
Specific Gene Duplication and Loss of Cytochrome P450 in Families 1-3 in Carnivora (Mammalia, Laurasiatheria)
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Retrieval for CYP Phylogenetic Analysis
2.2. Synteny Analysis of CYP Genes
3. Results
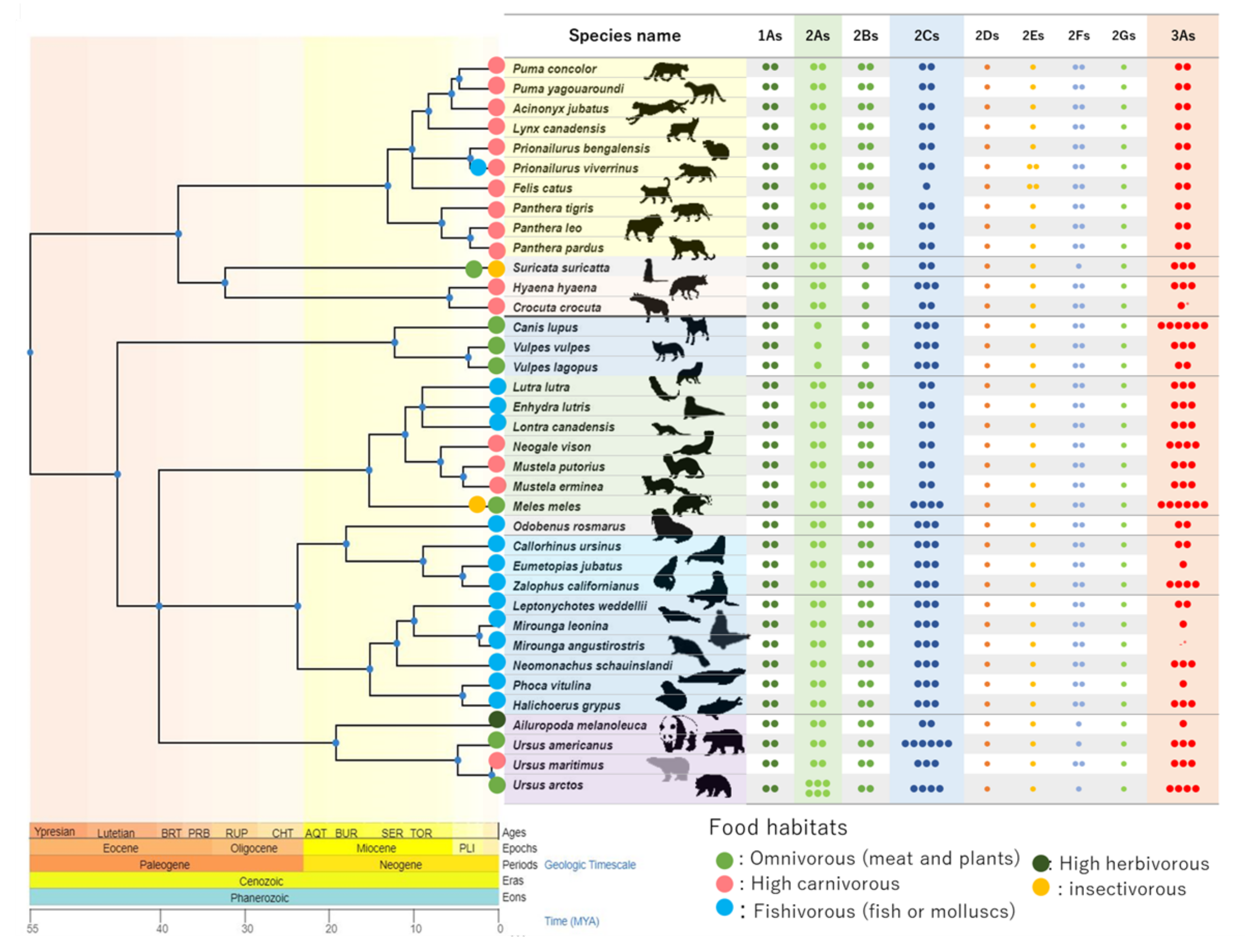
3.1. CYP Number Counts and Isoforms in CYP1As and 2ABGFSs Clusters
3.2. CYP Isoforms in CYP2Cs and CYP 2CEs Clusters
3.3. Synteny Analysis of the CYP3As Cluster
3.4. Phylogeny of CYP2Cs in Carnivorans
3.5. Phylogeny of CYP3As in Carnivorans
4. Discussion
4.1. CYPs Duplication and Loss in Mammals and Relationships with Foraging Habits
4.2. CYP2Cs in Carnivora
4.3. CYP2C23s Orthologous Genes in Carnivora
4.4. CYP3As in Mammals
4.5. Other CYP Isoforms
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelson, D.R.; Kamataki, T.; Waxman, D.J.; Guengerich, F.P.; Estabrook, R.W.; Feyereisen, R.; Gonzalez, F.J.; Coon, M.J.; Gunsalus, I.C.; Gotoh, O.; et al. The P450 Superfamily: Update on New Sequences, Gene Mapping, Accession Numbers, Early Trivial Names of Enzymes, and Nomenclature. DNA Cell Biol. 2009, 12, 1–51. [Google Scholar] [CrossRef] [PubMed]
- Zanger, U.M.; Schwab, M. Cytochrome P450 Enzymes in Drug Metabolism: Regulation of Gene Expression, Enzyme Activities, and Impact of Genetic Variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef] [PubMed]
- Verma, H.; Bahia, M.S.; Choudhary, S.; Singh, P.K.; Silakari, O. Drug Metabolizing Enzymes-Associated Chemo Resistance and Strategies to Overcome It. Drug Metab. Rev. 2019, 51, 196–223. [Google Scholar] [CrossRef] [PubMed]
- Ramaiahgari, S.C.; Waidyanatha, S.; Dixon, D.; DeVito, M.J.; Paules, R.S.; Ferguson, S.S. From the Cover: Three-Dimensional (3D) HepaRG Spheroid Model With Physiologically Relevant Xenobiotic Metabolism Competence and Hepatocyte Functionality for Liver Toxicity Screening. Toxicol. Sci. 2017, 159, 124–136. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.R.; Koymans, L.; Kamataki, T.; Stegeman, J.J.; Feyereisen, R.; Waxman, D.J.; Waterman, M.R.; Gotoh, O.; Coon, M.J.; Estabrook, R.W.; et al. P450 Superfamily: Update on New Sequences, Gene Mapping, Accession Numbers and Nomenclature. Pharmacogenetics 1996, 6, 1–42. [Google Scholar] [CrossRef]
- Zhao, M.; Ma, J.; Li, M.; Zhang, Y.; Jiang, B.; Zhao, X.; Huai, C.; Shen, L.; Zhang, N.; He, L.; et al. Cytochrome P450 Enzymes and Drug Metabolism in Humans. Int. J. Mol. Sci. 2021, 22, 12808. [Google Scholar] [CrossRef]
- Sabbagh, A.; Marin, J.; Veyssière, C.; Lecompte, E.; Boukouvala, S.; Poloni, E.S.; Darlu, P.; Crouau-Roy, B. Rapid Birth-and-Death Evolution of the Xenobiotic Metabolizing NAT Gene Family in Vertebrates with Evidence of Adaptive Selection. BMC Evol. Biol. 2013, 13, 62. [Google Scholar] [CrossRef] [Green Version]
- Kawashima, A.; Satta, Y. Substrate-Dependent Evolution of Cytochrome P450: Rapid Turnover of the Detoxification-Type and Conservation of the Biosynthesis-Type. PLoS ONE 2014, 9, e100059. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.H. Rapid Birth–Death Evolution Specific to Xenobiotic Cytochrome P450 Genes in Vertebrates. PLOS Genet. 2007, 3, e67. [Google Scholar] [CrossRef] [Green Version]
- Bock, K.W. Vertebrate UDP-Glucuronosyltransferases: Functional and Evolutionary Aspects. Biochem. Pharmacol. 2003, 66, 691–696. [Google Scholar] [CrossRef]
- Johnson, R.N.; O’Meally, D.; Chen, Z.; Etherington, G.J.; Ho, S.Y.W.; Nash, W.J.; Grueber, C.E.; Cheng, Y.; Whittington, C.M.; Dennison, S.; et al. Adaptation and Conservation Insights from the Koala Genome. Nat. Genet. 2018, 50, 1102–1111. [Google Scholar] [CrossRef] [PubMed]
- Greenhalgh, R.; Holding, M.L.; Orr, T.J.; Henderson, J.B.; Parchman, T.L.; Matocq, M.D.; Shapiro, M.D.; Dearing, M.D. Trio-Binned Genomes of the Woodrats Neotoma Bryanti and Neotoma Lepida Reveal Novel Gene Islands and Rapid Copy Number Evolution of Xenobiotic Metabolizing Genes. Mol. Ecol. Resour. 2022, 22, 2713–2731. [Google Scholar] [CrossRef]
- Watanabe, K.P.; Kawai, Y.K.; Ikenaka, Y.; Kawata, M.; Ikushiro, S.I.; Sakaki, T.; Ishizuka, M. Avian Cytochrome P450 (CYP) 1-3 Family Genes: Isoforms, Evolutionary Relationships, and MRNA Expression in Chicken Liver. PLoS ONE 2013, 8, e75689. [Google Scholar] [CrossRef] [Green Version]
- Radominska-Pandya, A.; Bratton, S.; Little, J. A Historical Overview of the Heterologous Expression of Mammalian UDP-Glucuronosyltransferase Isoforms Over the Past Twenty Years. Curr. Drug Metab. 2005, 6, 141–160. [Google Scholar] [CrossRef] [PubMed]
- Feng, P.; Liu, Z. Complex Gene Expansion of the CYP2D Gene Subfamily. Ecol. Evol. 2018, 8, 11022–11030. [Google Scholar] [CrossRef] [Green Version]
- Hassanin, A.; Veron, G.; Ropiquet, A.; van Vuuren, B.J.; Lécu, A.; Goodman, S.M.; Haider, J.; Nguyen, T.T. Evolutionary History of Carnivora (Mammalia, Laurasiatheria) Inferred from Mitochondrial Genomes. PLoS ONE 2021, 16, e0240770. [Google Scholar] [CrossRef]
- Werdelin, L.; Dehghani, R. Carnivora; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Kondo, T.; Ikenaka, Y.; Nakayama, S.M.M.; Kawai, Y.K.; Mizukawa, H.; Mitani, Y.; Nomiyama, K.; Tanabe, S.; Ishizuka, M. Uridine Diphosphate-Glucuronosyltransferase (UGT) 2B Subfamily Interspecies Differences in Carnivores. Toxicol. Sci. 2017, 158, 90–100. [Google Scholar] [CrossRef] [Green Version]
- Kakehi, M.; Ikenaka, Y.; Nakayama, S.M.M.; Kawai, Y.K.; Watanabe, K.P.; Mizukawa, H.; Nomiyama, K.; Tanabe, S.; Ishizuka, M. Uridine Diphosphate-Glucuronosyltransferase (UGT) Xenobiotic Metabolizing Activity and Genetic Evolution in Pinniped Species. Toxicol. Sci. 2015, 147, 360–369. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, B.; Reed, J.M.; Starks, P.T.; Kaufman, G.E.; Goldstone, J.V.; Roelke, M.E.; O’Brien, S.J.; Koepfli, K.P.; Frank, L.G.; Court, M.H. Evolution of a Major Drug Metabolizing Enzyme Defect in the Domestic Cat and Other Felidae: Phylogenetic Timing and the Role of Hypercarnivory. PLoS ONE 2011, 6, 221–237. [Google Scholar] [CrossRef] [Green Version]
- Genereux, D.P.; Serres, A.; Armstrong, J.; Johnson, J.; Marinescu, V.D.; Murén, E.; Juan, D.; Bejerano, G.; Casewell, N.R.; Chemnick, L.G.; et al. A Comparative Genomics Multitool for Scientific Discovery and Conservation. Nature 2020, 587, 240–245. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Suleski, M.; Blair Hedges, S. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef] [PubMed]
- Buels, R.; Yao, E.; Diesh, C.M.; Hayes, R.D.; Munoz-Torres, M.; Helt, G.; Goodstein, D.M.; Elsik, C.G.; Lewis, S.E.; Stein, L.; et al. JBrowse: A Dynamic Web Platform for Genome Visualization and Analysis. Genome Biol. 2016, 17, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lan, T.; Fang, D.; Li, H.; Sahu, S.K.; Wang, Q.; Yuan, H.; Zhu, Y.; Yang, Z.; Zhang, L.; Yang, S.; et al. Chromosome-Scale Genome of Masked Palm Civet (Paguma Larvata) Shows Genomic Signatures of Its Biological Characteristics and Evolution. Front. Genet. 2022, 12, 2699. [Google Scholar] [CrossRef]
- Uno, Y.; Jikuya, S.; Noda, Y.; Murayama, N.; Yamazaki, H. A Comprehensive Investigation of Dog Cytochromes P450 3A (CYP3A) Reveals A Functional Role of Newly Identified CYP3A98 in Small Intestine. Drug Metab. Dispos. 2022, 50, DMD-AR-2021-000749. [Google Scholar] [CrossRef]
- Honda, K.; Komatsu, T.; Koyama, F.; Kubota, A.; Kawakami, K.; Asakura, H.; Uno, Y.; Kitazawa, T.; Hiraga, T.; Teraoka, H. Expression of Two Novel Cytochrome P450 3A131 and 3A132 in Liver and Small Intestine of Domestic Cats. J. Vet. Med. Sci. 2011, 73, 1489–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haley, S.L.; Lamb, J.G.; Franklin, M.R.; Constance, J.E.; Denise Dearing, M. Xenobiotic Metabolism of Plant Secondary Compounds in Juniper (Juniperus Monosperma) by Specialist and Generalist Woodrat Herbivores, Genus Neotoma. Comp. Biochem. Physiol. Part. C Toxicol. Pharmacol. 2007, 146, 552–560. [Google Scholar] [CrossRef]
- Kitanovic, S.; Orr, T.J.; Spalink, D.; Cocke, G.B.; Schramm, K.; Wilderman, P.R.; Halpert, J.R.; Dearing, M.D. Role of Cytochrome P450 2B Sequence Variation and Gene Copy Number in Facilitating Dietary Specialization in Mammalian Herbivores. Mol. Ecol. 2018, 27, 723–736. [Google Scholar] [CrossRef]
- Mangipane, L.S.; Belant, J.L.; Lafferty, D.J.R.; Gustine, D.D.; Hiller, T.L.; Colvin, M.E.; Mangipane, B.A.; Hilderbrand, G.V. Dietary Plasticity in a Nutrient-Rich System Does Not Influence Brown Bear (Ursus Arctos) Body Condition or Denning. Polar Biol. 2018, 41, 763–772. [Google Scholar] [CrossRef]
- Costello, C.M.; Cain, S.L.; Pils, S.; Frattaroli, L.; Haroldson, M.A.; Van Manen, F.T. Diet and Macronutrient Optimization in Wild Ursids: A Comparison of Grizzly Bears with Sympatric and Allopatric Black Bears. PLoS ONE 2016, 11, e0153702. [Google Scholar] [CrossRef]
- Kruuk, H.; Parish, T. Feeding Specialization of the European Badger (Meles Meles) in Scotland. J. Anim. Ecol. 1981, 50, 773–788. [Google Scholar] [CrossRef]
- Balestrieri, A.; Remonti, L.; Saino, N.; Raubenheimer, D. The ‘Omnivorous Badger Dilemma’: Towards an Integration of Nutrition with the Dietary Niche in Wild Mammals. Mamm. Rev. 2019, 49, 324–339. [Google Scholar] [CrossRef]
- Davies, M. Veterinary Clinical Nutrition: Success Stories: An Overview. Proc. Nutr. Soc. 2016, 75, 392–397. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; Wang, X.; Hu, Y.; Wu, Q.; Nie, Y.; Dong, J.; Ding, Y.; Yan, L.; Wei, F. Diet Drives Convergent Evolution of Gut Microbiomes in Bamboo-Eating Species. Sci. China Life Sci. 2021, 64, 88–95. [Google Scholar] [CrossRef]
- Wei, F.; Hu, Y.; Yan, L.; Nie, Y.; Wu, Q.; Zhang, Z. Giant Pandas Are Not an Evolutionary Cul-de-Sac: Evidence from Multidisciplinary Research. Mol. Biol. Evol. 2015, 32, 4–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, G.; Wang, L.; Li, J.; Hou, R.; Wang, M.; Wang, Z.; Qu, Q.; Zhou, W.; Nie, Y.; Hu, Y.; et al. Seasonal Shift of the Gut Microbiome Synchronizes Host Peripheral Circadian Rhythm for Physiological Adaptation to a Low-Fat Diet in the Giant Panda. Cell Rep. 2022, 38, 110203. [Google Scholar] [CrossRef]
- Imaoka, S.; Wedlund, P.J.; Ogawa, H.; Kimura, S.; Gonzalez, F.J.; Kim, H.Y. Identification of CYP2C23 Expressed in Rat Kidney as an Arachidonic Acid Epoxygenase. J. Pharmacol. Exp. Ther. 1993, 267, 1012–1016. [Google Scholar]
- Shou, M.; Norcross, R.; Sandig, G.; Lu, P.; Li, Y.; Lin, Y.; Mei, Q.; Rodrigues, A.D.; Rushmore, T.H. Substrate Specificity and Kinetic Properties of Seven Heterologously Expressed Dog Cytochromes P450. Drug Metab. Dispos. 2003, 31, 1161–1169. [Google Scholar] [CrossRef]
- Blaisdell, J.; Goldstein, J.A.; Bai, S.A. Isolation of a New Canine Cytochrome P450 CDNA from the Cytochrome P450 2C Subfamily (CYP2C41) and Evidence for Polymorphic Differences in Its Expression. Drug Metab. Dispos. 1998, 26, 278–283. [Google Scholar]
- Zielinski, J.; Mevissen, M. Inhibition of in Vitro Metabolism of Testosterone in Human, Dog and Horse Liver Microsomes to Investigate Species Differences. Toxicol. Vitr. 2015, 29, 468–478. [Google Scholar] [CrossRef]
- Perez Jimenez, T.E.; Mealey, K.L.; Schnider, D.; Grubb, T.L.; Greene, S.A.; Court, M.H. Identification of Canine Cytochrome P-450s (CYPs) Metabolizing the Tramadol (+)-M1 and (+)-M2 Metabolites to the Tramadol (+)-M5 Metabolite in Dog Liver Microsomes. J. Vet. Pharmacol. Ther. 2018, 41, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Court, M.H. Canine Cytochrome P-450 Pharmacogenetics. Vet. Clin. North. Am. Small Anim. Pract. 2013, 43, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Capdevila, J.H.; Falck, J.R.; Harris, R.C. Cytochrome P450 and Arachidonic Acid Bioactivation: Molecular and Functional Properties of the Arachidonate Monooxygenase. J. Lipid Res. 2000, 41, 163–181. [Google Scholar] [CrossRef]
- Esteves, F.; Rueff, J.; Kranendonk, M. The Central Role of Cytochrome P450 in Xenobiotic Metabolism—A Brief Review on a Fascinating Enzyme Family. J. Xenobiotics 2021, 11, 7. [Google Scholar] [CrossRef]
- Miyazawa, M.; Shindo, M.; Shimada, T. Metabolism of (+)- and (−)-Limonenes to Respective Carveols and Perillyl Alcohols by CYP2C9 and CYP2C19 in Human Liver Microsomes. Drug Metab. Dispos. 2002, 30, 602–607. [Google Scholar] [CrossRef] [Green Version]
- Jiang, R.; Yamaori, S.; Takeda, S.; Yamamoto, I.; Watanabe, K. Identification of Cytochrome P450 Enzymes Responsible for Metabolism of Cannabidiol by Human Liver Microsomes. Life Sci. 2011, 89, 165–170. [Google Scholar] [CrossRef]
- Miyazawa, M.; Sugie, A.; Shimada, T. Roles of Human Cyp2a6 And 2b6 and Rat Cyp2c11 and 2b1 in the 10-Hydroxylation of (–)-Verbenone by Liver Microsomes. Drug Metab. Dispos. 2003, 31, 1049–1053. [Google Scholar] [CrossRef]
- Kendall, K.C. Use of Pine Nuts by Grizzly and Black Bears in the Yellowstone Area. Bears Biol. Manag. 1983, 5, 166. [Google Scholar] [CrossRef] [Green Version]
- Felicetti, L.A.; Schwartz, C.C.; Rye, R.O.; Haroldson, M.A.; Gunther, K.A.; Phillips, D.L.; Robbins, C.T. Use of Sulfur and Nitrogen Stable Isotopes to Determine the Importance of Whitebark Pine Nuts to Yellowstone Grizzly Bears. Can. J. Zool. 2011, 81, 763–770. [Google Scholar] [CrossRef]
- Zulak, K.G.; Bohlmann, J. Terpenoid Biosynthesis and Specialized Vascular Cells of Conifer Defense. J. Integr. Plant. Biol. 2010, 52, 86–97. [Google Scholar] [CrossRef]
- Kopaczyk, J.M.; Warguła, J.; Jelonek, T. The Variability of Terpenes in Conifers under Developmental and Environmental Stimuli. Environ. Exp. Bot. 2020, 180, 104197. [Google Scholar] [CrossRef]
- Karakus, E.; Prinzinger, C.; Leiting, S.; Geyer, J. Sequencing of the Canine Cytochrome P450 CYP2C41 Gene and Genotyping of Its Polymorphic Occurrence in 36 Dog Breeds. Front. Vet. Sci. 2021, 8, 663175. [Google Scholar] [CrossRef]
- DeLozier, T.C.; Tsao, C.C.; Coulter, S.J.; Foley, J.; Bradbury, J.A.; Zeldin, D.C.; Goldstein, J.A. CYP2C44, a New Murine CYP2C That Metabolizes Arachidonic Acid to Unique Stereospecific Products. J. Pharmacol. Exp. Ther. 2004, 310, 845–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capdevila, J.; Wang, W. Role of Cytochrome P450 Epoxygenase in Regulating Renal Membrane Transport and Hypertension. Curr. Opin. Nephrol. Hypertens. 2013, 22, 163–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingelman-Sundberg, M. Human Drug Metabolising Cytochrome P450 Enzymes: Properties and Polymorphisms. Naunyn. Schmiedebergs. Arch. Pharmacol. 2004, 369, 89–104. [Google Scholar] [CrossRef]
- Wright, W.C.; Chenge, J.; Chen, T. Structural Perspectives of the CYP3A Family and Their Small Molecule Modulators in Drug Metabolism. Liver Res. 2019, 3, 132–142. [Google Scholar] [CrossRef]
- Fraser, D.J.; Feyereisen, R.; Harlow, G.R.; Halpert, J.R. Isolation, Heterologous Expression and Functional Characterization of a Novel Cytochrome P450 3A Enzyme from a Canine Liver CDNA Library. J. Pharmacol. Exp. Ther. 1997, 283, 1425–1432. [Google Scholar]
- Locuson, C.W.; Ethell, B.T.; Voice, M.; Lee, D.; Feenstra, K.L. Evaluation of Escherichia Coli Membrane Preparations of Canine CYP1A1, 2B11, 2C21, 2C41, 2D15, 3A12, and 3A26 with Coexpressed Canine Cytochrome P450 Reductase. Drug Metab. Dispos. 2009, 37, 457–461. [Google Scholar] [CrossRef]
- Martinez, S.E.; Shi, J.; Zhu, H.J.; Jimenez, T.E.P.; Zhu, Z.; Court, M.H. Absolute Quantitation of Drug-Metabolizing Cytochrome P450 Enzymes and Accessory Proteins in Dog Liver Microsomes Using Label-Free Standard-Free Analysis Reveals Interbreed Variability. Drug Metab. Dispos. 2019, 47, 1314–1324. [Google Scholar] [CrossRef]
- Sugiyama, S.; Uno, Y.; Amano, T.; Kitazawa, T.; Teraoka, H. Genetic Diversity of Cytochrome P450 3A with Different Metabolic Activity in Domestic Cats. J. Vet. Med. Sci. 2019, 81, 598–600. [Google Scholar] [CrossRef] [Green Version]
- Visser, M.; Weber, K.L.; Lyons, L.A.; Rincon, G.; Boothe, D.M.; Merritt, D.A. Identification and Quantification of Domestic Feline Cytochrome P450 Transcriptome across Multiple Tissues. J. Vet. Pharmacol. Ther. 2019, 42, 7–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, P.P. Pyrrolizidine Alkaloids: Metabolic Activation Pathways Leading to Liver Tumor Initiation. Chem. Res. Toxicol. 2017, 30, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. Incorporation of Absorption and Metabolism into Liver Toxicity Prediction for Phytochemicals: A Tiered in Silico QSAR Approach. Food Chem. Toxicol. 2018, 118, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Huan, J.Y.; Miranda, C.L.; Buhler, D.R.; Cheeke, P.R. The Roles of CYP3A and CYP2B Isoforms in Hepatic Bioactivation and Detoxification of the Pyrrolizidine Alkaloid Senecionine in Sheep and Hamsters. Toxicol. Appl. Pharmacol. 1998, 151, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Sugiyama, S.; Matsushita, M.; Kitazawa, T.; Amano, T.; Uno, Y.; Ikushiro, S.; Teraoka, H. Limited Expression of Functional Cytochrome P450 2c Subtypes in the Liver and Small Intestine of Domestic Cats. Xenobiotica 2018, 49, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Shinkyo, R.; Sakaki, T.; Kasamastu, M.; Imaoka, S.; Funae, Y.; Yokota, H. Cytochrome P450 2E Polymorphism in Feline Liver. Biochim. Biophys. Acta Gen. Subj. 2005, 1726, 194–205. [Google Scholar] [CrossRef]
- Khidkhan, K.; Mizukawa, H.; Ikenaka, Y.; Nakayama, S.M.M.; Nomiyama, K.; Yokoyama, N.; Ichii, O.; Darwish, W.S.; Takiguchi, M.; Tanabe, S.; et al. Tissue Distribution and Characterization of Feline Cytochrome P450 Genes Related to Polychlorinated Biphenyl Exposure. Comp. Biochem. Physiol. Part. C Toxicol. Pharmacol. 2019, 226, 108613. [Google Scholar] [CrossRef]
- Rezaei Khozani, A.; Kaboli, M.; Ashrafi, S.; Akbari, H. Survey Diet of Asiatic Cheetah Acinonyx Jubatus Venaticus by Scat Analysis Method in Bafgh Protected Area, Central Iran. J. Anim. Environ. 2016, 8, 1–8. [Google Scholar]
- Mills, M.G.L.; Broomhall, L.S.; Du Toit, J.T. Cheetah Acinonyx Jubatus Feeding Ecology in the Kruger National Park and a Comparison across African Savanna Habitats: Is the Cheetah Only a Successful Hunter on Open Grassland Plains? Wildlife Biol. 2004, 10, 177–186. [Google Scholar] [CrossRef]
- Schaller, G.B. The Giant Pandas of Wolong; University of Chicago Press: Chicago, IL, USA, 1985; p. 298. [Google Scholar]
- Gudmundson, C.J.; Zeppelin, T.K.; Ream, R.R. Application of Two Methods for Determining Diet of Northern Fur Seals (Callorhinus Ursinus). Fish Bull. 2006, 104, 445–456. [Google Scholar]
- Berta, A.; Churchill, M.; Boessenecker, R.W. The Origin and Evolutionary Biology of Pinnipeds: Seals, Sea Lions, and Walruses. Annu. Rev. Earth Planet. Sci. 2018, 46, 203–228. [Google Scholar] [CrossRef]
- Costa, D.P.; McHuron, E.A. (Eds.) Ethology and Behavioral Ecology of Phocids; Springer: Cham, Switzerland, 2022; p. 645. [Google Scholar] [CrossRef]
- Pauly, D.; Trites, A.W.; Capuli, E.; Christensen, V. Diet Composition and Trophic Levels of Marine Mammals. ICES J. Mar. Sci. 1998, 55, 467–481. [Google Scholar] [CrossRef]
- Bosch, G.; Hagen-Plantinga, E.A.; Hendriks, W.H. Dietary Nutrient Profiles of Wild Wolves: Insights for Optimal Dog Nutrition? Br. J. Nutr. 2015, 113, S40–S54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soars, M.G.; Riley, R.J.; Findlay, K.A.; Coffey, M.J.; Burchell, B. Evidence for Significant Differences in Microsomal Drug Glucuronidation by Canine and Human Liver and Kidney. Drug Metab. Dispos. 2001, 29, 121–126. [Google Scholar]
- Baker, D.H.; Czarnecki-Maulden, G.L. Comparative Nutrition of Cats and Dogs. Annu. Rev. Nutr. 1991, 11, 239–263. [Google Scholar] [CrossRef]
- Montanari, S. Discrimination Factors of Carbon and Nitrogen Stable Isotopes in Meerkat Feces. PeerJ 2017, 2017, e3436. [Google Scholar] [CrossRef] [Green Version]
- Brox, B.W.; Edwards, K.; Buist, N.A.; Macaskill, A.C. Investigating Food Preference in Zoo-Housed Meerkats. Zoo Biol. 2021, 40, 517–526. [Google Scholar] [CrossRef]
- Johnson, C.K.; Tinker, M.T.; Estes, J.A.; Conrad, P.A.; Staedler, M.; Miller, M.A.; Jessup, D.A.; Mazet, J.A.K. Prey Choice and Habitat Use Drive Sea Otter Pathogen Exposure in a Resource-Limited Coastal System. Proc. Natl. Acad. Sci. USA 2009, 106, 2242–2247. [Google Scholar] [CrossRef] [Green Version]
- Rand, K.; McDermott, S.; Logerwell, E.; Matta, M.E.; Levine, M.; Bryan, D.R.; Spies, I.B.; Loomis, T. Higher Aggregation of Key Prey Species Associated with Diet and Abundance of the Steller Sea Lion Eumetopias Jubatus across the Aleutian Islands. Mar. Coast. Fish. 2019, 11, 472–486. [Google Scholar] [CrossRef] [Green Version]
- Edinboro, C.H.; Scott-Moncrieff, J.C.; Janovitz, E.; Thacker, H.L.; Glickman, L.T. Epidemiologic Study of Relationships between Consumption of Commercial Canned Food and Risk of Hyperthyroidism in Cats. J. Am. Vet. Med. Assoc. 2004, 224, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Hammill, M.O.; Stenson, G.B. Estimated Prey Consumption by Harp Seals (Phoca Groenlandica) & Hooded Seals (Cystophora Cristata) & Grey Seals (Halichoerus Grypus) and Harbour Seals (Phoca Vitulina) in Atlantic Canada. J. Northw. Atl. Fish. Sci 2000, 26, 1–23. [Google Scholar]
- Alam, M.S.; Khan, J.A. Food Habits of Striped Hyena (Hyaena Hyaena) in a Semi-Arid Conservation Area of India. J. Arid. Land 2015, 7, 860–866. [Google Scholar] [CrossRef]
- Bhandari, S.; Morley, C.; Aryal, A.; Shrestha, U.B. The Diet of the Striped Hyena in Nepal’s Lowland Regions. Ecol. Evol. 2020, 10, 7953–7962. [Google Scholar] [CrossRef] [PubMed]
- Goetz, K.T.; Burns, J.M.; Hückstӓdt, L.A.; Shero, M.R.; Costa, D.P. Temporal Variation in Isotopic Composition and Diet of Weddell Seals in the Western Ross Sea. Deep Res. Part II Top. Stud. Oceanogr. 2017, 140, 36–44. [Google Scholar] [CrossRef]
- Cote, D.; Stewart, H.M.J.; Gregory, R.S.; Gosse, J.; Reynolds, J.J.; Stenson, G.B.; Miller, E.H. Prey Selection by Marine-Coastal River Otters (Lontra Canadensis) in Newfoundland, Canada. J. Mammal. 2008, 89, 1001–1011. [Google Scholar] [CrossRef] [Green Version]
- Day, C.C.; Westover, M.D.; McMillan, B.R. Seasonal Diet of the Northern River Otter (Lontra Canadensis): What Drives Prey Selection? Can. J. Zool. 2015, 93, 197–205. [Google Scholar] [CrossRef]
- Krawczyk, A.J.; Bogdziewicz, M.; Majkowska, K.; Glazaczow, A. Diet Composition of the Eurasian Otter Lutra Lutra in Different Freshwater Habitats of Temperate Europe: A Review and Meta-Analysis. Mamm. Rev. 2016, 46, 106–113. [Google Scholar] [CrossRef]
- Burstahler, C.M.; Terwissen, C.V.; Roth, J.D. Latitudinal Gradient in Cortisol Concentrations in Canada Lynx (Lynx Canadensis) Is Not Explained by Diet. Can. J. Zool. 2019, 97, 748–753. [Google Scholar] [CrossRef]
- Parker, G.R.; Maxwell, J.W.; Morton, L.D.; Smith, G.E.J. The Ecology of the Lynx (Lynx Canadensis) on Cape Breton Island. Can. J. Zool. 2011, 61, 770–786. [Google Scholar] [CrossRef]
- Cleary, G.P.; Corner, L.A.L.; O’Keeffe, J.; Marples, N.M. The Diet of the Badger Meles Meles in the Republic of Ireland. Mamm. Biol. 2009, 74, 438–447. [Google Scholar] [CrossRef]
- Rita, D.; Drago, M.; Galimberti, F.; Cardona, L. Temporal Consistency of Individual Trophic Specialization in Southern Elephant Seals Mirounga Leonina. Mar. Ecol. Prog. Ser. 2017, 585, 229–242. [Google Scholar] [CrossRef]
- Spurlin, S.M.; Peterson, S.H.; Crocker, D.E.; Costa, D.P. Nitrogen and Carbon Stable-Isotope Ratios Change in Adult Northern Elephant Seals (Mirounga Angustirostris) during the Breeding and Molting Fasts. Mar. Mammal Sci. 2019, 35, 707–717. [Google Scholar] [CrossRef]
- Condit, R.; Le Boeuf, B.J. Feeding Habits and Feeding Grounds of the Northern Elephant Seal. J. Mammal. 1984, 65, 281–290. [Google Scholar] [CrossRef]
- Dell’Arte, G.L.; Laaksonen, T.; Norrdahl, K.; Korpimäki, E. Variation in the Diet Composition of a Generalist Predator, the Red Fox, in Relation to Season and Density of Main Prey. Acta Oecologica 2007, 31, 276–281. [Google Scholar] [CrossRef]
- McDonald, R.A.; Webbon, C.; Harris, S. The Diet of Stoats (Mustela Erminea) and Weasels (Mustela Nivalis) in Great Britain. J. Zool. 2000, 252, 363–371. [Google Scholar] [CrossRef]
- Smith, G.P.; Ragg, J.R.; Waldrup, K.A.; Moller, H. Diet of Feral Ferrets (Mustela Furo) from Pastoral Habitats in Otago and Southland, New Zealand. N. Z. J. Zool. 1995, 22, 363–369. [Google Scholar] [CrossRef]
- Vinke, C.M.; Schoemaker, N.J. The Welfare of Ferrets (Mustela Putorius Furo T): A Review on the Housing and Management of Pet Ferrets. Appl. Anim. Behav. Sci. 2012, 139, 155–168. [Google Scholar] [CrossRef]
- Huang, A.C.; Nelson, C.; Elliott, J.E.; Guertin, D.A.; Ritland, C.; Drouillard, K.; Cheng, K.M.; Schwantje, H.M. River Otters (Lontra Canadensis) “Trapped” in a Coastal Environment Contaminated with Persistent Organic Pollutants: Demographic and Physiological Consequences. Environ. Pollut. 2018, 238, 306–316. [Google Scholar] [CrossRef]
- Krawczyk, A.J.; Bogdziewicz, M.; Czyz, M.J. Diet of the American Mink Neovison Vison in an Agricultural Landscape in Western Poland. Folia Zool. 2013, 62, 304–310. [Google Scholar] [CrossRef]
- Magnusdottir, R.; von Schmalensee, M.; Stefansson, R.A.; Macdonald, D.W.; Hersteinsson, P. A Foe in Woe: American Mink (Neovison Vison) Diet Changes during a Population Decrease. Mamm. Biol. 2014 791 2013, 79, 58–63. [Google Scholar] [CrossRef]
- Robinson, S.; Barbieri, M.; Johanos, T. The Hawaiian Monk Seal: Ethology Applied to Endangered Species Conservation and Recovery. In Ethology and Behavioral Ecology of Phocids; Springer: Cham, Switzerland, 2022; pp. 599–635. [Google Scholar] [CrossRef]
- Sheffield, G.; Grebmeier, J.M. Pacific Walrus (Odobenus Rosmarus Divergens): Differential Prey Digestion and Diet. Mar. Mammal Sci. 2009, 25, 761–777. [Google Scholar] [CrossRef]
- Clark, C.T.; Horstmann, L.; de Vernal, A.; Jensen, A.M.; Misarti, N. Pacific Walrus Diet across 4000 Years of Changing Sea Ice Conditions. Quat. Res. 2022, 108, 26–42. [Google Scholar] [CrossRef]
- Beukes, M.; Radloff, F.G.T.; Ferreira, S.M. Estimating Lion’s Prey Species Profile in an Arid Environment. J. Zool. 2017, 303, 136–144. [Google Scholar] [CrossRef]
- Bothma, J.d.P.; Walker, C. The African Lion. In Larger Carnivores of the African Savannas; Springer: Berlin/Heidelberg, Germany, 1999; pp. 22–59. [Google Scholar] [CrossRef]
- Kshettry, A.; Vaidyanathan, S.; Athreya, V. Diet Selection of Leopards (Panthera Pardus) in a Human-Use Landscape in North-Eastern India. Trop. Conserv. Sci. 2018, 11, 1–8. [Google Scholar] [CrossRef]
- Andheria, A.P.; Karanth, K.U.; Kumar, N.S. Diet and Prey Profiles of Three Sympatric Large Carnivores in Bandipur Tiger Reserve, India. J. Zool. 2007, 273, 169–175. [Google Scholar] [CrossRef]
- Hayward, M.W.; Jedrzejewski, W.; Jedrzewska, B. Prey Preferences of the Tiger P Anthera Tigris. J. Zool. 2012, 286, 221–231. [Google Scholar] [CrossRef]
- Biswas, S.; Sankar, K. Prey Abundance and Food Habit of Tigers (Panthera Tigris Tigris) in Pench National Park, Madhya Pradesh, India. J. Zool. 2002, 256, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Toth, J.; Evert, S.; Zimmermann, E.; Sullivan, M.; Dotts, L.; Able, K.W.; Hagan, R.; Slocum, C. Annual Residency Patterns and Diet of Phoca Vitulina Concolor (Western Atlantic Harbor Seal) in a Southern New Jersey Estuary. Northeast. Nat. 2018, 25, 611–626. [Google Scholar] [CrossRef]
- Ganguly, D.; Adhya, T. How Fishing Cats Prionailurus Viverrinus Bennett, 1833 Fish: Describing a Felid’s Strategy to Hunt Aquatic Prey. Mammalia 2022, 86, 182–189. [Google Scholar] [CrossRef]
- Grassman, L.I.; Tewes, M.E.; Silvy, N.J.; Kreetiyutanont, K. Spatial Organization and Diet of the Leopard Cat (Prionailurus Bengalensis) in North-Central Thailand. J. Zool. 2005, 266, 45–54. [Google Scholar] [CrossRef]
- Rau, J.R.; Jiménez, J.E. Diet of Puma (Puma Concolor, Carnivora: Felidae) in Coastal and Andean Ranges of Southern Chile. Stud. Neotrop. Fauna Environ. 2002, 37, 201–205. [Google Scholar] [CrossRef]
- Aranda, M.; Sánchez-Cordero, V. Prey Spectra of Jaguar (Panthera Onca) and Puma (Puma Concolor) in Tropical Forests of Mexico. Stud. Neotrop. Fauna Environ. 1996, 31, 65–67. [Google Scholar] [CrossRef]
- Tófoli, C.F.; Rohe, F.; Setz, E.Z.F. Jaguarundi (Puma Yagouaroundi) (Geoffroy, 1803) (Carnivora, Felidae) Food Habits in a Mosaic of Atlantic Rainforest and Eucalypt Plantations of Southeastern Brazil. Braz. J. Biol. 2009, 69, 871–877. [Google Scholar] [CrossRef] [Green Version]
- De Cassia Bianchi, R.; Rosa, A.F.; Gatti, A.; Mendes, S.L. Diet of Margay, Leopardus Wiedii, and Jaguarundi, Puma Yagouaroundi, (Carnivora: Felidae) in Atlantic Rainforest, Brazil. Zoologia 2011, 28, 127–132. [Google Scholar] [CrossRef]
- Merkle, J.A.; Polfus, J.L.; Derbridge, J.J.; Heinemeyer, K.S. Dietary Niche Partitioning among Black Bears, Grizzly Bears, and Wolves in a Multiprey Ecosystem. Can. J. Zool. 2017, 95, 663–671. [Google Scholar] [CrossRef]
- Kirby, R.; Alldredge, M.W.; Pauli, J.N. The Diet of Black Bears Tracks the Human Footprint across a Rapidly Developing Landscape. Biol. Conserv. 2016, 200, 51–59. [Google Scholar] [CrossRef]
- Hatch, K.A.; Kester, K.A.; Auger, J.; Roeder, B.L.; Bunnell, K.; Black, H.L. The Effect of Sex, Age, and Location on Carnivory in Utah Black Bears (Ursus Americanus). Oecologia 2019, 189, 931–937. [Google Scholar] [CrossRef]
- Matsubayashi, J.; Otsubo, K.; Morimoto, J.O.; Nakamura, F.; Nose, T.; Tayasu, I. Feeding Habits May Explain the Morphological Uniqueness of Brown Bears on Etorofu Island, Southern Kuril Islands in East Asia. Biol. J. Linn. Soc. 2016, 119, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Mangipane, L.S.; Lafferty, D.J.R.; Joly, K.; Sorum, M.S.; Cameron, M.D.; Belant, J.L.; Hilderbrand, G.V.; Gustine, D.D. Dietary Plasticity and the Importance of Salmon to Brown Bear (Ursus Arctos) Body Size and Condition in a Low Arctic Ecosystem. Polar Biol. 2020, 43, 825–833. [Google Scholar] [CrossRef]
- Johnson, A.C.; Hobson, K.A.; Lunn, N.J.; McGeachy, D.; Richardson, E.S.; Derocher, A.E. Temporal and Intra-Population Patterns in Polar Bear Foraging Ecology in Western Hudson Bay. Mar. Ecol. Prog. Ser. 2019, 619, 187–199. [Google Scholar] [CrossRef] [Green Version]
- Elmhagen, B.; Tannerfeldt, M.; Verucci, P.; Angerbjörn, A. The Arctic Fox (Alopex Lagopus): An Opportunistic Specialist. J. Zool. 2000, 251, 139–149. [Google Scholar] [CrossRef]
- Food Habits of Arctic Foxes (Alopex Lagopus) on the Western Coast of Svalbard on JSTOR. Available online: https://0-www-jstor-org.brum.beds.ac.uk/stable/40511362 (accessed on 31 July 2022).
- Baltrūnaitė, L. Diet and Winter Habitat Use of the Red Fox, Pine Marten and Raccoon Dog in Dzūkija National Park, Lithuania. Acta Zool. Litu. 2006, 16, 46–53. [Google Scholar] [CrossRef]
- Castañeda, I.; Doherty, T.S.; Fleming, P.A.; Stobo-Wilson, A.M.; Woinarski, J.C.Z.; Newsome, T.M. Variation in Red Fox Vulpes Vulpes Diet in Five Continents. Mamm. Rev. 2022, 52, 328–342. [Google Scholar] [CrossRef]
- Weise, M.J.; Harvey, J.T. Temporal Variability in Ocean Climate and California Sea Lion Diet and Biomass Consumption: Implications for Fisheries Management. Mar. Ecol. Prog. Ser. 2008, 373, 157–172. [Google Scholar] [CrossRef]
- Bailey, K.M.; Ainley, D.G. The Dynamics of California Sea Lion Predation on Pacific Hake. Fish. Res. 1981, 1, 163–176. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondo, M.; Ikenaka, Y.; Nakayama, S.M.M.; Kawai, Y.K.; Ishizuka, M. Specific Gene Duplication and Loss of Cytochrome P450 in Families 1-3 in Carnivora (Mammalia, Laurasiatheria). Animals 2022, 12, 2821. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12202821
Kondo M, Ikenaka Y, Nakayama SMM, Kawai YK, Ishizuka M. Specific Gene Duplication and Loss of Cytochrome P450 in Families 1-3 in Carnivora (Mammalia, Laurasiatheria). Animals. 2022; 12(20):2821. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12202821
Chicago/Turabian StyleKondo, Mitsuki, Yoshinori Ikenaka, Shouta M. M. Nakayama, Yusuke K. Kawai, and Mayumi Ishizuka. 2022. "Specific Gene Duplication and Loss of Cytochrome P450 in Families 1-3 in Carnivora (Mammalia, Laurasiatheria)" Animals 12, no. 20: 2821. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12202821