
Heat Stress Induces Shifts in the Rumen Bacteria and Metabolome of Buffalo


 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Method
2.1. Animal Management
2.2. Sample Collection and Measurement
2.3. DNA Extraction, 16S rDNA Sequencing and Sequences Analysis
2.4. Metabolomics Profiling for Rumen Fluid and Data Analysis
2.5. Statistical Analysis
3. Results
3.1. Physiological Parameters, Animal Performance and Rumen Fermentation
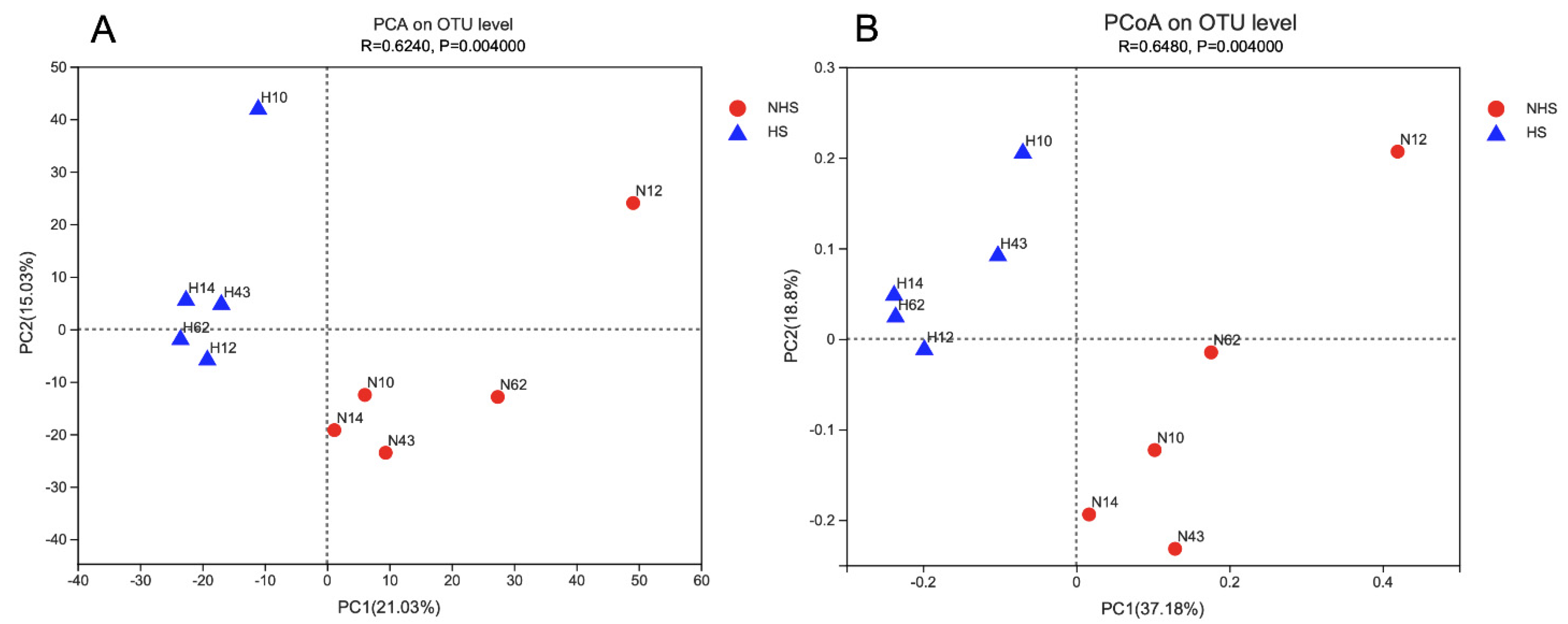
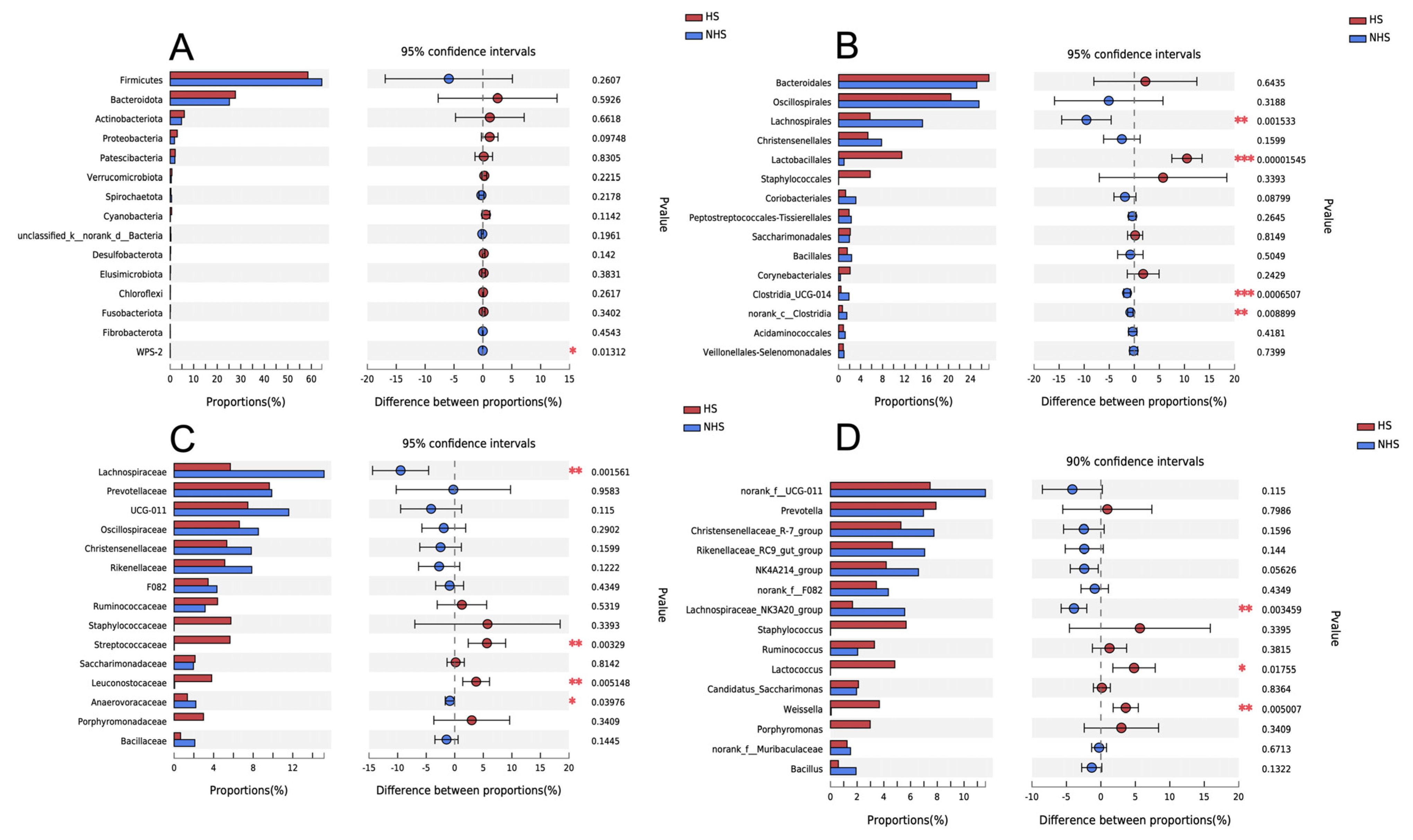
3.2. Rumen Bacteria Diversity and Composition
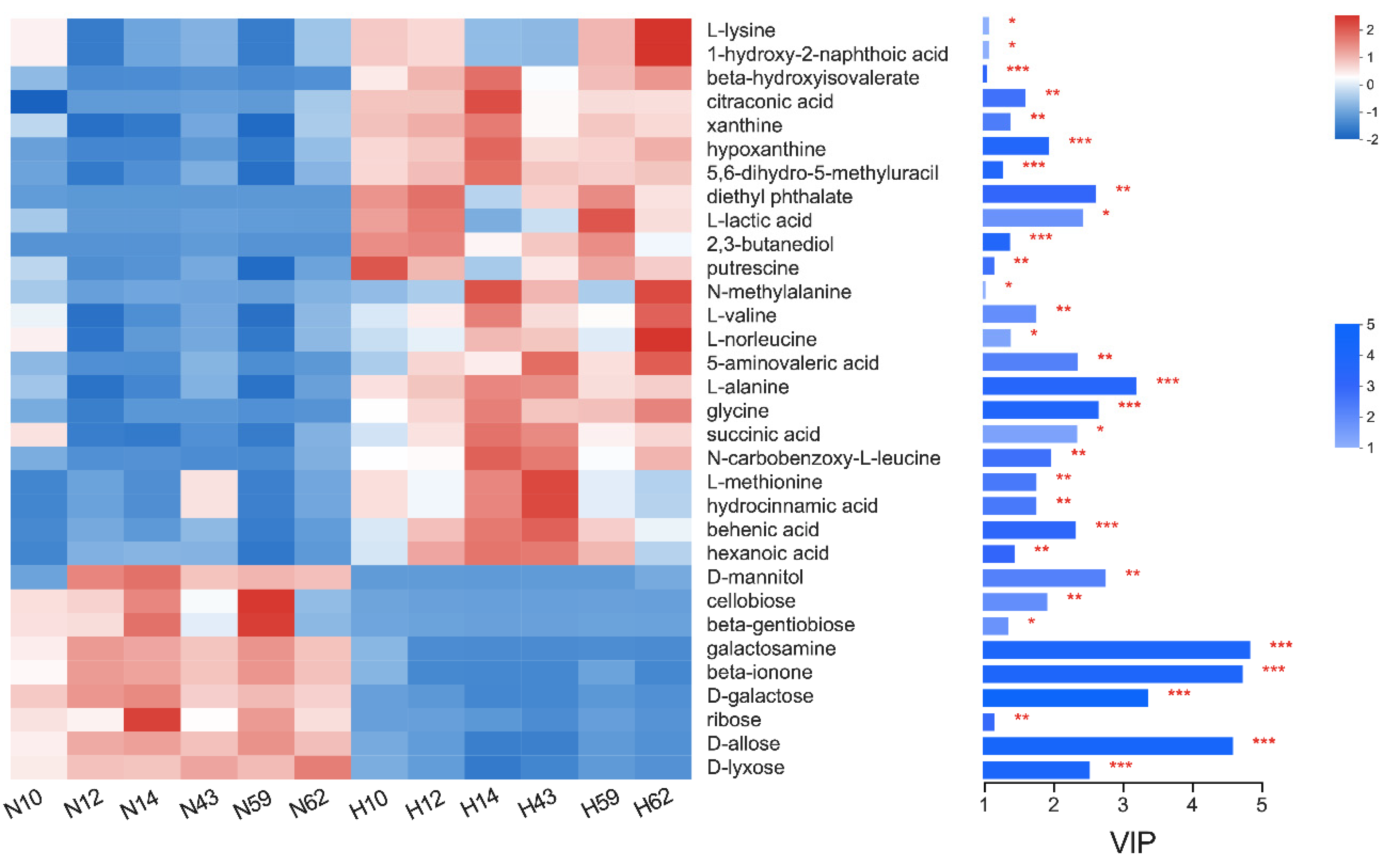
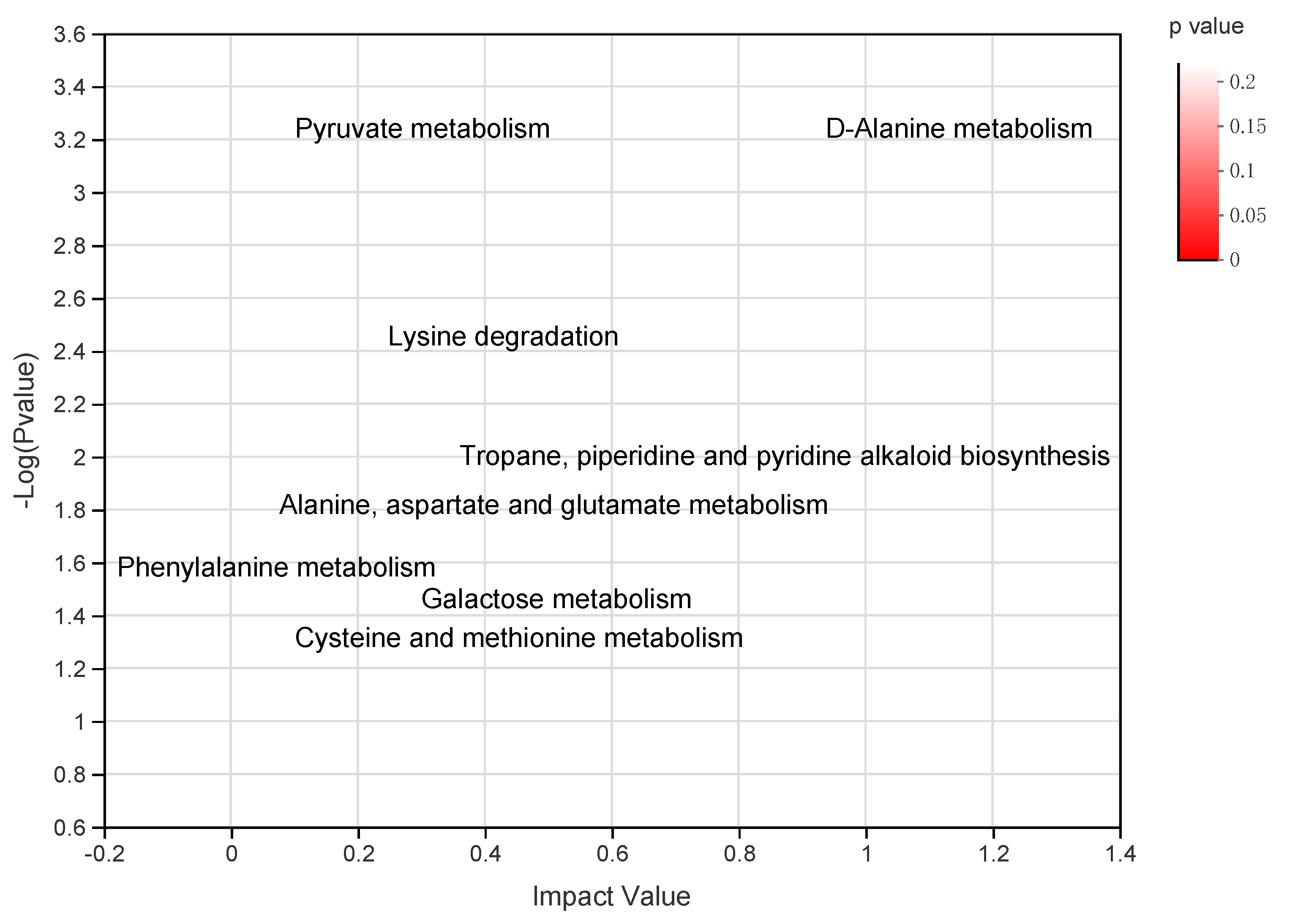
3.3. Identification of Different Metabolites and Metabolic Pathways
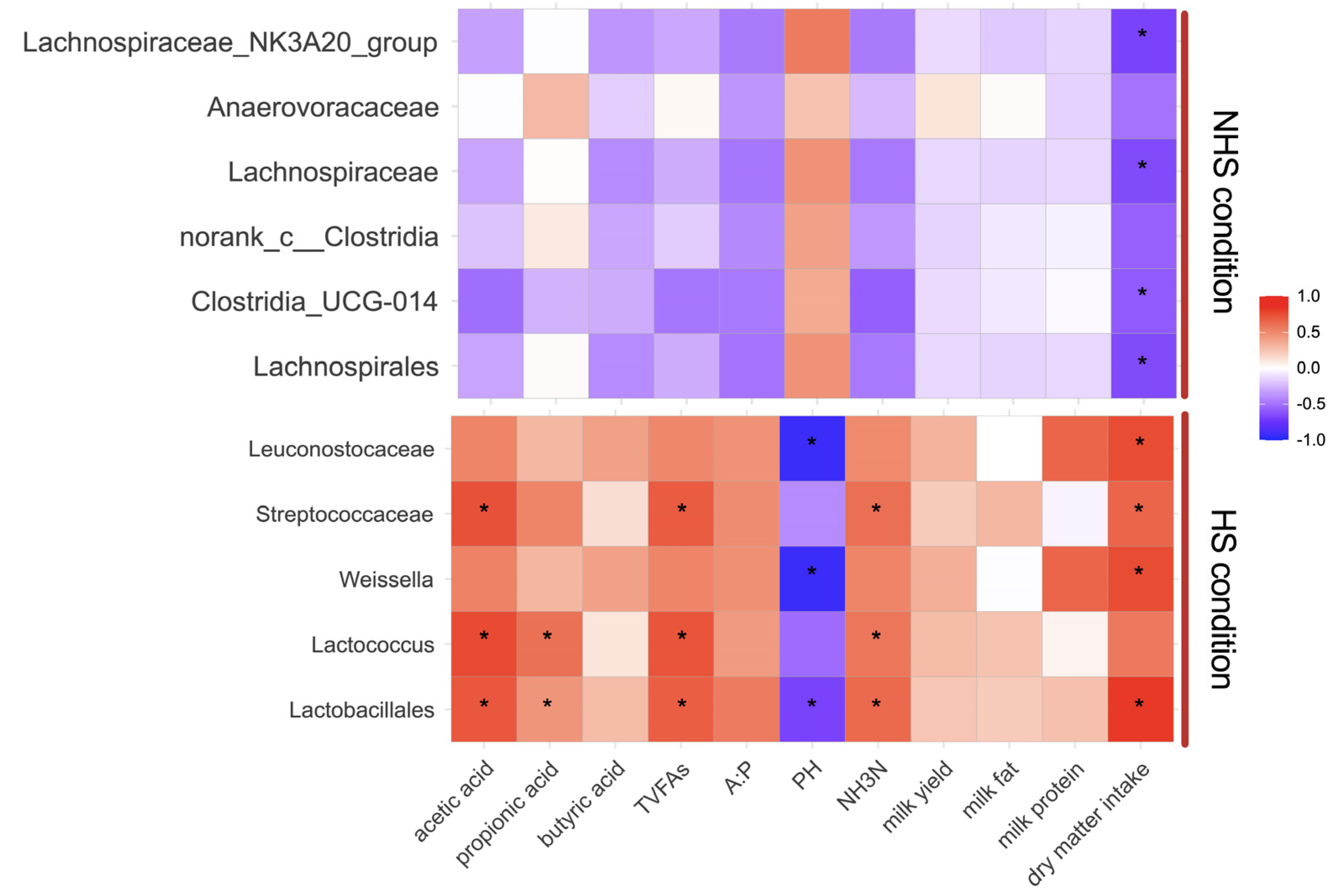
3.4. Correlation between Rumen Bacteria, Metabolites and Fermentation Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seerapu, S.R.; Kancharana, A.R.; Chappidi, V.S.; Bandi, E.R. Effect of microclimate alteration on milk production and composition in Murrah buffaloes. Vet. World 2015, 8, 1444–1452. [Google Scholar] [CrossRef] [Green Version]
- Wu, J. The response of the distributions of Asian buffalo breeds in China to climate change over the past 50 years. Livest. Sci. 2015, 180, 65–77. [Google Scholar] [CrossRef]
- Dayal, S.; Dey, A.; Pandian, S.J.; Gupta, J.J.; Chandran, P.C.; Ali, I. Effect of seasonal variation on physiological parameters in Murrah buffaloes. Indian J. Anim. Sci. 2017, 87, 965–967. [Google Scholar]
- Liu, S.; Li, J.; Li, Z.; Deng, T.; Rehman, Z.U.; Zichao, Z.; Liguo, Y. Effect of season and breed on physiological and blood parameters in buffaloes. J. Dairy Res. 2018, 85, 181–184. [Google Scholar]
- Bombade, K.; Kamboj, A.; Alhussien, M.N.; Mohanty, A.K.; Dang, A.K. Diurnal variation of milk somatic and differential leukocyte counts of Murrah buffaloes as influenced by different milk fractions, seasons and parities. Biol. Rhythm. Res. 2018, 49, 151–163. [Google Scholar] [CrossRef]
- Gu, Z.B.; Li, L.; Tang, S.K.; Liu, C.B.; Fu, X.H.; Shi, Z.; Mao, H. Metabolomics Reveals that Crossbred Dairy Buffaloes Are More Thermotolerant than Holstein Cows under Chronic Heat Stress. J. Agric. Food Chem. 2018, 66, 12889–12897. [Google Scholar] [CrossRef]
- Kumar, A.; Mehrotra, S.; Singh, G.; Narayanan, K.; Das, G.K.; Soni, Y.K.; Singh, M.; Mahla, A.S.; Srivastava, N.; Verma, M.R. Sustained delivery of exogenous melatonin influences biomarkers of oxidative stress and total antioxidant capacity in summer-stressed anestrous water buffalo (Bubalus bubalis). Theriogenology 2015, 83, 1402–1407. [Google Scholar] [CrossRef]
- Amamou, H.; Beckers, Y.; Mahouachi, M.; Hammami, H. Thermotolerance indicators related to production and physiological responses to heat stress of holstein cows. J. Therm. Biol. 2019, 82, 90–98. [Google Scholar] [CrossRef]
- Wu, X.H.; Sun, H.Z.; Xue, M.Y.; Wang, D.M.; Guan, L.L.; Liu, J. Serum metabolome profiling revealed potential biomarkers for milk protein yield in dairy cows. J. Proteom. 2018, 184, 54–61. [Google Scholar] [CrossRef]
- Sun, H.-Z.; Wang, D.-M.; Wang, B.; Wang, J.-K.; Liu, H.-Y.; Guan, L.L.; Liu, J.-X. Metabolomics of Four Biofluids from Dairy Cows: Potential Biomarkers for Milk Production and Quality. J. Proteome Res. 2015, 14, 1287–1298. [Google Scholar] [CrossRef]
- Zhao, S.; Min, L.; Zheng, N.; Wang, J. Effect of Heat Stress on Bacterial Composition and Metabolism in the Rumen of Lactating Dairy Cows. Animals 2019, 9, 925. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Ye, G.; Zhou, Y.; Liu, Y.; Zhao, L.; Liu, Y.; Chen, X.; Huang, D.; Liao, S.F.; Huang, K. Feeding glycerol-enriched yeast culture improves performance, energy status, and heat shock protein gene expression of lactating Holstein cows under heat stress. J. Anim. Sci. 2014, 92, 2494–2502. [Google Scholar] [CrossRef]
- Kim, D.-H.; Kim, M.-H.; Kim, S.-B.; Son, J.-K.; Lee, J.-H.; Joo, S.-S.; Gu, B.-H.; Park, T.; Park, B.-Y.; Kim, E.-T. Differential Dynamics of the Ruminal Microbiome of Jersey Cows in a Heat Stress Environment. Animals 2020, 10, 1127. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International, 17th ed.; AOAC: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Liang, Y.S.; Li, G.Z.; Li, X.Y.; Lü, J.Y.; Li, F.D.; Tang, D.F.; Li, F.; Deng, Y.; Zhang, H.; Wang, Z.L.; et al. Growth performance, rumen fermentation, bacteria composition, and gene expressions involved in intracellular pH regulation of rumen epithelium in finishing Hu lambs differing in residual feed intake phenotype. J. Anim. Sci. 2017, 95, 1727–1738. [Google Scholar]
- Shen, J.S.; Chai, Z.; Song, L.J.; Liu, J.X.; Wu, Y.M. Insertion depth of oral stomach tubes may affect the fermentation parameters of ruminal fluid collected in dairy cows. J. Dairy Sci. 2012, 95, 5978–5984. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Long, S.; Wang, Q.; Zhang, L.; Hu, J.; Yang, J.; Cheng, Z.; Piao, X. Mixed organic acids improve nutrients digestibility, volatile fatty acids composition and intestinal microbiota in growing-finishing pigs fed high-fiber diet. Asian-Australas. J. Anim. Sci. 2019, 32, 856–864. [Google Scholar] [CrossRef]
- Seankamsorn, A.; Cherdthong, A.; So, S.; Wanapat, M. Influence of chitosan sources on intake, digestibility, rumen fermentation, and milk production in tropical lactating dairy cows. Trop. Anim. Health Prod. 2021, 53, 241. [Google Scholar] [CrossRef]
- Liu, H.; Hu, L.; Han, X.; Zhao, N.; Xu, T.; Ma, L.; Wang, X.; Zhang, X.; Kang, S.; Zhao, X.; et al. Tibetan Sheep Adapt to Plant Phenology in Alpine Meadows by Changing Rumen Microbial Community Structure and Function. Front. Microbiol. 2020, 11, 2547. [Google Scholar] [CrossRef]
- Li, Y.Q.Q.; Xi, Y.M.M.; Wang, Z.D.D.; Zeng, H.F.F.; Han, Z.Y. Combined signature of rumen microbiome and metabolome in dairy cows with different feed intake levels. J. Anim. Sci. 2020, 98, skaa070. [Google Scholar] [CrossRef]
- Zhang, R.Y.; Liu, Y.J.; Yin, Y.Y.; Jin, W.; Mao, S.Y.; Liu, J.H. Response of rumen microbiota, and metabolic profiles of rumen fluid, liver and serum of goats to high-grain diets. Animal 2019, 13, 1855–1864. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Yuxia, C.; Zichen, W.; Zipeng, L.; Ahmad, M.J.; Ming, L.; Tengyun, G.; Shenhe, L. The Effect of Exogenous Melatonin on Milk Somatic Cell Count in Buffalo. Pak. Vet. J. 2021, 41, 152–155. [Google Scholar] [CrossRef]
- Hammami, H.; Bormann, J.; M’hamdi, N.; Montaldo, H.H.; Gengler, N. Evaluation of heat stress effects on production traits and somatic cell score of Holsteins in a temperate environment. J. Dairy Sci. 2013, 96, 1844–1855. [Google Scholar] [CrossRef] [Green Version]
- Lina, X.; Chaoyuan, W.; Luyu, D.; Zhiyuan, G.; Lu, Z.; Zhengxiang, S.; Baoming, L.; Chuntao, J. Heat stress alleviation for dairy cows housed in an open-sided barn by cooling fan and perforated air ducting (PAD) system. Int. J. Agric. Biol. Eng. 2017, 10, 1–10. [Google Scholar] [CrossRef]
- Becker, C.A.; Collier, R.J.; Stone, A.E. Invited review: Physiological and behavioral effects of heat stress in dairy cows. J. Dairy Sci. 2020, 103, 6751–6770. [Google Scholar] [CrossRef]
- Wheelock, J.B.; Rhoads, R.P.; VanBaale, M.J.; Sanders, S.R.; Baumgard, L.H. Effects of heat stress on energetic metabolism in lactating Holstein cows. J. Dairy Sci. 2010, 93, 644–655. [Google Scholar] [CrossRef]
- Gao, S.T.; Guo, J.; Quan, S.Y.; Nan, X.M.; Fernandez, M.S.; Baumgard, L.H.; Bu, D.P. The effects of heat stress on protein metabolism in lactating Holstein cows. J. Dairy Sci. 2017, 100, 5040–5049. [Google Scholar] [CrossRef] [Green Version]
- Khongdee, T.; Sripoon, S.; Vajrabukka, C. The effects of high temperature and roof modification on physiological responses of swamp buffalo (Bubalus bubalis) in the tropics. Int. J. Biometeorol. 2013, 57, 349–354. [Google Scholar] [CrossRef]
- Johnson, J.S.; Scharf, B.; Weaber, R.L.; Eichen, P.A.; Spiers, D.E. Patterns of heat response and adaptation on summer pasture: A comparison of heat-sensitive (Angus) and -tolerant (Romosinuano) cattle. J. Therm. Biol. 2012, 37, 344–350. [Google Scholar] [CrossRef]
- Wang, X.; Li, X.; Zhao, C.; Hu, P.; Chen, H.; Liu, Z.; Liu, G.; Wang, Z. Correlation between Composition of the Bacterial Community and Concentration of Volatile Fatty Acids in the Rumen during the Transition Period and Ketosis in Dairy Cows. Appl. Environ. Microbiol. 2012, 78, 2386–2392. [Google Scholar] [CrossRef] [Green Version]
- Ungerfeld, E.M. Shifts in metabolic hydrogen sinks in the methanogenesis-inhibited ruminal fermentation: A meta-analysis. Front. Microbiol. 2015, 6, 37. [Google Scholar] [CrossRef]
- Liu, L.; Sun, D.; Mao, S.; Zhu, W.; Liu, J. Infusion of sodium butyrate promotes rumen papillae growth and enhances expression of genes related to rumen epithelial VFA uptake and metabolism in neonatal twin lambs. J. Anim. Sci. 2019, 97, 909–921. [Google Scholar] [CrossRef]
- Hassan, F.; Tang, Z.; Ebeid, H.M.; Li, M.; Peng, K.; Liang, X.; Yang, C. Consequences of herbal mixture supplementation on milk performance, ruminal fermentation, and bacterial diversity in water buffaloes. PeerJ 2021, 9, e11241. [Google Scholar] [CrossRef]
- Liu, K.; Zhang, Y.; Yu, Z.; Xu, Q.; Zheng, N.; Zhao, S.; Huang, G.; Wang, J. Ruminal microbiota-host interaction and its effect on nutrient metabolism. Anim. Nutr. 2021, 7, 49–55. [Google Scholar] [CrossRef]
- Bailoni, L.; Carraro, L.; Cardin, M.; Cardazzo, B. Active Rumen Bacterial and Protozoal Communities Revealed by RNA-Based Amplicon Sequencing on Dairy Cows Fed Different Diets at Three Physiological Stages. Microorganisms 2021, 9, 754. [Google Scholar] [CrossRef]
- Aguilar-Marin, S.B.; Betancur-Murillo, C.L.; Isaza, G.A.; Mesa, H.; Jovel, J. Lower methane emissions were associated with higher abundance of ruminal Prevotella in a cohort of Colombian buffalos. BMC Microbiol. 2020, 20, 364. [Google Scholar] [CrossRef]
- Frantzen, C.A.; Kot, W.; Pedersen, T.B.; Ardö, Y.M.; Broadbent, J.R.; Neve, H.; Hansen, L.H.; Dal Bello, F.; Østlie, H.M.; Kleppen, H.P.; et al. Genomic Characterization of Dairy Associated Leuconostoc Species and Diversity of Leuconostocs in Undefined Mixed Mesophilic Starter Cultures. Front. Microbiol. 2017, 8, 132. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.; Harris, H.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Li, M.; Hassan, F.U.; Tang, Z.; Guo, Y.; Liang, X.; Peng, L.; Xie, H.; Yang, C. Physiological, oxidative and metabolic responses of lactating water buffaloes to tropical climate of South China. Vet. Med. Sci. 2021, 7, 1696–1706. [Google Scholar] [CrossRef]
- Boutard, M.; Cerisy, T.; Nogue, P.Y.; Alberti, A.; Weissenbach, J.; Salanoubat, M.; Tolonen, A.C. Functional Diversity of Carbohydrate-Active Enzymes Enabling a Bacterium to Ferment Plant Biomass. PLoS Genet. 2014, 10, e1004773. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Zhou, G.; Tian, G.; Liu, Y.; Dong, N.; Li, L.; Zhang, S.; Chai, H.; Chen, Y.; Yang, Y. Changes in Rumen Microbiota Affect Metabolites, Immune Responses and Antioxidant Enzyme Activities of Sheep under Cold Stimulation. Animals 2021, 11, 712. [Google Scholar] [CrossRef]
- Kavitake, D.; Devi, P.B.; Shetty, P.H. Overviewof exopolysaccharides produced by Weissella genus—A review. Int. J. Biol. Macromol. 2020, 164, 2964–2973. [Google Scholar] [CrossRef]
- Bainbridge, M.L.; Cersosimo, L.M.; Wright, A.-D.G.; Kraft, J. Rumen bacterial communities shift across a lactation in Holstein, Jersey and Holstein x Jersey dairy cows and correlate to rumen function, bacterial fatty acid composition and production parameters. FEMS Microbiol. Ecol. 2016, 92, fiw059. [Google Scholar] [CrossRef] [Green Version]
- Park, T.; Ma, L.; Ma, Y.; Zhou, X.; Bu, D.; Yu, Z. Dietary energy sources and levels shift the multi-kingdom microbiota and functions in the rumen of lactating dairy cows. J. Anim. Sci. Biotechnol. 2020, 11, 66. [Google Scholar] [CrossRef]
- Xue, M.-Y.; Sun, H.-Z.; Wu, X.-H.; Liu, J.-X.; Guan, L.L. Multi-omics reveals that the rumen microbiome and its metabolome together with the host metabolome contribute to individualized dairy cow performance. Microbiome 2020, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- Yue, S.; Ding, S.; Zhou, J.; Yang, C.; Hu, X.; Zhao, X.; Wang, Z.; Wang, L.; Peng, Q.; Xue, B. Metabolomics Approach Explore Diagnostic Biomarkers and Metabolic Changes in Heat-Stressed Dairy Cows. Animals 2020, 10, 1741. [Google Scholar] [CrossRef]
- Cowley, F.C.; Barber, D.G.; Houlihan, A.V.; Poppi, D.P. Immediate and residual effects of heat stress and restricted intake on milk protein and casein composition and energy metabolism. J. Dairy Sci. 2015, 98, 2356–2368. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Gao, S.; Quan, S.; Zhang, Y.; Bu, D.; Wang, J. Blood amino acids profile responding to heat stress in dairy cows. Asian-Australas. J. Anim. Sci. 2018, 31, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Eger, M.; Hussen, J.; Koy, M.; Dänicke, S.; Schuberth, H.J.; Breves, G. Glucose transporter expression differs between bovine monocyte and macrophage subsets and is influenced by milk production. J. Dairy Sci. 2016, 99, 2276–2287. [Google Scholar] [CrossRef] [Green Version]
- Meijer, A.J. Amino acids as regulators and components of nonproteinogenic pathways. J. Nutr. 2003, 133, 2057s–2062s. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Wang, C.; Liang, N.; Liu, Z.; Li, X.; Zhu, Z.J.; Merriman, T.R.; Dalbeth, N.; Terkeltaub, R.; Li, C.; et al. Serum Metabolomics Identifies Dysregulated Pathways and Potential Metabolic Biomarkers for Hyperuricemia and Gout. Arthritis Rheumatol. 2021, 73, 1738–1748. [Google Scholar] [CrossRef] [PubMed]
- Davoudi, P.; Daneshyar, M. Effect of different levels of tryptophane on performance, carcass characteristics and blood parameters of broiler chickens under heat stress condition. J. Vet. Res. 2017, 72, 157–164. [Google Scholar]
- Zhang, S.; Zeng, X.; Ren, M.; Mao, X.; Qiao, S. Novel metabolic and physiological functions of branched chain amino acids: A review. J. Anim. Sci. Biotechnol. 2017, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, H.; Zheng, N.; Wang, W.; Cheng, J.; Li, S.; Zhang, Y.; Wang, J. Integrated Metabolomics Study of the Milk of Heat-stressed Lactating Dairy Cows. Sci. Rep. 2016, 6, 24208. [Google Scholar] [CrossRef] [PubMed]
- Belenky, P.; Bogan, K.L.; Brenner, C. NAD(+) metabolism in health and disease. Trends Biochem. Sci. 2007, 32, 12–19. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Content |
|---|---|
| Ingredient (%, DM basis) | |
| Silage corn | 30 |
| Rice straw | 45 |
| Peanut vine | 10 |
| Corn | 7 |
| Rice bran | 2.5 |
| Soybean meal | 3 |
| Wheat bran | 2.5 |
| Mineral lick 1 | |
| Nutrient composition | |
| DM, % | 67.50 |
| NDF, % of DM | 32.74 |
| ADF, % of DM | 20.05 |
| CP, % of DM | 11.60 |
| Ca, % of DM | 0.70 |
| P, % of DM | 0.50 |
| GE, Mcal/kg of DM | 7.58 |
| Index | NHS | HS | SEM | p-Value |
|---|---|---|---|---|
| Physiological parameters | ||||
| Respiratory rate (bpm) | 46.39 | 66.27 | 6.74 | 0.005 |
| Rectal temperature (°C) | 38.74 | 38.80 | 0.14 | 0.831 |
| Skin temperature (°C) | 35.66 | 38.22 | 0.71 | 0.010 |
| Blood parameters | ||||
| HSP70 (ng/mL) | 395.91 | 568.79 | 117.77 | 0.126 |
| HSP90 (ng/mL) | 236.84 | 376.33 | 113.90 | 0.525 |
| Animal performance | ||||
| DMI (kg/d) | 13.03 | 11.57 | 1.29 | 0.456 |
| Milk yield (kg/d) | 4.10 | 2.93 | 1.72 | 0.480 |
| Fat (%) | 8.66 | 7.71 | 1.07 | 0.415 |
| Protein (%) | 4.76 | 4.47 | 0.49 | 0.064 |
| Milk urea nitrogen (mg/dL) | 14.64 | 20.48 | 3.42 | 0.060 |
| SCS | 3.68 | 3.88 | 1.26 | 0.581 |
| Rumen fermentation | ||||
| pH | 6.85 | 6.63 | 0.15 | 0.031 |
| Acetic acid (mmol/L) | 44.72 | 30.07 | 6.38 | 0.030 |
| Propionic acid (mmol/L) | 13.29 | 11.22 | 1.86 | 0.467 |
| Butyric acid (mmol/L) | 9.38 | 7.98 | 0.81 | 0.016 |
| TVFA (mmol/L) | 67.39 | 49.27 | 8.35 | 0.038 |
| A:P | 3.36 | 2.76 | 0.32 | 0.089 |
| NH3N (mg/dL) | 5.33 | 10.76 | 2.49 | 0.059 |
| Index | NHS | HS | SEM | p-Value |
|---|---|---|---|---|
| Sobs | 1487.40 | 1581.80 | 89.51 | 0.351 |
| ACE | 1738.79 | 1842.35 | 139.09 | 0.498 |
| Shannon | 5.43 | 5.68 | 0.15 | 0.170 |
| Coverage | 0.99 | 0.99 | 0.00 | 0.835 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Niu, K.; Rushdi, H.E.; Zhang, M.; Fu, T.; Gao, T.; Yang, L.; Liu, S.; Lin, F. Heat Stress Induces Shifts in the Rumen Bacteria and Metabolome of Buffalo. Animals 2022, 12, 1300. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101300
Wang Z, Niu K, Rushdi HE, Zhang M, Fu T, Gao T, Yang L, Liu S, Lin F. Heat Stress Induces Shifts in the Rumen Bacteria and Metabolome of Buffalo. Animals. 2022; 12(10):1300. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101300
Chicago/Turabian StyleWang, Zichen, Kaifeng Niu, Hossam E. Rushdi, Mingyue Zhang, Tong Fu, Tengyun Gao, Liguo Yang, Shenhe Liu, and Feng Lin. 2022. "Heat Stress Induces Shifts in the Rumen Bacteria and Metabolome of Buffalo" Animals 12, no. 10: 1300. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101300