
Dairy Buffalo Behavior: Calving, Imprinting and Allosuckling

 ,
,  ,
,  ,
,  ,
,  , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
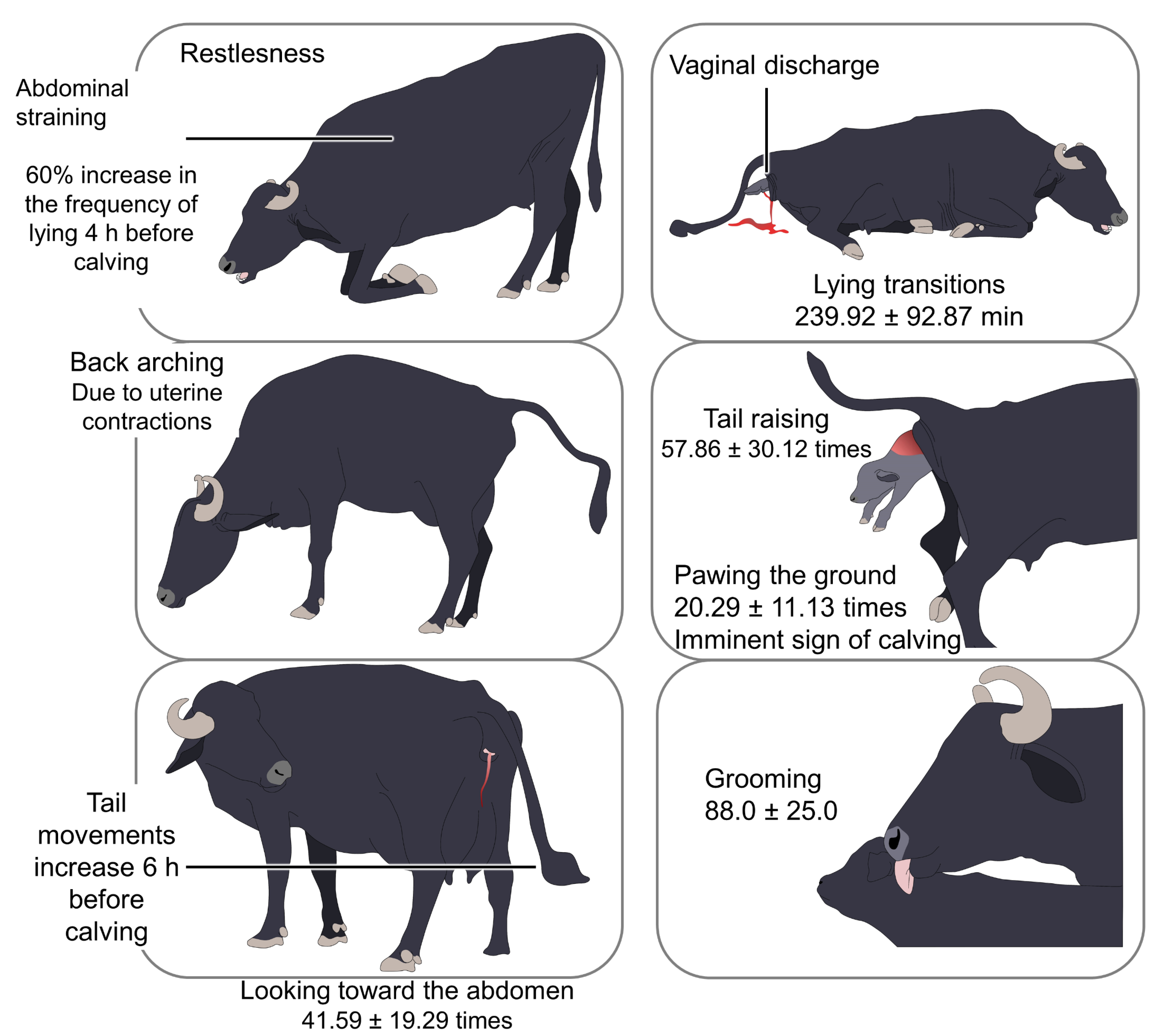
2. Calving Behavior
3. Calving Complication Behavior
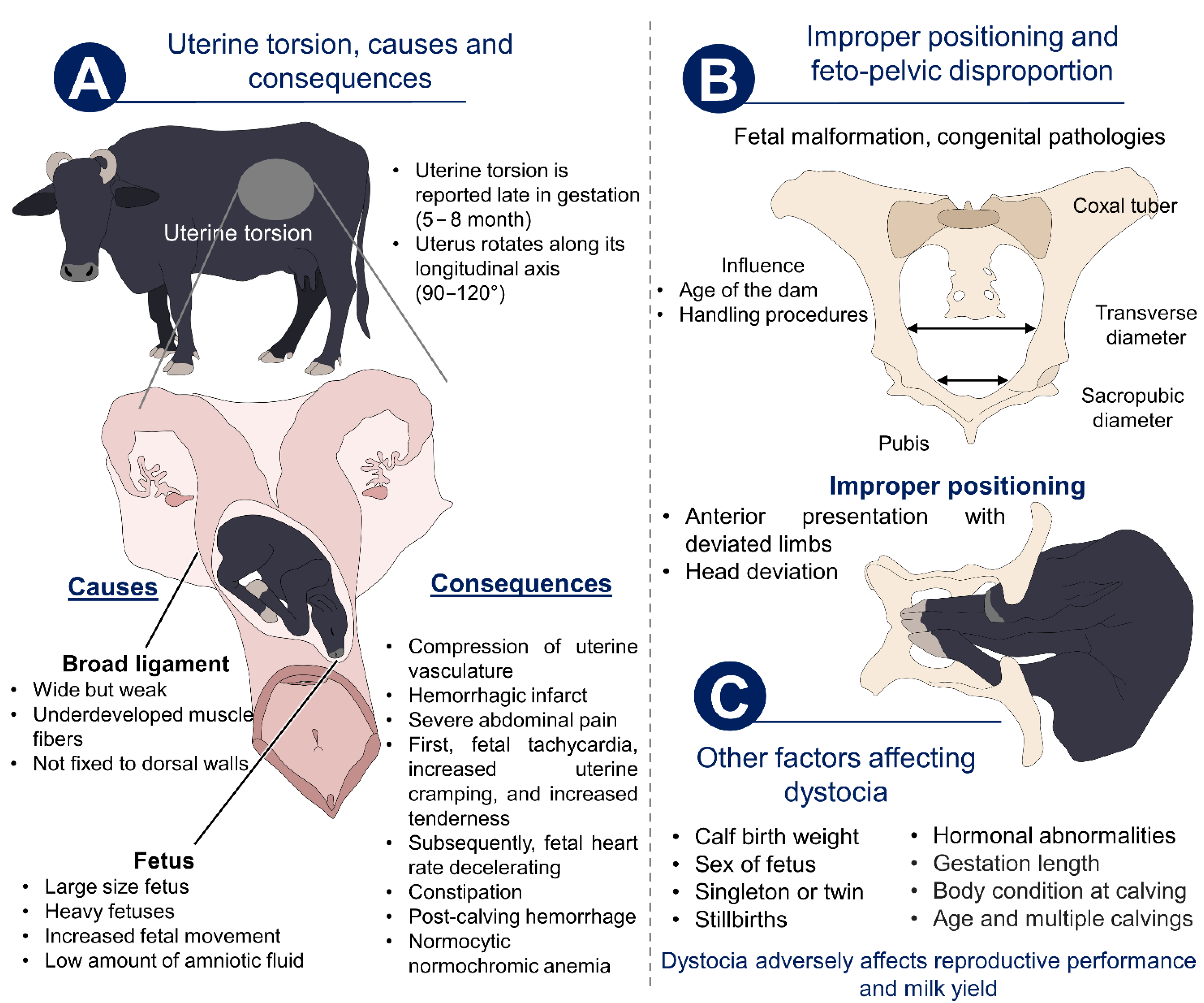
3.1. Maternal Causes
3.2. Fetal Complications
4. Imprinting Mechanisms
4.1. Tactile Stimulation
4.2. Olfactory Stimulation
4.3. Visual Stimulation
4.4. Auditory Stimulation
5. Factors That Contribute to the Survival of the Offspring
6. Maternal Care of the Newborn and Allosuckling
7. Perspectives
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.; Colli, L.; Barker, J.S.F. Asian water buffalo: Domestication, history and genetics. Anim. Genet. 2020, 51, 177–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertoni, A.; Álvarez-Macías, A.; Mota-Rojas, D.; Dávalos, J.L.; Minervino, A.H.H. Dual-purpose water buffalo production systems in tropical latin america: Bases for a sustainable model. Animals 2021, 11, 2910. [Google Scholar] [CrossRef] [PubMed]
- Zicarelli, L. Current trends in buffalo milk production. J. Buffalo Sci. 2020, 9, 121–132. [Google Scholar] [CrossRef]
- Catillo, G.; Macciotta, N.P.P.; Carretta, A.; Cappio-Borlino, A. Effects of age and calving season on lactation curves of milk production traits in italian water buffaloes. J. Dairy Sci. 2002, 85, 1298–1306. [Google Scholar] [CrossRef]
- Coria-Avila, G.A.; Pfaus, J.G.; Orihuela, A.; Domínguez-Oliva, A.; José-Pérez, N.; Hernández, L.A.; Mota-Rojas, D. The neurobiology of behavior and its applicability for animal welfare: A review. Animals 2022, 12, 928. [Google Scholar] [CrossRef]
- Poindron, P.; Lévy, F.; Krehbiel, D. Genital, olfactory, and endocrine interactions in the development of maternal behaviour in the parturient ewe. Psychoneuroendocrinology 1988, 13, 99–125. [Google Scholar] [CrossRef]
- Numan, M. Medial preoptic area and maternal behavior in the female rat. J. Comp. Physiol. Psychol. 1974, 87, 746–759. [Google Scholar] [CrossRef]
- Tulloch, G. The water buffalo, bubalus buba lis, in australia: Reproductive and parent-offspring behaviour. Aust. Wildl. Res 1979, 6, 265–287. [Google Scholar] [CrossRef]
- Das, G.; Kumar, A.; Dangi, S.; Khan, F. Parturition and puerperium in the buffalo. In Bubaline Theriogenology; Purohit, G., Ed.; International Veterinary Information Service: Ithaca, NY, USA, 2013; pp. 238–251. [Google Scholar]
- Poletto, R.; Meisel, R.L.; Richert, B.T.; Cheng, H.W.; Marchant-Forde, J.N. Behavior and peripheral amine concentrations in relation to ractopamine feeding, sex, and social rank of finishing pigs1. J. Anim. Sci. 2010, 88, 1184–1194. [Google Scholar] [CrossRef] [Green Version]
- Purohit, G.N.; Kumar, P.; Solanki, K.; Shekher, C.; Yadav, S.P. Perspectives of fetal dystocia in cattle and buffalo. Vet. Sci. Dev. 2012, 2, 8. [Google Scholar] [CrossRef]
- Ajith, B.; Aswathanarayanappa, V.; Pavankumar, K.; Vishwanath, B.; Ramesh, D.; Manjunatha, K. surgical intervention of dystocia in delayed condition of uterine torion in a buffalo-a case report. Vet. Pract. 2013, 14, 375–376. [Google Scholar]
- Megahed, G.A. Retrospective study on the fetal maldispositions as a cause of dystocia in egyptian buffalo-cows: Strategic plan to improve. J. Dairy, Vet. Anim. Res. 2016, 3, 1–3. [Google Scholar] [CrossRef]
- Srinivas, M.; Sreenu, M.; Rani, N.L.; Naidu, K.S.; Prasad, V.D. Studies on dystocia in graded Murrah buffaloes: A retrospective study. Buffalo Bull. 2007, 26, 40–45. [Google Scholar]
- Gloria, A.; Chincarini, M.; Vignola, G.; Ferri, N.; Contri, A. Venous blood gas parameters in healthy Mediterranean buffalo calves in the first 72 hours of life. Theriogenology 2020, 157, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Salem, M.M.I.; Amin, A.M.S. Risk factors and genetic evaluation of stillbirth trait in buffalo. Livest. Sci. 2017, 206, 132–134. [Google Scholar] [CrossRef]
- Nasr, M.A.F. The effect of stillbirth on reproductive and productive performance of pure Egyptian buffaloes and their crosses with Italian buffaloes. Theriogenology 2017, 103, 9–16. [Google Scholar] [CrossRef]
- Juozaitiene, V.; Juozaitis, A.; Kardisauskas, A.; Zymantiene, J.; Zilaitis, V.; Antanaitis, R.; Ruzauskas, M. Relationship between dystocia and the lactation number, stillbirth and mastitis prevalence in dairy cows. Acta Vet. Brno 2017, 86, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Hernández, H.; Terrazas, A.; Poindron, P.; Ramírez-Vera, S.; Flores, J.A.; Delgadillo, J.A.; Vielma, J.; Duarte, G.; Fernández, I.G.; Fitz-Rodríguez, G.; et al. Sensorial and physiological control of maternal behavior in small ruminants: Sheep and goats. Trop. Subtrop. Agroecosystems 2012, 15, S91–S102. [Google Scholar]
- Williams, G.L. Endocrine regulation of maternal behavior postpartum. In Endocrinology of Pregnancy; Bazer, F.W., Ed.; Springer: New York, NY, USA, 1988; pp. 555–568. [Google Scholar]
- Mora-Medina, P.; Napolitano, F.; Mota-Rojas, D.; Berdugo-Gutiérrez, J.; Ruiz-Buitrago, J.; Guerrero-Legarreta, I. Imprinting, sucking and allosucking behaviors in buffalo calves. J. Buffalo Sci. 2018, 7, 49–57. [Google Scholar] [CrossRef]
- Napolitano, F.; Pacelli, C.; Grasso, F.; Braghieri, A.; De Rosa, G. The behaviour and welfare of buffaloes (Bubalus bubalis) in modern dairy enterprises. Animal 2013, 7, 1704–1713. [Google Scholar] [CrossRef] [Green Version]
- Kharkar, K.P.; Raghuwanshi, D.S.; Thakre, P.D.; Lende, S.R.; Khati, B.M. Effect of Non-Genetic Parameters on Mortality Pattern in Nagpuri Buffalo Calves. J. Anim. Health Prod. 2019, 7, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Sreedhar, S.; Ranganadham, M.; Mohan, E.M. Calf mortality in indigenous buffaloes. Indian Vet. J. 2010, 87, 197–198. [Google Scholar]
- Jenny, B.; Cramling, G.; Glaze, T. Management factors associated with calf mortality in South Carolina dairy herds. J. Dairy Sci. 1981, 64, 2284–2289. [Google Scholar] [CrossRef]
- Dwyer, C.M.; Conington, J.; Corbiere, F.; Holmøy, I.H.; Muri, K.; Nowak, R.; Rooke, J.; Vipond, J.; Gautier, J.-M. Invited review: Improving neonatal survival in small ruminants: Science into practice. Animal 2016, 10, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Fordyce, G.; Olchowy, T.; Anderson, A. Hydration in non-suckling neonatal Brahman-cross calves. Aust. Vet. J. 2015, 93, 214–220. [Google Scholar] [CrossRef]
- Napolitano, F.; Cifuni, G.F.; Pacelli, C.; Riviezzi, A.M.; Girolami, A. Effect of artificial rearing on lamb welfare and meat quality. Meat Sci. 2002, 60, 307–315. [Google Scholar] [CrossRef]
- Shah, A.M.; Naeem, M.; Shah, M.G.; Haaroon, M.; Peng, Q.; Wang, Z. Effects of various colostrum feeding methods on growth performance and immunity of holstein-friesian calves. Pak. J. Zool. 2019, 51, 2161–2166. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Marcet-Rius, M.; Freitas-de-Melo, A.; Muns, R.; Mora-Medina, P.; Domínguez-Oliva, A.; Orihuela, A. Allonursing in wild and farm animals: Biological and physiological foundations and explanatory hypotheses. Animals 2021, 11, 3092. [Google Scholar] [CrossRef]
- Gloneková, M.; Brandlová, K.; Pluháček, J. Stealing milk by young and reciprocal mothers: High incidence of allonursing in giraffes, Giraffa camelopardalis. Anim. Behav. 2016, 113, 113–123. [Google Scholar] [CrossRef]
- Zapata, B.; Correa, L.; Soto-Gamboa, M.; Latorre, E.; González, B.A.; Ebensperger, L.A. Allosuckling allows growing offspring to compensate for insufficient maternal milk in farmed guanacos (Lama guanicoe). Appl. Anim. Behav. Sci. 2010, 122, 119–126. [Google Scholar] [CrossRef]
- Barrier, A.; Haskell, M. Calving difficulty in dairy cows has a longer effect on saleablemilk yield than on estimatedmilk production. J. Dairy Sci. 2011, 94, 1804–1812. [Google Scholar] [CrossRef] [Green Version]
- Barrier, A.; Haskell, M.; Dwyer, C. Dystocial dairy calves: Condemned to poor welfare? In Proceedings of the 5th WAFL Conference, Guelph, ON, Canada, 8–11 August 2011; p. 64. [Google Scholar]
- Mee, J. Prevalence and risk factors for dystocia in dairy cattle: A review. Veternary J. 2008, 176, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Barrier, A.C.; Haskell, M.J.; Macrae, A.I.; Dwyer, C.M. Parturition progress and behaviours in dairy cows with calving difficulty. Appl. Anim. Behav. Sci. 2012, 139, 209–217. [Google Scholar] [CrossRef]
- Roberts, S. Veterinary Obstetrics and Genital Disease; CBS Publishers and Distributors: New Delhi, India, 1971. [Google Scholar]
- Singh, R.; Khar, S.; Chander, S. Parturition in buffaloes. Indian J. Anim. Sci. 1994, 64, 1028–1033. [Google Scholar]
- Górriz-Martín, L.; Koenig, A.; Jung, K.; Bergforth, W.; von Soosten, D.; Hoedemaker, M.; Bajcsy, Á.C. Comparison between a calving predictive system and a routine prepartal examination in german holstein heifers and cows. Vet. Sci. 2022, 9, 192. [Google Scholar] [CrossRef]
- Deka, R.; Nath, K.C.; Bhuyan, M.; Bhuyan, D.; Das, G.C.; Dutta, L.; Borpujari, D. Parturition behavior of Swamp buffalo cows (Bubalus bubalis) under organized system of rearing. Biol. Rhythm Res. 2021, 52, 444–453. [Google Scholar] [CrossRef]
- Martínez-Burnes, J.; Muns, R.; Barrios-García, H.; Villanueva-García, D.; Domínguez-Oliva, A.; Mota-Rojas, D. Parturition in mammals: Animal models, pain and distress. Animals 2021, 11, 2960. [Google Scholar] [CrossRef]
- De Rosa, G.; Grasso, F.; Braghieri, A.; Bilancione, A.; Di Francia, A.; Napolitano, F. Behavior and milk production of buffalo cows as affected by housing system. J. Dairy Sci. 2009, 92, 907–912. [Google Scholar] [CrossRef] [Green Version]
- von Keyserlingk, M.A.G.; Weary, D.M. Maternal behavior in cattle. Horm. Behav. 2007, 52, 106–113. [Google Scholar] [CrossRef]
- Dietz, H.; Clarke, B.; Vancaillie, T. Vaginal childbirth and bladder neck mobility. Aust. New Zeal. J. Obstet. Gynaecol. 2002, 42, 522–525. [Google Scholar] [CrossRef]
- Lanzoni, L.; Chincarini, M.; Giammarco, M.; Fusaro, I.; Gloria, A.; Contri, A.; Ferri, N.; Vignola, G. Maternal and neonatal behaviour in italian mediterranean buffaloes. Animals 2021, 11, 1584. [Google Scholar] [CrossRef] [PubMed]
- Huzzey, J.M.; von Keyserlingk, M.A.G.; Weary, D.M. Changes in feeding, drinking, and standing behavior of dairy cows during the transition period. J. Dairy Sci. 2005, 88, 2454–2461. [Google Scholar] [CrossRef] [Green Version]
- Lanzoni, L.; Chincarini, M.; Giammarco, M.; Fusaro, I.; Iannotta, M.; Podaliri, M.; Contri, A.; Gloria, A.; Vignola, G. Changes in the behaviour before normal calving to predict its onset in Mediterranean buffaloes heifers. Appl. Anim. Behav. Sci. 2022, 254, 105721. [Google Scholar] [CrossRef]
- Dubey, P.; Singh, R.R.; Choudhary, S.S.; Verma, K.K.; Kumar, A.; Gamit, P.M.; Dubey, S.; Prajapati, K. Post parturient neonatal behaviour and their relationship with maternal behaviour score, parity and sex in surti buffaloes. J. Appl. Anim. Res. 2018, 46, 360–364. [Google Scholar] [CrossRef]
- Mainau, E.; Manteca, X. Pain and discomfort caused by parturition in cows and sows. Appl. Anim. Behav. Sci. 2011, 135, 241–251. [Google Scholar] [CrossRef]
- Weary, D.M.; Niel, L.; Flower, F.C.; Fraser, D. Identifying and preventing pain in animals. Appl. Anim. Behav. Sci. 2006, 100, 64–76. [Google Scholar] [CrossRef] [Green Version]
- Slater, R.; Cantarella, A.; Franck, L.; Meek, J.; Fitzgerald, M. How well do clinical pain assessment tools reflect pain in infants? PLoS Med. 2008, 5, e129. [Google Scholar] [CrossRef] [Green Version]
- Kebede, A.; Mohammed, A.; Tadessse, W.; Abera, D. Review on economic impacts of dystocia in dairy farm and its management and prevention methods. Nat. Sci. 2017, 15, 32–42. [Google Scholar] [CrossRef]
- Ahuja, A.K.; Dogra, P.; Kumar, S.; Dhindsa, S.S.; Singh, H. A case of dystocia due to fetal ascites in murrah buffalo. Int. J. Environ. Agric. Biotechnol. 2017, 2, 1767–1769. [Google Scholar] [CrossRef] [Green Version]
- Amin, S.M.; Amer, H.A.; Hussein, A.E.; Hazzaa, A.M. Creatine phosphokinase and aspartate aminotransverase profiles and its relation to the severity of uterine torsion in Egyptian buffalo. Anim. Reprod. Sci. 2011, 123, 163–168. [Google Scholar] [CrossRef]
- Meena, M.; Kumar, P.M.; Soni, Y.; Meena, M. Uterine torsion in cross bred murrah buffaloes and its clinical management. Pharma Innov. 2022, 11, 2246–2249. [Google Scholar]
- Qazi, M.; Shukla, S.; Nema, S.; Ali, R.; Mahor, S. Management of uterine torsion in a she buffalo. Buffalo Bull. 2010, 29, 75–77. [Google Scholar]
- Mohammad, D.R.I.; Abdel-Rahman, M.A.M. A comparative study on behavioral, physiological, and adrenal changes in buffaloes during the first stage of labor with normal and difficult parturition. J. Vet. Behav. 2013, 8, 46–50. [Google Scholar] [CrossRef]
- González-Lozano, M.; Mota-Rojas, D.; Orihuela, A.; Martínez-Burnes, J.; Di Francia, A.; Braghieri, A.; Berdugo-Gutiérrez, J.; Mora-Medina, P.; Ramírez-Necoechea, R.; Napolitano, F. Review: Behavioral, physiological, and reproductive performance of buffalo cows during eutocic and dystocic parturitions. Appl. Anim. Sci. 2020, 36, 407–422. [Google Scholar] [CrossRef]
- Samad, H.; Ali, C.; Rehman, N.; Ahmad, A.; Ahmad, N. Clinical incidence of reproductive disorders in buffaloes. Pak. Vet. J. 1987, 7, 16–19. [Google Scholar]
- Han, Y.K.; Kim, I.H. Risk factors for retained placenta and the effect of retained placenta on the occurrence of postpartum diseases and subsequent reproductive performance in dairy cows. J. Vet. Sci. 2005, 6, 53. [Google Scholar] [CrossRef]
- Mahmoud, E.A.; Essawi, W.M.; Neamat-Allah, A.N.F. Influence of uterine torsion in water buffaloes (Bubalus bubalis) with insights into the hematological and biochemical prognostic values regarding to manual correction. Trop. Anim. Health Prod. 2020, 52, 3165–3171. [Google Scholar] [CrossRef]
- Kumar, P.; Sharma, A.; Vasishta, N.; Khan, S.; Barman, P. Dystocia due to relative oversized fetus and fetal maldisposition in a buffalo. Vet. World 2011, 4, 569. [Google Scholar] [CrossRef]
- Das, G.; Dutt, R.; Kumar, R.; Deori, S.; Shanker, U. Dystocia due to fetal maldisposition in a buffalo. Buffalo Bull. 2009, 28, 59–60. [Google Scholar]
- Jensen, M.B. The effects of feeding method, milk allowance and social factors on milk feeding behaviour and cross-sucking in group housed dairy calves. Appl. Anim. Behav. Sci. 2003, 80, 191–206. [Google Scholar] [CrossRef]
- Charrier, I.; Mathevon, N.; Jouventin, P. Vocal signature recognition of mothers by fur seal pups. Anim. Behav. 2003, 65, 543–550. [Google Scholar] [CrossRef] [Green Version]
- Hudson, S.J.; Mullord, M.M. Investigations of maternal bonding in dairy cattle. Appl. Anim. Ethol. 1977, 3, 271–276. [Google Scholar] [CrossRef]
- Orihuela, A.; Mota-Rojas, D.; Strappini, A.; Serrapica, F.; Braghieri, A.; Mora-Medina, P.; Napolitano, F. Neurophysiological mechanisms of cow–calf bonding in buffalo and other farm animals. Animals 2021, 11, 1968. [Google Scholar] [CrossRef] [PubMed]
- Mandujano, H.; Mandujano-Camacho, H.; Mandujano, C.; Mandujano-Camacho, H. Ecología y sociobiología de la impronta: Perspectivas para su estudio en los Crocodylia. Cienc. y Mar 2010, 14, 49–54. [Google Scholar]
- Yamaguchi, S.; Aoki, N.; Kitajima, T.; Iikubo, E.; Katagiri, S.; Matsushima, T.; Homma, K.J. Thyroid hormone determines the start of the sensitive period of imprinting and primes later learning. Nat. Commun. 2012, 3, 1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knudsen, E.I. Sensitive periods in the development of the brain and behavior. J. Cogn. Neurosci. 2004, 16, 1412–1425. [Google Scholar] [CrossRef]
- Nowak, R.; Keller, M.; Val-Laillet, D.; Lévy, F. Perinatal visceral events and brain mechanisms involved in the development of mother–young bonding in sheep. Horm. Behav. 2007, 52, 92–98. [Google Scholar] [CrossRef]
- López-Arjona, M.; Mainau, E.; Navarro, E.; Contreras-Aguilar, M.D.; Escribano, D.; Mateo, S.V.; Manteca, X.; Cerón, J.J.; Martínez-Subiela, S. Oxytocin in bovine saliva: Validation of two assays and changes in parturition and at weaning. BMC Vet. Res. 2021, 17, 140. [Google Scholar] [CrossRef]
- Lévy, F. Neuroendocrine control of maternal behavior in non-human and human mammals. Ann. Endocrinol. 2016, 77, 114–125. [Google Scholar] [CrossRef]
- Mora-Medina, P.; Orihuela-Trujillo, A.; Arch-Tirado, E.; Santiago, R.; Terrazas, A.; Mota-Rojas, D. Sensory factors involved in mother-young bonding in sheep: A review. Vet. Med. 2016, 61, 595–611. [Google Scholar] [CrossRef]
- Dwyer, C.M. Maternal behaviour and lamb survival: From neuroendocrinology to practical application. Animal 2014, 8, 102–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Han, C.; Li, M.; Li, F.; Yang, Y.; Wang, Z.; Lv, S. Effects of parity, litter size and lamb sex on maternal behavior of small Tail Han sheep and their neuroendocrine mechanisms. Small Rumin. Res. 2021, 202, 106451. [Google Scholar] [CrossRef]
- Lv, S.-J.; Yang, Y.; Li, F.-K. Parity and litter size effects on maternal behavior of Small Tail Han sheep in China. Anim. Sci. J. 2016, 87, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Kristal, M.B. The biopsychology of maternal behavior in nonhuman mammals. ILAR J. 2009, 50, 51–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kendrick, K.M.; Da Costa, A.P.; Broad, K.D.; Ohkura, S.; Guevara, R.; Lévy, F.; Keverne, E.B. Neural control of maternal behaviour and olfactory recognition of offspring. Brain Res. Bull. 1997, 44, 383–395. [Google Scholar] [CrossRef]
- Niell, C.M.; Smith, S.J. Live optical imaging of nervous system development. Annu. Rev. Physiol. 2004, 66, 771–798. [Google Scholar] [CrossRef] [PubMed]
- Ruthazer, E.S.; Cline, H.T. Insights into activity-dependent map formation from the retinotectal system: A middle-of-the-brain perspective. J. Neurobiol. 2004, 59, 134–146. [Google Scholar] [CrossRef] [Green Version]
- Madigan, S.; Bakermans-Kranenburg, M.J.; Van Ijzendoorn, M.H.; Moran, G.; Pederson, D.R.; Benoit, D. Unresolved states of mind, anomalous parental behavior, and disorganized attachment: A review and meta-analysis of a transmission gap. Attach. Hum. Dev. 2006, 8, 89–111. [Google Scholar] [CrossRef]
- Rowson, L.E.A.; McNeilly, A.S.; O’Brien, C.A. The effect of vaginal and cervical stimulation on oxytocin release during the luteal phase of the cow’s oestrous cycle. Reproduction 1972, 30, 287–288. [Google Scholar] [CrossRef]
- Hedges, V.L.; Heaton, E.C.; Amaral, C.; Benedetto, L.E.; Bodie, C.L.; D’Antonio, B.I.; Davila Portillo, D.R.; Lee, R.H.; Levine, M.T.; O’Sullivan, E.C.; et al. Estrogen withdrawal increases postpartum anxiety via oxytocin plasticity in the paraventricular hypothalamus and dorsal raphe nucleus. Biol. Psychiatry 2021, 89, 929–938. [Google Scholar] [CrossRef]
- Murray, C.F.; Leslie, K.E. Newborn calf vitality: Risk factors, characteristics, assessment, resulting outcomes and strategies for improvement. Vet. J. 2013, 198, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Insel, T.R.; Young, L.J. The neurobiology of attachment. Nat. Rev. Neurosci. 2001, 2, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Newby, N.C.; Duffield, T.F.; Pearl, D.L.; Leslie, K.E.; LeBlanc, S.J.; von Keyserlingk, M.A.G. Short communication: Use of a mechanical brush by Holstein dairy cattle around parturition. J. Dairy Sci. 2013, 96, 2339–2344. [Google Scholar] [CrossRef] [Green Version]
- Lickliter, R.E. Behavior associated with parturition in the domestic goat. Appl. Anim. Behav. Sci. 1985, 13, 335–345. [Google Scholar] [CrossRef]
- Broom, D.; Fraser, A. Domestic Animal Behaviour and Welfare, 5th ed.; CABI: Wallingford, UK, 2015. [Google Scholar]
- Meaney, M.J. Maternal care, gene expression, and the transmission of individual differences in stress reactivity across generations. Annu. Rev. Neurosci. 2001, 24, 1161–1192. [Google Scholar] [CrossRef] [PubMed]
- Francis, D.D.; Young, L.J.; Meaney, M.J.; Insel, T.R. Naturally occurring differences in maternal care are associated with the expression of oxytocin and vasopressin (V1a) receptors: Gender differences. J. Neuroendocrinol. 2002, 14, 349–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, G.L.; Gazal, O.S.; Leshin, L.S.; Stanko, R.L.; Anderson, L.L. Physiological regulation of maternal behavior in heifers: Roles of genital stimulation, intracerebral oxytocin release, and ovarian steroids. Biol. Reprod. 2001, 65, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Jainudeen, M.R.; Hafez, E.S.E. Gestation, prenatal, physiology, and parturition. In Reproduction in Farm Animals; Hafez, E., Hafez, B., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2000; pp. 140–155. [Google Scholar]
- Metz, J.; Metz, J.H.M. Maternal influence on defecation and urination in the newborn calf. Appl. Anim. Behav. Sci. 1986, 16, 325–333. [Google Scholar] [CrossRef]
- Lezama-García, K.; Mariti, C.; Mota-Rojas, D.; Martínez-Burnes, J.; Barrios-García, H.; Gazzano, A. Maternal behaviour in domestic dogs. Int. J. Vet. Sci. Med. 2019, 7, 20–30. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Orihuela, A.; Napolitano, F.; Mora-Medina, P.; Gregorio, O.; Alonso-Spilsbury, M. Olfaction in animal behaviour and welfare. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2018, 13, 1–13. [Google Scholar] [CrossRef]
- Edwards, S.A.; Broom, D.M. Behavioural interactions of dairy cows with their newborn calves and the effects of parity. Anim. Behav. 1982, 30, 525–535. [Google Scholar] [CrossRef]
- Newberry, R.C.; Swanson, J.C. Implications of breaking mother–young social bonds. Appl. Anim. Behav. Sci. 2008, 110, 3–23. [Google Scholar] [CrossRef]
- Wyatt, T.D. Pheromones and animal behaviour. J. Exp. Biol. 2003, 206, 4382–4383. [Google Scholar] [CrossRef] [Green Version]
- Lévy, F.; Keller, M. Neurobiology of Maternal Behavior in Sheep. In Advances in the Study of Behavior; Brockmann, H.J., Roper, T., Naguib, M., Wynne-Edwards, K., Barnard, C., Mitani, J., Eds.; Academic Press: Cambridge, MA, USA, 2008; Volume 38, pp. 399–437. ISBN 9780120045389. [Google Scholar]
- Kendrick, K.M.; Guevara-Guzman, R.; Zorrilla, J.; Hinton, M.R.; Broad, K.D.; Mimmack, M.; Ohkura, S. Formation of olfactory memories mediated by nitric oxide. Nature 1997, 388, 670–674. [Google Scholar] [CrossRef] [PubMed]
- Brennan, P.A.; Kendrick, K.M. Mammalian social odours: Attraction and individual recognition. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 2061–2078. [Google Scholar] [CrossRef] [Green Version]
- Keller, M.; Lévy, F. The main but not the accessory olfactory system is involved in the processing of socially relevant chemosignals in ungulates. Front. Neuroanat. 2012, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Booth, K.K. The significance of the vomeronasal organ for offspring recognition in sheep. Small Rumin. Res. 2006, 62, 39–41. [Google Scholar] [CrossRef]
- Wenker, M.L.; Verwer, C.M.; Bokkers, E.A.M.; te Beest, D.E.; Gort, G.; de Oliveira, D.; Koets, A.; Bruckmaier, R.M.; Gross, J.J.; van Reenen, C.G. Effect of type of cow-calf contact on health, blood parameters, and performance of dairy cows and calves. Front. Vet. Sci. 2022, 9, 855086. [Google Scholar] [CrossRef]
- Napolitano, F.; Mota-Rojas, D.; Martínez-Burnes, J.; De Rosa, G.; Hufana Duran, D.; Braghieri, A.; González-Lozano, M.; Di Francia, A.; Serrapica, F.; Orihuela, A. Perinatology and gynecology of the water buffalo. In El búfalo de Agua en Latinoamerica, Hallazgos Recientes; Napolitano, F., Mota-Rojas, D., Guerrero- Legarreta, I., Orihuela, A., Eds.; BM Editores: Mexico City, Mexico, 2020; pp. 535–563. ISBN 9786079900816. Available online: https://lifescienceglobal.com/journals/journal-of-buffalo-science/97-abstract/jbs/4550-el-bufalo-de-agua-en-latinoamerica-hallazgos-recientes (accessed on 15 August 2020).
- Poindron, P.; Terrazas, A.; de la Luz Navarro Montes de Oca, M.; Serafín, N.; Hernández, H. Sensory and physiological determinants of maternal behavior in the goat (Capra hircus). Horm. Behav. 2007, 52, 99–105. [Google Scholar] [CrossRef]
- Rodríguez-González, D.; Minervino, A.H.H.; Orihuela, A.; Bertoni, A.; Morales-Canela, D.A.; Álvarez-Macías, A.; José-Pérez, N.; Domínguez-Oliva, A.; Mota-Rojas, D. Handling and physiological aspects of the dual-purpose water buffalo production system in the mexican humid tropics. Animals 2022, 12, 608. [Google Scholar] [CrossRef]
- Torriani, M.V.G.; Vannoni, E.; McElligott, A.G. Mother-Young recognition in an ungulate hider species: A unidirectional process. Am. Nat. 2006, 168, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Searby, A.; Jouventin, P. Mother-lamb acoustic recognition in sheep: A frequency coding. Proc. R. Soc. London. Ser. B Biol. Sci. 2003, 270, 1765–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sèbe, F.; Duboscq, J.; Aubin, T.; Ligout, S.; Poindron, P. Early vocal recognition of mother by lambs: Contribution of low- and high-frequency vocalizations. Anim. Behav. 2010, 79, 1055–1066. [Google Scholar] [CrossRef]
- Barfield, C.H.; Tang-Martinez, Z.; Trainer, J.M. Domestic calves (Bos taurus) recognize their own mothers by auditory cues. Ethology 2010, 97, 257–264. [Google Scholar] [CrossRef]
- Alexander, G. Role of auditory and visual cues in mutual recognition between ewes and lambs in merino sheep. In Reproductive and Developmental Behaviour in Sheep; Elsevier: Amsterdam, The Netherlands, 1985; pp. 321–337. [Google Scholar]
- Green, A.C.; Lidfors, L.M.; Lomax, S.; Favaro, L.; Clark, C.E.F. Vocal production in postpartum dairy cows: Temporal organization and association with maternal and stress behaviors. J. Dairy Sci. 2021, 104, 826–838. [Google Scholar] [CrossRef]
- Padilla de la Torre, M.; Briefer, E.F.; Ochocki, B.M.; McElligott, A.G.; Reader, T. Mother–offspring recognition via contact calls in cattle, Bos taurus. Anim. Behav. 2016, 114, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Khan, Z.Z.U.; Khan, S.; Ahmad, N.; Raziq, A. Investigation of mortality incidence and managemental practices in buffalo calves at commercial dairy farms in Peshawar City. J. Agric. 2007, 2, 16–22. [Google Scholar]
- Afzal, M.M.H.; Anjum, A. Calf mortality: Season pattern, age distribution and causes of calf mortality. Pak. Vet. J. 1983, 3, 30–33. [Google Scholar]
- Shakya, A.; Roy, B.; Baghel, R. Studies on buffalo calf mortality in Jabalpur district of Madhya Pradesh. Buffalo Bull. 2017, 36, 521–524. [Google Scholar]
- Khan, A.; Khan, M. Epidemiological aspects of neonatal calf mortality in the Nili-Ravi buffaloes. Pak. Vet. J. 1995, 15, 163–166. [Google Scholar]
- Hogan, L.A.; McGowan, M.R.; Johnston, S.D.; Lisle, A.T.; Schooley, K. Suckling behaviour of beef calves during the first five days postpartum. Ruminants 2022, 2, 22. [Google Scholar] [CrossRef]
- Whalin, L.; Weary, D.M.; von Keyserlingk, M.A.G. Understanding behavioural development of calves in natural settings to inform calf management. Animals 2021, 11, 2446. [Google Scholar] [CrossRef] [PubMed]
- Rowan, T.G. Thermoregulation in neonatal ruminants. BSAP Occas. Publ. 1992, 15, 13–24. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Pereira, A.M.F.; Wang, D.; Martínez-Burnes, J.; Ghezzi, M.; Hernández-Avalos, I.; Lendez, P.; Mora-Medina, P.; Casas, A.; Olmos-Hernández, A.; et al. Clinical applications and factors involved in validating thermal windows used in infrared thermography in cattle and river buffalo to assess health and productivity. Animals 2021, 11, 2247. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Napolitano, F.; Braghieri, A.; Guerrero-Legarreta, I.; Bertoni, A.; Martínez-Burnes, J.; Cruz-Monterrosa, R.; Gómez, J.; Ramírez-Bribiesca, E.; Barrios-García, H.; et al. Thermal biology in river buffalo in the humid tropics: Neurophysiological and behavioral responses assessed by infrared thermography. J. Anim. Behav. Biometeorol. 2021, 9, 1–12. [Google Scholar] [CrossRef]
- Lezama-García, K.; Mota-Rojas, D.; Martínez-Burnes, J.; Villanueva-García, D.; Domínguez-Oliva, A.; Gómez-Prado, J.; Mora-Medina, P.; Casas-Alvarado, A.; Olmos-Hernández, A.; Soto, P.; et al. Strategies for hypothermia compensation in altricial and precocial newborn mammals and their monitoring by infrared thermography. Vet. Sci. 2022, 9, 246. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Titto, C.G.; Orihuela, A.; Martínez-Burnes, J.; Gómez-Prado, J.; Torres-Bernal, F.; Flores-Padilla, K.; Carvajal-de la Fuente, V.; Wang, D.; la Fuente, V.C.; et al. Physiological and behavioral mechanisms of thermoregulation in mammals. Animals 2021, 11, 1733. [Google Scholar] [CrossRef] [PubMed]
- Hashmi, H.A.; Tarique, T.M.; Yang, S.; Zubair, M.; Qiu, J.; Chen, G.; Chen, A. Factors affecting mortality in buffaloes and calves. Int. J. Vet. Sci. Med. 2013, 1, 1–9. [Google Scholar]
- Nasr, M. The impact of cross-breeding Egyptian and Italian buffalo on reproductive and productive performance under a subtropical environment. Reprod. Domest. Anim. 2017, 52, 214–220. [Google Scholar] [CrossRef]
- Prasad, S.; Prasad, R.B. Measures of reproductive estimates in rural buffalo herds of Meerut district of Uttar Pradesh (India). Buffalo Bull. 1998, 17, 27–29. [Google Scholar]
- Boro, P.; Bharali, D.; Saharia, J.; Sarma, M.; Sonowal, M.; Brahma, J. Productive and reproductive performances of Murrah buffalo cows: A review. J. Entomol. Zool. Stud. 2020, 8, 290–293. [Google Scholar]
- Mota-Rojas, D.; Wang, D.; Titto, C.G.; Martínez-Burnes, J.; Villanueva-García, D.; Lezama, K.; Domínguez, A.; Hernández-Avalos, I.; Mora-Medina, P.; Verduzco, A.; et al. Neonatal infrared thermography images in the hypothermic ruminant model: Anatomical-morphological-physiological aspects and mechanisms for thermoregulation. Front. Vet. Sci. 2022, 9, 963205. [Google Scholar] [CrossRef]
- Terrien, J.; Perret, M.; Aujard, F. Behavioral thermoregulation in mammals: A review. Front. Biosci. 2011, 16, 1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, A.M.; Shah, A.R.; Hassan, M.F.; Yousif, M.; Wang, Z. Colostrum composition and its importance to the health of animals- a review. Punjab Univ. J. Zool. 2019, 34, 197–206. [Google Scholar] [CrossRef]
- Nagyová, V.; Tóthová, C.; Nagy, O. The impact of colostrum intake on the serum protein electrophoretic pattern in newborn ruminants. J. Appl. Anim. Res. 2017, 45, 498–504. [Google Scholar] [CrossRef] [Green Version]
- Ventorp, M.; Michanek, P. Cow-calf behaviour in relation to first suckling. Res. Vet. Sci. 1991, 51, 6–10. [Google Scholar] [CrossRef]
- Zhang, C.; Juniper, D.T.; McDonald, R.; Parsons, S.; Meagher, R.K. Holstein calves’ preference for potential physical enrichment items on different presentation schedules. J. Dairy Sci. 2022, 105, 8316–8327. [Google Scholar] [CrossRef]
- de Passillé, A.M.B.; Rushen, J. Calves’ behaviour during nursing is affected by feeding motivation and milk availability. Appl. Anim. Behav. Sci. 2006, 101, 264–275. [Google Scholar] [CrossRef]
- Mann, S.; Curone, G.; Chandler, T.L.; Sipka, A.; Cha, J.; Bhawal, R.; Zhang, S. Heat treatment of bovine colostrum: II. Effects on calf serum immunoglobulin, insulin, and IGF-I concentrations, and the serum proteome. J. Dairy Sci. 2020, 103, 9384–9406. [Google Scholar] [CrossRef]
- Abd El-Fattah, A.M.; Abd Rabo, F.H.; EL-Dieb, S.M.; El-Kashef, H.A. Changes in composition of colostrum of Egyptian buffaloes and Holstein cows. BMC Vet. Res. 2012, 8, 19. [Google Scholar] [CrossRef] [Green Version]
- Coroian, A.; Erler, S.; Matea, C.T.; Mireșan, V.; Răducu, C.; Bele, C.; Coroian, C.O. Seasonal changes of buffalo colostrum: Physicochemical parameters, fatty acids andcholesterol variation. Chem. Cent. J. 2013, 7, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huppertz, T.; Upadhyay, A.L.; Kelly, A.L.; Tamime, A.Y. Constituents and properties of milk from different species. In Brined Cheeses; Tamime, A., Ed.; Blackwell Publishing: Oxford, UK, 2006; pp. 1–42. [Google Scholar]
- Dang, A.K.; Kapila, S.; Purohit, M.; Singh, C. Changes in colostrum of Murrah buffaloes after calving. Trop. Anim. Health Prod. 2009, 41, 1213–1217. [Google Scholar] [CrossRef] [PubMed]
- Nowak, R.; Porter, R.H.; Lévy, F.; Orgeur, P.; Schaal, B. Role of mother-young interactions in the survival of offspring in domestic mammals. Rev. Reprod. 2000, 5, 153–163. [Google Scholar] [CrossRef]
- Andriolo, A.; Paranhos, M.J.R.; Costa, D.A.; Schmidek, W.R.; Costa, M.J.R.P.; De Estudos, E.-G. Suckling behaviour in water buffalo suckling behaviour in water buffalo (Bubalus bubalis): Development and individual differences. Rev. Etol. 2001, 3, 129–136. [Google Scholar]
- Ahmad, S.; Yaqoob, M.; Hashmi, N.; Zaman, M.A.; Amjad, M.S. Farmers’ attitude towards interventions regarding buffalo calf health care and management practices under field conditions. Pak. Vet. J. 2009, 29, 125–128. [Google Scholar]
- Besser, T.E.; Gay, C.C.; Pritchett, L. Comparison of three methods of feeding colostrum to dairy calves. J. Am. Vet. Med. Assoc. 1991, 198, 419–422. [Google Scholar]
- Trillmich, F. Parental care: Adjustments to conflict and cooperation. In Animal Behaviour: Evolution and Mechanisms; Kappeler, P., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 267–298. [Google Scholar]
- Lévy, F.; Fleming, A.S. The neurobiology of maternal behavior in mammals. In The Development of Social Engagement Neurobiological Perspectives; Marshall, P., Fox, N., Eds.; Oxford University Press: New York, NY, USA, 2006; pp. 197–246. [Google Scholar]
- Nagasawa, M.; Okabe, S.; Mogi, K.; Kikusui, T. Oxytocin and mutual communication in mother-infant bonding. Front. Hum. Neurosci. 2012, 6, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridges, R.S.; Nephew, B.C. Neuroendocrine control: Maternal behavior. In Encyclopedia of Neuroscience; Elsevier: Amsterdam, The Netherlands, 2009; pp. 333–342. [Google Scholar]
- Bridges, R.S. Neuroendocrine regulation of maternal behavior. Front. Neuroendocrinol. 2015, 36, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Murphey, R.M.; Paranhos da Costa, M.J.R.; Da Silva, R.G.; de Souza, R.C. Allonursing in river buffalo, Bubalus bubalis: Nepotism, incompetence, or thievery? Anim. Behav. 1995, 49, 1611–1616. [Google Scholar] [CrossRef]
- Pyzik, M.; Sand, K.M.K.; Hubbard, J.J.; Andersen, J.T.; Sandlie, I.; Blumberg, R.S. The Neonatal Fc Receptor (FcRn): A Misnomer? Front. Immunol. 2019, 10, 1540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, A.P.; Stabenfeldt, G. La glándula mamaria y la lactación. In Cunningham Fisiología Veterinaria; Klein, B.G., Ed.; Elsevier: Barcelona, España, 2020; pp. 458–470. [Google Scholar]
- Guzmán, V.; Olivera-Angel, M. Calostrogénesis, digestión y absorción del calostro. In La Lactancia, Vista Desde Múltiples Enfoques. Primera Parte: Biología e Inmunología; Olivera-Angel, M., Huertas-Molina, O., Eds.; Biogénesis fondeo editorial: Medellin, Colombia, 2020; pp. 17–29. [Google Scholar]
- Ni, Y.; Chen, Q.; Cai, J.; Xiao, L.; Zhang, J. Three lactation-related hormones: Regulation of hypothalamus-pituitary axis and function on lactation. Mol. Cell. Endocrinol. 2021, 520, 111084. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Idani, G. Suckling and allosuckling behavior in wild giraffe (Giraffa camelopardalis tippelskirchi). Mamm. Biol. 2018, 93, 1–4. [Google Scholar] [CrossRef]
- Skok, J. Suckling strategies in terrestrial ungulates (Artiodactyla, Perissodactyla ) and the factors influencing them: The offspring perspective. CABI Rev. 2022, 17, 1–12. [Google Scholar] [CrossRef]
- Murphey, R.M.; Paranhos da Costa, M.J.R.; de Souza Lima, L.O.; de Moura Duarte, F.A. Communal suckling in water buffalo (Bubalus bubalis). Appl. Anim. Behav. Sci. 1991, 28, 341–352. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.D.F.M.; Quirino, C.R.; Bastos, R. Effect of nursing behaviour, sex of the calf, and parity order on milk production of buffaloes. Rev. Colomb. Ciencias Pecu. 2017, 30, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Gračner, D.; Gilligan, G.; Garvey, N.; Moreira, L.; Harvey, P.; Tierney, A.; ZobeL, R. Correlation between the milk vein internal diameter surface andmilk yield in Simmental cows. Turkish J. Vet. Anim. Sci. 2015, 39, 741–744. [Google Scholar] [CrossRef]
- Paranhos Da Costa, M.J.R.; Andriolo, A.; Simplício De Oliveira, J.F.; Schmidek, W.R. Suckling and allosuckling in river buffalo calves and its relation with weight gain. Appl. Anim. Behav. Sci. 2000, 66, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lovarelli, D.; Bacenetti, J.; Guarino, M. A review on dairy cattle farming: Is precision livestock farming the compromise for an environmental, economic and social sustainable production? J. Clean. Prod. 2020, 262, 121409. [Google Scholar] [CrossRef]
- Edwards, E.M.; Krawczel, P.D.; Dann, H.M.; Schneider, L.G.; Whitlock, B.; Proudfoot, K.L. Calving location preference and changes in lying and exploratory behavior of preparturient dairy cattle with access to pasture. J. Dairy Sci. 2020, 103, 5455–5465. [Google Scholar] [CrossRef] [PubMed]
- Meo Zilio, D.; Steri, R.; Iacurto, M.; Catillo, G.; Barile, V.; Chiariotti, A.; Cenci, F.; La Mantia, M.C.; Buttazzoni, L. Precision livestock farming for mediterranean water buffalo: Some applications and opportunities from the agridigit project. In Safety, Health and Welfare in Agriculture and Agro-Food Systems; Biocca, M., Cavallo, E., Cecchini, M., Failla, S., Romano, E., Eds.; Springer: Cham, Switzerland, 2022; pp. 41–50. [Google Scholar]
- Vonnahme, K.A.; Lemley, C.O. Programming the offspring through altered uteroplacental hemodynamics: How maternal environment impacts uterine and umbilical blood flow in cattle, sheep and pigs. Reprod. Fertil. Dev. 2012, 24, 97. [Google Scholar] [CrossRef]
- Derar, R.; Hussein, H.; Fahmy, S.; El-Sherry, T.; Megahed, G. The effect of parity on the efficacy of an ovulation synchronization (Ovsynch) protocol in buffalo (Bubalus bubalis). Anim. Reprod. 2012, 9, 52–60. [Google Scholar]
- Nava-Trujillo, H.; Valeris-Chacin, R.; Quintero-Moreno, A.; Escalona-Muñoz, J. Milk yield at first lactation, parity, and season of calving affect the reproductive performance of water buffalo cows. Anim. Prod. Sci. 2020, 60, 1073. [Google Scholar] [CrossRef]
- Meurisse, M.; Gonzalez, A.; Delsol, G.; Caba, M.; Lévy, F.; Poindron, P. Estradiol receptor-α expression in hypothalamic and limbic regions of ewes is influenced by physiological state and maternal experience. Horm. Behav. 2005, 48, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Vandenheede, M.; Nicks, B.; Désiron, A.; Canart, B. Mother–young relationships in Belgian Blue cattle after a Caesarean section: Characterisation and effects of parity. Appl. Anim. Behav. Sci. 2001, 72, 281–292. [Google Scholar] [CrossRef]
- Numan, M.; Young, L.J. Neural mechanisms of mother–infant bonding and pair bonding: Similarities, differences, and broader implications. Horm. Behav. 2016, 77, 98–112. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, H.; Serafin, N.; Terrazas, A.M.; Marnet, P.G.; Kann, G.; Delgadillo, J.A.; Poindron, P. Maternal Olfaction Differentially Modulates Oxytocin and Prolactin Release during Suckling in Goats. Horm. Behav. 2002, 42, 232–244. [Google Scholar] [CrossRef]
- Derar, D.R.; Abdel-Rahman, M.A. A comparative study on behavioral, physiological and andrenal changes in blood cortisol of buffalo during actual labour. Buffalo Bull. 2012, 31, 46–50. [Google Scholar]
- Zipp, K.A.; Barth, K.; Rommelfanger, E.; Knierim, U. Responses of dams versus non-nursing cows to machine milking in terms of milk performance, behaviour and heart rate with and without additional acoustic, olfactory or manual stimulation. Appl. Anim. Behav. Sci. 2018, 204, 10–17. [Google Scholar] [CrossRef]
- MacLeod, K.J.; Clutton-Brock, T.H. Low costs of allonursing in meerkats: Mitigation by behavioral change? Behav. Ecol. 2015, 26, 697–705. [Google Scholar] [CrossRef] [Green Version]
- Zapata, B.; Gaete, G.; Correa, L.A.; González, B.A.; Ebensperger, L.A. A case of allosuckling in wild guanacos (Lama guanicoe). J. Ethol. 2009, 27, 295–297. [Google Scholar] [CrossRef]
- Becker, M.I.; De Ioannes, A.E.; León, C.; Ebensperger, L.A. Females of the communally breeding rodent, Octodon degus, transfer antibodies to their offspring during pregnancy and lactation. J. Reprod. Immunol. 2007, 74, 68–77. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mota-Rojas, D.; Bragaglio, A.; Braghieri, A.; Napolitano, F.; Domínguez-Oliva, A.; Mora-Medina, P.; Álvarez-Macías, A.; De Rosa, G.; Pacelli, C.; José, N.; et al. Dairy Buffalo Behavior: Calving, Imprinting and Allosuckling. Animals 2022, 12, 2899. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12212899
Mota-Rojas D, Bragaglio A, Braghieri A, Napolitano F, Domínguez-Oliva A, Mora-Medina P, Álvarez-Macías A, De Rosa G, Pacelli C, José N, et al. Dairy Buffalo Behavior: Calving, Imprinting and Allosuckling. Animals. 2022; 12(21):2899. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12212899
Chicago/Turabian StyleMota-Rojas, Daniel, Andrea Bragaglio, Ada Braghieri, Fabio Napolitano, Adriana Domínguez-Oliva, Patricia Mora-Medina, Adolfo Álvarez-Macías, Giuseppe De Rosa, Corrado Pacelli, Nancy José, and et al. 2022. "Dairy Buffalo Behavior: Calving, Imprinting and Allosuckling" Animals 12, no. 21: 2899. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12212899