
Effects of Dietary Fiber Type on Growth Performance, Serum Parameters and Fecal Microbiota Composition in Weaned and Growing-Finishing Pigs

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Diets and Experimental Design
2.2. Feeding and Management
2.3. Sample Collection
2.4. Lab Analysis
2.4.1. Chemical Composition
2.4.2. Serum Parameters
2.4.3. Fecal Microbiota Community
2.4.4. Fecal VFA Concentration
2.5. Statistical Analysis
3. Results
3.1. Effects of Different SDF to IDF Ratio on Growth Performance of Weaned and Growing-Finishing Pigs
3.2. Effects of Different SDF to IDF Ratio on Serum Immune and Antioxidance of Weaned and Growing-Finishing Pigs
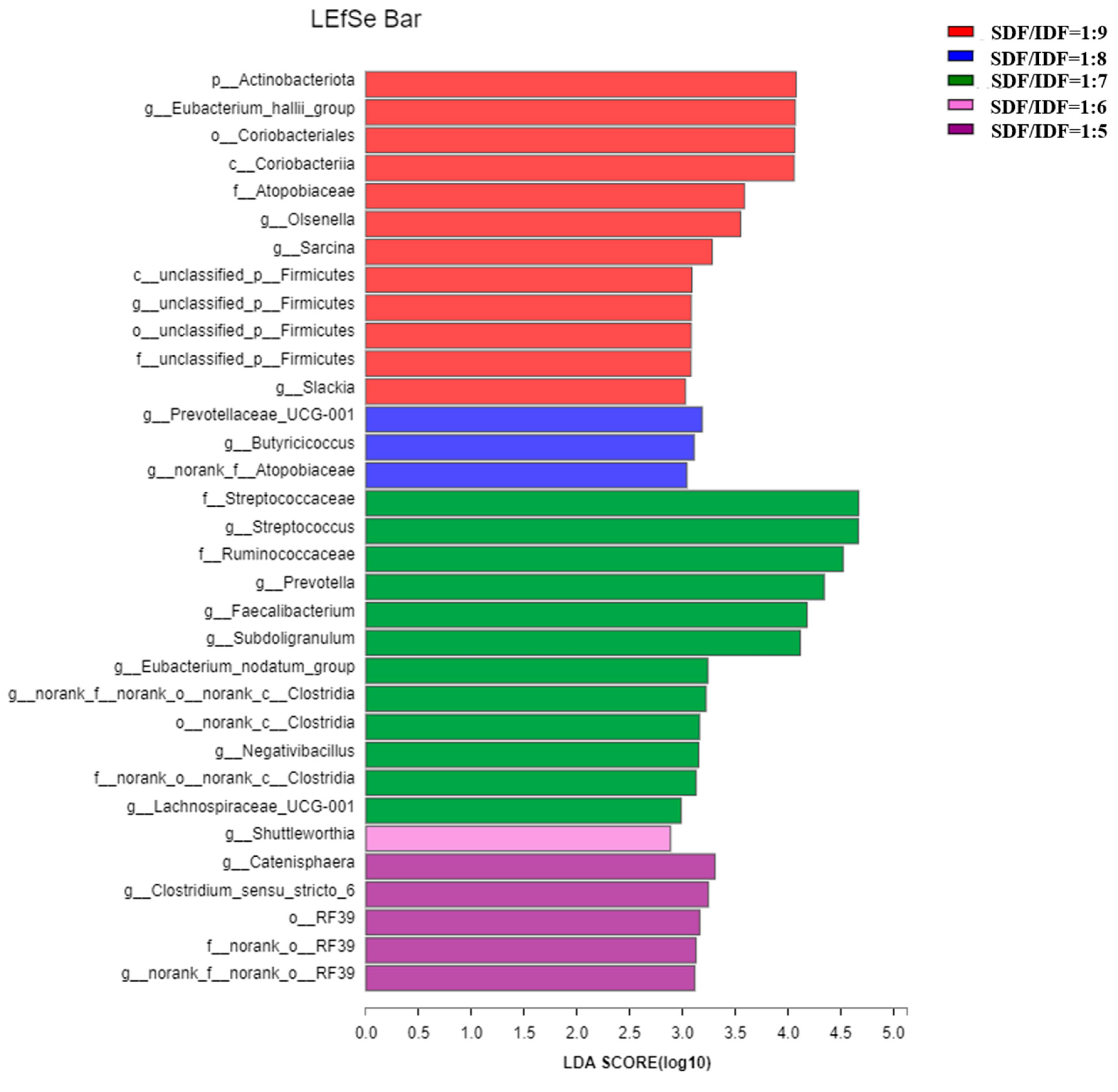
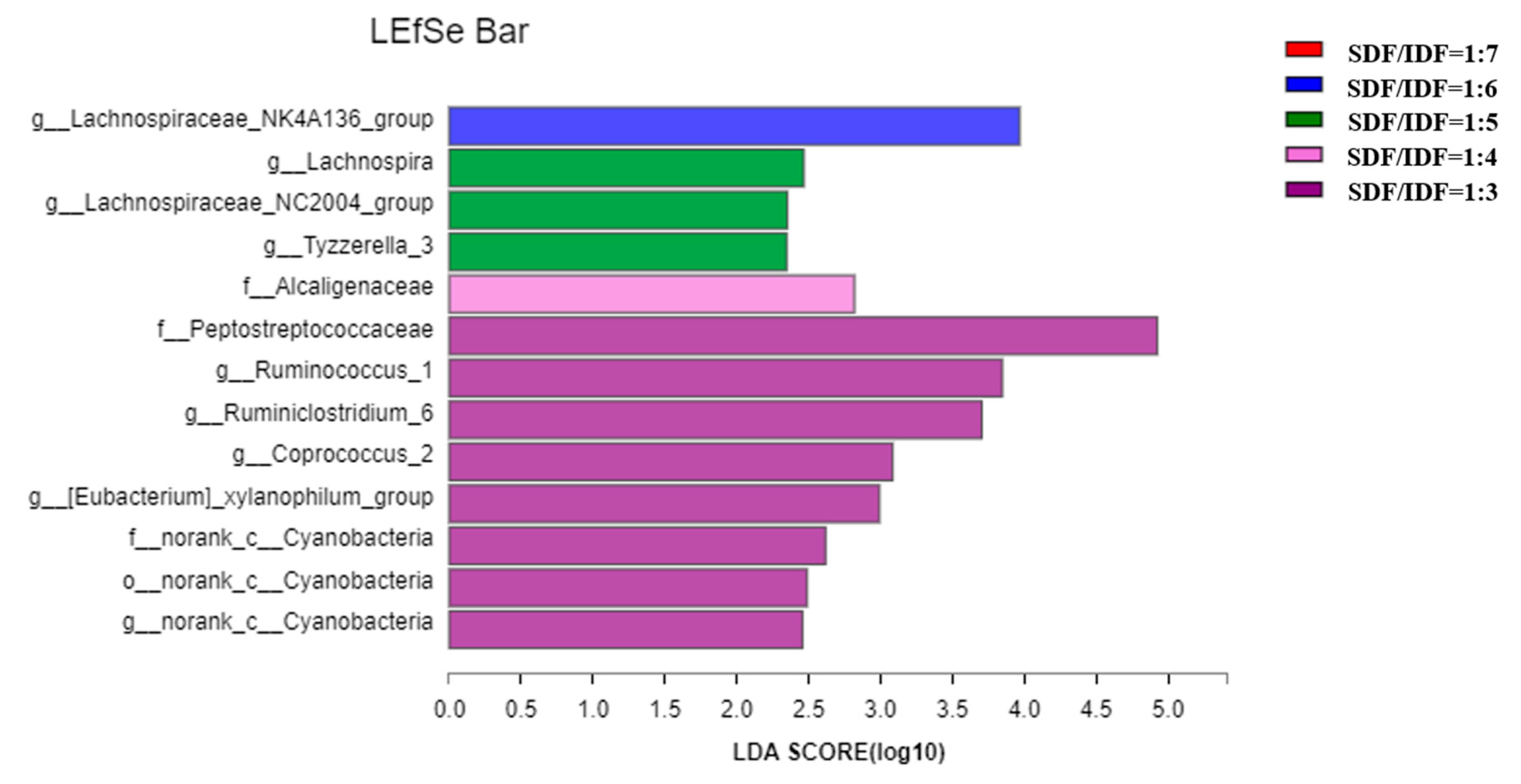
3.3. Effects of Different SDF to IDF Ratio on Fecal Microbiota Community of Weaned and Growing-Finishing Pigs
3.4. Effects of Different SDF to IDF Ratio on Fecal VFA Concentration of Weaned and Growing-Finishing Pigs
4. Discussion
4.1. Effects of Different SDF/IDF on Growth Performance
4.2. Effects of Different SDF/IDF on Serum Immune and Antioxidant Capacity
4.3. Effects of Different SDF/IDF on Fecal Microbiota and Their Metabolites
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Williams, B.A.; Verstegen, M.W.A.; Tamminga, S. Fermentation in the large intestine of single-stomached animals and its relationship to animal health. Nutr. Res. Rev. 2001, 14, 207–222. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Zac-Varghese, S.E.K.; MacDougall, K.; Preston, T.; Tedford, C.; Finlayson, G.S.; et al. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut 2015, 64, 1744–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.B.; Liu, P.; Wu, Y.; Guo, P.T.; Liu, L.; Ma, N.; Zhang, J.; Ma, X. Dietary fiber increases butyrate-producing bacteria and improves the growth performance of weaned piglets. J. Agric. Food Chem. 2018, 66, 7995–8004. [Google Scholar] [CrossRef]
- Yu, C.; Zhang, S.; Yang, Q.; Peng, Q.; Zhu, J.; Zeng, X.; Qiao, S.Y. Effect of high fibre diets formulated with different fibrous ingredients on performance, nutrient digestibility and faecal microbiota of weaned piglets. Arch. Anim. Nutr. 2016, 70, 263–277. [Google Scholar] [CrossRef]
- Knudsen, K.E.B. The nutritional significance of “dietary fibre” analysis. Anim. Feed Sci. Technol. 2001, 90, 3–20. [Google Scholar] [CrossRef]
- Yang, P.; Zhao, J.B. Fiber digestibility in growing pigs fed common fiber-rich ingredients: A systematic review. Ann. Anim. Sci. 2021, 22, 537–550. [Google Scholar] [CrossRef]
- Jha, R.; Berrocoso, J.D. Review: Dietary fiber utilization and its effects on physiological functions and gut health of swine. Animal 2015, 9, 1441–1452. [Google Scholar] [CrossRef] [Green Version]
- Jha, R.; Leterme, P. Feed ingredients differing in fermentable fibre and indigestible protein content affect fermentation metabolites and faecal nitrogen excretion in growing pigs. Animal 2012, 6, 603–612. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Piao, X.S.; Liu, L.; Li, D.F. Effects of inclusion level on nutrient digestibility and energy content of wheat middlings and soya bean meal for growing pigs. Arch. Anim. Nutr. 2013, 67, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.B.; Zhang, S.; Xie, F.; Li, D.F.; Huang, C.F. Effects of inclusion level and adaptation period on nutrient digestibility and digestible energy of wheat bran in growing-finishing pigs. Asian-Australas. J. Anim. Sci. 2018, 31, 116–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.F.; Zhang, S.; Stein, H.H.; Zhao, J.B.; Li, D.F.; Lai, C.H. Effect of inclusion level and adaptation duration on digestible energy and nutrient digestibility in palm kernel meal fed to growing-finishing pigs. Asian-Australas. J. Anim. Sci. 2018, 31, 395–402. [Google Scholar] [CrossRef] [PubMed]
- United States National Research Council. Nutrient Requirements of Swine, 11th ed.; National Academy Press: Washington, DC, USA, 2012. [Google Scholar]
- Hortwitz, W.; Latimer, G.W. Official Methods of Analysis of AOAC International, 18th ed.; Rev. 2. ed.; AOAC Int.: Gaithersburg, MD, USA, 2007. [Google Scholar]
- Porter, M.; Murray, R. The volatility of components of grass silage on oven drying and the inter-relationship between dry-matter content estimated by different analytical methods. Grass Forage Sci. 2001, 56, 405–411. [Google Scholar] [CrossRef] [Green Version]
- Weber, T.E.; Kerr, B.J. Metabolic effects of dietary sugar beet pulp or wheat bran in growing female pigs. J. Anim. Sci. 2012, 90, 523–532. [Google Scholar] [CrossRef]
- Wang, L.F.; Beltranena, E.; Zijlstra, R.T. Diet nutrient digestibility and growth performance of weaned pigs fed sugar beet pulp. Anim. Feed Sci. Technol. 2016, 211, 145–152. [Google Scholar] [CrossRef]
- Freire, J.P.B.; Guerreiro, A.J.G.; Cunha, L.F.; Aumaitre, A. Effect of dietary fibre source on total tract digestibility, caecum volatile fatty acids and digestive transit time in the weaned piglet. Anim. Feed Sci. Technol. 2000, 87, 71–83. [Google Scholar] [CrossRef]
- Molist, G.F.; Ywazaki, M.; Gómez, S.U.A.; Hermes, R.G.; Gasa, G.J.; Pérez, H.J.F. Administration of loperamide and addition of wheat bran to the diets of weaner pigs decrease the incidence of diarrhoea and enhance their gut maturation. Br. J. Nutr. 2010, 103, 879–885. [Google Scholar] [CrossRef] [Green Version]
- Varel, V.H.; Pond, W.G.; Pekas, J.C.; Yen, J.T. Influence of high-fiber diet on bacterial populations in gastrointestinal tracts of obese- and lean-genotype pigs. Appl. Environ. Microbiol. 1982, 44, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Magistrelli, D.; Galassi, G.; Crovetto, G.M.; Rosi, F. Influence of high levels of beet pulp in the diet on endocrine/metabolic traits, slaughter dressing percentage, and ham quality in Italian heavy pigs. Ital. J. Anim. Sci. 2009, 8, 37–49. [Google Scholar] [CrossRef]
- Hopwood, D.E.; Pethick, D.W.; Pluske, J.R.; Hampson, D.J. Addition of pearl barley to a rice-based diet for newly weaned piglets increases the viscosity of the intestinal contents, reduces starch digestibility and exacerbates post-weaning colibacillosis. Br. J. Nutr. 2004, 92, 419–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molist, F.; van Oostrum, M.; Pérez, J.F.; Mateos, G.G.; Nyachoti, C.M.; van der Aar, P.J. Relevance of functional properties of dietary fibre in diets for weanling pigs. Anim. Feed Sci. Technol. 2014, 189, 1–10. [Google Scholar] [CrossRef]
- Cox, M.A.; Jackson, J.; Stanton, M.; Rojas-Triana, A.; Bober, L.; Laverty, M.; Yang, X.; Zhu, F.; Liu, J.; Wang, S.; et al. Short-chain fatty acids act as antiinflammatory mediators by regulating prostaglandin E(2) and cytokines. World J. Gastroenterol. 2009, 15, 5549–5557. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The role of short-chain fatty acids in health and disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar]
- Macpherson, A.J.; McCoy, K.D.; Johansen, F.E.; Brandtzaeg, P. The immune geography of IgA induction and function. Mucosal Immunol. 2008, 1, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Cleophas, M.C.; Crişan, T.O.; Lemmers, H.; Toenhake-Dijkstra, H.; Fossati, G.; Jansen, T.L.; Dinarello, C.A.; Netea, M.G.; Joosten, L.A. Suppression of monosodium urate crystal-induced cytokine production by butyrate is mediated by the inhibition of class I histone deacetylases. Ann. Rheum. Dis. 2016, 75, 593–600. [Google Scholar] [CrossRef]
- Brahe, L.K.; Le Chatelier, E.; Prifti, E.; Pons, N.; Kennedy, S.; Hansen, T.; Pedersen, O.; Astrup, A.; Ehrlich, S.D.; Larsen, L.H. Specific gut microbiota features and metabolic markers in postmenopausal women with obesity. Nutr. Diabetes 2015, 5, e159. [Google Scholar] [CrossRef]
- Jha, R.; Rossnagel, B.; Pieper, R.; van Kessel, A.; Leterme, P. Barley and oat cultivars with diverse carbohydrate composition alter ileal and total tract nutrient digestibility and fermentation metabolites in weaned piglets. Animal 2010, 4, 724–731. [Google Scholar] [CrossRef] [Green Version]
- Jaworski, N.W.; Stein, H.H. Disappearance of nutrients and energy in the stomach and small intestine, cecum, and colon of pigs fed corn-soybean meal diets containing distillers dried grains with solubles, wheat middlings, or soybean hulls. J. Anim. Sci. 2017, 95, 727–739. [Google Scholar] [CrossRef]
- Urriola, P.E.; Shurson, G.C.; Stein, H.H. Digestibility of dietary fiber in distillers co-products fed to growing pigs. J. Anim. Sci. 2010, 88, 2373–2381. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.B.; Bai, Y.; Zhang, G.; Liu, L.; Lai, C.H. Relationship between dietary fiber fermentation and volatile fatty acids’ concentration in growing pigs. Animals 2020, 10, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Ivarsson, E.; Dicksved, J.; Lundh, T.; Lindberg, J.E. Inclusion of chicory (Cichorium intybus L.) in pigs’ diets affects the intestinal microenvironment and the gut microbiota. Appl. Environ. Microbiol. 2012, 78, 4102–4109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Zhang, M.; Wang, Y.; Dorfman, R.G.; Liu, H.; Yu, T.; Chen, X.; Tang, D.; Xu, L.; Yin, Y.; et al. Faecalibacterium prausnitzii produces butyrate to maintain Th17/Treg balance and to ameliorate colorectal colitis by inhibiting histone deacetylase 1. Inflamm. Bowel Dis. 2018, 24, 1926–1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Items | Corn | Soybean Meal | Corn Bran | Palm Kernel Meal | Sugar Beet Pulp |
|---|---|---|---|---|---|
| Total dietary fiber | 9.1 | 22.2 | 53.1 | 49.0 | 69.0 |
| Soluble dietary fiber | 1.0 | 4.2 | 5.0 | 3.1 | 27.4 |
| Insoluble dietary fiber | 8.1 | 17.9 | 48.1 | 46.2 | 42.5 |
| Neutral detergent fiber | 8.2 | 11.1 | 44.0 | 44.8 | 37.7 |
| Acid detergent fiber | 2.1 | 6.2 | 14.2 | 24.6 | 21.2 |
| Gross energy | 15.9 | 17.0 | 17.3 | 17.9 | 16.2 |
| Dry matter | 85.2 | 87.4 | 92.1 | 90.0 | 86.9 |
| Crude protein | 7.7 | 42.1 | 14.8 | 15.6 | 8.7 |
| Ether extract | 2.5 | 1.4 | 3.9 | 5.6 | 0.4 |
| Items, % | SDF/IDF | ||||
|---|---|---|---|---|---|
| 1:5 | 1:6 | 1:7 | 1:8 | 1:9 | |
| Sugar beet pulp | 14.00 | 8.00 | 5.00 | 2.00 | 0.00 |
| Corn bran | 3.00 | 12.00 | 14.00 | 16.70 | 18.00 |
| Palm kernel meal | 0.00 | 0.00 | 2.50 | 4.00 | 5.28 |
| Corn starch | 52.53 | 51.89 | 50.70 | 49.81 | 50.29 |
| Whey powder | 7.00 | 5.00 | 5.00 | 5.00 | 4.00 |
| Soy protein concentrate | 19.00 | 18.60 | 18.65 | 18.59 | 18.66 |
| Dicalcium phosphate | 1.60 | 1.65 | 1.50 | 1.40 | 1.35 |
| Limestone | 0.80 | 0.75 | 0.85 | 0.90 | 0.95 |
| NaCl | 0.40 | 0.40 | 0.40 | 0.40 | 0.40 |
| L-Lysine-HCl | 0.63 | 0.65 | 0.46 | 0.35 | 0.26 |
| DL-Methionine | 0.18 | 0.18 | 0.17 | 0.16 | 0.16 |
| L-Threonine | 0.30 | 0.32 | 0.23 | 0.17 | 0.14 |
| L-Tryptophan | 0.06 | 0.06 | 0.04 | 0.02 | 0.01 |
| Premix 1 | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 |
| Total | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 |
| Nutritive levels, % 2 | |||||
| DE, Kcal/kg | 3549 | 3546 | 3545 | 3540 | 3547 |
| CP | 15.33 | 15.30 | 15.33 | 15.30 | 15.32 |
| SID Lys | 1.35 | 1.35 | 1.35 | 1.35 | 1.35 |
| SID Met | 0.39 | 0.39 | 0.39 | 0.39 | 0.39 |
| SID Thr | 0.79 | 0.79 | 0.79 | 0.79 | 0.80 |
| SID Trp | 0.22 | 0.22 | 0.22 | 0.22 | 0.22 |
| TDF | 12.40 | 12.65 | 12.65 | 12.53 | 12.33 |
| SDF | 2.07 | 1.80 | 1.59 | 1.39 | 1.24 |
| IDF | 10.33 | 10.85 | 11.06 | 11.14 | 11.09 |
| Items, % | SDF/IDF | ||||
|---|---|---|---|---|---|
| 1:3 | 1:4 | 1:5 | 1:6 | 1:7 | |
| Corn | 71.33 | 74.17 | 72.59 | 73.59 | 73.92 |
| Soybean meal | 7 | 9 | 10 | 10 | 10 |
| Sugar beet pulp | 9.5 | 5.2 | 3.7 | 2 | 1 |
| Palm kernel meal | 0 | 1.8 | 5 | 6 | 7 |
| Soy protein concentrate | 4.8 | 3.5 | 3 | 3 | 3 |
| Soy oil | 2.3 | 1.6 | 1.3 | 1.1 | 0.9 |
| Dicalcium phosphate | 1.65 | 1.5 | 1.4 | 1.4 | 1.4 |
| Limestone | 0.95 | 1 | 1.1 | 1.1 | 1.1 |
| NaCl | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 |
| L-Lysine-HCl | 1.02 | 0.9 | 0.68 | 0.62 | 0.55 |
| DL-Methionine | 0.15 | 0.15 | 0.15 | 0.14 | 0.13 |
| L-Threonine | 0.4 | 0.3 | 0.22 | 0.2 | 0.16 |
| L-Tryptophan | 0.1 | 0.08 | 0.06 | 0.05 | 0.04 |
| Premix 1 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Total | 100 | 100 | 100 | 100 | 100 |
| Nutritive levels, % 2 | |||||
| DE, Kcal/kg | 3330 | 3331 | 3330 | 3335 | 3334 |
| CP | 14.80 | 14.73 | 14.76 | 14.75 | 14.70 |
| SID Lys | 1.34 | 1.34 | 1.35 | 1.36 | 1.36 |
| SID Met | 0.36 | 0.36 | 0.37 | 0.36 | 0.36 |
| SID Thr | 0.78 | 0.74 | 0.77 | 0.78 | 0.78 |
| SID Trp | 0.21 | 0.21 | 0.22 | 0.22 | 0.22 |
| TDF | 14.67 | 13.25 | 13.86 | 13.24 | 13.06 |
| SDF | 3.61 | 2.60 | 2.31 | 1.88 | 1.64 |
| IDF | 11.06 | 10.65 | 11.55 | 11.36 | 11.41 |
| Items | Different SDF/IDF Ratios | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 1:5 | 1:6 | 1:7 | 1:8 | 1:9 | Treatment | Linear | Quadratic | ||
| BW | |||||||||
| d 0 kg | 7.95 | 7.97 | 7.99 | 8.03 | 7.98 | 0.47 | 0.99 | 0.93 | 0.94 |
| d 14 kg | 12.12 | 12.11 | 12.32 | 12.41 | 12.07 | 0.59 | 0.99 | 0.91 | 0.73 |
| d 28 kg | 19.32 | 19.28 | 19.63 | 18.91 | 18.14 | 0.82 | 0.75 | 0.31 | 0.47 |
| d 0~14 | |||||||||
| ADG g/d | 299 | 296 | 309 | 314 | 293 | 13.43 | 0.77 | 0.90 | 0.38 |
| ADFI g/d | 442 | 416 | 444 | 442 | 430 | 15.23 | 0.71 | 0.96 | 0.99 |
| F/G | 1.48 | 1.41 | 1.46 | 1.41 | 1.47 | 0.05 | 0.77 | 0.89 | 0.36 |
| d 14~28 | |||||||||
| ADG g/d | 515 a | 512 a | 522 a | 464 a,b | 434 b | 21.91 | 0.03 | 0.01 | 0.15 |
| ADFI g/d | 796 | 781 | 829 | 741 | 726 | 32.72 | 0.22 | 0.09 | 0.23 |
| F/G | 1.55 | 1.53 | 1.59 | 1.60 | 1.68 | 0.04 | 0.13 | 0.02 | 0.38 |
| d 0~28 | |||||||||
| ADG g/d | 407 | 404 | 416 | 389 | 364 | 16.16 | 0.20 | 0.06 | 0.18 |
| ADFI g/d | 619 | 599 | 637 | 591 | 578 | 22.21 | 0.43 | 0.23 | 0.44 |
| F/G | 1.52 | 1.48 | 1.54 | 1.53 | 1.59 | 0.03 | 0.27 | 0.09 | 0.25 |
| Items | Different SDF/IDF Ratios | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 1:3 | 1:4 | 1:5 | 1:6 | 1:7 | Treatment | Linear | Quadratic | ||
| BW | |||||||||
| d 0 kg | 48.16 | 48.33 | 48.49 | 48.32 | 48.05 | 1.65 | 0.99 | 0.97 | 0.90 |
| d 28 kg | 74.27 | 75.74 | 77.24 | 75.44 | 74.49 | 3.54 | 0.99 | 0.99 | 0.64 |
| d 56 kg | 98.73 | 101.79 | 104.07 | 100.61 | 99.10 | 5.34 | 0.92 | 0.98 | 0.39 |
| d 0~28 | |||||||||
| ADG g/d | 933 | 979 | 1027 | 969 | 944 | 20.54 | 0.59 | 0.92 | 0.14 |
| ADFI g/d | 2477 | 2427 | 2537 | 2449 | 2414 | 37.54 | 0.97 | 0.82 | 0.75 |
| F/G | 2.65 | 2.47 | 2.47 | 2.54 | 2.55 | 0.07 | 0.34 | 0.51 | 0.09 |
| d 28–56 | |||||||||
| ADG g/d | 873 b | 930 a,b | 959 a | 899 a,b | 879 b | 21.65 | 0.04 | 0.75 | 0.01 |
| ADFI g/d | 2956 | 3041 | 3059 | 2899 | 2857 | 38.86 | 0.59 | 0.31 | 0.28 |
| F/G | 3.39 | 3.28 | 3.20 | 3.23 | 3.25 | 0.08 | 0.85 | 0.42 | 0.45 |
| d 0–56 | |||||||||
| ADG g/d | 903 | 955 | 993 | 934 | 912 | 24.65 | 0.08 | 0.95 | 0.01 |
| ADFI g/d | 2719 | 2737 | 2800 | 2676 | 2638 | 37.31 | 0.89 | 0.56 | 0.50 |
| F/G | 3.01 | 2.86 | 2.82 | 2.87 | 2.89 | 0.10 | 0.49 | 0.32 | 0.16 |
| Items | Different SDF/IDF Ratios | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 1:5 | 1:6 | 1:7 | 1:8 | 1:9 | Treatment | Linear | Quadratic | ||
| IgM g/L | 1.59 | 1.58 | 1.66 | 1.68 | 1.58 | 0.02 | 0.73 | 0.72 | 0.36 |
| IgA g/L | 2.52 | 2.48 | 2.52 | 2.49 | 2.59 | 0.02 | 0.94 | 0.62 | 0.61 |
| IgG g/L | 18.29 | 16.69 | 16.94 | 19.03 | 18.24 | 1.03 | 0.10 | 0.30 | 0.18 |
| SOD U/mL | 94.88 | 81.78 | 87.87 | 75.93 | 87.53 | 7.45 | 0.21 | 0.26 | 0.15 |
| GSH-Px U/mL | 309 | 314 | 331 | 310 | 317 | 16.07 | 0.20 | 0.50 | 0.28 |
| T-AOC U/mL | 7.54 | 7.70 | 8.54 | 8.68 | 7.63 | 0.32 | 0.13 | 0.51 | 0.08 |
| MDA nmol/mL | 5.61 | 5.08 | 5.27 | 5.31 | 5.79 | 0.18 | 0.52 | 0.39 | 0.41 |
| Items | Different SDF/IDF Ratios | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 1:3 | 1:4 | 1:5 | 1:6 | 1:7 | Treatment | Linear | Quadratic | ||
| Ig M g/L | 1.42 | 1.52 | 1.49 | 1.42 | 1.43 | 0.05 | 0.94 | 0.83 | 0.41 |
| Ig A g/L | 3.41 | 3.29 | 3.33 | 3.28 | 3.32 | 0.16 | 0.91 | 0.55 | 0.47 |
| Ig G g/L | 11.66 | 12.18 | 11.79 | 10.63 | 11.69 | 0.63 | 0.15 | 0.39 | 0.12 |
| SOD U/mL | 42.52 | 34.23 | 42.77 | 38.42 | 47.77 | 3.45 | 0.40 | 0.36 | 0.21 |
| GSH-Px U/mL | 197 | 195 | 225 | 229 | 201 | 13.34 | 0.21 | 0.50 | 0.07 |
| T-AOC U/mL | 6.36 | 5.24 | 6.28 | 5.58 | 6.62 | 0.38 | 0.18 | 0.55 | 0.12 |
| MDA nmol/mL | 3.65 b | 4.34 a | 3.66 b | 4.13 a,b | 3.89 a,b | 0.16 | 0.03 | 0.30 | 0.27 |
| Items | SDF/IDF | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 1:5 | 1:6 | 1:7 | 1:8 | 1:9 | One-Way | Linear | Quadratic | ||
| Shannon | 3.68 a,b | 3.60 a,b | 4.01 a | 3.97 a | 3.46 b | 0.16 | 0.01 | 0.43 | 0.02 |
| Chao | 614 | 582 | 590 | 579 | 611 | 22.59 | 0.41 | 0.72 | 0.64 |
| Items | SDF/IDF | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 1:3 | 1:4 | 1:5 | 1:6 | 1:7 | One-Way | Linear | Quadratic | ||
| Shannon | 2.80 b | 3.28 a,b | 3.41 a,b | 3.67 a | 3.55 a | 0.21 | 0.01 | 0.03 | 0.76 |
| Chao | 512 b | 501 b | 598 a | 570 a,b | 562 a,b | 22.59 | 0.01 | 0.02 | 0.39 |
| Items | SDF/IDF | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 1:5 | 1:6 | 1:7 | 1:8 | 1:9 | One-Way | Linear | Quadratic | ||
| Acetate | 5.25 | 5.00 | 4.60 | 4.65 | 4.32 | 0.54 | 0.21 | 0.01 | 0.45 |
| Propionate | 3.16 | 3.17 | 2.83 | 2.36 | 2.03 | 0.37 | 0.06 | 0.01 | 0.64 |
| Butyrate | 1.67 a | 1.57 a | 1.66 a | 1.19 a,b | 0.87 b | 0.15 | 0.01 | 0.01 | 0.56 |
| Total VFA | 10.08 a | 9.74 a | 9.09 a,b | 8.19 a,b | 7.22 b | 0.82 | 0.03 | 0.01 | 0.35 |
| Items | SDF/IDF | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 1:3 | 1:4 | 1:5 | 1:6 | 1:7 | One-Way | Linear | Quadratic | ||
| Acetate | 12.02 a | 11.25 a | 9.93 a,b | 9.49 a,b | 6.26 b | 1.23 | 0.01 | 0.01 | 0.43 |
| Propionate | 7.53 a,b | 8.99 a | 5.34 b,c | 4.79 b,c | 4.14 c | 0.87 | 0.01 | 0.01 | 0.52 |
| Butyrate | 6.31 a | 4.71 a | 4.28 b | 3.32 b,c | 1.80 c | 0.63 | 0.01 | 0.01 | 0.49 |
| Total VFA | 25.86 a | 24.96 a | 19.54 a,b | 17.60 b | 12.21 c | 1.59 | 0.01 | 0.01 | 0.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, Z.; Zhang, Z.; Wang, F.; Guo, J.; Zhao, X.; Zhao, J. Effects of Dietary Fiber Type on Growth Performance, Serum Parameters and Fecal Microbiota Composition in Weaned and Growing-Finishing Pigs. Animals 2022, 12, 1579. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121579
Lv Z, Zhang Z, Wang F, Guo J, Zhao X, Zhao J. Effects of Dietary Fiber Type on Growth Performance, Serum Parameters and Fecal Microbiota Composition in Weaned and Growing-Finishing Pigs. Animals. 2022; 12(12):1579. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121579
Chicago/Turabian StyleLv, Zhiqian, Zeyu Zhang, Fenglai Wang, Jiyu Guo, Xiaogang Zhao, and Jinbiao Zhao. 2022. "Effects of Dietary Fiber Type on Growth Performance, Serum Parameters and Fecal Microbiota Composition in Weaned and Growing-Finishing Pigs" Animals 12, no. 12: 1579. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12121579