreira, M.Z.; Fonseca, R.C.B.; Ribeiro, M.C.; Rodrigues, M.G.; de Barros, K.M.P.M. Human-modified landscapes alter mammal resource and habitat use and trophic structure. Proc. Natl. Acad. Sci. USA 2019, 116, 18466–18472. [Google Scholar] [CrossRef] [Green Version]
- Balčiauskas, L.; Balčiauskienė, L. On the Doorstep, Rodents in Homesteads and Kitchen Gardens. Animals 2020, 10, 856. [Google Scholar] [CrossRef] [PubMed]
- Stirkė, V.; Balčiauskas, L.; Balčiauskienė, L. Common Vole as a Focal Small Mammal Species in Orchards of the Northern Zone. Diversity 2021, 13, 134. [Google Scholar] [CrossRef]
- Winfree, R.; Bartomeus, I.; Cariveau, D.P. Native pollinators in anthropogenic habitats. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Guzmán, J.M.; Morán, R.; Masero, J.A.; Corbacho, C.; Costillo, E.; Villegas, A.; Santiago-Quesada, F. Identifying new buffer areas for conserving waterbirds in the Mediterranean basin: The importance of the rice fields in Extremadura, Spain. Biodivers. Conserv. 2007, 16, 3333–3344. [Google Scholar] [CrossRef]
- Pianka, E.R. Niche overlap and diffuse competition. Proc. Natl. Acad. Sci. USA 1974, 71, 2141–2145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa-Pereira, R.; Araújo, M.S.; Souza, F.L.; Ingram, T. Competition and resource breadth shape niche variation and overlap in multiple trophic dimensions. Proc. R. Soc. B 2019, 286, 20190369. [Google Scholar] [CrossRef] [Green Version]
- Manlick, P.J.; Pauli, J.N. Human disturbance increases trophic niche overlap in terrestrial carnivore communities. Proc. Natl. Acad. Sci. USA 2020, 117, 26842–26848. [Google Scholar] [CrossRef]
- Krause, A.; Sandmann, D.; Bluhm, S.L.; Ermilov, S.; Widyastuti, R.; Haneda, N.F.; Scheu, S.; Maraun, M. Shift in trophic niches of soil microarthropods with conversion of tropical rainforest into plantations as indicated by stable isotopes (15N, 13C). PLoS ONE 2019, 14, e0224520. [Google Scholar] [CrossRef] [Green Version]
- Koplin, J.R.; Hoffmann, R.S. Habitat overlap and competitive exclusion in voles (Microtus). Am. Midl. Nat. 1968, 80, 494–507. [Google Scholar] [CrossRef]
- Myllymäki, A. Interactions between the field vole Microtus agrestis and its microtine competitors in Central-Scandinavian populations. Oikos 1977, 29, 570–580. [Google Scholar] [CrossRef]
- Santos, S.M.; da Luz Mathias, M.; Mira, A.P. Local coexistence and niche differences between the Lusitanian and Mediterranean pine voles (Microtus lusitanicus and M. duodecimcostatus). Ecol. Res. 2010, 25, 1019–1031. [Google Scholar] [CrossRef] [Green Version]
- Jancewicz, E.; Gliwicz, J. Niche dynamics and biodiversity: Many rodent species on one marshy meadow. Pol. J. Ecol. 2017, 65, 371–379. [Google Scholar] [CrossRef]
- Koivisto, E.; Hoset, K.S.; Huitu, O.; Korpimäki, E. Habitat use of coexisting Microtus vole species under competition and predation risk. Can. J. Zool. 2018, 96, 237–244. [Google Scholar] [CrossRef]
- Bolnick, D.I.; Ingram, T.; Stutz, W.E.; Snowberg, L.K.; Lau, O.L.; Paull, J.S. Ecological release from interspecific competition leads to decoupled changes in population and individual niche width. Proc. R. Soc. B Biol. Sci. 2010, 277, 1789–1797. [Google Scholar] [CrossRef]
- DeNiro, M.J.; Epstein, S. Influence of diet on the distribution of carbon isotopes in animals. Geochim. Cosmochim. Acta 1978, 42, 495–506. [Google Scholar] [CrossRef]
- Matthews, D.E.; Bier, D.M. Stable Isotope Methods for Nutritional Investigation. Annu. Rev. Nutr. 1983, 3, 309–339. [Google Scholar] [CrossRef]
- Crawford, K.; Mcdonald, R.A.; Bearhop, S. Applications of stable isotope techniques to the ecology of mammals. Mammal Rev. 2008, 38, 87–107. [Google Scholar] [CrossRef]
- Polis, G.A. Age structure component of niche width and intraspecific resource partitioning: Can age groups function as ecological species? Am. Nat. 1984, 123, 541–564. [Google Scholar] [CrossRef]
- Sheppard, C.E.; Inger, R.; McDonald, R.A.; Barker, S.; Jackson, A.L.; Thompson, F.J.; Vitikainen, E.I.K.; Cant, M.A.; Marshall, H.H. Intragroup competition predicts individual foraging specialisation in a group-living mammal. Ecol. Lett. 2018, 21, 665–673. [Google Scholar] [CrossRef]
- Costa, A.F.; Botta, S.; Siciliano, S.; Giarrizzo, T. Resource partitioning among stranded aquatic mammals from Amazon and Northeastern coast of Brazil revealed through Carbon and Nitrogen Stable Isotopes. Sci. Rep. 2020, 10, 12897. [Google Scholar] [CrossRef]
- de Camargo, N.F.; Reis, G.G.; Camargo, A.C.L.; Nardoto, G.B.; Kneitel, J.M.; Vieira, E.M. Seasonal isotopic niche of a rodent: High between-individual variation but no changes in individual niche width during the rich-resource period. Biotropica 2021, 53, 966–975. [Google Scholar] [CrossRef]
- Koike, S.; Nakashita, R.; Kozakai, C.; Nakajima, A.; Nemoto, Y.; Yamazaki, K. Baseline characterization of the diet and stable isotope signatures of bears that consume natural foods in central Japan. Eur. J. Wildl. Res. 2016, 62, 23–31. [Google Scholar] [CrossRef]
- Hopkins, J.B.; Kurle, C.M. Measuring the realized niches of animals using stable isotopes: From rats to bears. Methods Ecol. Evol. 2016, 7, 210–221. [Google Scholar] [CrossRef]
- Bearhop, S.; Adams, C.E.; Waldron, S.; Fuller, R.A.; MacLeod, H. Determining trophic niche width: A novel approach using stable isotope analysis. J. Anim. Ecol. 2004, 73, 1007–1012. [Google Scholar] [CrossRef] [Green Version]
- Hette-Tronquart, N. Isotopic niche is not equal to trophic niche. Ecol. Lett. 2019, 22, 1987–1989. [Google Scholar] [CrossRef]
- Marshall, H.H.; Inger, R.; Jackson, A.L.; McDonald, R.A.; Thompson, F.J.; Cant, M.A. Stable isotopes are quantitative indicators of trophic niche. Ecol. Lett. 2019, 22, 1990–1992. [Google Scholar] [CrossRef]
- Balčiauskas, L. Methods of Investigation of Terrestrial Ecosystems. Part. I. Animal Surveys; VU Leidykla: Vilnius, Lithuania, 2004; p. 183. [Google Scholar]
- Prūsaitė, J. (Comp.). Fauna of Lithuania. Mammals; Mokslas: Vilnius, Lithuania, 1988; p. 295. [Google Scholar]
- Balciauskas, L.; Balciauskiene, L.; Janonyte, A. Reproduction of the root vole (Microtus oeconomus) at the edge of its distribution range. Turk. J. Zool. 2012, 36, 668–675. [Google Scholar]
- Metcalf, D. Multiple Thymus Grafts in Aged Mice. Nature 1965, 208, 87–88. [Google Scholar] [CrossRef] [PubMed]
- Gui, J.; Mustachio, L.M.; Su, D.M.; Craig, R.W. Thymus Size and Age-related Thymic Involution: Early Programming, Sexual Dimorphism, Progenitors and Stroma. Aging Dis. 2012, 3, 280–290. [Google Scholar]
- Rezzani, R.; Nardo, L.; Favero, G.; Peroni, M.; Rodella, L.F. Thymus and aging: Morphological, radiological, and functional overview. Age 2014, 36, 313–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balčiauskas, L.; Skipitytė, R.; Jasiulionis, M.; Balčiauskienė, L.; Remeikis, V. Immediate increase in isotopic enrichment in small mammals following the expansion of a great cormorant colony. Biogeosciences 2018, 15, 3883–3891. [Google Scholar] [CrossRef] [Green Version]
- Balčiauskas, L.; Skipitytė, R.; Jasiulionis, M.; Trakimas, G.; Balčiauskienė, L.; Remeikis, V. The impact of Great Cormorants on biogenic pollution of land ecosystems: Stable isotope signatures in small mammals. Sci. Total Environ. 2016, 565, 376–383. [Google Scholar] [CrossRef]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER—Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Yeakel, J.D.; Bhat, U.; Elliott Smith, E.A.; Newsome, S.D. Exploring the isotopic niche: Isotopic variance, physiological incorporation, and the temporal dynamics of foraging. Front. Ecol. Evol. 2016, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Missagia, R.V.; Patterson, B.D.; Perini, F.A. Stable isotope signatures and the trophic diversification of akodontine rodents. Evol. Ecol. 2019, 33, 855–872. [Google Scholar] [CrossRef]
- Ribeiro, J.F.; Guaraldo, A.; Nardoto, G.B.; Santoro, G.; Vieira, E.M. Habitat type and seasonality influence the isotopic trophic niche of small mammals in a neotropical savanna. Hystrix Ital. J. Mammal. 2019, 30, 30–38. [Google Scholar] [CrossRef]
- Mathias, M.D.L.; Hart, E.B.; Ramalhinho, M.D.G.; Jaarola, M. Microtus agrestis (Rodentia: Cricetidae). Mamm. Species 2017, 49, 23–39. [Google Scholar] [CrossRef]
- Faber, J.; Ma, W.C. Observations on seasonal dynamics in diet composition of the field vole, Microtus agrestis, with some methodological remarks. Acta Theriol. 1986, 31, 479–490. [Google Scholar] [CrossRef] [Green Version]
- Landry, S.O. The Rodentia as omnivores. Q. Rev. Biol. 1970, 45, 351–372. [Google Scholar] [CrossRef]
- Northern Zone Pesticide Risk Assessment for Birds and Mammals. Selection of Relevant Species and Development of Standard Scenarios for Higher Tier Risk Assessment in the Northern Zone in Accordance with Regulation EC 1107/2009. Version 2.1, December 2021. Available online: https://eng.mst.dk/media/211955/birds-and-mammals-higher-tier-risk-assesment-northern-zone-april-2020-ver-2-0.docx (accessed on 15 February 2021).
- Ozaki, S.; Fritsch, C.; Valot, B.; Mora, F.; Cornier, T.; Scheifler, R.; Raoul, F. Does pollution influence small mammal diet in the field? A metabarcoding approach in a generalist consumer. Mol. Ecol. 2018, 27, 3700–3713. [Google Scholar] [CrossRef]
- Hope, A.G.; Gragg, S.F.; Nippert, J.B.; Combe, F.J. Consumer roles of small mammals within fragmented native tallgrass prairie. Ecosphere 2021, 12, e03441. [Google Scholar] [CrossRef]
- Cotes, B.; Castro, J.; Cardenas, M.; Campos, M. Responses of epigeal beetles to the removal of weed cover crops in organic olive orchards. Bull. Insectol. 2009, 62, 47–52. [Google Scholar]
- Sullivan, T.P.; Sullivan, D.S.; Granatstein, D.M. Influence of living mulches on vole populations and feeding damage to apple trees. Crop. Prot. 2018, 108, 78–86. [Google Scholar] [CrossRef]
- Samnegård, U.; Alins, G.; Boreux, V.; Bosch, J.; García, D.; Happe, A.K.; Klein, A.-M.; Miñarro, M.; Mody, K.; Mario Porcel, M.; et al. Management trade-offs on ecosystem services in apple orchards across Europe: Direct and indirect effects of organic production. J. Appl. Ecol. 2019, 56, 802–811. [Google Scholar] [CrossRef] [Green Version]
- Das, B.; Kandpal, B.K.; Devi, H.L. Cover Crops for Orchard Soil Management. In Cover Crops and Sustainable Agriculture; Islam, R., Sherman, B., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 147–168. [Google Scholar]
- Reid, R.E.; Greenwald, E.N.; Wang, Y.; Wilmers, C.C. Dietary niche partitioning by sympatric Peromyscus boylii and P. californicus in a mixed evergreen forest. J. Mammal. 2013, 94, 1248–1257. [Google Scholar] [CrossRef]
- Lanszki, Z.; Horváth, G.F.; Bende, Z.; Lanszki, J. Differences in the diet and trophic niche of three sympatric carnivores in a marshland. Mammal. Res. 2020, 65, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Tucker, M.A.; Böhning-Gaese, K.; Fagan, W.F.; Fryxell, J.M.; van Moorter, B.; Alberts, S.C.; Ali, A.H.; Allen, A.M.; Attias, N.; Avgar, T.; et al. Moving in the Anthropocene: Global reductions in terrestrial mammalian movements. Science 2018, 359, 466–469. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.A.; Thomas, A.C.; Levi, T.; Wang, Y.; Wilmers, C.C. Human activity reduces niche partitioning among three widespread mesocarnivores. Oikos 2018, 127, 890–901. [Google Scholar] [CrossRef]
- Stenseth, N.C.; Hansson, L.; Myllymäki, A.; Myllymaki, A. Food Selection of the Field Vole Microtus agrestis. Oikos 1977, 29, 511. [Google Scholar] [CrossRef]
- de Jonge, G. Aggression and Group Formation in the Voles Microtus agrestis, M. arvalis and Clethrionomys glareolus in Relation to Intra- and Interspecific Competition. Behaviour 1983, 84, 1–73. [Google Scholar] [CrossRef]
- Bergeron, J.M.; Joudoin, L. Patterns of resource use, food quality, and health status of voles (Microtus pennsylvanicus) trapped from fluctuating populations. Oecologia 1989, 79, 306–314. [Google Scholar] [CrossRef]
- Primdahl, J.; Peco, B.; Schramek, J.; Andersen, E.; Oñate, J.J. Environmental effects of agri-environmental schemes in Western Europe. J. Environ. Manag. 2003, 67, 129–138. [Google Scholar] [CrossRef]
- Albert, L.; Franck, P.; Gilles, Y.; Plantegenest, M. Impact of agroecological infrastructures on the dynamics of Dysaphis plantaginea (hemiptera: Aphididae) and its natural enemies in apple orchards in northwestern France. Environ. Entomol. 2017, 46, 528–537. [Google Scholar] [CrossRef]
- van der Meer, M.; Kay, S.; Lüscher, G.; Jeanneret, P. What evidence exists on the impact of agricultural practices in fruit orchards on biodiversity? A systematic map. Environ. Evid. 2020, 9, 2. [Google Scholar] [CrossRef]
- Prieur, A.-G.A.; Swihart, R.K. Field attributes and farming practices associated with vole (Microtus) damage in cover-cropped fields. Agric. Ecosyst. Environ. 2020, 300, 106950. [Google Scholar] [CrossRef]
- Khlyap, L.A.; Warshavsky, A.A. Synanthropic and agrophilic rodents as invasive alien mammals. Russ. J. Biol. Invasions 2010, 1, 301–312. [Google Scholar] [CrossRef]
- Shiels, A.B.; Flores, C.A.; Khamsing, A.; Krushelnycky, P.D.; Mosher, S.M.; Drake, D.R. Dietary niche differentiation among three species of invasive rodents (Rattus rattus, R. exulans, Mus musculus). Biol. Invasions 2013, 15, 1037–1048. [Google Scholar] [CrossRef]
- Biró, Z.; Lanszki, J.; Szemethy, L.; Heltai, M.; Randi, E. Feeding habits of feral domestic cats (Felis catus), wild cats (Felis silvestris) and their hybrids: Trophic niche overlap among cat groups in Hungary. J. Zool. 2005, 266, 187–196. [Google Scholar] [CrossRef]
- Eccard, J.A.; Ylönen, H. Interspecific competition in small rodents: From populations to individuals. Evol. Ecol. 2003, 17, 423–440. [Google Scholar] [CrossRef]
- Heroldová, M.; Tkadlec, E.; Bryja, J.; Zejda, J. Wheat or barley? Appl. Anim. Behav. Sci. 2008, 110, 354–362. [Google Scholar] [CrossRef]
- Hansson, L. An Interpretation of Rodent Dynamics as Due to Trophic Interactions. Oikos 1987, 50, 308–318. [Google Scholar] [CrossRef]
- Myllymäki, A. Outbreaks and Damage by the Field Vole, Microtus agrestis (L.), since World War II in Europe. EPPO Bull. 1977, 7, 177–207. [Google Scholar] [CrossRef]
- Wheeler, P. The diet of field voles Microtus agrestis at low population density in upland Britain. Acta Theriol. 2005, 50, 483–492. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Trapped | Analysed | Adults | Sub-Adults | Juveniles | Males | Females |
|---|---|---|---|---|---|---|---|
| M. arvalis | 436 | 306 | 133 | 36 | 137 | 129 | 177 |
| M. agrestis | 31 | 31 | 21 | 6 | 4 | 13 | 18 |
| M. oeconomus | 42 | 39 * | 22 | 6 | 11 | 16 | 23 |
| Habitat | N | NONE | CV | FV | RV | CV + FV | CV + RV | FV + RV |
|---|---|---|---|---|---|---|---|---|
| Apple orchards (n = 10) | 46 | 21 | 15 | 1 | 2 | 2 | 4 | 1 |
| Plum orchards (n = 2) | 8 | 3 | 5 | |||||
| Raspberry plantations (n = 3) | 15 | 7 | 7 | 1 | ||||
| Currant plantations (n = 3) | 14 | 5 | 9 | |||||
| High blueberry plantations (n = 1) | 3 | 3 | ||||||
| Control meadows (n = 16) | 82 | 39 | 26 | 4 | 5 | 1 | 6 | 1 |
| Species | δ13C Values, ‰ | δ15N Values, ‰ | ||||
|---|---|---|---|---|---|---|
| Mean ± SE | Min–Max | Range | Mean ± SE | Min–Max | Range | |
| M. arvalis | −27.08 ± 0.04 | −28.58–−24.58 | 4.00 | 4.93 ± 0.08 | 0.52–14.50 | 13.98 |
| M. agrestis | −27.54 ± 0.10 | −29.00–−26.48 | 2.52 | 5.16 ± 0.33 | 1.91–9.13 | 7.22 |
| M. oeconomus | −27.59 ± 0.11 | −28.88–−26.37 | 2.51 | 5.77 ± 0.25 | 2.16–9.01 | 6.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balčiauskas, L.; Skipitytė, R.; Garbaras, A.; Stirkė, V.; Balčiauskienė, L.; Remeikis, V. Stable Isotopes Reveal the Dominant Species to Have the Widest Trophic Niche of Three Syntopic Microtus Voles. Animals 2021, 11, 1814. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11061814
Balčiauskas L, Skipitytė R, Garbaras A, Stirkė V, Balčiauskienė L, Remeikis V. Stable Isotopes Reveal the Dominant Species to Have the Widest Trophic Niche of Three Syntopic Microtus Voles. Animals. 2021; 11(6):1814. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11061814
Chicago/Turabian StyleBalčiauskas, Linas, Raminta Skipitytė, Andrius Garbaras, Vitalijus Stirkė, Laima Balčiauskienė, and Vidmantas Remeikis. 2021. "Stable Isotopes Reveal the Dominant Species to Have the Widest Trophic Niche of Three Syntopic Microtus Voles" Animals 11, no. 6: 1814. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11061814