
Comparative Histology of the Cornea and Palisades of Vogt in the Different Wild Ruminants (Bovidae, Camelidae, Cervidae, Giraffidae, Tragulidae)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Specimen and Conservation Status
2.2. Ethical Statement
2.3. Histological Study
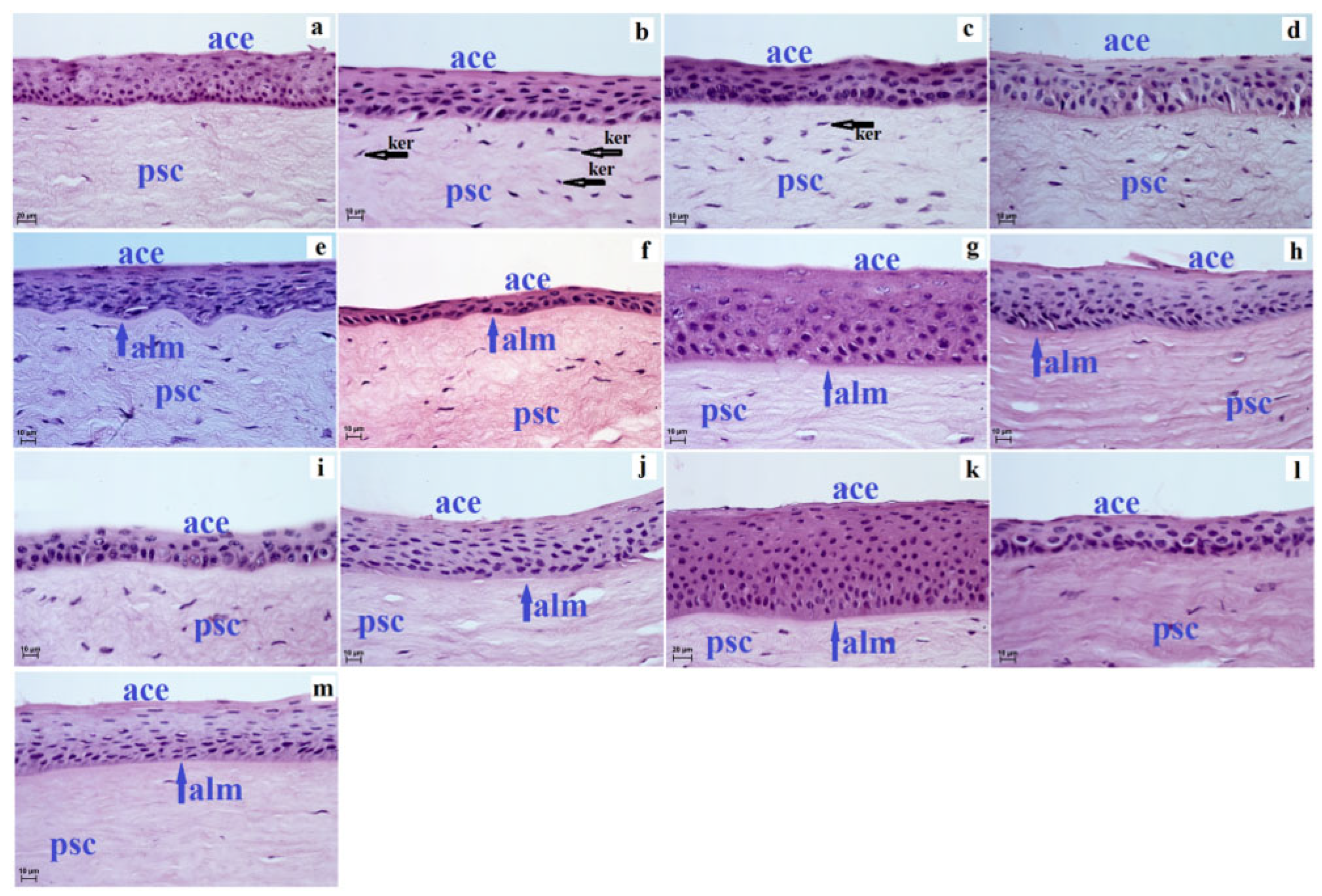
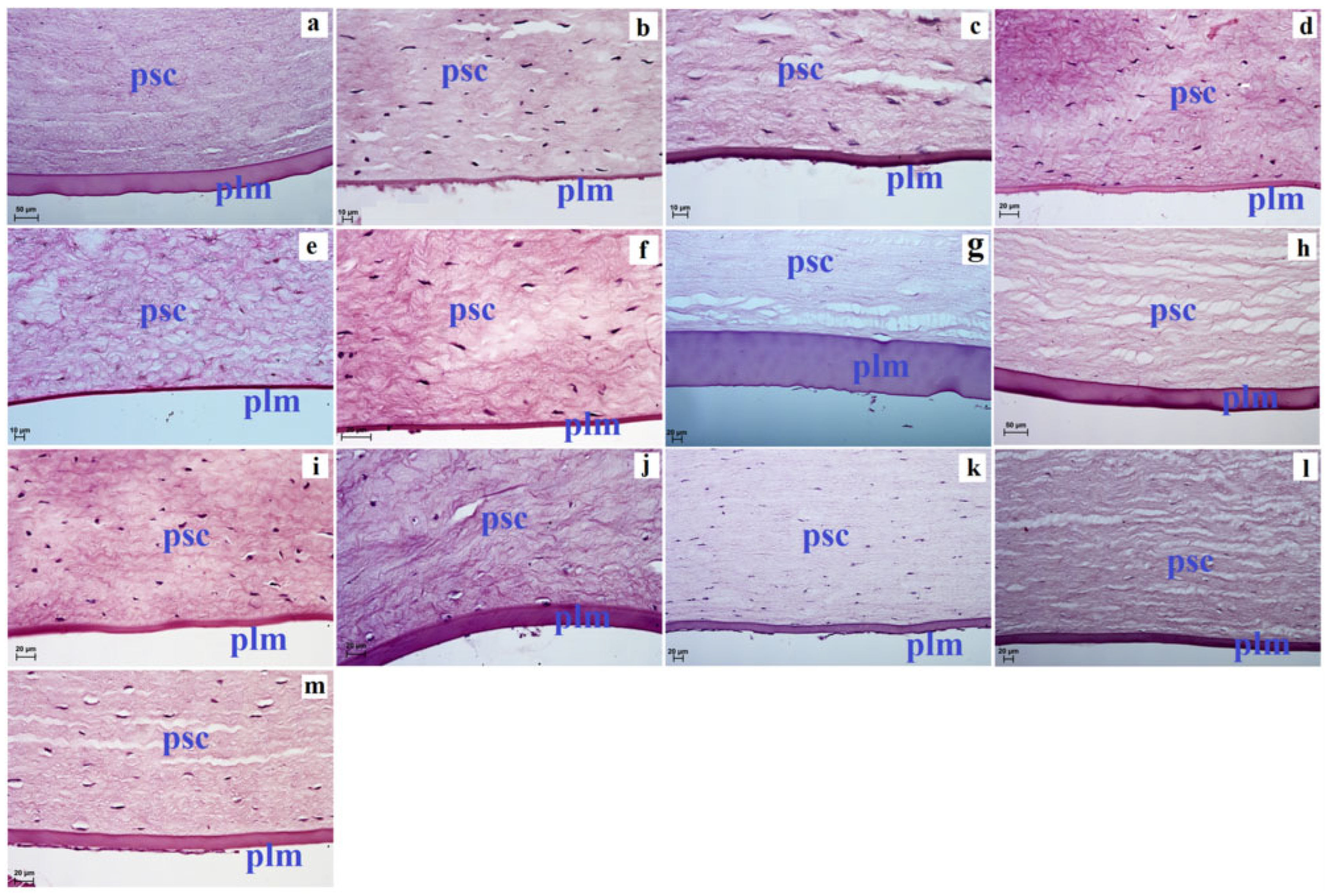
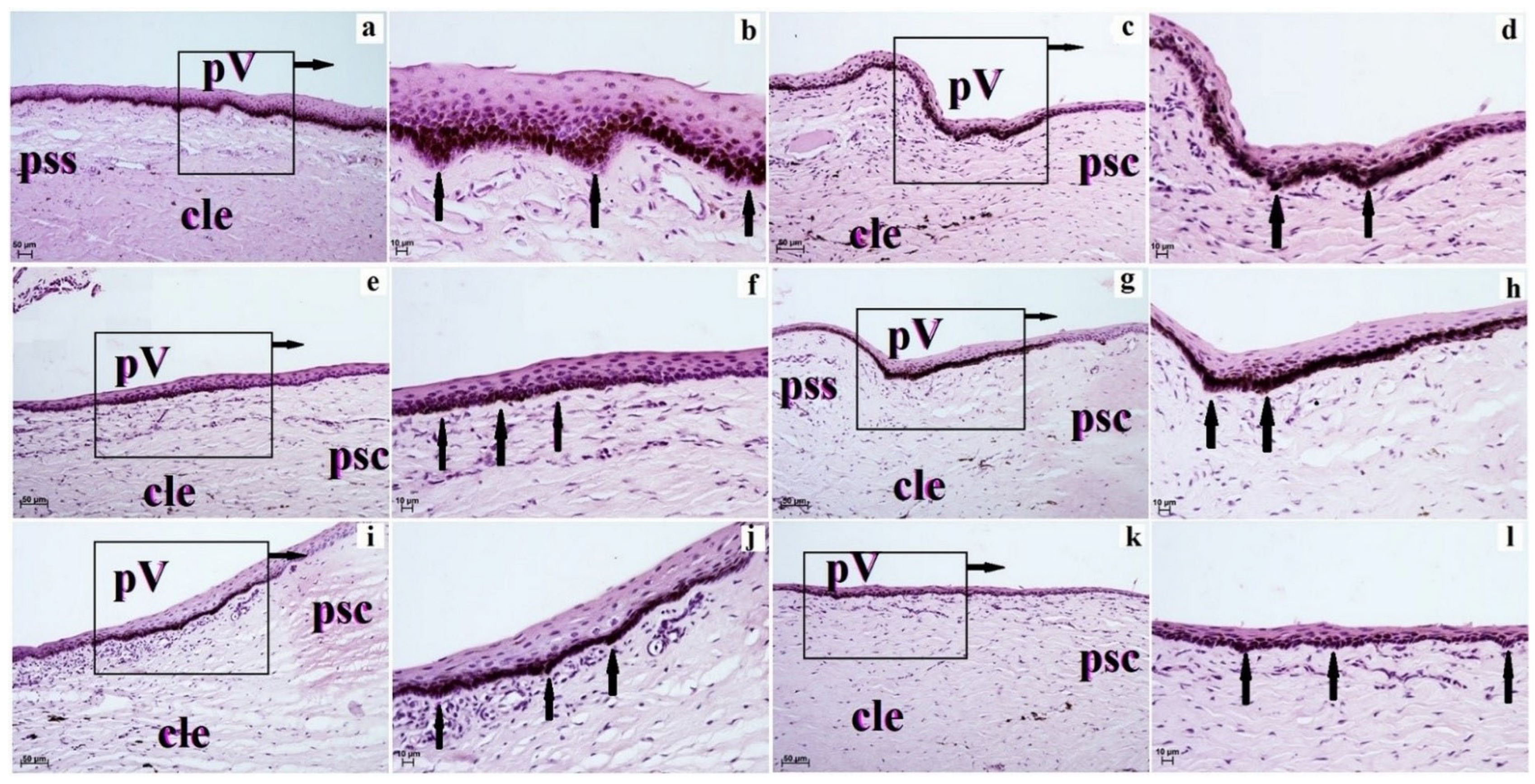
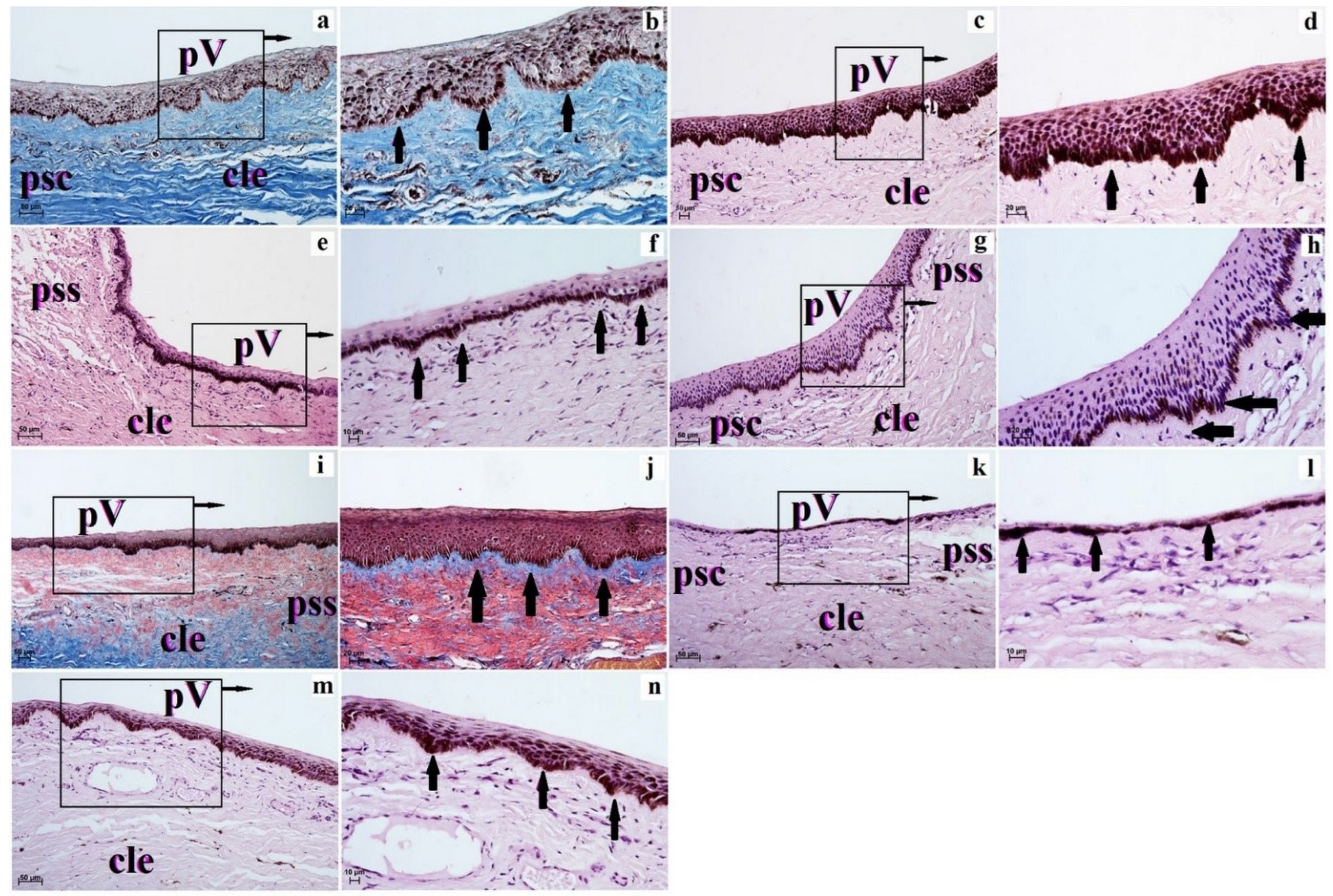
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Almubrad, T.; Akhtar, S. Ultrastructure features of camel cornea—Collagen fibril and proteoglycans. Vet. Ophthalmol. 2012, 15, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Doughty, M.J. A physiological perspective on the swelling properties of the mammalian corneal stroma. Contact Lens Anterior Eye 2003, 26, 117–129. [Google Scholar] [CrossRef]
- Nautscher, N.; Bauer, A.; Steffl, M.; Amselgruber, W.M. Comparative morphological evaluation of domestic animal cornea. Vet. Ophthalmol. 2016, 19, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, K. Histology; PZWL: Warszawa, Poland, 1988. [Google Scholar]
- Marciniak, T.; Ziółkowski, M. Anatomia Prawidłowa Człowieka; RU ZSP AM: Wrocław, Poland, 1992. [Google Scholar]
- Meek, K.M.; Fullwood, N.J. Corneal and scleral collagens—A microscopist’s perspective. Micron 2001, 32, 261–272. [Google Scholar] [CrossRef]
- Czaińska, M. Astygmatyzm—Charakterystyka wady. Optyka 2016, 5, 28–31. [Google Scholar]
- Roszkowska, A.M. Aktualności w diagnostyce i leczeniu stożka rogówki. Okul. Dyplomie 2013, 4, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Szaflik, J.; Izdebska, J. Choroby Aparatu Cchronnego Oka i Rogówki; American Academy of Ophthalmology: Wrocław, Poland, 2019. [Google Scholar]
- Maurice, D.M. The structure and transparency of the cornea. J. Physiol. 1957, 136, 263–286. [Google Scholar] [CrossRef]
- Pedler, C. The fine structure of the corneal epithelium. Exp. Eye Res. 1962, 1, 286–289. [Google Scholar] [CrossRef]
- Merindano, M.D.; Costa, J.; Canals, M.; Potau, J.M.; Ruano, D. A comparative study of Bowman’s layer in some mammals: Relationships with other constituent corneal structures. Eur. J. Anat. 2002, 6, 133–139. [Google Scholar]
- Dua, H.S.; Faraj, L.A.; Said, D.G.; Gray, T.; Lowe, J. Human corneal anatomy redefined: A novel pre-Descemet’s layer (Dua’s layer). Ophthalmol. 2013, 120, 1778–1785. [Google Scholar] [CrossRef]
- Koçluk, Y.; Burcu, A.; Sukgen, E.A. Demonstration of cornea Dua’s layer at a deep anterior lamellar keratoplasty surgery. Oman J. Ophthalmol. 2016, 9, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ederra, J.; Garcıa, M.; Hernandez, M.; Urcola, H.; Hernández-Barbáchano, E.; Araiz, J.; Vecino, E. The pig eye as a novel model of glaucoma. Exp. Eye Res. 2005, 81, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Brunette, I.; Rosolen, S.G.; Carrier, M.; Abderrahman, M.; Nada, O.; Germain, L.; Proulx, S. Comparison of the pig and feline models for full thickness corneal transplantation. Vet. Ophthalmol. 2011, 14, 365–377. [Google Scholar] [CrossRef]
- Cafaro, T.A.; Suarez, M.F.; Maldonado, C.; Croxatto, J.O.; Insfran, C.; Urrets-Zavalıa, J.A.; Serra, H.M. On the Cornea of Healthy Merino Sheep: A Detailed Ex Vivo Confocal, Histological and Ultrastructural Study. Anat. Histol. Embryol. 2015, 44, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Calmettes, L.; Deodati, F.; Pianel, H.; Bec, C. Etude Histologque de L’Epithelium Anterieure de a1 Cornee et de ses Basales. Arch. Ophtal. Paris 1956, 16, 481–506. [Google Scholar]
- Ehlers, N. Some Comparative Studies on the mammalian corneal epithelium. Acta Ophthalmol. 1970, 48, 821–828. [Google Scholar]
- Lee, R.E.; Davison, P.F. Collagen composition and turnover in ocular tissues of the rabbit. Exp. Eye Res. 1984, 32, 737–745. [Google Scholar] [CrossRef]
- Merindano, M.D.; Canals, M.; Potau, J.M.; Costa, J.; Ruan, D. Morphometrical features of the corneal epithelium in mammals. Anat. Histol. Embryol. 1998, 27, 105–110. [Google Scholar] [CrossRef]
- Rahi, A.H.S.; Sheikh, H.; Morgan, G. Histology of the camel eye. Acta Anat. 1980, 106, 345–350. [Google Scholar] [CrossRef]
- Rehbinder, C.; Winquist, G.; Roos, C. Structure of the cornea in some cervidae. Acta Vet. Scand. 1977, 18, 152–158. [Google Scholar] [CrossRef]
- Reichard, M.; Hovakimyan, M.; Wree, A.; Meyer-Lindenberg, A.; Nolte, I.; Junghans, C.; Guthoff, R.; Stachs, O. Comparative in vivo confocal microscopical study of the cornea anatomy of different laboratory animals. Curr. Eye Res. 2010, 35, 1072–1080. [Google Scholar] [CrossRef] [PubMed]
- Winquist, G.; Rehbinder, C. Fine structure of the reindeer cornea in normal conditions and in keratitis. Acta Vet. Scand. 1973, 14, 292–300. [Google Scholar] [CrossRef]
- Sanchez, R.F.; Daniels, J.T. Mini-Review: Limbal stem cells deficiency in companion animals: Time to give something back? Curr. Eye Res. 2016, 41, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Dua, H.S. The conjunctiva in corneal epithelial wound healing. Br. J. Ophthalmol. 1998, 82, 1407–1411. [Google Scholar] [CrossRef]
- Dua, H.S.; Saini, J.S.; Azuara-Blanco, A.; Gupta, P. Limbal stem cell deficiency: Concept, aetiology, clinical presentation, diagnosis and management. Indian J. Ophthalmol. 2000, 48, 83–92. [Google Scholar]
- Kenyon, K.R.; Tseng, S.C.G. Limbal autograft transplantation for ocular surface disorder. Ophthalmology 1989, 96, 709–723. [Google Scholar] [PubMed]
- Kruse, F.E. Classification of ocular surface disease. In Ocular Surface Disease. Medical and Surgical Management; Holland, E.J., Mannis, M.J., Eds.; Springer: New York, NY, USA, 2001. [Google Scholar]
- IUCN Red List of Threatened Species. 2022. Available online: https://www.iucnredlist.org (accessed on 23 September 2022).
- Burck, N.C. Technika Histologiczna; PZWL: Warszawa, Poland, 1975. [Google Scholar]
- Movat, H.Z. Demonstration of all connective tissue elements in a single section. AMA Arch. Pathol. 1955, 60, 289. [Google Scholar]
- International Committee on Veterinary Gross Anatomical Nomenclature (ICVGAN). Nomina Anatomica Veterinaria, 6th ed.; Editoral Committee: Hanover, Germany; Ghent, Belgium; Columbia, MO, USA; Rio de Janeiro, Brazil, 2017. [Google Scholar]
- Nomina Histologica Veterinaria. Submitted by the International Committee on Veterinary Histological Nomenclature (ICVHN) to the World Association Veterinary Anatomists. 2017. Available online: https://www.wava-amav.org/downloads/NHV_2017.pdf (accessed on 13 October 2022).
- Ang, M.; Konstantopoulos, A.; Goh, G.; Htoon, H.M.; Seah, X.; Lwin, N.C.; Liu, X.; Chen, S.; Liu, L.; Mehta, J.S. Evaluation of a micro-optical coherence tomography for the corneal endothelium in an animal model. Sci. Rep. 2016, 6, 29769. [Google Scholar] [CrossRef] [Green Version]
- Ibares-Fríasa, L.; Gallego, P.; Cantalapiedra-Rodriguezc, R.; Merayo-Llovesa, J.; Martínez-García, M.C. Clinical, refractive and histological reversibility of corneal additive surgery in deep stroma in an animal model. Curr. Eye Res. 2016, 41, 1192–1201. [Google Scholar] [CrossRef] [Green Version]
- Kethiria, A.R.; Rajua, E.; Bokarac, K.K.; Mishrad, D.K.; Basua, S.; Raoc, C.M.; Sangwana, V.S.; Singha, V. Inflammation, vascularization and goblet cell differences in LSCD: Validating animal models of corneal alkali burns. Exp. Eye Res. 2019, 185, 1–8. [Google Scholar] [CrossRef]
- Kim, W.-J.; Marco, C.H.; Mohan, R.R.; Wilson, E.E. Changes in corneal morphology associated with chronic epithelial injury. IOVS 1999, 40, 35–42. [Google Scholar]
- Rodriguez-Ramos, F.J.; Dubielzig, R.R. Ocular comparative anatomy of the family Rodentia. Vet. Ophthalmol. 2013, 16, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Sitalakshmi, G.; Sudha, B.; Madhavan, H.N.; Vinay, S.; Krishnakumar, S.; Mori, Y.; Yoshioka, H.; Abraham, S. Ex vivo cultivation of corneal limbal epithelial cells in a thermoreversible Polymer (Mebiol Gel) and Their Transplantation in Rabbits: An Animal Model. Tissue Eng. 2009, 15, 407–415. [Google Scholar] [CrossRef]
- Buck, R.C. Ultrastructural characteristics associated with the anchoring of corneal epithelium in several classes of vertebrates. J. Anat. 1983, 137, 743–756. [Google Scholar]
- Gronkiewicz, K.M.; Giuliano, E.A.; Kuroki, K.; Bunyak, F.; Sharma, A.; Teixeira, L.B.C.; Hamm, C.W.; Mohan, R.R. Development of a novel in vivo corneal fibrosis model in the dog. Exp. Eye Res. 2016, 14375, e88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marlo, T.L.; Giuliano, E.A.; Sharma, A.; Mohan, R.R. Development of a novel ex vivo equine corneal model. Vet. Ophthalmol. 2017, 20, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Napoli, P.E.; Nioi, M.; d’Aloja, E.; Loy, F.; Fossarello, M. The architecture of corneal stromal striae on optical coherence tomography and histology in an animal model and in humans. Sci. Rep. 2020, 10, 19861. [Google Scholar] [CrossRef]
- Proulx, S.; Bensaoula, T.; Nada, O.; Audet, C.; d’Arc Uwamaliya, J.; Devaux, A.; Allaire, G.; Germain, L.; Brunette, I. Transplantation of a Tissue-Engineered Corneal Endothelium Reconstructed on a Devitalized Carrier in the Feline Model. IOVS 2009, 50, 2686–2694. [Google Scholar] [CrossRef]
- Subasinghe, S.K.; Ogbuehi, K.C.; Mitchell, L.; Dias, G.J. Animal model with structural similarity to human corneal collagen fibrillar arrangement. Anat. Sci. Int. 2021, 96, 286–293. [Google Scholar] [CrossRef]
- Jacobsen, I.E.; Jensen, O.A.; Prause, J.U. Structure and composition of Bowman‘s membrane: Study by frozen resin cracking. Acta Ophthalmol. 1984, 62, 39–53. [Google Scholar] [CrossRef]
- Germundsson, J.; Fagerholm, P.; Koulikovska, M.; Lagali, N.S. An accurate method to determine Bowman’s layer thickness in vivo in the human cornea. IOVS 2012, 53, 2354–2359. [Google Scholar]
- Wislocki, G.B. The anterior segment of the eye of the rhesus monkey investigated by histochemical means. Am. J. Anat. 1951, 91, 233–261. [Google Scholar] [CrossRef] [PubMed]
- Abrams, G.A.; Bentley, E.; Nealey, P.F.; Murphy, C.J. Electron microscopy of the canine corneal basement membranes. Cells Tissues Organs 2002, 170, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Ledbetter, E.C.; Scarlett, J.M. In vivo confocal microscopy of the normal equine cornea and limbus. Vet. Ophthalmol. 2009, 12, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Gwin, R.M.; Lerner, I.; Warren, J.K.; Gum, G. Decrease in canine endothelial cell density and increase in corneal thickness with age. IOVS 1982, 22, 267–271. [Google Scholar]
- Lavker, R.M.; Tseng, S.C.G.; Sun, T.T. Corneal epithelial stem cells at the limbus: Looking at some old problems from a new angle. Exp. Eye Res. 2004, 78, 433–446. [Google Scholar] [CrossRef]
- Patruno, M.; Perazzi, A.; Martinello, T.; Blaseotto, A.; Di Iorio, E.; Iacopetti, I. Morphological description of limbal epithelium: Searching for steam cells crypts in the dog, cat, pig, cow, sheep and horse. Vet. Res. Commun. 2017, 41, 169–173. [Google Scholar] [CrossRef]
- Schermer, A.; Galvin, S.; Sun, T.T. Differentiation-related expression of a major 64K corneal keratin in vivo and in culture suggests limbal location of corneal epithelial stem cells. J. Cell Biol. 1986, 103, 49–62. [Google Scholar] [CrossRef]
- Di Iorio, E.; Barbaro, V.; Ruzza, A.; Ponzin, D.; Pellegrini, G.; De Luca, M. Isoforms of DeltaNp63 and the migration of ocular limbal cells in human corneal regeneration. Proc. Natl. Acad, Sci. USA 2005, 102, 9523–9528. [Google Scholar] [CrossRef] [Green Version]
- Haagdorens, M.; Behaegel, J.; Rozema, J.; Van Gerwen, V.; Michiels, S.; Dhubhghaill, S.N.; Tassignon, M.-J.; Zakaria, N. A method for quantifying limbal stem cell niches using OCT imaging. Br. J. Ophthalmol. 2017, 101, 1250–1255. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Infraorder | Family | Subfamily | Tribus | Genus | Species/ Subspecies | IUCN (2022-1) | Number of Collection/ Date of Collection | Source of Collection |
|---|---|---|---|---|---|---|---|---|
| Pecora | Bovidae | Antilopinae | Alcelaphini | Connochaetes | common wildebeest Connochaetes taurinus | LC stable | 2 /2017 | Wroclaw Zoological Garden |
| Antilopini | Madoqua | Kirk’s dik-dik Madoqua kirkii | LC stable | 7 /2016, 2017, 2018, 2019, 2021 | Wroclaw Zoological Garden | |||
| Cephalophini | Cephalophus | Natal red duiker Cephalophus natalensis | LC decreasing | 6 /2016, 2017, 2020, 2021 | Wroclaw Zoological Garden | |||
| Hippotragini | Oryx | scimitar oryx Oryx dammah | EW | 2 /2017 | Wroclaw Zoological Garden | |||
| Bovinae | Tragelaphini | Tragelaphus | sitatunga Tragelaphus spekii | LC decreasing | 11 /2016, 2017, 2018, 2019 | Wroclaw Zoological Garden | ||
| Cervidae | Cervinae | Cervini | Rusa | Philippine spotted deer Rusa alfredi | EN decreasing | 1 /2016 | Wroclaw Zoological Garden | |
| Elaphurus | Père David’s deer Elaphurus davidianus | EW | 1 /2017 | Wroclaw Zoological Garden | ||||
| Capreolinae | Alceini | Alces | moose Alces alces | LC increasing | 1 /2017 | Warsaw Zoological Garden | ||
| Odocoileini | Rangifer | reindeer Rangifer tarandus | VU decreasing | 3 /2018, 2019 | Wroclaw Zoological Garden | |||
| Giraffidae | Giraffinae | – | Giraffa | reticulated giraffe Giraffa Camelopardalis reticulata | VU decreasing | 1 /2016 | Wroclaw Zoological Garden | |
| – | Okapia | okapi Okapia johnstoni | EN decreasing | 1 /2017 | Wroclaw Zoological Garden | |||
| Tragulina | Tragulidae | – | – | Tragulus | Balabac mouse-deer Tragulus nigricans | EN decreasing | 8 /2016, 2018, 2019, 2022 | Wroclaw Zoological Garden |
| Cameliformes | Camelidae | – | Lamini | Vicunia pacos | alpaca Vicugna pacos | LC MD increasing | 6 /2013, 2014 | Wroclaw Zoological Garden, Own collection of Division of Animal Anatomy |
| Anterior Corneal Epithelium | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Number of Cellular Layers in the Central Cornea | Number of the Superficial Cell Layers | Number of the Intermediate Cell Layers | Number of the Basal Cell Layers | Thickness of the Central Part | Number of Cellular Layers in the Peripheral Cornea | Number of the Superficial Cell Layers | Number of the Intermediate Cell Layers | Number of the Basal Cell Layers | Thickness of the Peripheral Part | |
| common wildebeest | 8 – 9 | 3 | 4 – 5 | 1 | 47.493 ( ± 4.9) | 7 – 8 | 1 – 2 | 5 | 1 | 35.948 ( ± 4.1) |
| Kirk’s dik-dik | 6 – 7 | 2 – 3 | 3 | 1 | 28.873 ( ± 3.2) | 6 – 7 | 2 – 3 | 3 | 1 | 29.596 ( ± 4.6) |
| Natal red duiker | 5 – 6 | 1 | 3 | 2 | 35.208 ( ± 3.3) | 3 – 4 | 1 | 1 – 2 | 1 | 19.255 ( ± 2.2) |
| scimitar oryx | 5 – 6 | 2 – 3 | 2 | 1 | 35.606 ( ± 3.2) | 5 – 6 | 2 – 3 | 2 | 1 | 35.577 ( ± 4.4) |
| sitatunga | 8 – 9 | 3 – 4 | 2 – 5 | 1 | 51.218 ( ± 5.7) | 9 – 10 | 1 | 6–7 | 3 | 42.712 (±4.2) |
| Philippine spotted deer | 3 – 4 | 1 – 3 | 1 | 1 | 15.456 ( ± 2.2) | 6 – 7 | 3 | 1 – 2 | 1 – 2 | 16.891 ( ± 2.7) |
| Père David’s deer | 12 – 13 | 1 – 2 | 7 – 8 | 2 – 3 | 80.99 ( ± 1.8) | 11 – 12 | 1 – 2 | 6 – 7 | 2 – 3 | 74.163 ( ± 3.7) |
| moose | 10 – 11 | 1 – 2 | 7 – 8 | 1 | 42.073 ( ± 1.3) | 14 – 15 | 1 – 3 | 12 | 1 – 2 | 75.63 ( ± 3.0) |
| reindeer | 7 – 8 | 1 – 2 | 5 – 6 | 1 | 33.461 ( ± 3.1) | 4 – 5 | 1 | 2 – 3 | 1 | 21.712 ( ± 3.4) |
| reticulated giraffe | 9 – 10 | 2 – 3 | 6 – 7 | 1 | 42.93 ( ± 3.1) | 10 – 11 | 2 – 3 | 6 – 7 | 2 | 42.072 ( ± 3.4) |
| okapi | 15 – 16 | 2 | 11 – 12 | 1 – 2 | 130.315 ( ± 7.6) | 14 – 15 | 2–3 | 10 | 2 | 115.923 ( ± 6.9) |
| Balabac mouse-deer | 4 – 5 | 1 – 2 | 2 | 1 | 24.778 ( ± 3.8) | 3 – 4 | 1 | 1 – 2 | 1 | 13.127 ( ± 0.9) |
| alpaca | 7 – 8 | 2 – 3 | 4 – 5 | 1 | 50.563 ( ± 3.5) | 13 – 14 | 1 – 2 | 9 | 2 – 3 | 55.651 ( ± 4.1) |
| Species | Bowman’s Layer | Proper Substance of Cornea | Descemet’s Membrane |
|---|---|---|---|
| common wildebeest | – | 1461.735 (±57.7) | 43.181 (±2.3) |
| Kirk’s dik-dik | – | 651.772 (±19.4) | 4.563 (±0.8) |
| Natal red duiker | – | 669.987 (±34.9) | 8.097 (±0.6) |
| scimitar oryx | – | 697.172 (±17.7) | 4.487 (±1.1) |
| sitatunga | 4.04 (±0.5) | 885.483 (±20.5) | 5.028 (±0.8) |
| Philippine spotted deer | 2.5 (±0.4) | 645.331 (±14.6) | 4.32 (±0.4) |
| Père David’s deer | 8.401 (±1.1) | 1347.154 (±30.7) | 118.571 (±3.3) |
| moose | 3.286 (±0.7) | 1668.633 (±36.5) | 47.366 (±2.3) |
| reindeer | – | 869.367 (±33.1) | 7.14 (±0.6) |
| reticulated giraffe | 5.055 (±0.7) | 1971.645 (±194.1) | 20.387 (±2.1) |
| okapi | 4.691 (±0.9) | 1417.026 (±46.1) | 27.661 (±1.3) |
| Balabac mouse-deer | – | 571.808 (±47.5) | 19.458 (±1.2) |
| alpaca | 4.295 (±1.1) | 679.217 (±19.2) | 14.922 (±0.7) |
| Corneal Limbus Epithelium | |||||
|---|---|---|---|---|---|
| Total Number of Cell Layers | Number of Superficial Cell Layers | Number of Intermediate Cell Layers | Number of Basal Cell Layers | Obecność Melanocytes | |
| common wildebeest | 15–16 | 3 | 3–4–6 | 6–7 | +++ (basal cell layers) |
| Kirk’s dik-dik | 7–8 | 3 | 2–3 | 2–3 | + (all cell layers) |
| Natal red duiker | 7–9 | 1–2 | 4 | 2–3 | + (basal cell layers) |
| scimitar oryx | 7–8 | 2–3 | 3 | 2 | ++ (all cell layers) |
| sitatunga | 15–19–20 | 1 | 13–16 | 1–2–3 | + (all cell layers) |
| Philippine spotted deer | 4–6 | 1–2 | 2–3 | 1 | −/+ (all cell layers) |
| Père David’s deer | 17–18 | 3–4 | 12–13 | 1–2 | ++ (intermediate and basal cell layers) |
| moose | 17 –18 | 2–3 | 13 | 2 | +++ (all cell layers) |
| reindeer | 8–9 | 1 | 5–6 | 2 | + (basal cell layers) |
| reticulated giraffe | 17–18 | 2 | 14–15 | 1–2 | −/+ (intermediate and basal cell layers) |
| okapi | 19–20 | 1–2 | 13–16 | 1–2 | −/+ (basal cell layers) |
| Balabac mouse-deer | 3–4 | 1 | 1 | 1–2 | + (all cell layers) |
| alpaca | 13–14 | 1 | 10–11 | 2–3 | – |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klećkowska-Nawrot, J.; Goździewska-Harłajczuk, K.; Barszcz, K. Comparative Histology of the Cornea and Palisades of Vogt in the Different Wild Ruminants (Bovidae, Camelidae, Cervidae, Giraffidae, Tragulidae). Animals 2022, 12, 3188. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12223188
Klećkowska-Nawrot J, Goździewska-Harłajczuk K, Barszcz K. Comparative Histology of the Cornea and Palisades of Vogt in the Different Wild Ruminants (Bovidae, Camelidae, Cervidae, Giraffidae, Tragulidae). Animals. 2022; 12(22):3188. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12223188
Chicago/Turabian StyleKlećkowska-Nawrot, Joanna, Karolina Goździewska-Harłajczuk, and Karolina Barszcz. 2022. "Comparative Histology of the Cornea and Palisades of Vogt in the Different Wild Ruminants (Bovidae, Camelidae, Cervidae, Giraffidae, Tragulidae)" Animals 12, no. 22: 3188. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12223188