
Taxonomic Reassessment of the Izumo Lineage of Hynobius utsunomiyaorum: Description of a New Species from Chugoku, Japan

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Molecular Analysis
2.2. Morphological Analysis
2.3. Sampling and Measurements of Type Specimens
3. Results
3.1. Molecular Analysis
3.2. Morphological Analysis
4. Species Account
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frost, D.R. Amphibian Species of the World: An Online Reference; American Museum of Natural History: New York, NY, USA, 2021; Available online: http://research.amnh.org/herpetology/amphibia/index.html (accessed on 15 February 2021).
- Matsui, M.; Nishikawa, K.; Misawa, Y.; Kakegawa, M.; Sugahara, T. Taxonomic relationships of an endangered Japanese salamander Hynobius hidamontanus Matsui, 1987 with H. tenuis Nambu, 1991 (Amphibia: Caudata). Curr. Herpetol. 2002, 21, 25–34. [Google Scholar] [CrossRef]
- Matsui, M.; Okawa, H.; Nishikawa, K.; Aoki, G.; Eto, K.; Yoshikawa, N.; Tanabe, S.; Misawa, Y.; Tominaga, A. Systematics of the widely distributed Japanese clouded salamander, Hynobius nebulosus (Amphibia: Caudata: Hynobiidae), and Its Closest Relatives. Curr. Herpetol. 2019, 38, 32–90. [Google Scholar] [CrossRef]
- Tominaga, A.; Matsui, M.; Nishikawa, K. Two new species of lotic breeding Salamanders (Amphibia, Caudata, Hynobiidae) from Western Japan. Zootaxa 2019, 4550, 525–544. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.; Ooi, K. Molecular Study on Hynobius in East Area of Shimane Prefecture, Japan: Notes on Haplotype Diversity of mtDNA and Their Distribution. Bull. Hoshizaki Green Found. 2020, 23, 99–104. (In Japanese) [Google Scholar]
- Matsui, M.; Nishikawa, K.; Utsunomiya, T.; Tanabe, S. Geographic allozyme variation in the Japanese clouded Salamander, Hynobius nebulosus (Amphibia: Urodela). Biol. J. Linn. Soc. 2006, 89, 311–330. [Google Scholar] [CrossRef] [Green Version]
- Dubois, A. The International code of zoological nomenclature must be drastically improved before it is too late. Bionomina 2011, 2, 1–104. [Google Scholar] [CrossRef] [Green Version]
- Frost, D.R.; Hillis, D.M. Species in concept and practice: Herpetological applications. Herpetologica 1990, 46, 87–104. [Google Scholar]
- Matsui, M.; Tominaga, A.; Hayashi, T.; Misawa, Y.; Tanabe, S. Phylogenetic Relationship and Phylogeography of Hynobius tokyoensis (Amphibia: Caudata) using complete sequences of cytochrome b and control reason genes of mitochondrial DNA. Mol. Phylogenet. Evol. 2007, 44, 204–216. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiura, N. Further analysts of the data by akaike’s information criterion and the finite corrections. Commun. Stat. Theor. Methods 1978, 7, 13–26. [Google Scholar] [CrossRef]
- Schwarz, G. Estimating the dimension of a model. Ann. Statist. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsenbeck, J.P.; Rannala, B. Frequentist properties of Bayesian posterior probabilities of phylogenetic trees under simple and complex substitution models. Syst. Biol. 2004, 53, 904–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regier, J.C.; Mitter, C.; Solis, M.A.; Hayden, J.E.; Landry, B.; Nuss, M.; Simonsen, T.J.; Yen, S.H.; Zwick, A.; Cummings, M.P. A molecular phylogeny for the pyraloid moths (Lepidoptera: Pyraloidea) and its implications for higher-level classification. Syst. Entomol. 2012, 37, 635–656. [Google Scholar] [CrossRef]
- Bennett, R.A. A Review of anesthesia and chemical restraint in reptiles. J. Zoo Wildl. Med. 1991, 22, 282–303. [Google Scholar]
- Ihaka, R.; Gentleman, R. R: A language for data analysis and graphics. J. Comp. Graph. Stat. 1996, 5, 299–314. [Google Scholar]
- Dubois, A.; Raffaëlli, J. A new ergotaxonomy of the order Urodela Duméril, 1805 (Amphibia, Batrachia). Alytes 2012, 28, 77–161. [Google Scholar]
- Fujita, H.; Iwata, T.; Teraoka, S. Damage example of eggs of Hynobius nebulosus (high ground type) by overfishing in Eastern Shimane Prefecture. Bull. Hoshizaki Green Found. 2016, 19, 253–255. (In Japanese) [Google Scholar]










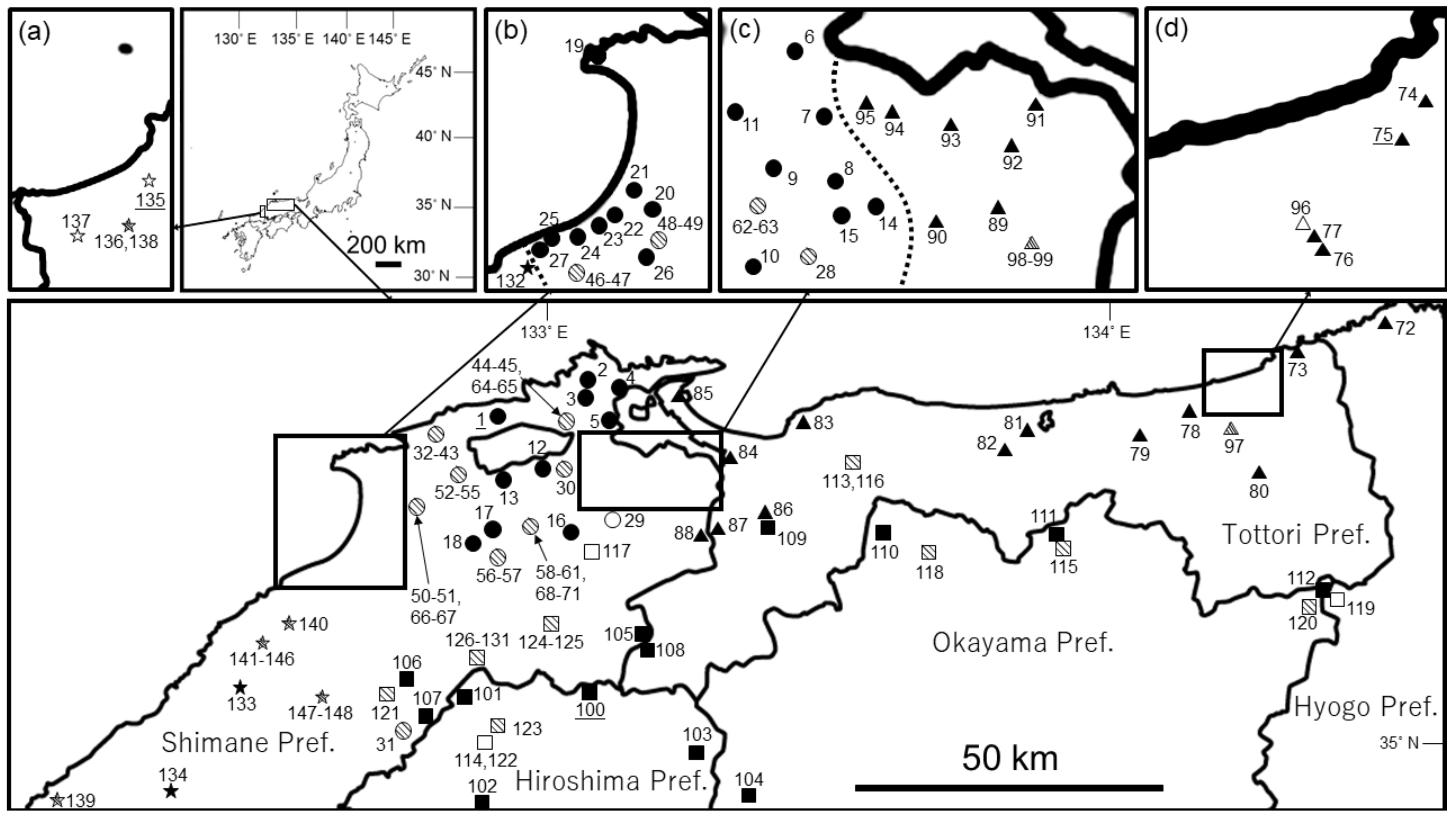{kind=link}
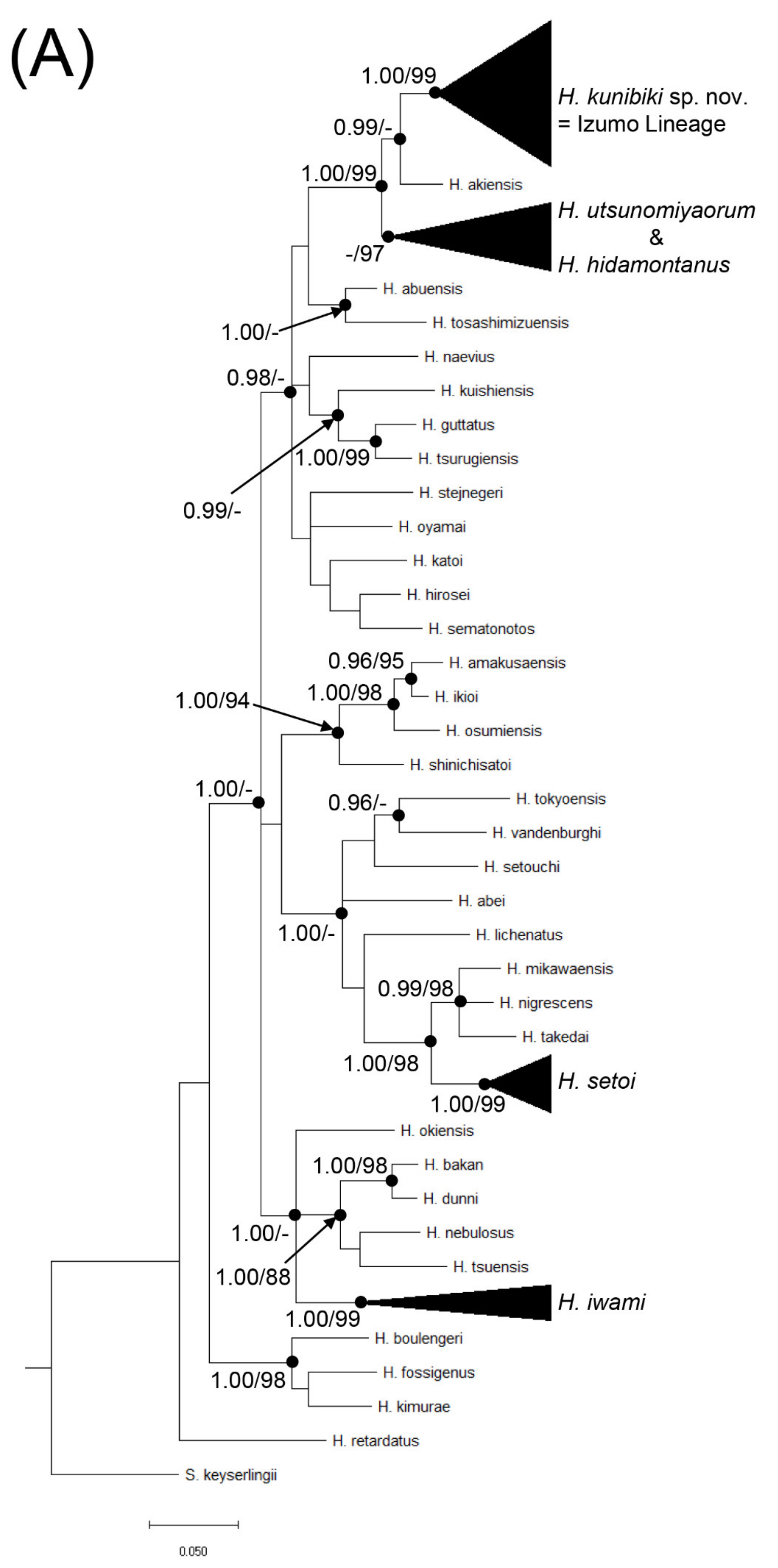{kind=link}
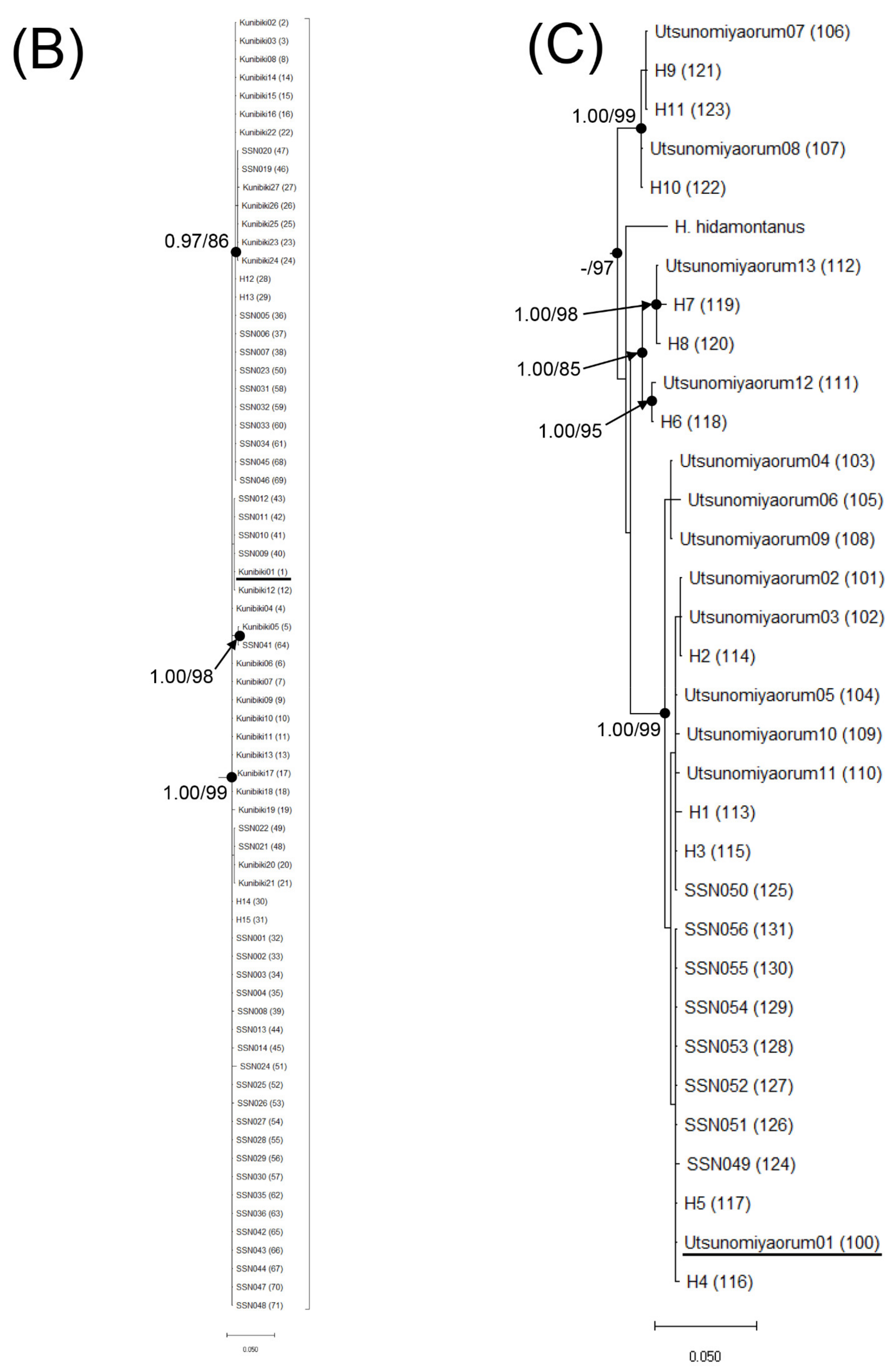{kind=link}
{kind=link}
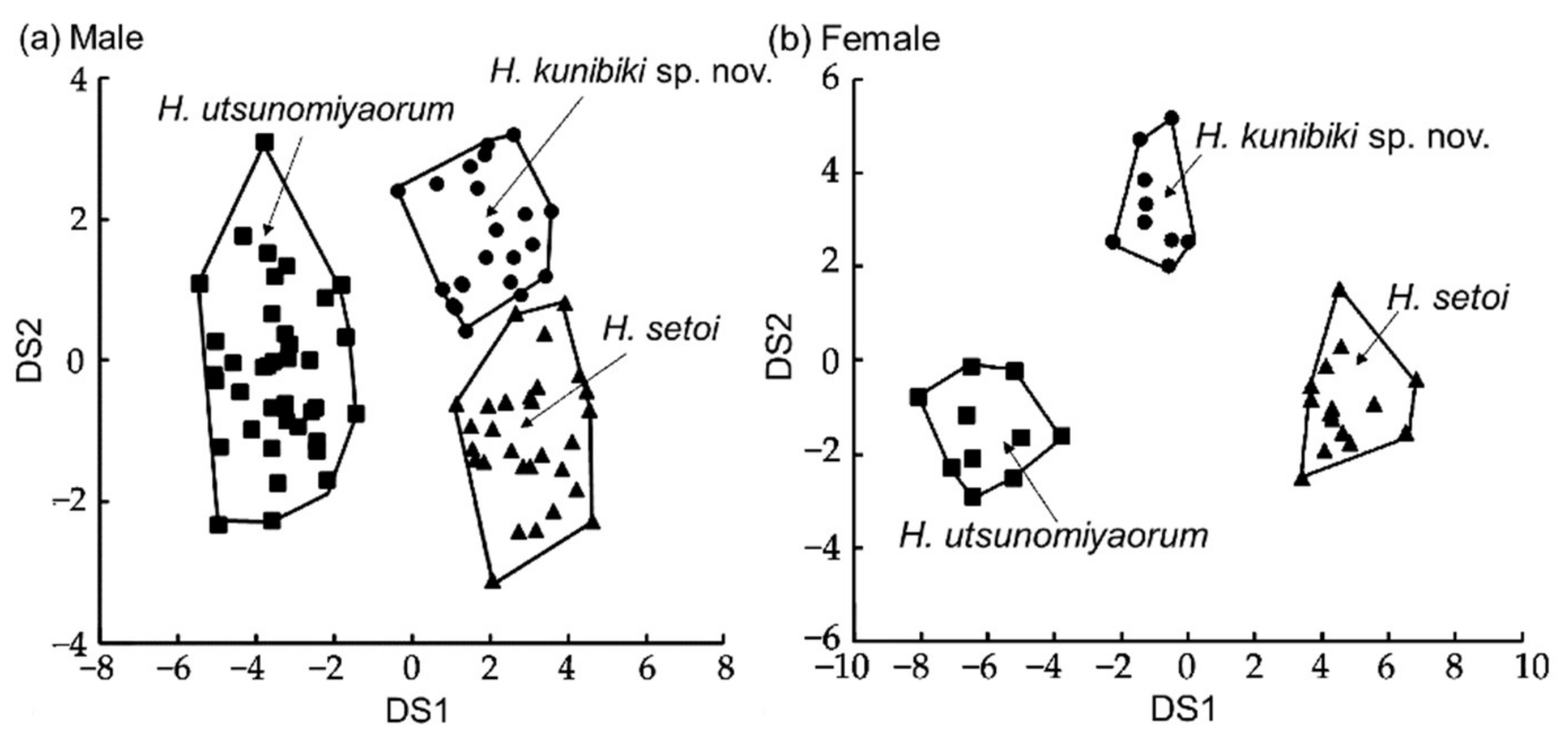{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H. kunibiki sp. nov. | H. setoi | H. utsunomiyaorum | ||||
|---|---|---|---|---|---|---|
| Male | Female | Male | Female | Male | Female | |
| Trait | n = 22 | n = 9 | n = 30 | n = 15 | n = 38 | n = 10 |
| SVL | 58.9 ± 3.15 | 58.2 ± 4.24 | 60.7 ± 6.37 | 63.0 ± 6.16 | 53.4 ± 4.10 | 54.6 ± 4.19 |
| (52.7–63.9) | (51.0–66.6) | (49.0–70.8) | (50.5–72.5) | (40.0–59.8) | (48.0–60.5) | |
| RTRL | 76.5 ± 1.00 | 77.6 ± 1.24 | 77.4 ± 1.00 | 77.7 ± 2.20 | 76.7 ± 1.65 | 77.0 ± 0.62 |
| (75.7–78.4) | (75.6–79.2) | (75.7–80.1) | (71.0–80.8) | (74.3–84.8) | (76.2–77.8) | |
| RAGD | 51.3 ± 2.06 | 55.7 ± 2.35 | 52.3 ± 1.72 | 55.5 ± 2.63 | 52.1 ± 1.68 | 52.8 ± 2.27 |
| (47.7–54.9) | (50.8–59.2) | (48.1–55.9) | (50.7–59.1) | (48.9–56.5) | (49.0–55.9) | |
| RHL | 24.2 ± 0.84 | 23.3 ± 0.91 | 23.5 ± 0.99 | 23.0 ± 1.10 | 24.4 ± 1.03 | 24.2 ± 0.73 |
| (22.6–25.7) | (21.8–24.6) | (22.0–26.1) | (21.2–24.8) | (22.0–26.5) | (22.6–25.0) | |
| RTAL | 82.0 ± 4.09 | 71.3 ± 4.28 | 78.3 ± 6.15 | 69.5 ± 5.47 | 73.1 ± 7.28 | 70.5 ± 6.51 |
| (74.1–93.1) | (62.8–76.4) | (67.5–94.7) | (59.7–77.0) | (52.9–86.0) | (58.8–81.4) | |
| RMTAW | 6.9 ± 0.84 | 5.8 ± 0.76 | 6.6 ± 1.01 | 6.5 ± 0.87 | 6.7 ± 0.96 | 6.6 ± 0.59 |
| (5.5–8.3) | (4.9–7.4) | (4.8–8.8) | (4.8–7.7) | (4.4–8.8) | (5.7–7.5) | |
| RMTAH | 13.0 ± 0.95 | 10.8 ± 1.38 | 13.0 ± 1.36 | 11.3 ± 1.17 | 10.2 ± 1.37 | 9.3 ± 0.75 |
| (11.4–14.6) | (7.5–12.2) | (10.8–15.9) | (9.1–13.7) | (6.8–12.9) | (8.3–10.6) | |
| RVTL | 5.1 ± 0.46 | 5.3 ± 0.50 | 4.9 ± 0.48 | 4.6 ± 0.63 | 4.5 ± 0.50 | 4.7 ± 0.38 |
| (4.3–6.1) | (4.3–5.9) | (3.9–6.1) | (3.5–5.6) | (3.6–5.9) | (4.1–5.4) | |
| RVTW | 5.4 ± 0.51 | 5.4 ± 0.48 | 5.4 ± 0.47 | 5.1 ± 0.61 | 5.7 ± 0.50 | 5.6 ± 0.28 |
| (4.4–6.3) | (4.7–6.0) | (4.4–6.1) | (4.3–6.4) | (4.7–6.7) | (5.0–6.0) | |
| RHW | 17.7 ± 0.62 | 17.0 ± 0.87 | 17.5 ± 0.87 | 16.6 ± 0.77 | 17.3 ± 0.73 | 16.7 ± 0.71 |
| (16.6–19.2) | (15.8–18.2) | (16.2–19.0) | (15.3–17.6) | (15.7–18.8) | (15.5–18.2) | |
| RFLL | 25.5 ± 1.49 | 24.0 ± 1.45 | 24.8 ± 1.71 | 22.3 ± 0.92 | 24.8 ± 1.69 | 25.0 ± 2.50 |
| (23.1–29.5) | (21.7–26.1) | (22.6–30.6) | (20.7–24.2) | (21.0–27.9) | (21.1–28.8) | |
| RHLL | 32.4 ± 1.60 | 31.0 ± 1.87 | 32.1 ± 1.31 | 29.4 ± 1.75 | 31.8 ± 1.30 | 31.8 ± 1.50 |
| (29.9–35.2) | (27.7–34.8) | (29.5–36.3) | (25.9–31.9) | (29.3–35.6) | (29.3–34.2) | |
| R2FL | 5.2 ± 0.62 | 4.7 ± 0.69 | 4.9 ± 0.71 | 4.3 ± 0.68 | 5.1 ± 0.71 | 4.9 ± 0.65 |
| (3.9–6.3) | (4.1–6.0) | (2.9–6.5) | (3.1–5.5) | (2.9–6.6) | (4.0–5.9) | |
| R3FL | 4.1 ± 0.54 | 3.7 ± 0.87 | 4.4 ± 0.67 | 4.0 ± 0.58 | 3.6 ± 0.67 | 3.6 ± 0.58 |
| (2.5–4.9) | (2.3–5.0) | (2.8–5.5) | (3.0–5.1) | (1.4–4.9) | (2.8–4.7) | |
| R3TL | 7.9 ± 0.64 | 7.5 ± 0.61 | 8.0 ± 0.56 | 7.3 ± 0.84 | 8.3 ± 0.82 | 8.4 ± 0.59 |
| (7.0–9.1) | (6.4–8.3) | (7.0–9.1) | (5.9–9.1) | (5.5–10.4) | (7.4–9.4) | |
| R5TL | 1.9 ± 0.62 | 1.8 ± 0.69 | 2.2 ± 0.62 | 2.3 ± 0.69 | 0.2 ± 0.48 | 0.3 ± 0.48 |
| (0.3–3.0) | (0.5–2.6) | (1.1–3.3) | (1.1–3.5) | (0.0–2.2) | (0.0–1.2) | |
| RIND | 5.3 ± 0.57 | 4.9 ± 0.52 | 4.7 ± 0.48 | 4.6 ± 0.29 | 5.2 ± 0.45 | 4.9 ± 0.67 |
| (3.9–6.3) | (4.2–6.1) | (3.7–5.7) | (4.1–5.2) | (4.4–6.1) | (4.3–6.3) | |
| RIOD | 6.4 ± 0.51 | 6.0 ± 0.37 | 5.9 ± 0.63 | 5.6 ± 0.38 | 6.3 ± 0.38 | 6.1 ± 0.46 |
| (5.3–7.5) | (5.6–6.6) | (4.9–7.1) | (5.0–6.3) | (5.4–6.9) | (5.5–7.0) | |
| RUEL | 3.2 ± 0.25 | 3.2 ± 0.19 | 3.2 ± 0.30 | 3.2 ± 0.28 | 3.3 ± 0.28 | 3.3 ± 0.27 |
| (2.8–3.8) | (2.9–3.4) | (2.5–3.9) | (2.8–3.8) | (2.8–3.9) | (2.9–3.8) | |
| RSL | 6.7 ± 0.32 | 6.4 ± 0.35 | 6.0 ± 0.34 | 5.7 ± 0.29 | 6.4 ± 0.38 | 6.1 ± 0.53 |
| (6.1–7.3) | (5.7–6.8) | (5.2–6.7) | (5.3–6.3) | (5.4–7.4) | (5.0–6.6) | |
| RUEW | 4.6 ± 0.20 | 4.3 ± 0.29 | 4.4 ± 0.28 | 4.2 ± 0.29 | 4.6 ± 0.37 | 4.4 ± 0.25 |
| (4.2–5.1) | (3.9–4.7) | (3.8–4.9) | (3.7–5.0) | (4.1–5.5) | (4.1–4.9) | |
| RLJL | 13.9 ± 0.69 | 13.7 ± 0.69 | 13.4 ± 0.75 | 13.1 ± 0.85 | 14.1 ± 0.73 | 13.4 ± 0.61 |
| (12.9–15.7) | (13.1–15.4) | (11.5–14.9) | (11.5–14.6) | (12.7–16.4) | (12.2–14.1) | |
| H. kunibiki sp. nov. | H. setoi | H. utsunomiyaorum | |||||
|---|---|---|---|---|---|---|---|
| Male | Female | Male | Female | Male | Female | ||
| Character | Condition | n = 22 | n = 9 | n = 30 | n = 15 | n = 38 | n = 10 |
| DWSV | Absent | 21 (95.5%) | 5 (55.6%) | 21 (70.0%) | 7 (46.7%) | 8 (21.1%) | 1 (10.0%) |
| Present | 1 (4.5%) | 4 (44.4%) | 9 (30.0%) | 8 (53.3%) | 30 (78.9%) | 9 (90.0%) | |
| DWSL | Absent | 21 (95.5%) | 9 (100%) | 28 (93.3%) | 13 (86.7%) | 4 (10.5%) | 1 (10.0%) |
| Present | 1 (4.5%) | 0 (0%) | 2 (6.7%) | 2 (13.3%) | 34 (89.5%) | 9 (90.0%) | |
| DCTYLD | Absent | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 33 (86.8%) | 6 (60.0%) |
| Present | 22 (100%) | 9 (100%) | 30 (100%) | 15 (100%) | 5 (13.2%) | 4 (40.0%) | |
| DTYLV | Absent | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 38 (100%) | 10 (100%) |
| Present | 22 (100%) | 9 (100%) | 30 (100%) | 15 (100%) | 0 (0%) | 0 (0%) | |
| DGM | Absent | 4 (18.2%) | 9 (100%) | 14 (46.7%) | 15 (100%) | 29 (76.3%) | 10 (100%) |
| Present | 18 (81.8%) | 0 (0%) | 16 (53.3%) | 0 (0%) | 9 (23.7%) | 0 (0%) | |
| CGN | 11 | 1 (4.5%) | 1 (11.1%) | 2 (6.7%) | 0 (0%) | 7 (18.4%) | 1 (10.0%) |
| 12 | 18 (81.8%) | 4 (44.4%) | 24 (80.0%) | 7 (46.7%) | 23 (60.5%) | 8 (80.0%) | |
| 13 | 3 (13.6%) | 4 (44.4%) | 4 (13.3%) | 8 (53.3%) | 8 (21.1%) | 1 (10.0%) | |
| CFBALN | 2.0 | 0 (0%) | 0 (0%) | 1 (3.3%) | 0 (0%) | 0 (0%) | 0 (0%) |
| 1.5 | 1 (4.5%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | |
| 1.0 | 5 (22.7%) | 0 (0%) | 2 (6.7%) | 0 (0%) | 3 (7.9%) | 0 (0%) | |
| 0.5 | 4 (18.2%) | 2 (22.2%) | 2 (6.7%) | 0 (0%) | 7 (18.4%) | 1 (10.0%) | |
| 0.0 | 6 (27.3%) | 0 (0%) | 8 (26.7%) | 0 (0%) | 9 (23.7%) | 3 (30.0%) | |
| −0.5 | 4 (18.2%) | 0 (0%) | 5 (16.7%) | 0 (0%) | 9 (23.7%) | 1 (10.0%) | |
| −1.0 | 2 (9.1%) | 3 (33.3%) | 5 (16.7%) | 0 (0%) | 8 (21.1%) | 3 (30.0%) | |
| −1.5 | 0 (0%) | 0 (0%) | 5 (16.7%) | 2 (13.3%) | 1 (2.6%) | 1 (10.0%) | |
| −2.0 | 0 (0%) | 3 (33.3%) | 2 (6.7%) | 8 (53.3%) | 1 (2.6%) | 1 (10.0%) | |
| −2.5 | 0 (0%) | 0 (0%) | 0 (0%) | 2 (13.3%) | 0 (0%) | 0 (0%) | |
| −3.0 | 0 (0%) | 1 (11.1%) | 0 (0%) | 3 (20.0%) | 0 (0%) | 0 (0%) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sugawara, H.; Iwata, T.; Yamashita, H.; Nagano, M. Taxonomic Reassessment of the Izumo Lineage of Hynobius utsunomiyaorum: Description of a New Species from Chugoku, Japan. Animals 2021, 11, 2187. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11082187
Sugawara H, Iwata T, Yamashita H, Nagano M. Taxonomic Reassessment of the Izumo Lineage of Hynobius utsunomiyaorum: Description of a New Species from Chugoku, Japan. Animals. 2021; 11(8):2187. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11082187
Chicago/Turabian StyleSugawara, Hirotaka, Takayuki Iwata, Hitoshi Yamashita, and Masahiro Nagano. 2021. "Taxonomic Reassessment of the Izumo Lineage of Hynobius utsunomiyaorum: Description of a New Species from Chugoku, Japan" Animals 11, no. 8: 2187. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11082187