
Comparative Proteomic Analyses of Poorly Motile Swamp Buffalo Spermatozoa Reveal Low Energy Metabolism and Deficiencies in Motility-Related Proteins

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sperm Preparation
2.2. Protein Extraction
2.3. Protein Digestion and TMT Labeling
2.4. Peptide Prefractionation and On-Line LC–MS/MS Analysis
2.5. Data Analysis
2.6. Western Blot Analysis
2.7. Quantitative Reverse Transcription Polymerase Chain Reaction (RT–qPCR)
3. Results
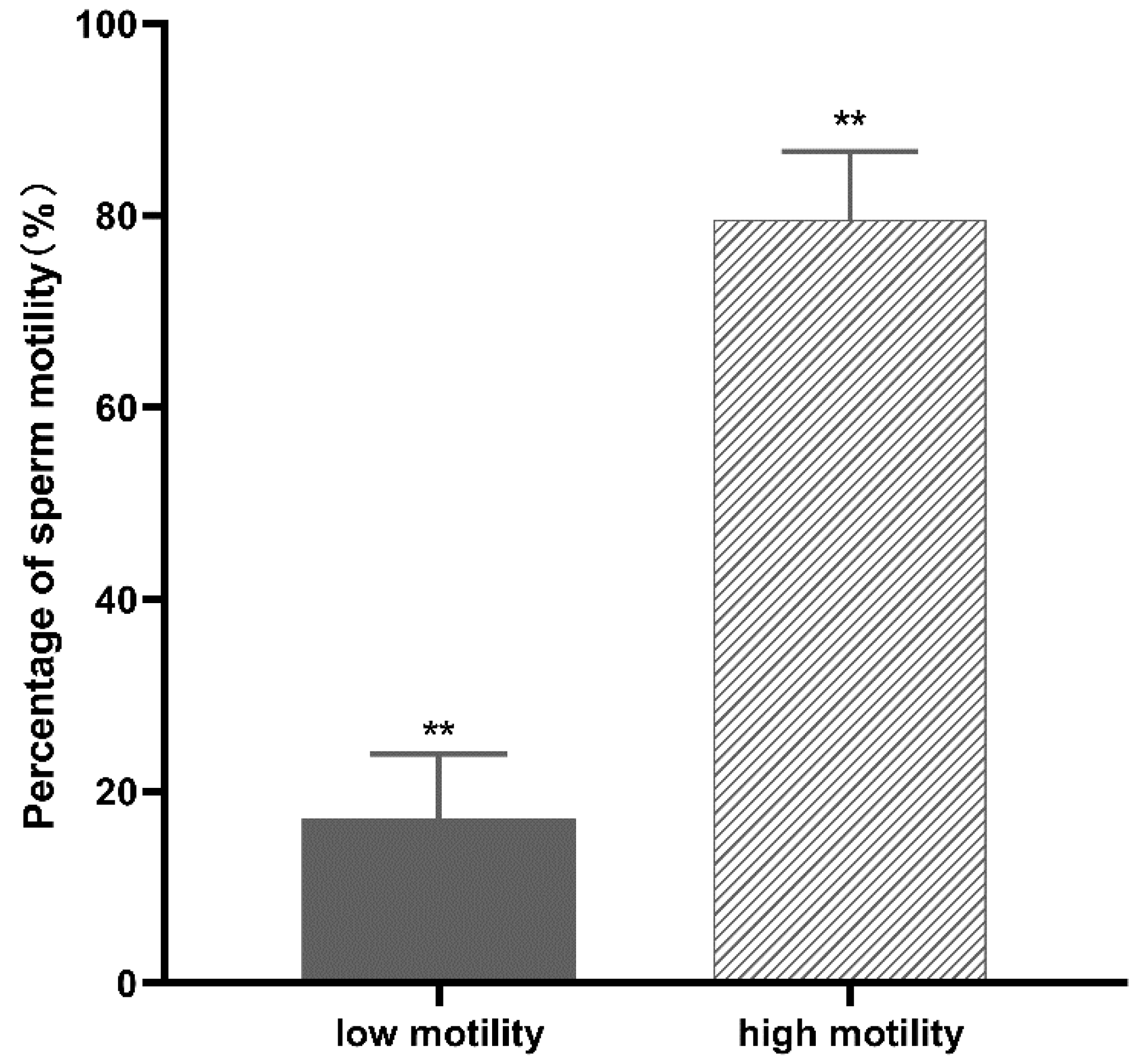
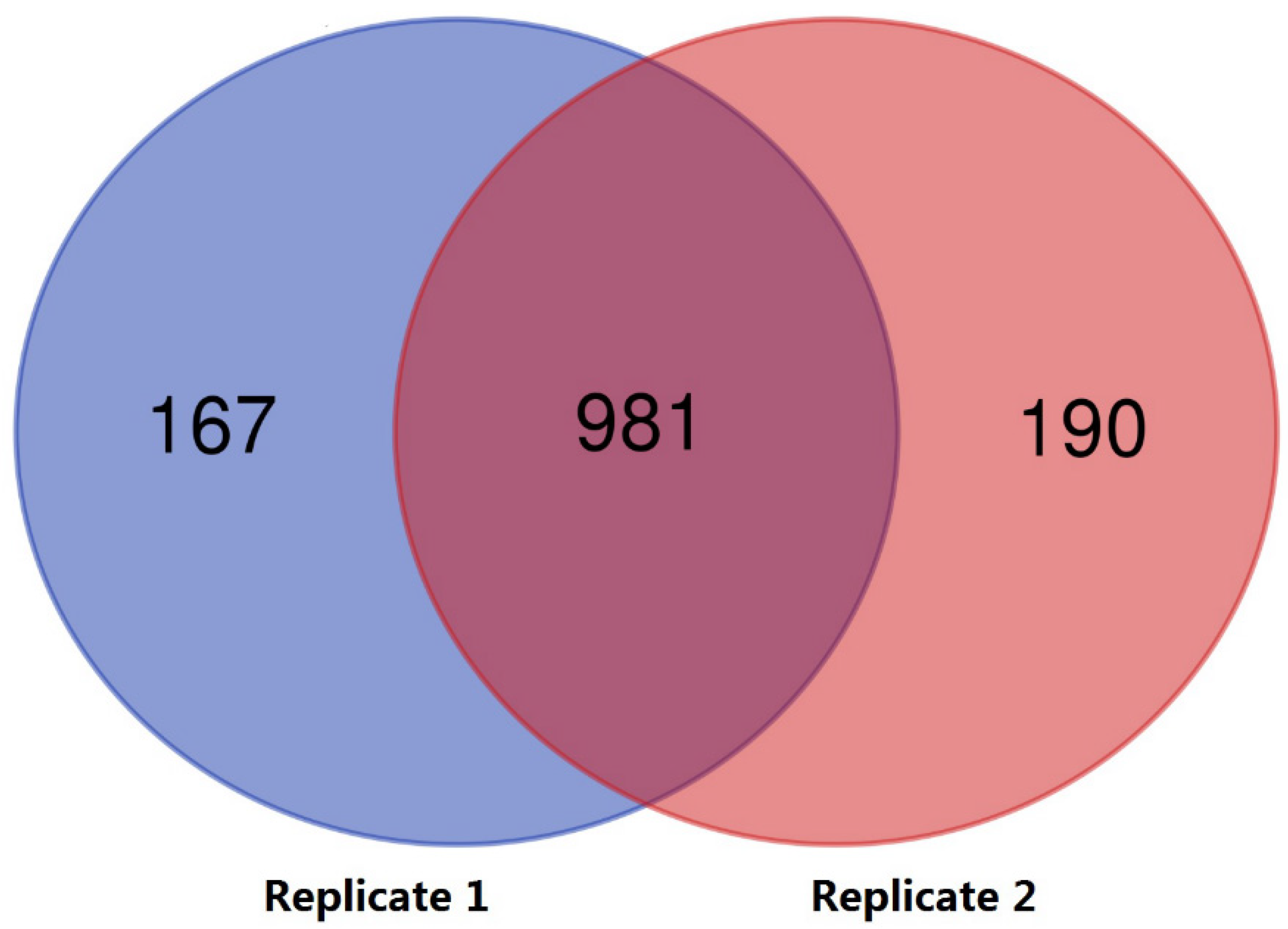
3.1. Proteomic Profile Analysis
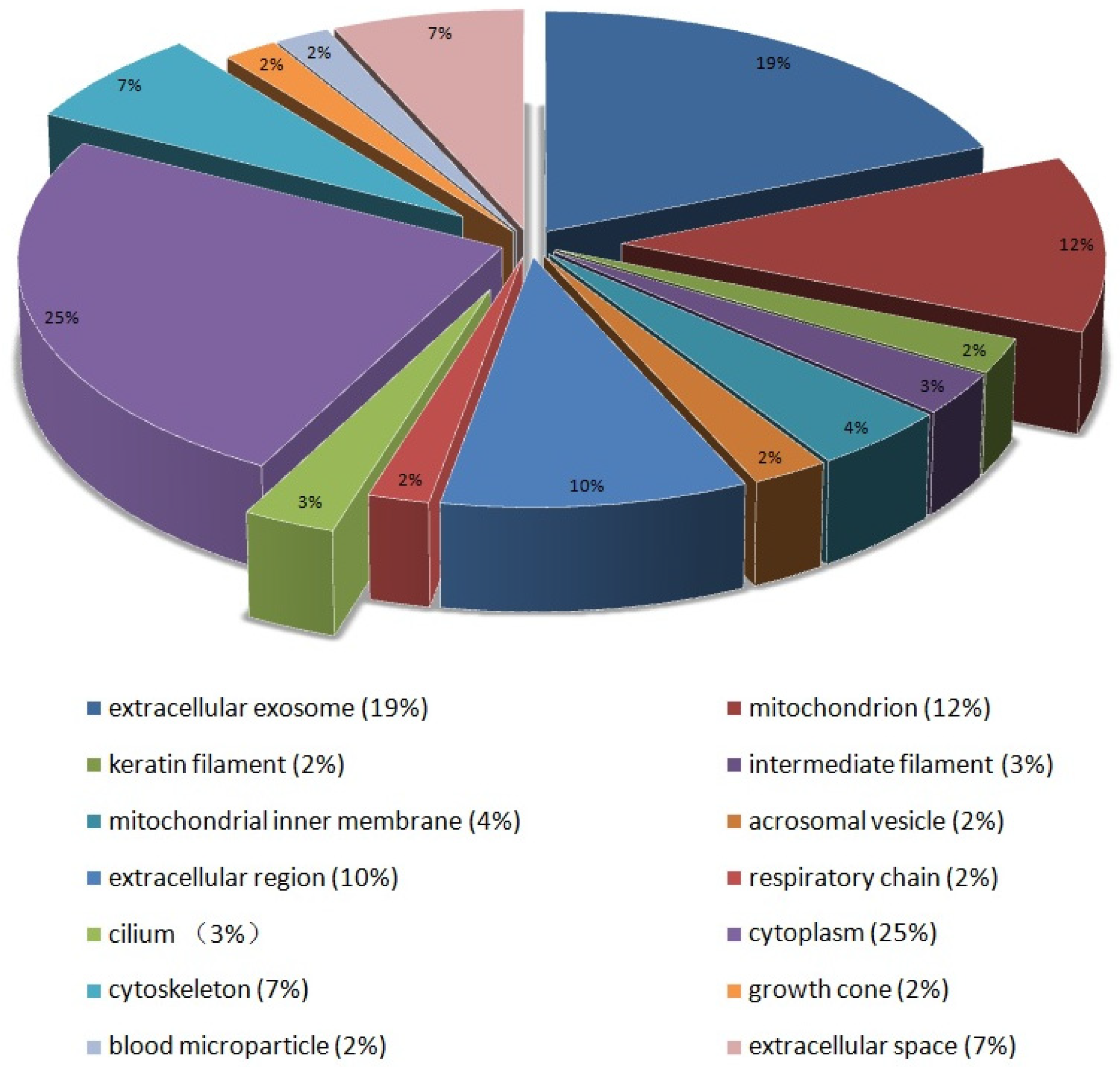
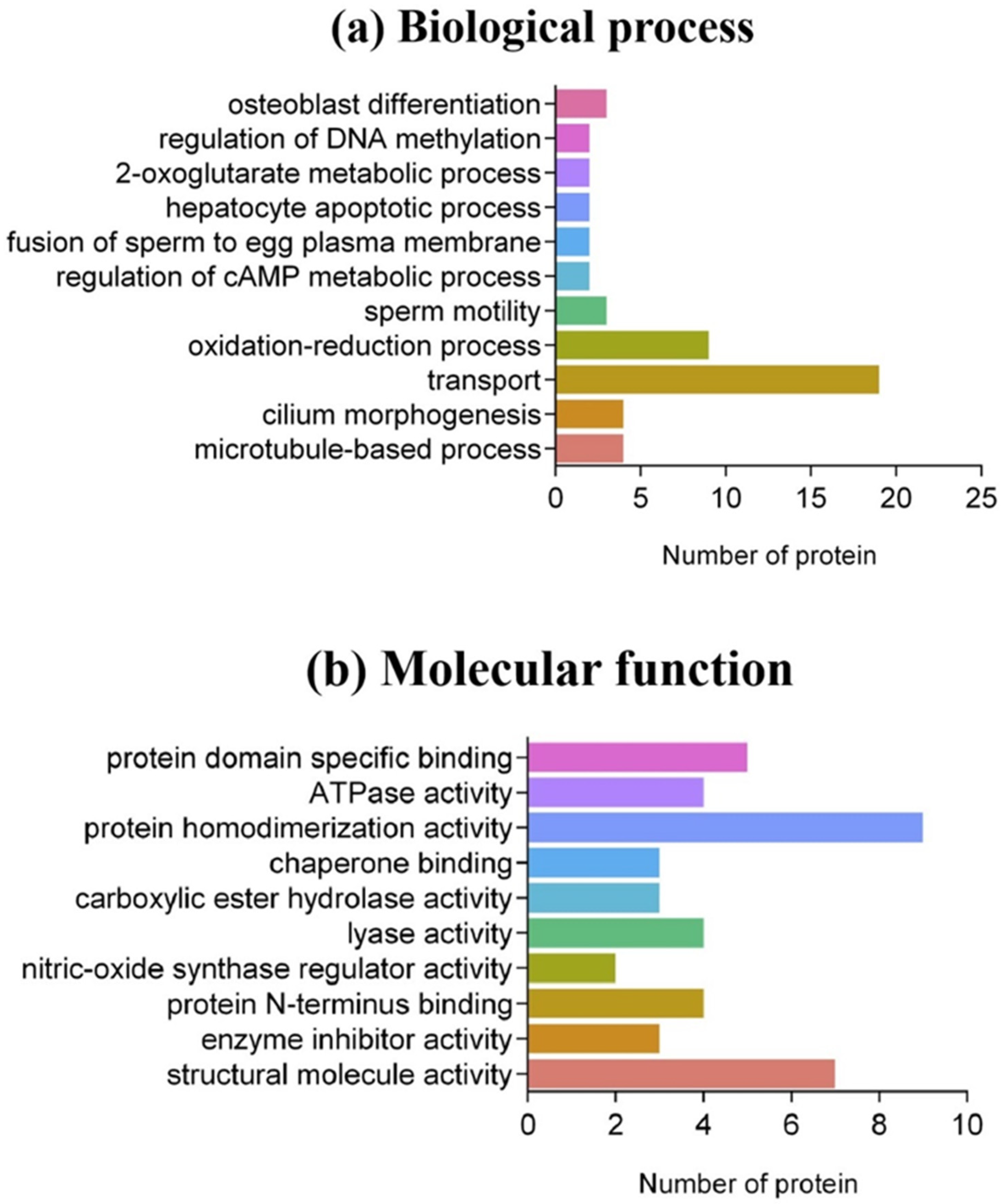
3.2. Bioinformatics Analysis
3.3. KEGG Pathway Analysis of DEPs
3.4. MGI Reproductive Phenotype Analysis
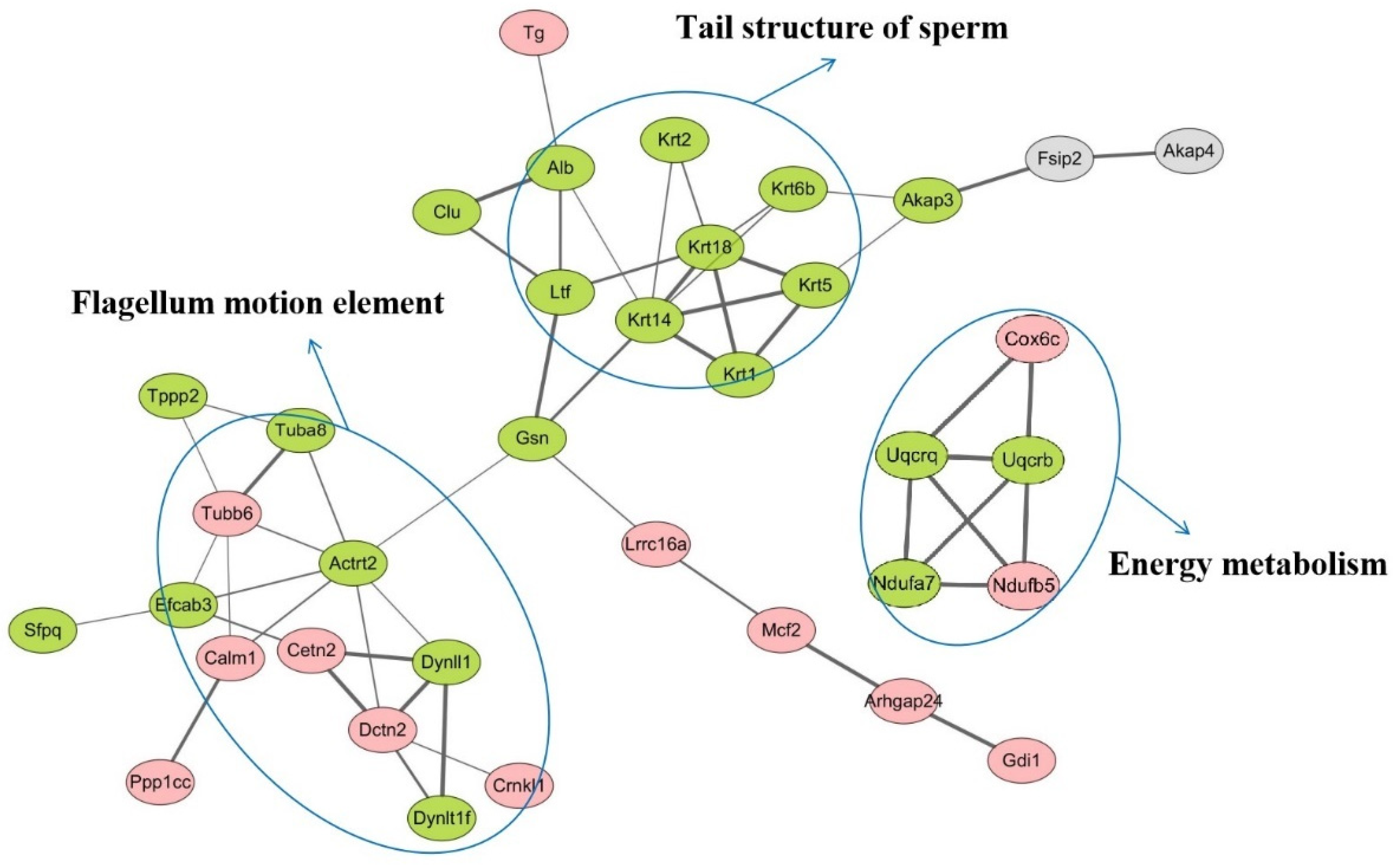
3.5. Protein–Protein Interaction (PPI) Analysis
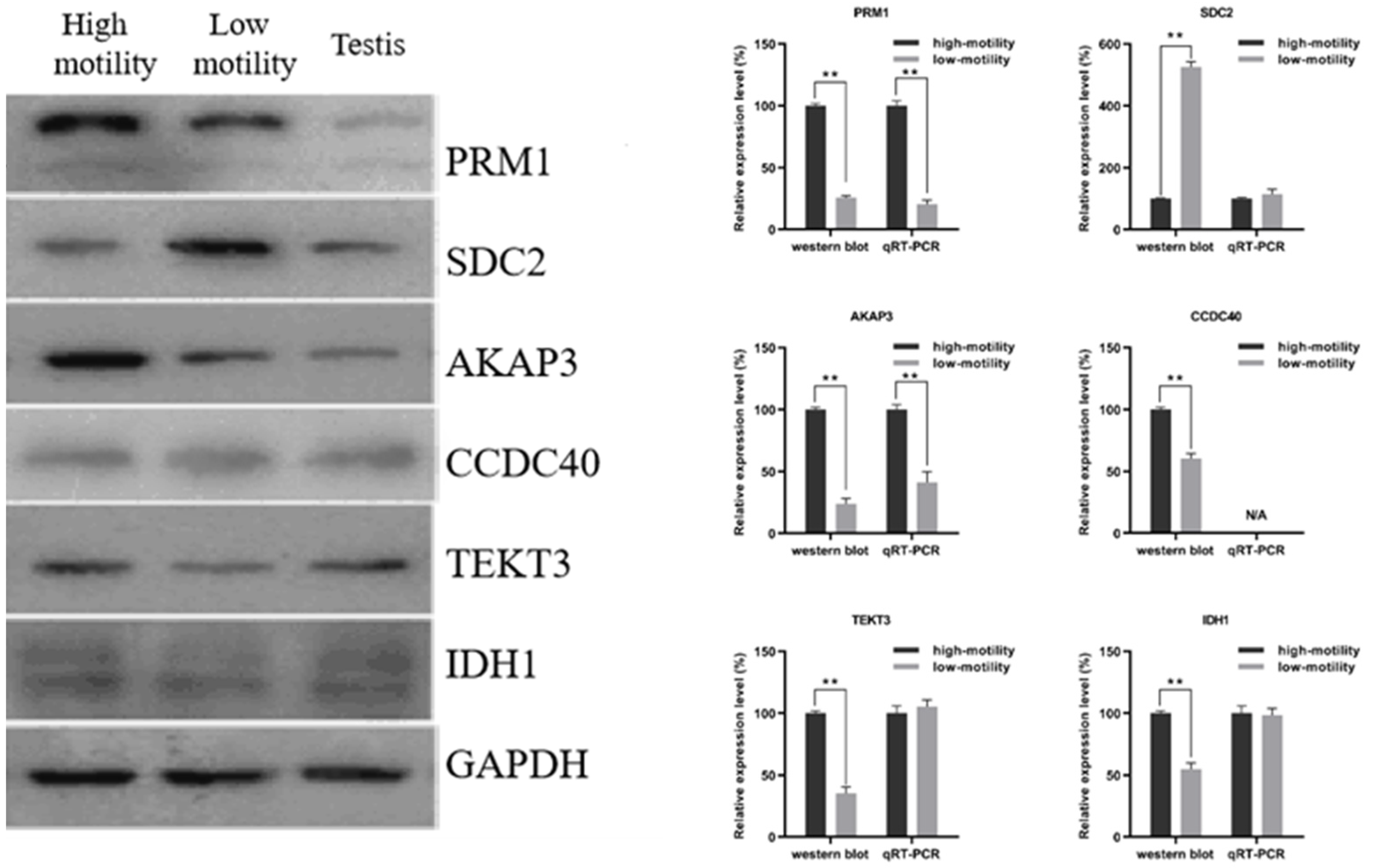
3.6. Western Blot and RT–qPCR Analysis
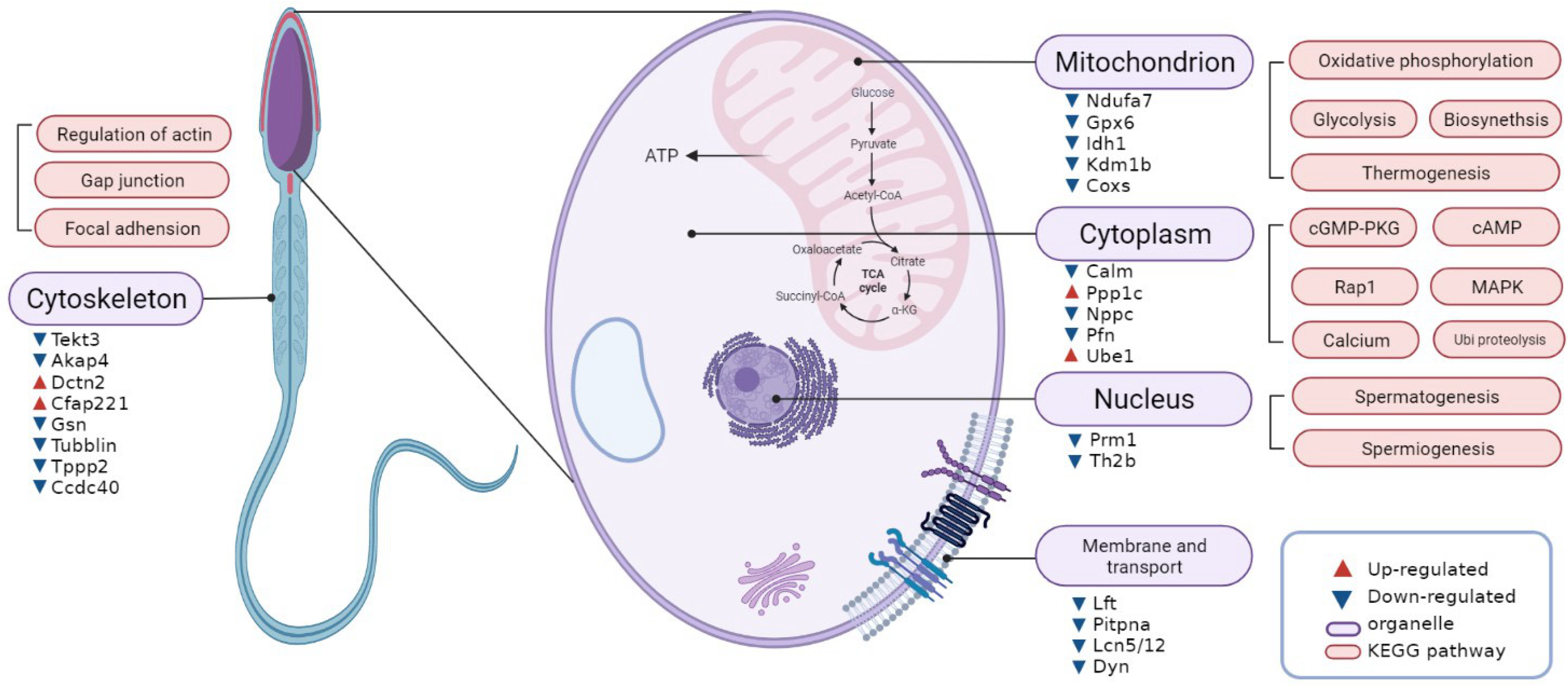
4. Discussion
4.1. Abnormal Nuclear Chromatin of Poorly Motile Buffalo Spermatozoa
4.2. Decreased Mitochondrial Energy Metabolism in Poorly Motile Sperm Samples
4.3. Relationship between Tail Structure Proteins and Motion in Poorly Motile Spermatozoa
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Austin, C.R. Observations on the penetration of the sperm in the mammalian egg. Aust. J. Sci. Res. B 1951, 4, 581–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, M.C. Fertilizing capacity of spermatozoa deposited into the fallopian tubes. Nature 1951, 168, 697–698. [Google Scholar] [CrossRef] [PubMed]
- Moscatelli, N.; Lunetti, P.; Braccia, C.; Armirotti, A.; Pisanello, F.; De Vittorio, M.; Zara, V.; Ferramosca, A. Comparative Proteomic Analysis of Proteins Involved in Bioenergetics Pathways Associated with Human Sperm Motility. Int. J. Mol. Sci. 2019, 20, 3000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piomboni, P.; Focarelli, R.; Stendardi, A.; Ferramosca, A.; Zara, V. The role of mitochondria in energy production for human sperm motility. Int. J. Androl. 2012, 35, 109–124. [Google Scholar] [CrossRef] [Green Version]
- Andras, P.; Yueming, Q.; Chohan, K.R.; Shirley, C.R.; Wendy, A.; Sanjay, B.; Middleton, F.A.; Conkrite, K.L.; Maureen, B.; Nick, G. Transaldolase is essential for maintenance of the mitochondrial transmembrane potential and fertility of spermatozoa. Proc. Natl. Acad. Sci. USA 2006, 103, 14813–14818. [Google Scholar]
- Minghai, S.; Ananya, G.; Cooke, V.G.; Naik, U.P.; Martin-Deleon, P.A. JAM-A is present in mammalian spermatozoa where it is essential for normal motility. Dev. Biol. 2008, 313, 246–255. [Google Scholar]
- Oliva, R.; Martínezheredia, J.; Estanyol, J.M. Proteomics in the Study of the Sperm Cell Composition, Differentiation and Function. Syst. Biol. Reprod. Med. 2008, 54, 23–36. [Google Scholar] [CrossRef]
- Rafael, O.; Sara, D.M.; Josep Maria, E. Sperm cell proteomics. Proteomics 2009, 9, 1004–1017. [Google Scholar]
- Huang, Y.L.; Fu, Q.; Yang, L.; Guan, J.L.; Pan, H.; Chen, F.M.; Lu, K.L.; Zhang, M. Differences between high- and low-motility buffalo sperm identified by comparative proteomics. Reprod. Domest. Anim. 2015, 50, 443–451. [Google Scholar] [CrossRef]
- Zhao, C.; Huo, R.; Wang, F.Q.; Lin, M.; Zhou, Z.M.; Sha, J.H. Identification of several proteins involved in regulation of sperm motility by proteomic analysis. Fertil. Steril. 2007, 87, 436–438. [Google Scholar] [CrossRef]
- Shen, S.; Wang, J.; Liang, J.; He, D. Comparative proteomic study between human normal motility sperm and idiopathic asthenozoospermia. World J. Urol. 2013, 31, 1395–1401. [Google Scholar] [CrossRef] [PubMed]
- Hsin-Chih Albert, C.; Chia-Ling, C.; Hsien-An, P.; Pao-Chi, L.; Pao-Lin, K.; Chao-Chin, H. Protein tyrosine phosphatase non-receptor type 14 is a novel sperm-motility biomarker. J. Assist. Reprod. Genet. 2011, 28, 851–861. [Google Scholar]
- Carballada, R.; Esponda, P. Binding of seminal vesicle proteins to the plasma membrane of rat spermatozoa in vivo and in vitro. Int. J. Androl. 2010, 21, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.A.M.; Souza, C.E.A.; Martins, J.A.M.; Rego, J.P.A.; Oliveira, J.T.A.; Domont, G.; Nogueira, F.C.S.; Moura, A.A. Seminal plasma proteins and their relationship with sperm motility in Santa Ines rams. Small Rumin. Res. 2013, 109, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Ding, Z.; Qu, F.; Guo, W.; Ying, X.; Wu, M.; Zhang, Y. Identification of sperm forward motility-related proteins in human seminal plasma. Mol. Reprod. Dev. 2010, 74, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; He, Y.; Yan, J.; Zhao, Q.; Di, C.; Zhang, H. Comparative proteomics reveals the underlying toxicological mechanism of low sperm motility induced by iron ion radiation in mice. Reprod. Toxicol. 2016, 65, 148–158. [Google Scholar] [CrossRef]
- Hou, Z.; Fu, Q.; Huang, Y.; Zhang, P.; Chen, F.; Li, M.; Xu, Z.; Yao, S.; Chen, D.; Zhang, M. Erratum to Comparative proteomic identification of capacitation and noncapacitation swamp buffalo spermatozoa. Theriogenology 2019, 128, R1. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.L.; Zhang, P.F.; Fu, Q.; He, W.T.; Xiao, K.; Zhang, M. Novel targets identified by integrated proteomic and phosphoproteomic analysis in spermatogenesis of swamp buffalo (Bubalus bubalis). Sci. Rep. 2020, 10, 15659. [Google Scholar] [CrossRef]
- Huang, Y.L.; Zhang, P.F.; Hou, Z.; Fu, Q.; Li, M.X.; Huang, D.L.; Deng, T.X.; Lu, Y.Q.; Liang, X.W.; Zhang, M. Ubiquitome analysis reveals the involvement of lysine ubiquitination in the spermatogenesis process of adult buffalo (Bubalus bubalis) testis. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef]
- Zhang, P.F.; Huang, Y.L.; Fu, Q.; He, W.T.; Xiao, K.; Zhang, M. Integrated analysis of phosphoproteome and ubiquitylome in epididymal sperm of buffalo (Bubalus bubalis). Mol. Reprod. Dev. 2021, 88, 15–33. [Google Scholar] [CrossRef]
- Almadaly, E.; El-Kon, I.; Heleil, B.; Fattouh, E.S.; Mukoujima, K.; Ueda, T.; Hoshino, Y.; Takasu, M.; Murase, T. Methodological factors affecting the results of staining frozen-thawed fertile and subfertile Japanese Black bull spermatozoa for acrosomal status. Anim. Reprod. Sci. 2012, 136, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Pan, L.; Huang, D.; Wang, Z.; Hou, Z.; Zhang, M. Proteomic profiles of buffalo spermatozoa and seminal plasma. Theriogenology 2019, 134, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Gygi, S.P. Proteomics: The move to mixtures. J. Mass Spectrom. 2001, 36, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xu, H.; Aronow, B.J.; Jegga, A.G. Improved human disease candidate gene prioritization using mouse phenotype. BMC Bioinform. 2007, 8, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Tolić, N.; Masselon, C.; Pašatolić, L.; Ii, D.G.C.; Lipton, M.S.; Anderson, G.A.; Smith, R.D. Nanoscale proteomics. Anal. Bioanal. Chem. 2004, 378, 1037–1045. [Google Scholar] [CrossRef]
- Nice, E.C.; Rothacker, J.; Weinstock, J.; Lim, L.; Catimel, B. Use of multidimensional separation protocols for the purification of trace components in complex biological samples for proteomics analysis. J. Chromatogr. A 2007, 1168, 190–210. [Google Scholar] [CrossRef]
- Lower, D.R.; Cropcho, L.; Rosendorff, A. Comparison of CEDIA FK506 Assay With HPLC/MS/MS in a Large Cohort of Pediatric Patients. Am. J. Clin. Pathol. 2013, 139, 788–792. [Google Scholar] [CrossRef] [Green Version]
- De Godoy, L.M.F.; Olsen, J.V.; Jürgen, C.; Nielsen, M.L.; Hubner, N.C.; Florian, F.H.; Walther, T.C.; Matthias, M. Comprehensive mass-spectrometry-based proteome quantification of haploid versus diploid yeast. Nature 2008, 455, 1251–1254. [Google Scholar] [CrossRef]
- Peddinti, D.; Memili, E.; Burgess, S.C. Proteomics-Based Systems Biology Modeling of Bovine Germinal Vesicle Stage Oocyte and Cumulus Cell Interaction. PLoS ONE 2010, 5, e11240. [Google Scholar] [CrossRef] [Green Version]
- Kistler, W.S.; Henriksén, K.; Mali, P.; Parvinen, M. Sequential expression of nucleoproteins during rat spermiogenesis. Exp. Cell Res. 1996, 225, 374–381. [Google Scholar] [CrossRef]
- Cho, C.; Willis, W.D.; Goulding, E.H.; Jung-Ha, H.; Choi, Y.C.; Hecht, N.B.; Eddy, E.M. Haploinsufficiency of protamine-1 or -2 causes infertility in mice. Nat. Genet. 2001, 28, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Tüttelmann, F.; Krenková, P.; Römer, S.; Nestorovic, A.R.; Ljujic, M.; Stambergová, A.; Macek, M.; Macek, M.; Nieschlag, E.; Gromoll, J. A common haplotype of protamine 1 and 2 genes is associated with higher sperm counts. Int. J. Androl. 2010, 33, e240–e248. [Google Scholar] [CrossRef] [PubMed]
- D’Occhio, M.J.; Hengstberger, K.J.; Johnston, S.D. Biology of sperm chromatin structure and relationship to male fertility and embryonic survival. Anim. Reprod. Sci. 2007, 101, 1–17. [Google Scholar] [CrossRef]
- Ford, W.C. Glycolysis and sperm motility: Does a spoonful of sugar help the flagellum go round? Hum. Reprod. Update 2006, 12, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Barazzoni, R.; Short, K.R.; Nair, K.S. Effects of aging on mitochondrial DNA copy number and cytochrome c oxidase gene expression in rat skeletal muscle, liver, and heart. J. Biol. Chem. 2000, 275, 3343–3347. [Google Scholar] [CrossRef] [Green Version]
- Huttemann, M.; Jaradat, S.; Grossman, L.I. Cytochrome c oxidase of mammals contains a testes-specific isoform of subunit VIb--the counterpart to testes-specific cytochrome c? Mol. Reprod. Dev. 2003, 66, 8–16. [Google Scholar] [CrossRef]
- Coenen, M.J.; van den Heuvel, L.P.; Smeitink, J.A. Mitochondrial oxidative phosphorylation system assembly in man: Recent achievements. Curr. Opin. Neurol. 2001, 14, 777–781. [Google Scholar] [CrossRef]
- Skowron, B.; Juszczak, K.; Thor, P.J. Drug induced man infertility. Folia Med. Cracov. 2011, 51, 99–106. [Google Scholar]
- Suarez, S.S. How Do Sperm Get to the Egg? Bioengineering Expertise Needed! Exp. Mech. 2010, 50, 1267–1274. [Google Scholar] [CrossRef]
- Larsson, M.; Norrander, J.; Gräslund, S.; Brundell, E.; Linck, R.; Ståhl, S.; Höög, C. The spatial and temporal expression of Tekt1, a mouse tektin C homologue, during spermatogenesis suggest that it is involved in the development of the sperm tail basal body and axoneme. Eur. J. Cell Biol. 2000, 79, 718–725. [Google Scholar] [CrossRef]
- Murayama, E.; Yamamoto, E.; Kaneko, T.; Shibata, Y.; Inai, T.; Iida, H. Tektin5, a new Tektin family member, is a component of the middle piece of flagella in rat spermatozoa. Mol. Reprod. Dev. 2010, 75, 650–658. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Ijiri, T.W.; Huang, A.P.; Gerton, G.L. Characterization of a novel tektin member, TEKT5, in mouse sperm. J. Androl. 2011, 32, 55–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oiki, S.; Hiyama, E.; Gotoh, T.; Iida, H. Localization of Tektin 1 at both acrosome and flagella of mouse and bull spermatozoa. Zool. Sci. 2014, 31, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Gerton, G.L.; Moss, S.B. Proteomic profiling of accessory structures from the mouse sperm flagellum. Mol. Cell. Proteom. MCP 2006, 5, 801. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Lin, Y.N.; Julio, E.; Demayo, F.J.; Matzuk, M.M. Tektin 3 is Required for Progressive Sperm Motility in Mice. Mol. Reprod. Dev. 2010, 76, 453–459. [Google Scholar] [CrossRef] [Green Version]
- Takiguchi, H.; Murayama, E.; Kaneko, T.; Kurio, H.; Toshimori, K.; Iida, H. Characterization and subcellular localization of Tektin 3 in rat spermatozoa. Mol. Reprod. Dev. 2011, 78, 611–620. [Google Scholar] [CrossRef]
- Adachi, J.; Tate, S.; Miyake, M.; Harayama, H. Effects of protein phosphatase inhibitor calyculin a on the postacrosomal protein serine/threonine phosphorylation state and acrosome reaction in boar spermatozoa incubated with a cAMP analog. J. Reprod. Dev. 2008, 54, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Anita, B.H.; Zohn, I.E.; Noriko, O.; Andrew, P.; Kari Baker, L.; Jessica, S.B.; Jason, M.S.; Loges, N.T.; Heike, O.; Karsten, H. The coiled-coil domain containing protein CCDC40 is essential for motile cilia function and left-right axis formation. Nat. Genet. 2011, 43, 79–84. [Google Scholar]
- Dinu, A.; Anita, B.H.; Zariwala, M.A.; Miriam, S.; Alexandros, O.; Mitra, F.; Robert, W.; Theresa, T.C.; Ann, D.; Claire, J. Mutations in CCDC39 and CCDC40 are the major cause of primary ciliary dyskinesia with axonemal disorganization and absent inner dynein arms. Hum. Mutat. 2013, 34, 462–472. [Google Scholar]
- Sylvain, B.; Marie, L.; Bruno, C.; Philippe, D.; Guy, M.; Esther, K.; Florence, D.; Ludovic, J.; Marine, C.; Alexandra, R. Delineation of CCDC39/CCDC40 mutation spectrum and associated phenotypes in primary ciliary dyskinesia. J. Med. Genet. 2012, 49, 410–416. [Google Scholar]
- Sui, W.; Hou, X.; Che, W.; Ou, M.; Sun, G.; Huang, S.; Liu, F.; Chen, P.; Wei, X.; Dai, Y. CCDC40 mutation as a cause of primary ciliary dyskinesia: A case report and review of literature. Clin. Respir. J. 2016, 10, 614–621. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KEGG ID | Pathway Name | Protein Counts | Protein Names |
|---|---|---|---|
| ko01100 | Metabolic pathways | 11 | IDH1; QCR7; QCR8; ALD0; E4.1.3.4; ATPeF0F6; COX6C; NDUFA7; NDUFB5; ECHS1; ASRGL1 |
| ko04714 | Thermogenesis | 8 | QCR7; QCR8; ATPeF0F6; COX6C; NDUFA7; NDUFB5; NDUFAF5; COA1 |
| ko00190 | Oxidative phosphorylation | 6 | QCR7; QCR8; ATPeF0F6; COX6C; NDUFA7; NDUFB5; |
| ko01110 | Biosynthesis of secondary metabolites | 4 | IDH1; ALDO; ECHS1; ASRGL1 |
| ko04022 | cGMP-PKG signaling pathway | 3 | CALM; PPP1C; NPPC |
| ko04979 | Cholesterol metabolism | 2 | VAPB; NPC2 |
| ko04218 | Cellular senescence | 2 | CALM; PPP1C |
| ko04120 | Ubiquitin mediated proteolysis | 2 | UBE1; ELOC |
| ko04024 | cAMP signaling pathway | 2 | CALM; PPP1C |
| ko01230 | Biosynthesis of amino acids | 2 | IDH1; ALDO |
| ko04015 | Rap1 signaling pathway | 2 | CALM; PFN |
| ko04810 | Regulation of actin cytoskeleton | 2 | PFN; PPP1C |
| ko04540 | Gap junction | 2 | TUBA; TUBB |
| ko04510 | Focal adhesion | 1 | PPP1C |
| ko04013 | MAPK signaling pathway | 1 | PFN |
| ko01212 | Fatty acid metabolism | 1 | ECHS1 |
| ko00010 | Glycolysis/Gluconeogenesis | 1 | ALDO |
| ko04020 | Calcium signaling pathway | 1 | CALM |
| ko00072 | Synthesis and degradation of ketone bodies | 1 | E4.1.3.4 |
| Mammalian Phenotype ID | Mammalian Phenotype Term | Gene Names |
|---|---|---|
| MP: 0001925 | male infertility | Akap4, Ppp1cc, Ddx25, Izumo1, Adam1a, S1c9c1, Odf1 |
| MP: 0002675 | asthenozoospermia | Ppp1cc, Tekt3, Adam1a, S1c9c1, Odf1 |
| MP: 0003984 | Embryonic growth retardation | Lig3, C1qbp, Rbpj |
| MP: 0005389 | Abnormal Reproductive system phenotype | Rdh11, Akap4, Ppp1cc, Cd46, Tekt3, Mcf2, Nppc, Odf1 |
| MP: 0005410 | Abnormal fertilization | Cd46, Adam1a |
| MP: 0009238 | Coiled sperm flagellum | Akap4, Odf1, Dynll1 |
| MP: 0009832 | Abnormal sperm mitochondrial sheath morphology | Akap4, Ppp1cc, Odf1 |
| MP: 0009836 | Abnormal sperm principal piece morphology | Akap4, Ppp1cc |
| MP: 0011092 | Embryonic lethality | Zfp335, Slc3a2, Rbpj, C1qbp, Glrx3, Gsn, Lig3, Hmgc1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, R.; Huang, X.; Sun, Q.; Hou, Z.; Yang, W.; Zhang, J.; Zhang, P.; Huang, L.; Lu, Y.; Fu, Q. Comparative Proteomic Analyses of Poorly Motile Swamp Buffalo Spermatozoa Reveal Low Energy Metabolism and Deficiencies in Motility-Related Proteins. Animals 2022, 12, 1706. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12131706
Liu R, Huang X, Sun Q, Hou Z, Yang W, Zhang J, Zhang P, Huang L, Lu Y, Fu Q. Comparative Proteomic Analyses of Poorly Motile Swamp Buffalo Spermatozoa Reveal Low Energy Metabolism and Deficiencies in Motility-Related Proteins. Animals. 2022; 12(13):1706. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12131706
Chicago/Turabian StyleLiu, Runfeng, Xingchen Huang, Qinqiang Sun, Zhen Hou, Weihan Yang, Junjun Zhang, Pengfei Zhang, Liangfeng Huang, Yangqing Lu, and Qiang Fu. 2022. "Comparative Proteomic Analyses of Poorly Motile Swamp Buffalo Spermatozoa Reveal Low Energy Metabolism and Deficiencies in Motility-Related Proteins" Animals 12, no. 13: 1706. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12131706