
Effects of Vitamin A on Yanbian Yellow Cattle and Their Preadipocytes by Activating AKT/mTOR Signaling Pathway and Intestinal Microflora

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Adipogenic Differentiation
2.2. Measurement of Cell Proliferation
2.3. Cell Cycle Analysis
2.4. Oil Red O Staining
2.5. RNA Extraction and Quantitative Polymerase Chain Reaction Analysis
2.6. Protein Extraction and Western Blotting Analysis
2.7. Animals and Feeding
2.8. Diversity Analysis of Intestinal Flora of Yanbian Yellow Cattle
2.9. Statistical Analysis
3. Results
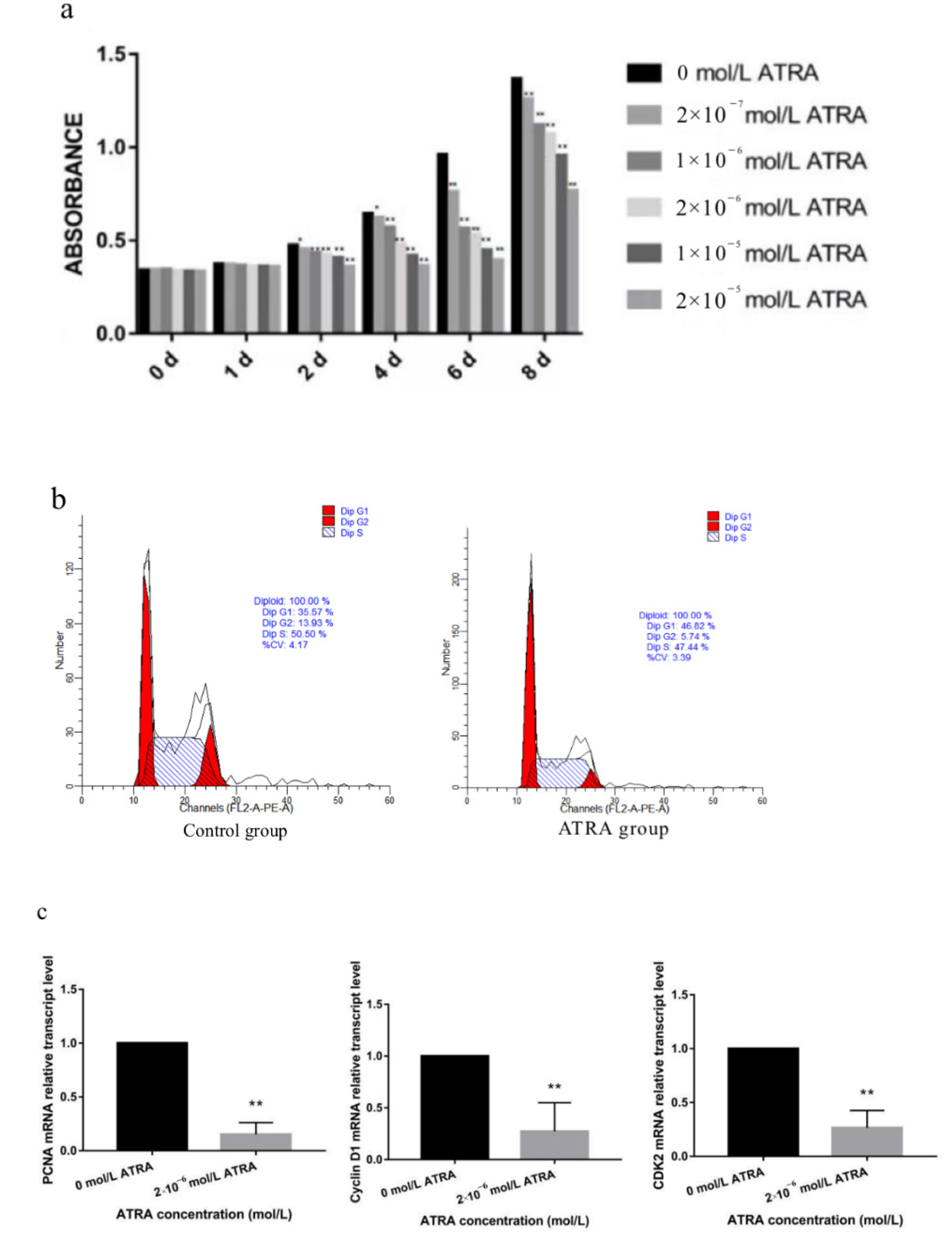
3.1. Effect of ATRA on the Proliferation of Preadipocytes from Yanbian Yellow Cattle
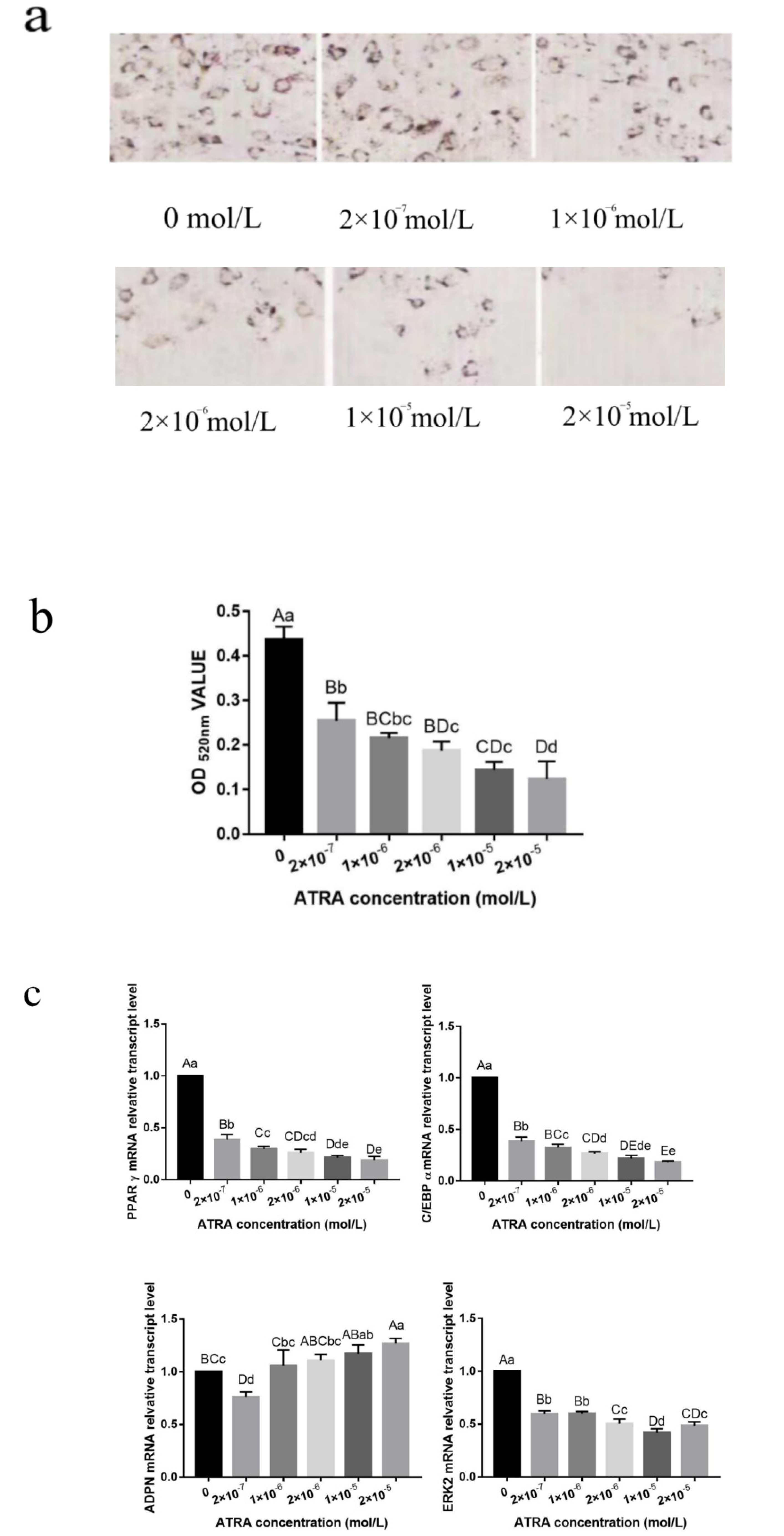
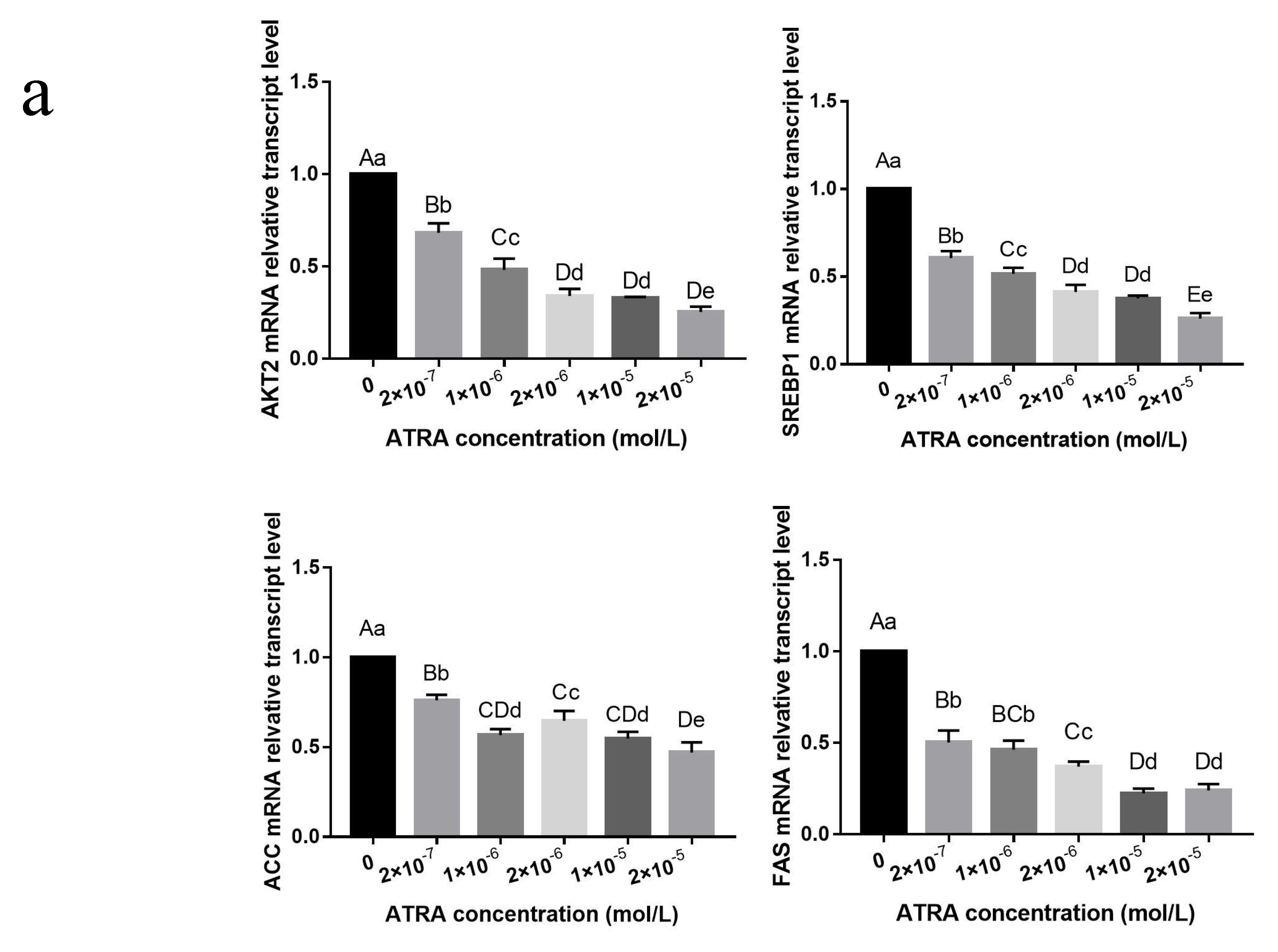
3.2. Effect of ATRA on the Differentiation of Preadipocytes from Yanbian Yellow Cattle
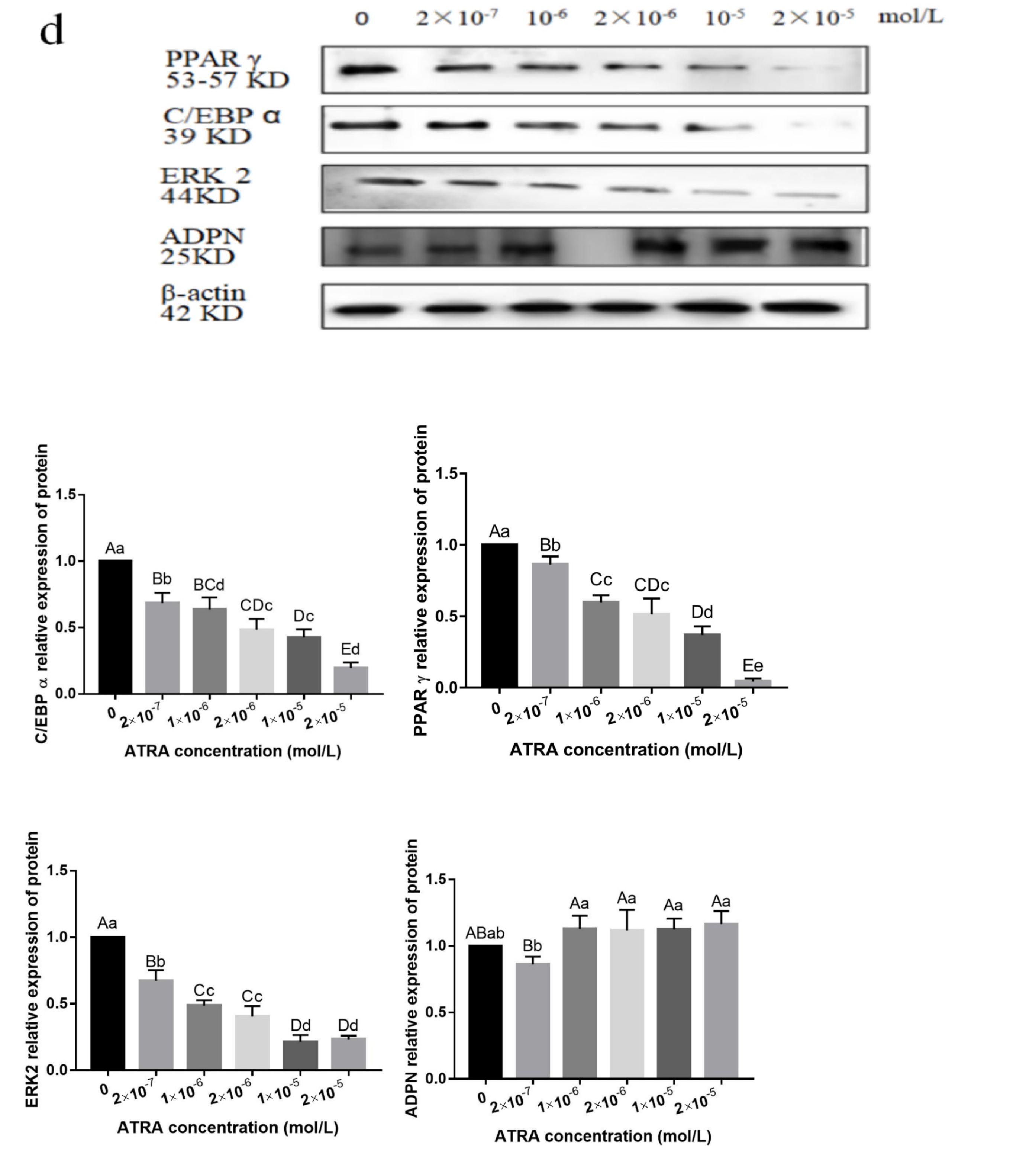
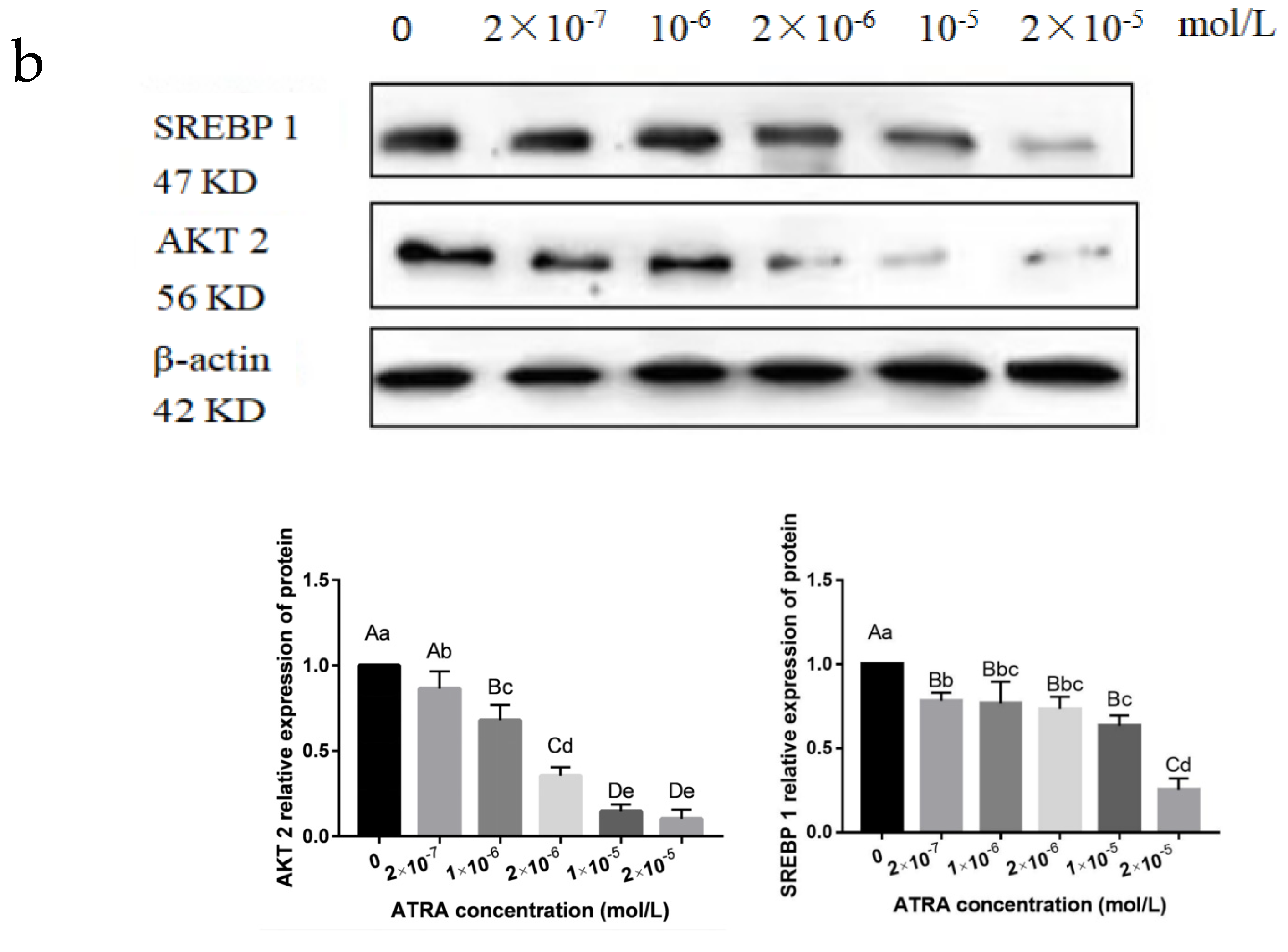
3.3. ATRA Regulated the Differentiation of Preadipocytes from Yanbian Yellow Cattle through the mTOR Signaling Pathway
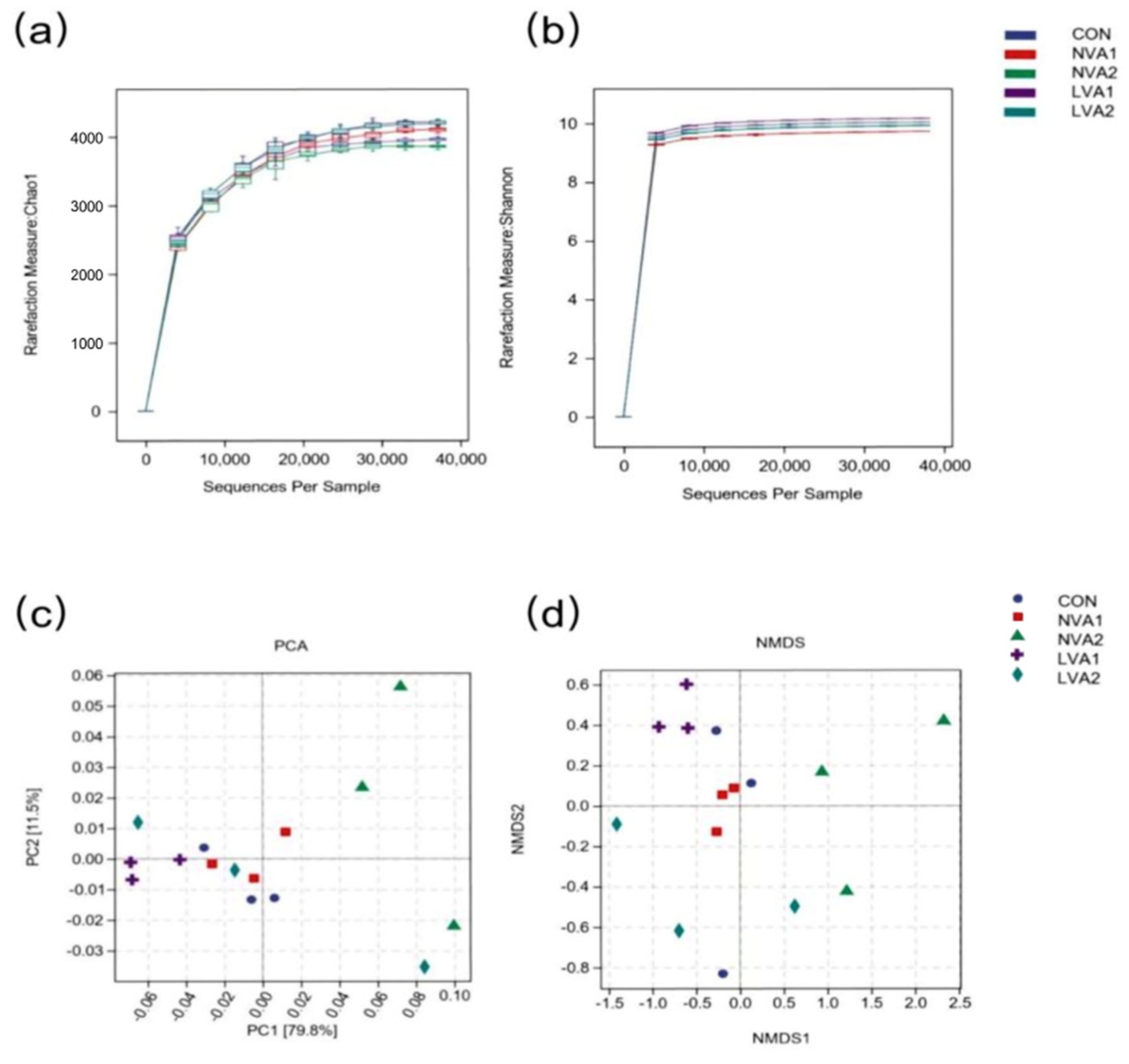
3.4. Effect of Vitamin A on the Diversity of the Intestinal Flora of Yanbian Yellow Cattle
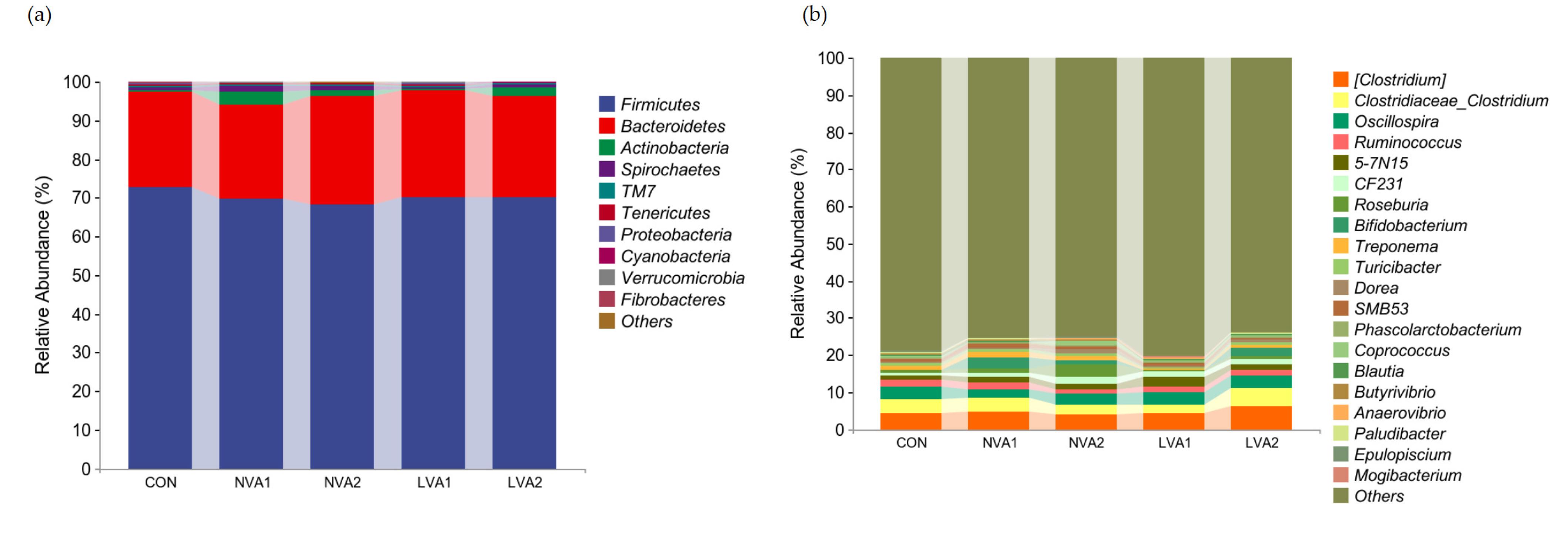
3.5. Vitamin A Regulated the Bacterial Composition in Yanbian Yellow Cattle at Different Taxa Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cappai, M.G.; Lunesu, M.G.; Accioni, F.; Liscia, M.; Pusceddu, M.; Burrai, L.; Nieddu, M.; Dimauro, C.; Boatto, G.; Pinna, W. Blood serum retinol levels in Asinara white donkeys reflect albinism-induced metabolic adaptation to photoperiod at Mediterranean latitudes. Ecol. Evol. 2016, 7, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Chen, G. The Roles of Vitamin A in the Regulation of Carbohydrate, Lipid, and Protein Metabolism. J. Clin. Med. 2014, 3, 453–479. [Google Scholar] [CrossRef] [Green Version]
- Oka, A.; Maruo, Y.; Miki, T.; Yamasaki, T.; Saito, T. Influence of Vitamin A on the quality of beef from the Tajima strain of Japanese Black cattle. Meat Sci. 1998, 48, 159–167. [Google Scholar] [CrossRef]
- Knutson, E.E.; Menezes, A.C.B.; Sun, X.; Fontoura, A.B.P.; Liu, J.H.; Bauer, M.L.; Maddock-Carlin, K.R.; Swanson, K.C.; Ward, A.K. Effect of feeding a low-vitamin A diet on carcass and production characteristics of steers with a high or low propensity for marbling. Animal 2020, 14, 2308–2314. [Google Scholar] [CrossRef]
- Lee, S.-A.; Jiang, H.; Feranil, J.B.; Brun, P.-J.; Blaner, W.S. Adipocyte-Specific Expression of a Retinoic Acid Receptor α Dominant Negative Form Causes Glucose Intolerance and Hepatic Steatosis in Mice. Biochem. Biophys. Res. Commun. 2019, 514, 1231–1237. [Google Scholar] [CrossRef]
- Xu, Q.; Fan, Y.; Loor, J.J.; Liang, Y.; Sun, X.; Jia, H.; Zhao, C.; Xu, C. All-Trans Retinoic Acid Controls Differentiation, Proliferation, and Lipolysis in Isolated Subcutaneous Adipocytes from Peripartal Holstein Cows. J. Dairy Sci. 2021, 104, 4999–5008. [Google Scholar] [CrossRef]
- Yan, C.; Wang, Y.; Piao, S.; Piao, X.; Piao, H.; Li, W. Study on the Quality Characteristics of Beef of Yanbian Yellow Cattle. China Cattle Sci. 2004, 30, 5–7. [Google Scholar]
- Hausman, G.J.; Dodson, M.V.; Ajuwon, K.; Azain, M.; Barnes, K.M.; Guan, L.L.; Jiang, Z.; Poulos, S.P.; Sainz, R.D.; Smith, S.; et al. BOARD-INVITED REVIEW: The Biology and Regulation of Preadipocytes and Adipocytes in Meat Animals1,2. J. Anim. Sci. 2009, 87, 1218–1246. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhang, X.; Guo, S.; Zhang, J.; Yin, B.; Zhang, L.; Xia, G. Research Progress on Vitamin a on Fat Deposition and Its Regulation in Beef Cattle. China Anim. Husb. Vet. Med. 2021, 48, 3652–3659. [Google Scholar]
- Ren, X. Biological Function of Adipose Tissue. J. Shangqiu Norm. Univ. 2013, 29, 87–89. [Google Scholar]
- Ali, A.T.; Hochfeld, W.E.; Myburgh, R.; Pepper, M.S. Adipocyte and Adipogenesis. Eur. J. Cell Biol. 2013, 92, 229–236. [Google Scholar] [CrossRef]
- Waldén, T.B.; Hansen, I.R.; Timmons, J.A.; Cannon, B.; Nedergaard, J. Recruited vs. Nonrecruited Molecular Signatures of Brown, “Brite,” and White Adipose Tissues. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E19–E31. [Google Scholar] [CrossRef] [Green Version]
- Hagiwara, A.; Cornu, M.; Cybulski, N.; Polak, P.; Betz, C.; Trapani, F.; Terracciano, L.; Heim Markus, H.; Rüegg Markus, A.; Hall Michael, N. Hepatic MTORC2 Activates Glycolysis and Lipogenesis through Akt, Glucokinase, and SREBP1c. Cell Metab. 2012, 15, 725–738. [Google Scholar] [CrossRef] [Green Version]
- Schoeler, M.; Caesar, R. Dietary Lipids, Gut Microbiota and Lipid Metabolism. Rev. Endocr. Metab. Disord. 2019, 20, 461–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishimwe, N.; Daliri, E.B.; Lee, B.H.; Fang, F.; Du, G. The perspective on cholesterol-lowering mechanisms of probiotics. Mol. Nutr. Food Res. 2015, 59, 94–105. [Google Scholar] [CrossRef]
- Hooper, L.V.; Wong, M.H.; Thelin, A.; Hansson, L.; Falk, P.G.; Gordon, J.I. Molecular analysis of commensal host-microbial relationships in the intestine. Science 2001, 291, 881–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullish, B.H.; Pechlivanis, A.; Barker, G.F.; Thursz, M.R.; Marchesi, J.R.; McDonald, J.A.K. Functional Microbiomics: Evaluation of Gut Microbiota-Bile Acid Metabolism Interactions in Health and Disease. Methods 2018, 149, 49–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tourniaire, F.; Gouranton, E.; von Lintig, J.; Keijer, J.; Luisa Bonet, M.; Amengual, J.; Lietz, G.; Landrier, J.-F. β-Carotene Conversion Products and Their Effects on Adipose Tissue. Genes Nutr. 2009, 4, 179–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, F. Effects of Vitamin a on Beef Quality of Limousin × Luxi Crossbred Steers and Action Mechanisms. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2005. [Google Scholar]
- Wang, D.; Jin, Y.; Shan, A.; Cheng, B.; Fang, H.; Zhao, Y.; Han, Y.; Wu, G.; Shen, J.; Zhang, J. Effects of All Trans Retinoic Acid on Differentiation and Expression of PPARγ and C/EBPα MRNA in 3T3-L1 Preadipocytes. Chin. J. Vet. Sci. 2015, 35, 647–649. [Google Scholar]
- Wang, P.; Xiong, Y.; Yang, G.; Shen, Q.; Pang, W. Knockdown of Akt2 by Lentivirus Vector-Mediated RNA Interference Suppresses Differentiation of Porcine Preadipocytes. Chin. J. Biochem. Mol. Biol. 2012, 28, 53–60. [Google Scholar]
- Jideonwo, V.; Hou, Y.; Ahn, M.; Surendran, S.; Morral, N. Impact of Silencing Hepatic SREBP-1 on Insulin Signaling. PLoS ONE 2018, 13, e0196704. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.B.; Moon, H.M.; Kim, W.S.; Lee, Y.S.; Jeong, H.W.; Yoo, E.J.; Ham, J.; Kang, H.; Park, M.-G.; Steffensen, K.R.; et al. Activated Liver X Receptors Stimulate Adipocyte Differentiation through Induction of Peroxisome Proliferator-Activated Receptor γ Expression. Mol. Cell. Biol. 2004, 24, 3430–3444. [Google Scholar] [CrossRef] [Green Version]
- Novakofski, J. Adipogenesis: Usefulness of in Vitro and in Vivo Experimental Models1,2. J. Anim. Sci. 2004, 82, 905–915. [Google Scholar] [CrossRef]
- Wang, Y.; Viscarra, J.; Kim, S.-J.; Sul, H.S. Transcriptional Regulation of Hepatic Lipogenesis. Nat. Rev. Mol. Cell Biol. 2015, 16, 678–689. [Google Scholar] [CrossRef] [Green Version]
- Brandebourg, T.D.; Hu, C.Y. Regulation of Differentiating Pig Preadipocytes by Retinoic Acid. J. Anim. Sci. 2005, 83, 98–107. [Google Scholar] [CrossRef]
- Komiya, Y.; Habas, R. Wnt Signal Transduction Pathways. Organogenesis 2008, 4, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X. The Influence of Intramuscular Fat Deposit and Gene Expression of ACC/HSL and PPARγ with the Supplemetation of Vitamin a in Beef Cattle. Master Thesis, Northwest A&F University, Shanxi, China, 2005. [Google Scholar]
- Simpson, K.L.; Chichester, C.O. Metabolism and Nutritional Significance of Carotenoids. Annu. Rev. Nutr. 1981, 1, 351–374. [Google Scholar] [CrossRef] [PubMed]
- Yan, H. Study of β-Carotene Digestion and Uptake in Rumen and Intestinal of Goats and Its Anti-Oxidative Effect. Master Thesis, Yangzhou University, Jiangsu, China, 2007. [Google Scholar]
- Saeed, A.; Hoekstra, M.; Hoeke, M.O.; Heegsma, J.; Faber, K.N. The Interrelationship between Bile Acid and Vitamin a Homeostasis. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2017, 1862, 496–512. [Google Scholar] [CrossRef]
- Sun, X.; Yang, G. Effect of Bile Acids on Glucose and Lipid Metabolism. Int. J. Endocrinol. Metab. 2008, 5, 323–325. [Google Scholar]
- Chiang, J.Y. Bile Acid Metabolism and Signaling. Compr. Physiol. 2013, 3, 1191–1212. [Google Scholar] [PubMed] [Green Version]
- Zeng, Z.; Liu, R.; Tang, L.; Li, W. Interaction between Gut Microbiota and Bile Acid Metabolism. Chin. J. Microecol. 2021, 33, 849–856. [Google Scholar]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial Signature of Dysbiosis in Gut Microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Human Microbiome Jumpstart Reference Strains Consortium; Nelson, K.E.; Weinstock, G.M.; Highlander, S.K.; Worley, K.C.; Creasy, H.H.; Wortman, J.R.; Rusch, D.B.; Mitreva, M.; Sodergren, E.; et al. A Catalog of Reference Genomes from the Human Microbiome. Science 2010, 328, 994–999. [Google Scholar]
- Yang, J.; Li, Y.; Wen, Z.; Liu, W.; Meng, L.; Huang, H. Oscillospira—A Candidate for the Next-Generation Probiotics. Gut Microbes 2021, 13, 1987783. [Google Scholar] [CrossRef]
- Wan, Y.; Wang, F.; Yuan, J.; Li, J.; Jiang, D.; Zhang, J.; Li, H.; Wang, R.; Tang, J.; Huang, T.; et al. Effects of Dietary Fat on Gut Microbiota and Faecal Metabolites, and Their Relationship with Cardiometabolic Risk Factors: A 6-Month Randomised Controlled-Feeding Trial. Gut 2019, 68, 1417–1429. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | GenBank Accession No. | Primer Sequence (5′-3′) |
|---|---|---|
| Cyclin D1-F | NM_001046273.2 | GCGTACCCTGACACCAATCTC |
| Cyclin D1-R | CTCCTCTTCGCACTTCTGCTC | |
| CDK 2-F | NM_001014934.1 | GGGTCCCTGTTCGTACTTATAC |
| CDK 2-R | CCACTGCTGTGGAGTAGTATTT | |
| PCNA-F | NM_001034494.1 | CCTTGGTGCAGCTAACCCTT |
| PCNA-R | TTGGACATGCTGGTGAGGTT | |
| β-actin-F | NM_173979.3 | AGGCATCCTGACCCTCAAGTA |
| β-actin-R | GCTCGTTGTAGAAGGTGTGGT | |
| ERK 2-F | NM_175793.2 | AAGACGCAACACCTCAGCA |
| ERK 2-R | AAGACGCAACACCTCAGCA | |
| PPAR γ-F | NM_181024.2 | CGAGAAGGAGAAGCTGTTGG |
| PPAR γ-R | TCAGCGGGAAGGACTTTATG | |
| C/EBP α-F | NM_176784.2 | TGGACAAGAACAGCAACGAG |
| C/EBP α-R | TCACTGGTCAACTCCAGCAC | |
| ADPN-F | NM_015462053.1 | AGGCAGAAAGGGAGAACC |
| ADPN-R | GTCGTGGTGAAGAGCAG | |
| FAS-F | NM_001012669 | AGGACCTCGTGAAGGCTGTGA |
| FAS-R | CCAAGGTCTGAAAGCGAGCTG | |
| ACC-F | AJ132890 | CATCTTGTCCGAAACGTCGAT |
| ACC-R | CCCTTCGAACATACACCTCCA | |
| SREBP 1-F | NM_001113302 | CTTGGAGCGAGCACTGAATT |
| SREBP 1-R | GGGCATCTGAGAACTCCTTGTC | |
| AKT 2-F | NM_001206146.2 | GCCGAATAGGAGAACTGGGG |
| AKT 2-R | CACGTCTGAGGTCGACACAA |
| Raw Materials | Content (%) | Nutritional Ingredient | Content (%) |
|---|---|---|---|
| corn | 35.52 | dry matter | 87.63 |
| soybean meal | 14.40 | crude protein | 13.70 |
| wheat bran | 6.00 | crude fat | 3.20 |
| straw | 40.00 | crude ash | 10.69 |
| salt | 0.48 | crude fibre | 11.81 |
| sodium bicarbonate | 0.60 | calcium | 0.96 |
| premix | 3.00 | phosphorus | 0.62 |
| total | 100 | Nacl | 0.50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Xu, H.; Zhang, C.; Bai, J.; Song, J.; Hao, B.; Zhang, L.; Xia, G. Effects of Vitamin A on Yanbian Yellow Cattle and Their Preadipocytes by Activating AKT/mTOR Signaling Pathway and Intestinal Microflora. Animals 2022, 12, 1477. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121477
Zhang X, Xu H, Zhang C, Bai J, Song J, Hao B, Zhang L, Xia G. Effects of Vitamin A on Yanbian Yellow Cattle and Their Preadipocytes by Activating AKT/mTOR Signaling Pathway and Intestinal Microflora. Animals. 2022; 12(12):1477. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121477
Chicago/Turabian StyleZhang, Xinxin, Hongyan Xu, Congcong Zhang, Jinhui Bai, Jixuan Song, Beibei Hao, Luomeng Zhang, and Guangjun Xia. 2022. "Effects of Vitamin A on Yanbian Yellow Cattle and Their Preadipocytes by Activating AKT/mTOR Signaling Pathway and Intestinal Microflora" Animals 12, no. 12: 1477. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121477