
Gut Microbiome Studies in Livestock: Achievements, Challenges, and Perspectives
by
 , and
, and
Giovanni Forcina
1,2,3,*,
Lucía Pérez-Pardal
1,2,
Júlio Carvalheira
1,2,4 and
Albano Beja-Pereira
1,2,5,6,* 1
CIBIO, Centro de Investigação em Biodiversidade e Recursos Genéticos, InBIO Laboratório Associado, Campus de Vairão, Universidade do Porto, 4485-661 Vairão, Portugal
2
BIOPOLIS Program in Genomics, Biodiversity and Land Planning, CIBIO, Campus de Vairão, Universidade do Porto, 4485-661 Vairão, Portugal
3
Universidad de Alcalá, Global Change Ecology and Evolution Research Group (GloCEE), Departamento de Ciencias de la Vida, 28805 Alcalá de Henares, Spain
4
Abel Salazar Institute of Biomedical Sciences, University of Porto, Rua de Jorge Viterbo Ferreira 228, 4050-313 Porto, Portugal
5
DGAOT, Faculty of Sciences, Universidade do Porto, Rua Campo Alegre 687, 4169-007 Porto, Portugal
6
Sustainable Agrifood Production Research Centre (GreenUPorto), Universidade do Porto, Rua da Agrária 747, 4485-646 Vairão, Portugal
*
Authors to whom correspondence should be addressed.
Animals 2022, 12(23), 3375; https://0-doi-org.brum.beds.ac.uk/10.3390/ani12233375
Submission received: 13 September 2022
/
Revised: 16 November 2022
/
Accepted: 28 November 2022
/
Published: 30 November 2022
(This article belongs to the Special Issue New Tools for Monitoring Genetic Diversity in Animals)
{kind=link}
{kind=link}
Abstract
:Simple Summary
The relentless capacity of sequencing every bit of DNA at low cost has been fueling major advances in several research areas. This also applies to the animal sciences, which witnessed unprecedented progresses in fields such as animal nutrition, health, and breeding. Particular attention has been paid to the gut microbiome, the community of microorganisms inhabiting the digestive tract of livestock species, and unforeseen developments have arisen. Nonetheless, such efforts have not been equal for the different livestock species, and the vast majority rely on widely-used standard techniques through which taxonomically useful genetic data are generated rather than more informative—yet computationally demanding—organismal genome-wide variation data. This review offers a glimpse of the gut microbiome research on five emblematic livestock species touching on the limitations regarding (i) the major methodological frameworks, (ii) species or breed, (iii) and spatial reach of these studies, thus providing valuable indications to fill current knowledge gaps and hopefully lay the basis for the planning of concerted research efforts. In this respect, we conclude that future studies should extend shotgun sequencing and transcriptomic approaches primarily to largely neglected ovicaprine and chicken breeds from rural areas of developing countries and microbial groups other than bacteria.
Abstract
The variety and makeup of the gut microbiome are frequently regarded as the primary determinants of health and production performances in domestic animals. High-throughput DNA/RNA sequencing techniques (NGS) have recently gained popularity and permitted previously unheard-of advancements in the study of gut microbiota, particularly for determining the taxonomic composition of such complex communities. Here, we summarize the existing body of knowledge on livestock gut microbiome, discuss the state-of-the-art in sequencing techniques, and offer predictions for next research. We found that the enormous volumes of available data are biased toward a small number of globally distributed and carefully chosen varieties, while local breeds (or populations) are frequently overlooked despite their demonstrated resistance to harsh environmental circumstances. Furthermore, the bulk of this research has mostly focused on bacteria, whereas other microbial components such as protists, fungi, and viruses have received far less attention. The majority of these data were gathered utilizing traditional metabarcoding techniques that taxonomically identify the gut microbiota by analyzing small portions of their genome (less than 1000 base pairs). However, to extend the coverage of microbial genomes for a more precise and thorough characterization of microbial communities, a variety of increasingly practical and economical shotgun techniques are currently available.
1. Introduction
The massive decrease in sequencing costs associated with the generalization of high-throughput or next-generation sequencing (NGS) techniques has enabled unprecedented advances in microbiome studies spanning throughout the life sciences fields [1]. The strong bond between public health and the economy has been propelling the interest in microbiome research, which is deemed to hold a huge applicative potential under the One Health strategy [2] and other similar initiatives. Most of such studies addressing health and animal production have been mostly focused on gut microbiota, which is justified by the crucial role of these microorganisms in nutrition, fitness, and performance traits [3,4,5]. It is generally expected that advancing knowledge of the ruminant microbiome [6] bears a huge potential in terms of boosting animal production and health while lessening environmental pollution [7,8]. This promise seems of utmost importance when considering forecasts predicting an almost two-fold increase in the current production and consumption of meat in 30 years from now, with changes in dietary habits in developing countries—on top of human population growth—which will boost the demand for dairy products [9].
Some terms will be used extensively in this review and, for the benefit of the readers, their definition is provided as follows. The community of microorganisms inhabiting a given environment is referred to as a microbiota, while the term microbiome is used to indicate microbiota’s collective genomes [10]. On the other hand, the concept of metagenomics, first defined as “the direct genetic analysis of genomes contained in an environmental sample” [11], has been elaborated further as the “study of the structure and function of entire nucleotide sequences isolated and analyzed from all the organisms (typically microbes) in a bulk sample” [12].
However, the fast-increasing body of research produced in the wake of the above-mentioned compelling socioeconomic reasons has yet to cover much ground. Among the main shortfalls of this kind of study is the almost exclusive focus on cosmopolitan and highly selected breeds, which lacks representativeness both in terms of diversity and functionality, as the most promising knowledge may come from locally adapted native breeds. The role of microorganisms in the resilience and performance of livestock species is of paramount interest for their potential commercial value, especially in a time of rampant global change.
Another critical factor is the uneven attention being paid to bacteria, which include taxa that cause significant economic losses in addition to being a serious hazard to public health, e.g., [13]. On the other hand, the other microorganisms, such as fungi, archaea, protozoa [14], and viruses, have received far less attention. Yet another weakness is that the classical metabarcoding approach is still largely used, as opposed to increasingly feasible and affordable shotgun approaches that are now available (although only for a low number of samples) for a more precise and extensive characterization of microbial communities.
The goal of this review, which was prompted by the steadily increasing number of articles devoted to the livestock gut microbiome, is to assess the primary body of research in this subject, provide an overview of the state-of-the-art regarding sequencing approaches and knowledge produced, and then offer suggestions for future studies.
2. Next-Generation Sequencing Techniques
2.1. Amplicon Metabarcoding
This technique, known as the large-scale taxonomic identification of biological samples through the analysis of short DNA fragments of one or more genes (known as DNA barcodes), has benefited significantly from the development of high-throughput sequencing, which made it possible to process complex environmental samples [https://metazoogene.org/metabarcoding (accessed on 10 September 2022)].
According to the taxonomic group being targeted, different barcodes are preferred. For example, the V2-V3 and V3-V4 16S rRNA hypervariable regions have being traditionally used for bacteria [15,16], the V4 and V9 18S rRNA hypervariable regions for protists [17], and the internal transcribed spacer (ITS) rRNA regions (ITS1 and ITS2) for fungi [18,19]. This PCR-based method relies on a dual indexing mechanism to simultaneously process huge numbers of individual libraries (i.e., samples) covering many taxa at a low cost.
The operational taxonomic units (OTUs) characterized by means of these amplicon libraries, however, are an underrepresentation of the true microbial diversity in the community of interest because DNA barcoding has lower sensitivity and limited resolution when compared to metagenomic data (i.e., spanning entire genomes). Instead, PCR and sequencing errors may result in its overrepresentation [20]. Nevertheless, since amplicon metabarcoding has been widely used by the scientific community worldwide for almost 20 years, a large number of homologous sequences are available for free download from GenBank as well as from other widely used public repositories, including Greengenes v13_8 [21], SILVA 138 [22], and RDP18 (Ribosomal Database Project) [23].
However, it is important to note that these databases are not always regularly updated (the most recent updates were made in 2013, 2019, and 2020, respectively), which can be problematic for users. Another reason to rely on this locus over others is the accessibility of user-friendly software such as the Quantitative Insights into Microbial Ecology pipeline (QIIME) [24], which implements 16S-based tools for taxonomic assignment. This even inspired the development of software that predicts functional profiles of bacterial populations based on their 16S sequences, such as Tax4Fun [25]. Overall, this locus has been used in the majority of livestock microbiome investigations carried out until now, necessitating the establishment of recommendations and best practices for the benefit of the animal science community as a whole [26].
2.2. Shotgun Sequencing
This technique entails randomly shearing one or multiple genomes (for instance, in the case of an environmental sample) into small DNA fragments that are then individually sequenced, mapped to reference genomes and then reassembled in the proper order (for a quick explanation, see [27]). Such a technique, which is not based on PCR, has the benefit of avoiding the formation of amplification artifacts and, by being not reliant on taxon-specific primers, may produce more thorough and reliable results in terms of the overall microbial diversity associated with a given sample thanks to its high sensitivity and resolution power.
The provision of knowledge on the biological functions encoded by the genome(s) being sequenced is another significant benefit [28]. However, because of its high sequencing costs, this technique is not yet scalable for large-scale surveys based on high numbers of samples, despite being simple and fast to execute [29]. The difficulties in recreating the microbial composition in the case of complex and large communities and the high computing expenses connected with data storage and processing are additional limitations of this technique [28,29]. The rapidly expanding community of scientists using shotgun sequencing is fortunate to have access to powerful bioinformatics tools that are being made available, with some like BLAST+ [30] allowing the buildup of customized reference databases based on the inclusion of freely available nucleotide and protein sequences from public repositories.
2.3. Metatranscriptomics
Referred to as the study of genes that are transcribed in microbial communities at a given moment and under certain environmental conditions as measured by the abundance of collective RNA transcripts [31], this culture-independent approach has delivered major insights into niche-specific transcript expression patterns and the ecological functions of microbial taxa within their community [32]. Overall, a common drawback of this suite of techniques, especially RNASeq, is the high costs, which are nonetheless expected to drop in the coming years in parallel with an increase in computational power and specific software such as HISAT [33] or ABioTrans [34].
3. The Significance of Microbiome Studies in Livestock Species
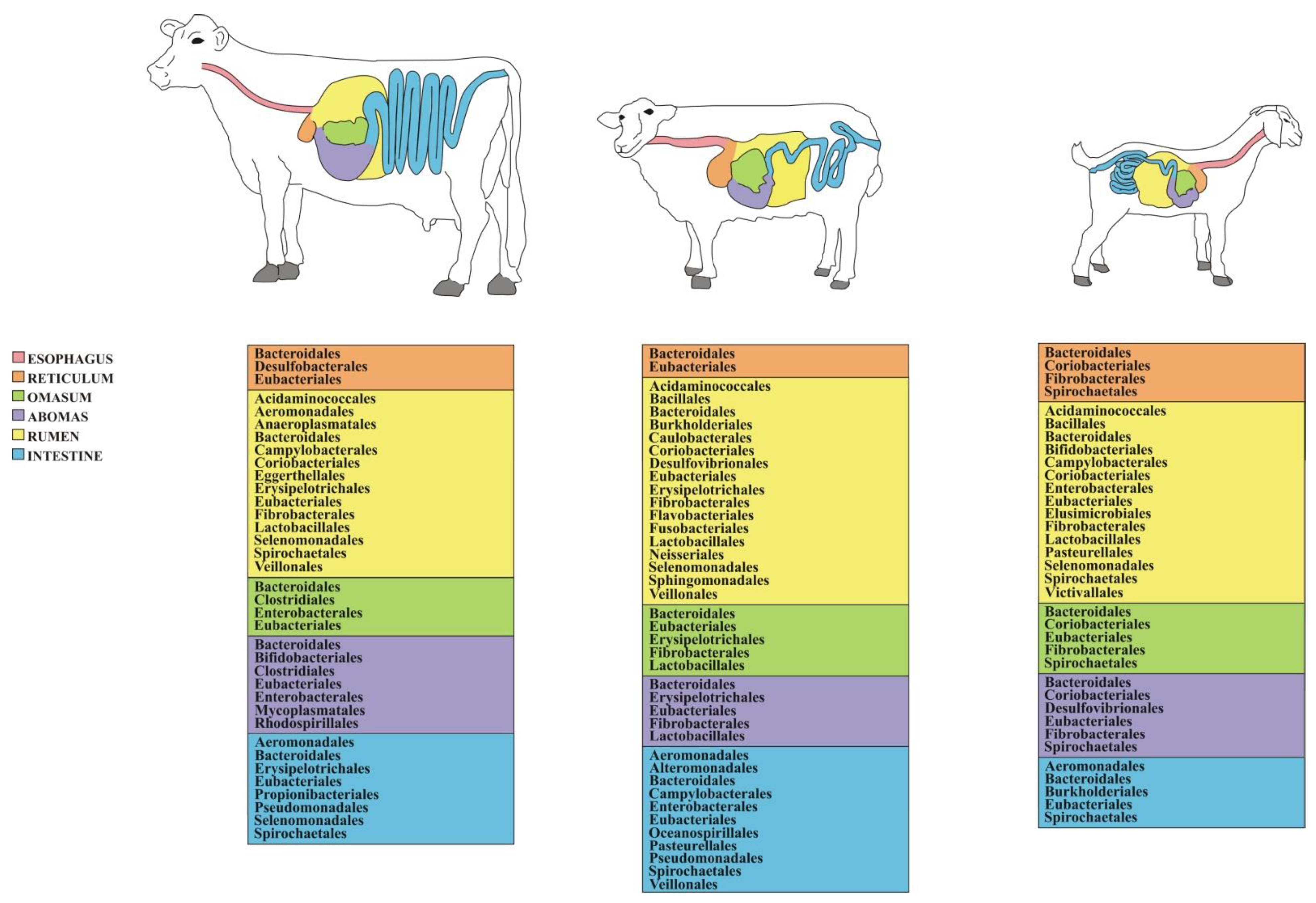
Only five animal species—cows, chickens, goats, pigs, and sheep—produce the great majority of the animal products that humans consume (meat, milk, eggs) [35,36,37]. Each of these species has its own evolutionary history that can be very deep, as is the case of the chicken—a bird—when compared with the other four, which are mammals. Even among the latter, there are notable differences not only in their digestive tracts, but also in terms of physiological aspects such as growth and lifespan as well as reproduction and behavior [38]. There are significant differences between ruminants, which possess a multi-chambered stomach (consisting of reticulum, rumen, omasum and abomasum) used to digest plant materials through fermentation, and monogastric animals, whose stomach is a simple structure made of a single compartment. The advantages of the ruminants regarding their capacity to obtain energy from poor-quality food and the limitations experienced in maintaining a balanced and healthy ruminal flora do not apply to the monogastrics, which are characterized by a faster development and a shorter lifespan.
Indeed, the digestive tract is the most structural factor in animal production as its functionality and health determine most of the individual’s performance [39], and is therefore the region that has been attracting the vast majority of microbiome research, followed by the reproductive tract [40]. Yet, more recently, environmental and public health concerns have also been the focus of a growing number of these studies, namely regarding greenhouse gas emissions [41], the spread of food-borne pathogens [42], and the rise of antibiotic resistance [43].
4. Microbiome Studies in Livestock Species
4.1. Ruminants
4.1.1. Cattle
The majority of gut microbiome studies in cattle have focused on the characterization of microbial communities by 16S rRNA gene amplicon sequencing as a consequence of different animal diet composition [44,45], gastrointestinal tract (GIT) location [46], feed efficiency [47], breed-specificity [48], metabolic disturbs [49], changes over time [50] and individual specificities [51,52], as well as across housing types and farms [53]. Interestingly, special attention has been devoted to identifying individual-based differences irrespective of age, sex, breed, or environment [54], with patterns of similarity and dissimilarities helping to define the core microbiome in the bovine rumen [51] as well as other livestock [55]. At the same time, it was evidenced that differences in taxonomic composition and the underlying community metabolic networks may still result in functional similarity [56], as well as that the metabolic potential of the rumen microbiome may be diet-driven [57]. A large-scale survey of dairy cows indicated that the core rumen microbiome composition underlies not only animal productivity but also the nature of their emissions [58]. It was only recently that shotgun metagenomics opened the door to a thorough exploration of the rumen microbiome composition in cattle, enabling the assembly of entire bacterial genomes (most of which belong to new taxa), and the identification of new enzymes [59]. This approach has also allowed for the elucidation of the interplay between the rumen microbiome along with its metabolome and the host metabolome, shedding new light on the finest mechanisms underlying production performances in dairy cows [60].
4.1.2. Cattle Microbiome Profiling
Compared to other ruminant livestock species, the cattle gut microbiome is probably the one that has been explored more intensively, which provides an exhaustive picture of the bacterial communities inhabiting different GIT locations. The most abundant phyla are represented by Bacteroidetes and Firmicutes, which may account for more than 90% of the entire GIT bacterial community, with Actinobacteria, Proteobacteria, Spirochaetes, and Tenericutes representing other major yet comparatively less abundant taxa [49,53,61,62,63,64,65]. Bacteroidetes and Firmicutes are dominated, respectively, by classes Bacterioidia and Clostridia along with Bacilli. Concerning the major orders (Figure 1), the former class mostly consists of Bacteroidales, while the latter one of Clostridiales [59]. The most abundant families include Bacteroidaceae, Clostridiaceae, Lachnospiraceae, Peptostreptococcaceae, Rikenellaceae, and Ruminococcaceae [53,55], while dominant genera—not only in cattle but in adult ruminants as a whole [55]—are Butyrivibrio, Prevotella and Ruminococcus [51,60,63,64,66]. Genus Clostridium is also abundant in cattle rumen [65] along with Acetitomaculum, Acinetobacter, Mogibacterium, Succiniclasticum, and Treponema [46]. Based on recent studies, genera like Fibrobacter and Ruminococcus are among the core heritable bacteria transferred vertically across generations in the light of their primary role in cellulolysis [58]. A detailed list of the GIT-associated bacterial taxa and the pertinent bibliographic references in cattle is reported in Table S1.
4.1.3. Sheep
The last decade has witnessed a mounting interest in sheep microbiome research. A recent study based on bacterial 16S has confirmed that, similar to what was found in cows, the microbial hosts may be responsible for alterations in terms of feed efficiency [67], while other works have suggested that feeding strategies may promote a more or less diverse microbial community [68,69]. Additionally, compositional changes in the microbiome have been observed along the GIT [70,71] and as an effect of parasite infections [72]. In sheep, however, the compositional changes of the archaeal rather than the eubacterial community play a main role in feed efficiency, with the latter exerting its main influence in terms of the presence/absence pattern of only a few specific taxa [67]. Another recent 16S study compared the microbiome composition in sheep and goats, finding no substantial differences between the two taxa; however, variation did occur depending on age, with older individuals hosting a higher microbial diversity [73], similar to what has been found in Tibetan sheep [71]. Interestingly, differences in gut bacterial compositions have been observed among different Chinese sheep breeds from the Tibetan Plateau [74], contradicting what was found in a similar study on Italian sheep where microbiome differences were mostly due to different husbandry practices [75]. Like in cattle, however, feed efficiency turned out to be related to a higher abundance and diversity of rumen microbiomes [76]. Other studies have been carried out on local breeds of high socioeconomic relevance, often revealing a fairly diverse composition, as in the case of the Chinese Mongolian sheep [77], or, similar to what was found in cattle and goats [78], a marked heterogeneity across different GIT locations as in the case of the Qinghai semi-fine wool sheep [71]. Notably, some recent studies addressing a likely association between host genetics and rumen microbiota in local sheep breeds have unveiled the modulating effect of ovine candidate genes on its composition [79] and the interplay between this and host gene expression in maintaining homeostasis in extreme environments [80]. Nevertheless, all the previous studies are based on 16S metabarcoding, while applications of shotgun metagenomics to characterize the gut microbial composition in sheep are still scant. In this respect, however, it is worth mentioning a study combining the two approaches with metaproteomics to explore the link between microbial communities and biochemical pathways [81].
4.1.4. Sheep Microbiome Profiling
The characterization of the sheep GIT microbiome has revealed its substantial similarity in composition with that of cattle and other ruminants, with Bacteroidetes and Firmicutes making up more than 80 to 90 percent of the gut microbial community [67,69], followed by the phyla Actinobacteria, Proteobacteria, Spirochaetes, and Verrucomicrobia [68,74,82]. Bacterioidia and Clostridia are the dominant classes [75]. Moreover, Bacteroidales and Clostridiales figure among the most abundant orders (Figure 1), while, similar to what is observed in cattle, Eubacteriales and Lactobacillales stand out among Firmicutes. As far as the family-level is concerned, Ruminococcaceae and Lachnospiraceae emerge [74] along with Prevotellaceae, Rikenellaceae, and Succinivibrionaceae [67,76,80]. Concerning the most prevalent genera, Prevotella outstands [80], followed by Acinetobacter [79], Campylobacter [75], Bacteroides, Desulfovibrio, Oscillospira, Ruminococcus, Treponema [77], Fibrobacter, and Succinivibrio [76]. A detailed list of the GIT-associated bacterial taxa and the associated bibliographic references in sheep is provided in Table S1.
4.1.5. Goat
Molecular studies aimed at characterizing gut microbiome composition in this livestock species are still scarce in comparison to sheep, despite the economic relevance of goat meat and dairy products. Interesting exceptions, however, do occur, such as a work exploring the effects of dietary nitrate addition on microbial composition and ruminal fermentation based on a combined metabarcoding approach employing 16S and 18S amplicon libraries to characterize bacteria and protists along with fungi, respectively [83]. Other studies have evidenced the role played by fat acid supplementation [84] and a grain-rich diet [85] in shaping the bacterial and fungal diversity of rumen microbiome based on 16S and ITS metabarcoding, respectively. Interestingly, a recent study based on amplicon libraries of the three loci mentioned before evidenced the role played by specific fungal and bacterial consortia in enabling lignocellulose breakdown by means of the production and interaction of a suite of specific metabolites [86]. Consistently, the 16S-based methanogenic archaea diversity has turned out to be associated with a diet rich in condensed tannin-containing pine bark [87]. Current investigations have evidenced that, in goats as well, the microbial community varies throughout different GIT sectors [88] and tends to increase with age in young individuals [89,90], improving their productive performances [88,91,92]. Concordantly, the inoculation of rumen fluids during early life stages was found to boost the development of the rumen microbiome and even accelerate weaning [93], while the occurrence of apicomplexan parasites in goat kids was found to be associated with a decrease in the abundance of butyrate-producing bacteria, leading to an increase in mucosal inflammation and tissue repair [94]. Contrarily, it was discovered that antibiotic-induced gut microbiota dysbiosis likely worsened disease by encouraging inflammatory immune responses. [95].
Differences in the microbial composition have emerged when comparing adults belonging to different goat breeds [96], even if diet and environment seem to be the more important drivers of microbial diversity than genotype [97]. The occurrence of given bacterial hosts, in turn, was found to be associated with the digestibility of dietary phosphorus [98]. However, the exploration of gut microbiome components other than bacteria is quite limited in goats, with one of the few exceptions being represented by a study employing 16S and 18S amplicon libraries to explore the bacterial and ciliate protozoal diversity, respectively, in relation to the effects of antibacterial peptides on rumen fermentation function [99]. Moreover, the application of shotgun approaches to the characterization of the gut microbiome in goats is still limited to a single recent study [78].
4.1.6. Goat Microbiome Profiling
Compared to other ruminant livestock species, goats are probably those that have so far received less attention concerning gut microbiome studies. Bacteroidetes and Firmicutes are the dominant bacterial phyla (i.e., accounting for more than 80% of the GIT bacterial community), followed by Proteobacteria [89,90,98] and Verrumicrobia [84,88] along with Fibrobacteres, Spirochaetes, and Tenericutes [73,85]. As far as the most abundant orders are concerned, Bacteroidales and Clostridiales—similar to what is observed in cows and sheep—prevail over others (Figure 1) [96]. The dominant families include Prevotellaceae, Veillonellaceae, and, to a lesser extent, Lachnospiraceae, Rikenellaceae, and Ruminococcaceae [84,98]. Among the dominant genera, Prevotella stands out along with Bacteroides, Butyrivibrio, Clostridium, Oscillospira, Ruminococcus, Succiniclasticum, and Succinivibrio [73,84,85,88,90,92,96,100]. A list of the GIT-associated bacterial taxa and the related literature in goat is reported in Table S1.
4.2. Monogastric
4.2.1. Pig
Microbiome research in the pig industry has been propelled by the need to reduce animal stress that may otherwise turn into economic losses for farmers [101]. In this context, weaning is a critical life stage in which the piglet diet undergoes a sharp change. Studies on the swine gut microbiome have largely benefited from the establishment of a reference gene catalogue by means of deep metagenome sequencing of fecal samples [102] and have confirmed that also in this livestock species the interplay between diet and gut physiology across different growth stages is intimately associated with animal health and production performance [103], including fat deposition [104]. Other than varying on the basis of the food provided [105,106,107,108], GIT location [109,110,111], behavior [112], parasite infections [113], breed affiliation, and sex [114], the microbial diversity was found to correlate positively with piglet weight [115] and age [116]. Likewise, studies combining 16S rRNA metabarcoding and shotgun metagenomic sequencing revealed that the composition of the pig gut microbiome varies considerably and predictably across the lifespan [117]. This is particularly evident postweaning [118], when a higher microbial diversity underlies an increase in the genes associated with oxidative stress and heat shock compared to nursing piglets [119]. Interestingly, some studies evidenced that the combination of culturomics and shotgun metagenomics—an approach seldom applied to other livestock species—may deliver a more exhaustive picture of gut [120,121] and antimicrobial resistance [122]. Investigations based on the combination of 16S rRNA metabarcoding and shotgun metagenomics have delivered insights into antimicrobial resistance dynamics in pig farms [108], while 18S rRNA metabarcoding of fecal samples allowed to draw up a detailed list of intestinal protist parasites [123]. The combination of 18S and ITS amplicon libraries has been recently used to characterize the pig gut microbial eukaryote community, finding the association of some of its members with host body weight [124], while that of 16S amplicon data and metagenomics has delivered unprecedented insights into the functional and taxonomic diversity of the pig gut microbiome [123].
4.2.2. Pig Microbiome Profiling
Notwithstanding the pronounced GIT structural differences between ruminants and monogastric animals such as pigs, the gut microbiome of the latter is also dominated by phyla Bacteroidetes and Firmicutes [115], followed by Proteobacteria [103,112], with Bacteroidia and Clostridia being the most abundant classes along with Bacilli [112,124]. Similar to what was observed in other livestock species, the dominant orders are Bacteroidales and Clostridiales (Figure 2), while the most abundant families are Bacteroidaceae, Enterobacteriaceae, Lachnospiraceae, Lactobacillaceae, Prevotellaceae, and Ruminococcaceae [106,108,116]. The genera most commonly found in the GIT of adult pigs are Alloprevotella, Bacteroides, Escherichia, Lactobacillus, and Prevotella [110,120,125,126] along with Clostridium, Desulfovibrio, Enterococcus, Fusobacterium, and Streptococcus [127,128,129]. A list of the GIT-associated bacterial taxa and the pertinent bibliographic references in pig is provided in Table S2.
4.2.3. Chicken
Microbiome research in chicken has made great strides since the advent of NGS techniques, as testified by the studies based on comparative metagenomic pyrosequencing to characterize the cecal microbiome in pathogen-free and infected individuals [130] and to explore the effect of antimicrobials on its communities as well as in relation to the abundance of antimicrobial resistance genes [131]. Nevertheless, most of these investigations are based on 16S rRNA metabarcoding [132], while shotgun metagenomics is just taking its first steps in the poultry sector, with comparative studies applying the two approaches pointing to the much higher resolution power of the latter [133]. Shotgun metagenomics has also recently been employed to assess the role of dietary supplementation in improving the health status—and hence the productive performances—in broiler chickens by fostering the diversity of their cecum microbiome [134], also in the form of in ovo supplementation [92], as well as to characterize new bacterial, archaeal, and bacteriophage taxa of the chicken gut microbiome [135], thus shedding light on their biological function [136]. However, 16S rRNA metabarcoding alone is still widely used to compare the microbiome composition of healthy versus unhealthy individuals as a consequence of viral, e.g., [137], or bacterial [138] infections, of individuals subjected to different dietary treatments [139], as well as across different indigenous breeds [140], GIT locations [141], rearing systems [142] and individual lifetimes [143], with a special focus on improving growth performance by transplanting cecal [144] or fecal [92] material between individuals of different age groups. However, compared to other livestock species, the non-bacterial component of the gut microbiome has been given more attention and most of the studies focus on possible pathogens such as Cryptosporidium [145].
4.2.4. Chicken Microbiome Profiling
Similar to what occurs in the GIT of other livestock species, the most abundant microbial phyla in chicken are Bacteroidetes and Firmicutes [136,146], even though sometimes Proteobacteria are more abundant than the former [135,138,145], while Bacilli, Clostridia, and Gammaproteobacteria are the dominant classes [134]. At the order level, Bacillales, Enterobacteriales, Lactobacillales, and Campylobacterales are the most common groups (Figure 2), while the most prevalent families include Enterobacteriaceae and Lactobacillaceae [139]. As far as the dominant genera are concerned, Alistipes, Bacteroides, Clostridium, Helicobacter, Lactobacillus, and Ruminococcus [133,143,144,146,147,148,149] stand out as well as Flavobacterium [139], Campylobacter, and Veillonella [150]. A detailed list of the GIT-associated bacterial taxa and the related bibliographic references in pig is reported in Table S2.
5. Resistome
The term “resistome” was introduced approximately two decades ago to indicate “the resistance determinants present in the soil” associated with bacterial populations living therein and showing multidrug resistance higher than expected [151]. The expression “bacterial resistome” has since become increasingly popular, while its meaning has evolved into the suite of all antibiotic resistance genes (ARGs) and their precursors in both pathogenic and nonpathogenic bacteria as well as antibiotic producers [152]. With a fast-growing body of research published on this topic, the concept of resistome has further evolved to incorporate different types of resistance and is now a key element in the framework of the One Health approach [153].
The identification of antimicrobial ARGs in bacteria inhabiting livestock GITs is crucial in animal science. An investigation specifically focused on the fecal bacterial resistome used a combination of the two approaches, traditional 16S metabarcoding and shotgun metagenomics, evidencing the strong link between diet and antimicrobial resistance [154]. Moreover, the specificity of the microbial hosts in different GIT locations has emerged in a study on wild and domestic ungulates including cattle and goats [78]. This result serves as a model for future association research by highlighting the significance of local physiological changes along the GIT for various hosts. The advent of innovative nanopore technology, which enables large-scale research to highlight the most abundant resistance genes that may have a significant influence on animal, human, and environmental health, has spurred the rapidly expanding interest in the cow resistome [155]. On the other hand, studies on the bacterial resistome associated with the sheep gut microbiome are still scant when compared to those carried out in cattle or other livestock [156], and no specific investigation has been carried out on goats, but a recent study flagged as many as 30 ARGs in the sheep rumen, most of which related to daptomycin and colistin [157].
As far as non-ruminant livestock species are concerned, the scenario is even more complex. Pigs have received special attention in terms of characterization of the gut bacterial resistome, with recent studies demonstrating differential expression in humans, chickens, and specifically pigs [158]. Yet in chickens, the investigation into the ARGs associated with the gut microbiome has shown that the predominant classes are largely the same as those detected in pigs, including tetracycline, aminoglycoside and macrolide–lincosamide–streptogramin [159]. Of particular interest and utmost topicality is the risk of the potential transmission of ARGs from poultry meat to humans [160].
6. Metagenome and Functional Profile Prediction
Over the last years, a plethora of bioinformatics tools, including PICRUSt (Phylogenetic Investigation of Communities by Reconstruction of Unobserved States [161]), PICRUSt2 [162] along with FaproTax [163], the already mentioned Tax4Fun [25] and Tax4fun2 [164], have been made available to the scientific community for the purpose of predicting the functional profiles of the microbiota investigated in different studies. Moreover, Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis is often used in combination with these software to predict their metagenomic contributions. Since the vast majority of microbiome investigations have so far relied on 16S rRNA, the algorithms of this type of program map the copies of this gene that were obtained in a given study to its homologs in the phylogenetically closest taxa with fully sequenced genomes. In other words, this approach allows predicting the functional metagenomic content without sequencing the entire genomes of the taxa which are actually present in the sample analyzed. Noteworthy, these software can work not only with amplicon metabarcoding but also shotgun sequencing data, even though their accuracy largely relies on available reference genomes and, as of now, it is still severely biased toward human datasets [165]. In addition, it is worth mentioning that a recent soil microbiome study comparing amplicon and shotgun sequencing functional profiling suggested that PICRUSt performs better than Tax4Fun to detect omnipresent functions, whereas Tax4Fun predicted greater abundances of functions from more specialized pathways [166]. Since the predictive tool used can lead to making different inferences, the authors suggested to reap the benefits of combining them rather than relying on either one or another [166].
The paucity of available reference genomes has been specifically invoked by some authors as the reason preventing them from performing functional prediction, e.g., [53], but others who nonetheless opted to perform it still detected significant differences in the predicted metagenomic profiles among GIT locations in dairy cows [46], sheep [77], goats [82], and pigs [126], thus pinpointing the most important metabolic pathways across different gastrointestinal microbial ecosystems. Additionally, functional prediction and KEGG analysis have been applied to unveil the differences in terms of metabolic pathways in the rumen of sheep cohorts with different feed efficiency [76] as well as in the same sheep sampled in different periods of the year [80], goats of different age [90], piglets with different body conditions [115], adult pigs of different breed and sex [71], and chickens of different age [167], breeds [140] or with a different health status [137]. It is conceivable that with the fast-growing increase in reference genomes available, metagenome and functional profile prediction tools will become more and more accurate in the next future, and their employment should be envisaged in any gut microbiome study.
7. Gut Microbiome, Health, and Welfare in Livestock
The positive or negative roles played by gut microbial taxa on health, welfare, behavior and performances in livestock exposed to different production conditions have been extensively discussed in several studies, e.g., [168,169,170,171,172,173,174]. It is worth touching on here why the studies addressing this topic are in such high demand nowadays and how crucial they are in animal farming as well as to broader society. In this respect, it is pertinent to mention their contribution to the development of non-antibiotic microbial therapies based on probiotics [175] as well as in pinpointing biomarkers of feed efficiency to deploy strategies that can notably improve livestock production performances [67,68,76,84,85,98,126,134,148,176] and growth [69,93,115]. Additionally, gut microbial profiling is paramount to monitor livestock health status and set up treatments to boost it, e.g., [113,125,139,144,165,177] as well as to prevent the establishment or aggravation of pathologic states [64,137,138,150] and evidence peculiar adaptations of local breeds to harsh environments [80].
8. Conclusions
The major advances in high-throughput sequencing technology have opened a new era in the study of the livestock gut microbiome, the composition and function of which are tightly associated with animal health and productive performance. The information produced has a profound social and economic impact. Previous attempts to take stock of available gut microbiome studies in livestock were mostly focused on cattle and chickens, or on the microbial groups rather than their hosts. In cattle, microbiome composition has been widely investigated in terms of feeding-related changes and their impacts on production strategies or environmental issues associated with ruminal methane emissions. We have expanded this review to other livestock animals, trying to make the point about what has been mostly performed so far and what is still lacking. A first consideration deals with the subjects of the microbiome studies carried out so far, in which priority was given to some species (such as cattle and pigs) rather than others (such as goats). Additionally, there is a clear bias in terms of the breeds investigated: expectedly, most studies are focused on a few cosmopolitan and highly selected breeds, while local breeds from rural areas are largely neglected, even though livestock research is a fundamental component to boost development strategies and the socioeconomic level of associated human communities. Characterizing the microbiome composition and its interaction with the host in non-intensive husbandry systems might, for instance, provide useful information on how to optimize livestock productivity through nutrient supplementation. Furthermore, it should be noted that a considerable number of microbiome studies, also on local breeds, have been carried out in China or Europe, while much less attention has been devoted to Africa and the tropical and subtropical regions as a whole.
Overall, the body of literature examined in this review allows us to conclude that livestock microbiome composition is affected by age, food, sex, and taxonomy, even if core bacteria occur across the GITs of different species. Admittedly, however, knowledge of other microbial groups is scant. As far as the methodological approach is concerned, shotgun metagenomics is still insipid when compared to amplicon metabarcoding sequencing, even though the few comparative studies employing both approaches on the same datasets evidenced the tremendously higher detection power of the former one, which is much more efficient in identifying underrepresented taxa whose detectability may be biased by the failure of universal primers to hybridize all templates as well as by its reliance on the number of hypervariable regions targeted, e.g., [133,178]. Conventional wisdom suggests that comparisons between studies based on either amplicon metabarcoding or shotgun sequencing on different datasets and with different experimental settings should not be made, but in general it can be stated that the latter is more reliable when estimating the absolute abundance of different microbial taxa. In addition, it is worth mentioning that in an increasing number of studies the two approaches are combined to first obtain a general picture of microbial diversity in the entire sample via amplicon metabarcoding and then, on this basis, select samples for shotgun sequencing to carry out functional analysis with higher accuracy, e.g., [108,127]. It is conceivable that with the fast-decreasing sequencing cost and the increasing suite of powerful bioinformatic tools available, the much more insightful shotgun sequencing will replace amplicon metabarcoding in most gut microbiome studies. This will presumably translate into expanding not only the taxonomic breadth and resolution but also the focus of the research. Indeed, bacteria is the most-studied microbial group compared to the others, which are most often given some attention only when represented by parasites of commercial relevance, but having a large amount of information encompassing other microbial groups as well may trigger interest in promoting research on them.
Indeed, focus on commensal protozoa, which nonetheless may still play a major role in regulating bacterial populations they feed on, is still limited, as is research on the fungal component of the gut microbiome. To achieve a comprehensive knowledge of the function of the microbiome and its underlying dynamics, the characterization of microbial groups other than bacteria is of key importance and should be addressed in future studies. As far as the NGS approach is concerned, it is important to note that the choice is often based on, other than budgetary issues, the availability and accessibility of comparative data as well as on the bioinformatics hurdles associated with the newest and most comprehensive techniques, which may prevent some research groups from applying them due to their limited computational resources and/or expertise. Enhancing the integration of metatranscriptomic studies—which are particularly scant for non-bacterial components—into microbiome research would allow a better understanding of the functional role of different microbial groups in the gastrointestinal tract. Having such valuable tools should not deter researchers from embracing more exhaustive approaches such as those based on shotgun genomics or metatranscriptomics, which, on the one hand, are less affordable and more computationally intensive, yet, on the other hand, may deliver much larger and more accurate amounts of information. These efforts are fully justifiable if we consider that a major application of genomic data in relation to livestock studies is on animal and human health, where epidemiological investigations are fueled by the prospect of threats to human activity and public health with great impact on state wealth. Moreover, the integration of metagenomics and metatranscriptomics with metabolomics and proteomics (multi-omics sequencing) could provide more valuable information about the interaction of the complex “host-microbiota-environment”, which could be useful for deploying future applications and interventions. In this context, there is a pressing need to better our understanding of the reciprocal influence of coexisting humans and livestock on each other’s gut microbiome and resistome.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/ani12233375/s1, Table S1: List of most abundant eubacterial genera found across the GIT of ruminant livestock species; Table S2: List of most abundant eubacterial genera found across the GIT of monogastric livestock species.
Author Contributions
Conceptualization, A.B.-P.; methodology, G.F., L.P.-P.; validation, A.B.-P., J.C.; writing—original draft preparation, G.F.; writing—review and editing, G.F., L.P.-P. and A.B.-P.; visualization, G.F., L.P.-P.; supervision, A.B.-P., L.P.-P.; project administration, L.P.-P., J.C.; funding acquisition, L.P.-P., J.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Portuguese Science and Technology foundation FCT/MCTES, through the grants PTDC/BAA-AGR/28866 and PTDC/BAA-AGR/028575, and from the European Regional Development Fund FEDER (contracts POCI-01-0145-FEDER-028866 and POCI-01-0145-FEDER-028575). LP-P holds a research contract from FCT/MCTES, while G.F. held a research contract through the project PTDC/BAA-AGR/28866 and is currently supported by the Ministry of Universities of the Spanish Government (María Zambrano/Next Generation EU). The article processing charge (APC) exemption was applied thanks to a waiver associated with the special issue ”New Tools for Monitoring Genetic Diversity in Animals”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Amos, G.C.A.; Logan, A.; Anwar, S.; Fritzsche, M.; Mate, R.; Bleazard, T.; Rijpkema, S. Developing standards for the microbiome field. Microbiome 2020, 8, 98. [Google Scholar] [CrossRef] [PubMed]
- European OneHealth/EcoHealth Workshop. Available online: http://www.biodiversity.be/health/58 (accessed on 20 May 2022).
- Park, W. Gut microbiomes and their metabolites shape human and animal health. J. Microbiol. 2018, 56, 151–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, G.O.; Gruninger, R.J.; Badhan, A.; McAllister, T.A. Mining the rumen for fibrolytic feed enzymes. Anim. Front. 2016, 6, 20–26. [Google Scholar] [CrossRef] [Green Version]
- Weimer, P.J. Redundancy, resilience, and host specificity of the ruminal microbiota: Implications for engineering improved ruminal fermentations. Front. Microbiol. 2015, 6, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malmuthuge, N.; Guan, L.L. Gut microbiome and omics: A new definition to ruminant production and health. Anim. Front. 2016, 6, 8–12. [Google Scholar] [CrossRef]
- Alexander, T.W.; Plaizier, J.C. From the Editors: The importance of microbiota in ruminant production. Anim. Front. 2016, 6, 4–7. [Google Scholar] [CrossRef] [Green Version]
- Cholewinska, P.; Czyz, K.; Nowakowski, P.; Wyrostek, A. The microbiome of the digestive system of ruminants—A review. Anim. Health Res. Rev. 2020, 21, 3–14. [Google Scholar] [CrossRef]
- Alexandratos, N.; Bruinsma, J. World Agriculture towards 2030/2050: The 2012 Revision; ESA Working Paper 12-03; FAO: Rome, Italy, 2012. [Google Scholar]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Verges, M.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome definition re-visited: Old concepts and new challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef]
- Thomas, T.; Gilbert, J.; Meyer, F. Metagenomics—A guide from sampling to data analysis. Microb. Inform. Exp. 2012, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- National Human Genome Research Institute. Metagenomics. Available online: https://www.genome.gov/genetics-glossary/Metagenomics (accessed on 12 June 2022).
- Wareth, G.; El-Diasty, M.; Abdel-Hamid, N.H.; Holzer, K.; Hamdy, M.E.R.; Moustafa, S.; Shahein, M.A.; Melzer, F.; Beyer, W.; Pletz, M.W.; et al. Molecular characterization and antimicrobial susceptibility testing of clinical and non-clinical Brucella melitensis and Brucella abortus isolates from Egypt. One Health 2021, 13, 100255. [Google Scholar] [CrossRef]
- Hristov, A.N.; Ivan, M.; Rode, L.M.; McAllister, T.A. Fermentation characteristics and ruminal ciliate protozoal populations in cattle fed medium- or high-concentrate barley-based diets. J. Anim. Sci. 2001, 79, 515–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drancourt, M.; Bollet, C.; Carlioz, A.; Martelin, R.; Gayral, J.-P.; Raoult, R. 16S ribosomal DNA sequence analysis of a large collection of environmental and clinical unidentifiable bacterial isolates. J. Clin. Microbiol. 2000, 38, 3623–3630. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Wang, Y.; Qian, P.Y. Sensitivity and correlation of hypervariable regions in 16S rRNA genes in phylogenetic analysis. BMC Bioinform. 2016, 17, 135. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Park, J.S. Comparative analyses of the V4 and V9 regions of 18S rDNA for the extant eukaryotic community using the Illumina platform. Sci. Rep. 2020, 10, 6519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellemain, E.; Carlsen, T.; Brochmann, C.; Coissac, E.; Taberlet, P.; Kauserud, H. ITS as an environmental DNA barcode for fungi: An in silico approach reveals potential PCR biases. BMC Microbiol. 2010, 10, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [Green Version]
- Poretsky, R.; Rodriguez, R.L.; Luo, C.; Tsementzi, D.; Konstantinidis, K.T. Strengths and limitations of 16S rRNA gene amplicon sequencing in revealing temporal microbial community dynamics. PLoS ONE 2014, 9, e93827. [Google Scholar] [CrossRef] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Maidak, B.L.; Olsen, G.J.; Larsen, N.; Overbeek, R.; McCaughey, M.J.; Woese, C.R. The RDP (Ribosomal Database Project). Nucleic Acids Res. 1997, 25, 109–111. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Asshauer, K.P.; Wemheuer, B.; Daniel, R.; Meinicke, P. Tax4Fun: Predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinroth, M.D.; Belk, A.D.; Dean, C.; Noyes, N.; Dittoe, D.K.; Rothrock, M.J.; Ricke, S.C.; Myer, P.R.; Henniger, M.T.; Ramirez, G.A.; et al. Considerations and best practices in animal science 16S ribosomal RNA gene sequencing microbiome studies. J. Anim. Sci. 2022, 100, skab346. [Google Scholar] [CrossRef] [PubMed]
- National Human Genome Research Institute. Shotgun Sequencing. Available online: https://www.genome.gov/genetics-glossary/Shotgun-Sequencing (accessed on 12 June 2022).
- Sharpton, T.J. An introduction to the analysis of shotgun metagenomic data. Front. Plant. Sci. 2014, 5, 209. [Google Scholar] [CrossRef] [Green Version]
- Peabody, M.A.; Van Rossum, T.; Lo, R.; Brinkman, F.S. Evaluation of shotgun metagenomics sequence classification methods using in silico and in vitro simulated communities. BMC Bioinform. 2015, 16, 363. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- O’Malley, M.A. Metatranscriptomics. In Encyclopedia of Systems Biology; Dubitzky, W., Wolkenhauer, O., Cho, K.H., Yokota, H., Eds.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Waikel, R.L.; Waikel, P.A. Metatranscriptomics. AccessScience. 2011. Available online: https://www.accessscience.com/content/article/aYB110044 (accessed on 27 November 2022).
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Bui, T.T.; Selvarajoo, K. ABioTrans: A Biostatistical Tool for Transcriptomics Analysis. Front. Genet. 2019, 10, 499. [Google Scholar] [CrossRef] [Green Version]
- FAO. How to feed the world in 2050. In Proceedings of the Expert Meeting on How to Feed the World in 2050; Food and Agriculture Organization of the United Nations: Rome, Italy, 2009. [Google Scholar]
- FAO. FAOSTAT. Available online: http://www.fao.org/faostat/en/#home (accessed on 18 March 2022).
- Pulina, G.; Milan, M.J.; Lavin, M.P.; Theodoridis, A.; Morin, E.; Capote, J.; Thomas, D.L.; Francesconi, A.H.D.; Caja, G. Invited review: Current production trends, farm structures, and economics of the dairy sheep and goat sectors. J. Dairy Sci. 2018, 101, 6715–6729. [Google Scholar] [CrossRef] [Green Version]
- Reece, W.O. Functional Anatomy and Physiology of Domestic Animals, 4th ed.; Wiley-Blackwell: Ames, IA, USA, 2009. [Google Scholar]
- Celi, P.; Cowieson, A.J.; Fru-Nji, F.; Steinert, R.E.; Kluenter, A.M.; Verlhac, V. Gastrointestinal functionality in animal nutrition and health: New opportunities for sustainable animal production. Anim. Feed Sci. Technol. 2017, 234, 88–100. [Google Scholar] [CrossRef]
- Heil, B.A.; Paccamonti, D.L.; Sones, J.L. Role for the mammalian female reproductive tract microbiome in pregnancy outcomes. Physiol. Genom. 2019, 51, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Min, B.R.; Solaiman, S.; Waldrip, H.M.; Parker, D.; Todd, R.W.; Brauer, D. Dietary mitigation of enteric methane emissions from ruminants: A review of plant tannin mitigation options. Anim. Nutr. 2020, 6, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Black, Z.; Balta, I.; Black, L.; Naughton, P.J.; Dooley, J.S.G.; Corcionivoschi, N. The Fate of Foodborne Pathogens in Manure Treated Soil. Front. Microbiol. 2021, 12, 781357. [Google Scholar] [CrossRef]
- Zalewska, M.; Blazejewska, A.; Czapko, A.; Popowska, M. Antibiotics and Antibiotic Resistance Genes in Animal Manure—Consequences of Its Application in Agriculture. Front. Microbiol. 2021, 12, 610656. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, J.; Kuehn, L.A.; Bono, J.L.; Berry, E.D.; Kalchayanand, N.; Freetly, H.C.; Benson, A.K.; Wells, J.E. Investigation of bacterial diversity in the feces of cattle fed different diets. J. Anim. Sci. 2014, 92, 683–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palumbo, F.; Squartini, A.; Barcaccia, G.; Macolino, S.; Pornaro, C.; Pindo, M.; Sturaro, E.; Ramanzin, M. A multi-kingdom metabarcoding study on cattle grazing Alpine pastures discloses intra-seasonal shifts in plant selection and faecal microbiota. Sci. Rep. 2021, 11, 889. [Google Scholar] [CrossRef]
- Mao, S.; Zhang, M.; Liu, J.; Zhu, W. Characterising the bacterial microbiota across the gastrointestinal tracts of dairy cattle: Membership and potential function. Sci. Rep. 2015, 5, 16116. [Google Scholar] [CrossRef] [Green Version]
- McGovern, E.; Kenny, D.A.; McCabe, M.S.; Fitzsimons, C.; McGee, M.; Kelly, A.K.; Waters, S.M. 16S rRNA Sequencing Reveals Relationship Between Potent Cellulolytic Genera and Feed Efficiency in the Rumen of Bulls. Front. Microbiol. 2018, 9, 1842. [Google Scholar] [CrossRef]
- Gonzalez-Recio, O.; Zubiria, I.; Garcia-Rodriguez, A.; Hurtado, A.; Atxaerandio, R. Short communication: Signs of host genetic regulation in the microbiome composition in 2 dairy breeds: Holstein and Brown Swiss. J. Dairy Sci. 2018, 101, 2285–2292. [Google Scholar] [CrossRef] [Green Version]
- Plaizier, J.C.; Li, S.; Tun, H.M.; Khafipour, E. Nutritional Models of Experimentally-Induced Subacute Ruminal Acidosis (SARA) Differ in Their Impact on Rumen and Hindgut Bacterial Communities in Dairy Cows. Front. Microbiol. 2016, 7, 2128. [Google Scholar] [CrossRef]
- Rudi, K.; Moen, B.; Sekelja, M.; Frisli, T.; Lee, M.R. An eight-year investigation of bovine livestock fecal microbiota. Vet. Microbiol. 2012, 160, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Jami, E.; Mizrahi, I. Composition and similarity of bovine rumen microbiota across individual animals. PLoS ONE 2012, 7, e33306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.W.; Connor, E.E.; Li, C.; Baldwin Vi, R.L.; Sparks, M.E. Characterization of the rumen microbiota of pre-ruminant calves using metagenomic tools. Environ. Microbiol. 2012, 14, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Hagey, J.V.; Bhatnagar, S.; Heguy, J.M.; Karle, B.M.; Price, P.L.; Meyer, D.; Maga, E.A. Fecal Microbial Communities in a Large Representative Cohort of California Dairy Cows. Front. Microbiol. 2019, 10, 1093. [Google Scholar] [CrossRef] [PubMed]
- Durso, L.M.; Harhay, G.P.; Smith, T.P.; Bono, J.L.; Desantis, T.Z.; Harhay, D.M.; Andersen, G.L.; Keen, J.E.; Laegreid, W.W.; Clawson, M.L. Animal-to-animal variation in fecal microbial diversity among beef cattle. Appl. Environ. Microbiol. 2010, 76, 4858–4862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Global Rumen Census Collaborators; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef] [Green Version]
- Taxis, T.M.; Wolff, S.; Gregg, S.J.; Minton, N.O.; Zhang, C.; Dai, J.; Schnabel, R.D.; Taylor, J.F.; Kerley, M.S.; Pires, J.C.; et al. The players may change but the game remains: Network analyses of ruminal microbiomes suggest taxonomic differences mask functional similarity. Nucleic Acids Res. 2015, 43, 9600–9612. [Google Scholar] [CrossRef] [Green Version]
- Brulc, J.M.; Antonopoulos, D.A.; Miller, M.E.; Wilson, M.K.; Yannarell, A.C.; Dinsdale, E.A.; Edwards, R.E.; Frank, E.D.; Emerson, J.B.; Wacklin, P.; et al. Gene-centric metagenomics of the fiber-adherent bovine rumen microbiome reveals forage specific glycoside hydrolases. Proc. Natl. Acad. Sci. USA 2009, 106, 1948–1953. [Google Scholar] [CrossRef] [Green Version]
- Wallace, R.J.; Sasson, G.; Garnsworthy, P.C.; Tapio, I.; Gregson, E.; Bani, P.; Huhtanen, P.; Bayat, A.R.; Strozzi, F.; Biscarini, F.; et al. A heritable subset of the core rumen microbiome dictates dairy cow productivity and emissions. Sci. Adv. 2019, 5, eaav8391. [Google Scholar] [CrossRef] [Green Version]
- Stewart, R.D.; Auffret, M.D.; Warr, A.; Walker, A.W.; Roehe, R.; Watson, M. Compendium of 4941 rumen metagenome-assembled genomes for rumen microbiome biology and enzyme discovery. Nat. Biotechnol. 2019, 37, 953–961. [Google Scholar] [CrossRef]
- Xue, M.Y.; Sun, H.Z.; Wu, X.H.; Liu, J.X.; Guan, L.L. Multi-omics reveals that the rumen microbiome and its metabolome together with the host metabolome contribute to individualized dairy cow performance. Microbiome 2020, 8, 64. [Google Scholar] [CrossRef]
- Khafipour, E.; Li, S.; Plaizier, J.C.; Krause, D.O. Rumen microbiome composition determined using two nutritional models of subacute ruminal acidosis. Appl. Environ. Microbiol. 2009, 75, 7115–7124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petri, R.M.; Schwaiger, T.; Penner, G.B.; Beauchemin, K.A.; Forster, R.J.; McKinnon, J.J.; McAllister, T.A. Characterization of the Core Rumen Microbiome in Cattle during Transition from Forage to Concentrate as Well as during and after an Acidotic Challenge. PLoS ONE 2013, 8, e83424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jami, E.; Israel, A.; Kotser, A.; Mizrahi, I. Exploring the bovine rumen bacterial community from birth to adulthood. ISME J. 2013, 7, 1069–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, S.; Zhang, R.; Wang, D.; Zhu, W. Impact of subacute ruminal acidosis (SARA) adaptation on rumen microbiota in dairy cattle using pyrosequencing. Anaerobe 2013, 24, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Nagata, R.; Ohkubo, A.; Ohtani, N.; Kushibiki, S.; Ichijo, T.; Sato, S. Changes in ruminal and reticular pH and bacterial communities in Holstein cattle fed a high-grain diet. BMC Vet. Res. 2018, 14, 310. [Google Scholar] [CrossRef]
- Myer, P.; Freetly, H.; Wells, J.; Smith, T.; Kuehn, L. Analysis of the gut bacterial communities in beef cattle and their association with feed intake, growth, and efficiency. J. Anim. Sci. 2017, 95, 3215–3224. [Google Scholar] [CrossRef]
- McLoughlin, S.; Spillane, C.; Claffey, N.; Smith, P.E.; O’Rourke, T.; Diskin, M.G.; Waters, S.M. Rumen Microbiome Composition Is Altered in Sheep Divergent in Feed Efficiency. Front. Microbiol. 2020, 11, 1981. [Google Scholar] [CrossRef]
- Fu, Z.; Xu, X.; Zhang, J.; Zhang, L. Effect of different feeding methods on rumen microbes in growing Chinese Tan sheep. Rev. Bras. Zootec. 2020, 49, e20190258. [Google Scholar] [CrossRef]
- Yu, S.; Zhang, G.; Liu, Z.; Wu, P.; Yu, Z.; Wang, J. Repeated inoculation with fresh rumen fluid before or during weaning modulates the microbiota composition and co-occurrence of the rumen and colon of lambs. BMC Microbiol. 2020, 20, 29. [Google Scholar] [CrossRef]
- Wang, J.; Fan, H.; Han, Y.; Zhao, J.; Zhou, Z. Characterization of the microbial communities along the gastrointestinal tract of sheep by 454 pyrosequencing analysis. Asian-Australas. J. Anim. Sci. 2017, 30, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hu, L.; Liu, H.; Xu, T.; Zhao, N.; Zhang, X.; Geng, Y.; Kang, S.; Xu, S. Characterization of the bacterial microbiota across the different intestinal segments of the Qinghai semi-fine wool sheep on the Qinghai-Tibetan Plateau. Anim. Biosci. 2021, 34, 1921–1929. [Google Scholar] [CrossRef] [PubMed]
- Cortes, A.; Wills, J.; Su, X.; Hewitt, R.E.; Robertson, J.; Scotti, R.; Price, D.R.G.; Bartley, Y.; McNeilly, T.N.; Krause, L.; et al. Infection with the sheep gastrointestinal nematode Teladorsagia circumcincta increases luminal pathobionts. Microbiome 2020, 8, 60. [Google Scholar] [CrossRef] [PubMed]
- Shabana, I.I.; Albakri, N.N.; Bouqellah, N.A. Metagenomic investigation of faecal microbiota in sheep and goats of the same ages. J. Taibah Univ. Sci. 2020, 15, 1–9. [Google Scholar] [CrossRef]
- Chang, J.; Yao, X.; Zuo, C.; Qi, Y.; Chen, D.; Ma, W. The gut bacterial diversity of sheep associated with different breeds in Qinghai province. BMC Vet. Res. 2020, 16, 254. [Google Scholar] [CrossRef]
- Minozzi, G.; Biscarini, F.; Dalla Costa, E.; Chincarini, M.; Ferri, N.; Palestrini, C.; Minero, M.; Mazzola, S.; Piccinini, R.; Vignola, G.; et al. Analysis of Hindgut Microbiome of Sheep and Effect of Different Husbandry Conditions. Animals 2020, 11, 4. [Google Scholar] [CrossRef]
- Zhang, Y.K.; Zhang, X.X.; Li, F.D.; Li, C.; Li, G.Z.; Zhang, D.Y.; Song, Q.Z.; Li, X.L.; Zhao, Y.; Wang, W.M. Characterization of the rumen microbiota and its relationship with residual feed intake in sheep. Animal 2021, 15, 100161. [Google Scholar] [CrossRef]
- Zeng, Y.; Zeng, D.; Ni, X.; Zhu, H.; Jian, P.; Zhou, Y.; Xu, S.; Lin, Y.; Li, Y.; Yin, Z.; et al. Microbial community compositions in the gastrointestinal tract of Chinese Mongolian sheep using Illumina MiSeq sequencing revealed high microbial diversity. AMB Express 2017, 7, 75. [Google Scholar] [CrossRef] [Green Version]
- Xie, F.; Jin, W.; Si, H.; Yuan, Y.; Tao, Y.; Liu, J.; Wang, X.; Yang, C.; Li, Q.; Yan, X.; et al. An integrated gene catalog and over 10,000 metagenome-assembled genomes from the gastrointestinal microbiome of ruminants. Microbiome 2021, 9, 137. [Google Scholar] [CrossRef]
- Mani, S.; Aiyegoro, O.A.; Adeleke, M.A. Association between host genetics of sheep and the rumen microbial composition. Trop. Anim. Health Prod. 2022, 54, 109. [Google Scholar] [CrossRef]
- Lv, W.; Liu, X.; Sha, Y.; Shi, H.; Wei, H.; Luo, Y.; Wang, J.; Li, S.; Hu, J.; Guo, X.; et al. Rumen Fermentation-Microbiota-Host Gene Expression Interactions to Reveal the Adaptability of Tibetan Sheep in Different Periods. Animals 2021, 11, 3529. [Google Scholar] [CrossRef] [PubMed]
- Tanca, A.; Fraumene, C.; Manghina, V.; Palomba, A.; Abbondio, M.; Deligios, M.; Pagnozzi, D.; Addis, M.F.; Uzzau, S. Diversity and functions of the sheep faecal microbiota: A multi-omic characterization. Microb. Biotechnol. 2017, 10, 541–554. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, K.; Zhang, C.; Feng, Y.; Zhang, X.; Wang, X.; Wu, G. Dynamics and stabilization of the rumen microbiome in yearling Tibetan sheep. Sci. Rep. 2019, 9, 19620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asanuma, N.; Yokoyama, S.; Hino, T. Effects of nitrate addition to a diet on fermentation and microbial populations in the rumen of goats, with special reference to Selenomonas ruminantium having the ability to reduce nitrate and nitrite. Anim. Sci. J. 2015, 86, 378–384. [Google Scholar] [CrossRef]
- Cremonesi, P.; Conte, G.; Severgnini, M.; Turri, F.; Monni, A.; Capra, E.; Rapetti, L.; Colombini, S.; Chessa, S.; Battelli, G.; et al. Evaluation of the effects of different diets on microbiome diversity and fatty acid composition of rumen liquor in dairy goat. Animal 2018, 12, 1856–1866. [Google Scholar] [CrossRef] [PubMed]
- Fliegerova, K.O.; Podmirseg, S.M.; Vinzelj, J.; Grilli, D.J.; Kvasnova, S.; Schierova, D.; Sechovcova, H.; Mrazek, J.; Siddi, G.; Arenas, G.N.; et al. The Effect of a High-Grain Diet on the Rumen Microbiome of Goats with a Special Focus on Anaerobic Fungi. Microorganisms 2021, 9, 157. [Google Scholar] [CrossRef]
- Peng, X.; Wilken, S.E.; Lankiewicz, T.S.; Gilmore, S.P.; Brown, J.L.; Henske, J.K.; Swift, C.L.; Salamov, A.; Barry, K.; Grigoriev, I.V.; et al. Genomic and functional analyses of fungal and bacterial consortia that enable lignocellulose breakdown in goat gut microbiomes. Nat. Microbiol. 2021, 6, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Min, B.R.; Solaiman, S.; Shange, R.; Eun, J.S. Gastrointestinal Bacterial and Methanogenic Archaea Diversity Dynamics Associated with Condensed Tannin-Containing Pine Bark Diet in Goats Using 16S rDNA Amplicon Pyrosequencing. Int. J. Microbiol. 2014, 2014, 141909. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Zhang, K.; Li, C.; Wang, X.; Chen, Y.; Yang, Y. Characterization and Comparison of Microbiota in the Gastrointestinal Tracts of the Goat (Capra hircus) During Preweaning Development. Front. Microbiol. 2019, 10, 2125. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Xu, Q.; Kong, F.; Yang, Y.; Wu, D.; Mishra, S.; Li, Y. Exploring the Goat Rumen Microbiome from Seven Days to Two Years. PLoS ONE 2016, 11, e0154354. [Google Scholar] [CrossRef]
- Zou, X.; Liu, G.; Meng, F.; Hong, L.; Li, Y.; Lian, Z.; Yang, Z.; Luo, C.; Liu, D. Exploring the Rumen and Cecum Microbial Community from Fetus to Adulthood in Goat. Animals 2020, 10, 1639. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Yang, Y.; Yan, H.; Wang, X.; Qu, L.; Chen, Y. Rumen bacterial diversity of 80 to 110-day-old goats using 16S rRNA sequencing. PLoS ONE 2015, 10, e0117811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Cai, K.; Mishra, R.; Jha, R. In ovo supplementation of chitooligosaccharide and chlorella polysaccharide affects cecal microbial community, metabolic pathways, and fermentation metabolites in broiler chickens. Poult. Sci. 2020, 99, 4776–4785. [Google Scholar] [CrossRef] [PubMed]
- Palma-Hidalgo, J.M.; Jimenez, E.; Popova, M.; Morgavi, D.P.; Martin-Garcia, A.I.; Yanez-Ruiz, D.R.; Belanche, A. Inoculation with rumen fluid in early life accelerates the rumen microbial development and favours the weaning process in goats. Anim. Microbiome 2021, 3, 11. [Google Scholar] [CrossRef]
- Mammeri, M.; Obregon, D.A.; Chevillot, A.; Polack, B.; Julien, C.; Pollet, T.; Cabezas-Cruz, A.; Adjou, K.T. Cryptosporidium parvum Infection Depletes Butyrate Producer Bacteria in Goat Kid Microbiome. Front. Microbiol. 2020, 11, 548737. [Google Scholar] [CrossRef]
- Tong, J.; Ma, W.; Yang, R.; Wang, T.; Chen, X.; Zhang, X.; Tang, X.; Wen, Y.; Chang, J.; Chen, D. Dysbiosis of the gut microbiota maybe exacerbate orf pathology by promoting inflammatory immune responses. Vet. Microbiol. 2020, 251, 108884. [Google Scholar] [CrossRef]
- Wang, L.; Jin, L.; Xue, B.; Wang, Z.; Peng, Q. Characterizing the bacterial community across the gastrointestinal tract of goats: Composition and potential function. Microbiologyopen 2019, 8, e00820. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Huo, D.; You, Z.; Peng, Q.; Ma, C.; Chang, H.; Lin, X.; Wang, L.; Zhang, J. The distal intestinal microbiome of hybrids of Hainan black goats and Saanen goats. PLoS ONE 2020, 15, e0228496. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Shah, A.M.; Liu, Y.; Jin, L.; Wang, Z.; Xue, B.; Peng, Q. Relationship between true digestibility of dietary phosphorus and gastrointestinal bacteria of goats. PLoS ONE 2020, 15, e0225018. [Google Scholar] [CrossRef]
- Ren, Z.; Yao, R.; Liu, Q.; Deng, Y.; Shen, L.; Deng, H.; Zuo, Z.; Wang, Y.; Deng, J.; Cui, H.; et al. Effects of antibacterial peptides on rumen fermentation function and rumen microorganisms in goats. PLoS ONE 2019, 14, e0221815. [Google Scholar] [CrossRef]
- Zhuang, Y.; Chai, J.; Cui, K.; Bi, Y.; Diao, Q.; Huang, W.; Usdrowski, H.; Zhang, N. Longitudinal Investigation of the Gut Microbiota in Goat Kids from Birth to Postweaning. Microorganisms 2020, 8, 1111. [Google Scholar] [CrossRef] [PubMed]
- Dou, S.; Gadonna-Widehem, P.; Rome, V.; Hamoudi, D.; Rhazi, L.; Lakhal, L.; Larcher, T.; Bahi-Jaber, N.; Pinon-Quintana, A.; Guyonvarch, A.; et al. Characterisation of Early-Life Fecal Microbiota in Susceptible and Healthy Pigs to Post-Weaning Diarrhoea. PLoS ONE 2017, 12, e0169851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Estelle, J.; Kiilerich, P.; Ramayo-Caldas, Y.; Xia, Z.; Feng, Q.; Liang, S.; Pedersen, A.O.; Kjeldsen, N.J.; Liu, C.; et al. A reference gene catalogue of the pig gut microbiome. Nat. Microbiol. 2016, 1, 16161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Tsai, T.; Deng, F.; Wei, X.; Chai, J.; Knapp, J.; Apple, J.; Maxwell, C.V.; Lee, J.A.; Li, Y.; et al. Longitudinal investigation of the swine gut microbiome from birth to market reveals stage and growth performance associated bacteria. Microbiome 2019, 7, 109. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Xiang, Y.; Wang, X.; Dai, B.; Zhang, X.; Ma, L.; Yang, H.; Lyu, W. Exploring the Possible Link between the Gut Microbiome and Fat Deposition in Pigs. Oxid. Med. Cell. Longev. 2022, 2022, 1098892. [Google Scholar] [CrossRef] [PubMed]
- Frese, S.A.; Parker, K.; Calvert, C.C.; Mills, D.A. Diet shapes the gut microbiome of pigs during nursing and weaning. Microbiome 2015, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Klinsoda, J.; Votterl, J.; Koger, S.; Metzler-Zebeli, B.U. Dietary Phytase- and Lactic Acid-Treated Cereals Caused Greater Taxonomic Adaptations than Functional Adaptations in the Cecal Metagenome of Growing Pigs. Appl. Environ. Microbiol. 2020, 87, e02240-20. [Google Scholar] [CrossRef]
- Petry, A.L.; Patience, J.F.; Koester, L.R.; Huntley, N.F.; Bedford, M.R.; Schmitz-Esser, S. Xylanase modulates the microbiota of ileal mucosa and digesta of pigs fed corn-based arabinoxylans likely through both a stimbiotic and prebiotic mechanism. PLoS ONE 2021, 16, e0246144. [Google Scholar] [CrossRef]
- Pollock, J.; Muwonge, A.; Hutchings, M.R.; Mainda, G.; Bronsvoort, B.M.; Gally, D.L.; Corbishley, A. Resistance to change: AMR gene dynamics on a commercial pig farm with high antimicrobial usage. Sci. Rep. 2020, 10, 1708. [Google Scholar] [CrossRef] [Green Version]
- Crespo-Piazuelo, D.; Estelle, J.; Revilla, M.; Criado-Mesas, L.; Ramayo-Caldas, Y.; Ovilo, C.; Fernandez, A.I.; Ballester, M.; Folch, J.M. Characterization of bacterial microbiota compositions along the intestinal tract in pigs and their interactions and functions. Sci. Rep. 2018, 8, 12727. [Google Scholar] [CrossRef]
- De Rodas, B.; Youmans, B.P.; Danzeisen, J.L.; Tran, H.; Johnson, T.J. Microbiome profiling of commercial pigs from farrow to finish. J. Anim. Sci. 2018, 96, 1778–1794. [Google Scholar] [CrossRef]
- Motta, V.; Trevisi, P.; Bertolini, F.; Ribani, A.; Schiavo, G.; Fontanesi, L.; Bosi, P. Exploring gastric bacterial community in young pigs. PLoS ONE 2017, 12, e0173029. [Google Scholar] [CrossRef]
- Verbeek, E.; Keeling, L.; Landberg, R.; Lindberg, J.E.; Dicksved, J. The gut microbiota and microbial metabolites are associated with tail biting in pigs. Sci. Rep. 2021, 11, 20547. [Google Scholar] [CrossRef]
- Borewicz, K.A.; Kim, H.B.; Singer, R.S.; Gebhart, C.J.; Sreevatsan, S.; Johnson, T.; Isaacson, R.E. Changes in the Porcine Intestinal Microbiome in Response to Infection with Salmonella enterica and Lawsonia intracellularis. PLoS ONE 2015, 10, e0139106. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Chen, J.; Gan, M.; Chen, L.; Zhao, Y.; Zhu, Y.; Niu, L.; Zhang, S.; Zhu, L.; Shen, L. Gut Microbiota Composition and Diversity in Different Commercial Swine Breeds in Early and Finishing Growth Stages. Animals 2022, 12, 1607. [Google Scholar] [CrossRef] [PubMed]
- Han, G.G.; Lee, J.Y.; Jin, G.D.; Park, J.; Choi, Y.H.; Chae, B.J.; Kim, E.B.; Choi, Y.J. Evaluating the association between body weight and the intestinal microbiota of weaned piglets via 16S rRNA sequencing. Appl. Microbiol. Biotechnol. 2017, 101, 5903–5911. [Google Scholar] [CrossRef] [PubMed]
- Cremonesi, P.; Biscarini, F.; Castiglioni, B.; Sgoifo, C.A.; Compiani, R.; Moroni, P. Gut microbiome modifications over time when removing in-feed antibiotics from the prophylaxis of post-weaning diarrhea in piglets. PLoS ONE 2022, 17, e0262199. [Google Scholar] [CrossRef] [PubMed]
- Holman, D.B.; Gzyl, K.E.; Mou, K.T.; Allen, H.K. Weaning Age and Its Effect on the Development of the Swine Gut Microbiome and Resistome. Msystems 2021, 6, e0068221. [Google Scholar] [CrossRef]
- Gaio, D.; DeMaere, M.Z.; Anantanawat, K.; Chapman, T.A.; Djordjevic, S.P.; Darling, A.E. Post-weaning shifts in microbiome composition and metabolism revealed by over 25 000 pig gut metagenome-assembled genomes. Microb. Genom. 2021, 7, 000501. [Google Scholar] [CrossRef]
- Guevarra, R.B.; Hong, S.H.; Cho, J.H.; Kim, B.R.; Shin, J.; Lee, J.H.; Kang, B.N.; Kim, Y.H.; Wattanaphansak, S.; Isaacson, R.E.; et al. The dynamics of the piglet gut microbiome during the weaning transition in association with health and nutrition. J Anim. Sci. Biotechnol. 2018, 9, 54. [Google Scholar] [CrossRef] [PubMed]
- Fenske, G.J.; Ghimire, S.; Antony, L.; Christopher-Hennings, J.; Scaria, J. Integration of culture-dependent and independent methods provides a more coherent picture of the pig gut microbiome. FEMS Microbiol. Ecol. 2020, 96, fiaa022. [Google Scholar] [CrossRef]
- Munk, P.; Andersen, V.D.; de Knegt, L.; Jensen, M.S.; Knudsen, B.E.; Lukjancenko, O.; Mordhorst, H.; Clasen, J.; Agerso, Y.; Folkesson, A.; et al. A sampling and metagenomic sequencing-based methodology for monitoring antimicrobial resistance in swine herds. J. Antimicrob. Chemother. 2017, 72, 385–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gweon, H.S.; Shaw, L.P.; Swann, J.; De Maio, N.; AbuOun, M.; Niehus, R.; Hubbard, A.T.M.; Bowes, M.J.; Bailey, M.J.; Peto, T.E.A.; et al. The impact of sequencing depth on the inferred taxonomic composition and AMR gene content of metagenomic samples. Environ. Microbiome 2019, 14, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wylezich, C.; Belka, A.; Hanke, D.; Beer, M.; Blome, S.; Hoper, D. Metagenomics for broad and improved parasite detection: A proof-of-concept study using swine faecal samples. Int. J. Parasitol. 2019, 49, 769–777. [Google Scholar] [CrossRef]
- Ramayo-Caldas, Y.; Prenafeta-Boldu, F.; Zingaretti, L.M.; Gonzalez-Rodriguez, O.; Dalmau, A.; Quintanilla, R.; Ballester, M. Gut eukaryotic communities in pigs: Diversity, composition and host genetics contribution. Anim. Microbiome 2020, 2, 18. [Google Scholar] [CrossRef] [PubMed]
- Mann, E.; Schmitz-Esser, S.; Zebeli, Q.; Wagner, M.; Ritzmann, M.; Metzler-Zebeli, B.U. Mucosa-associated bacterial microbiome of the gastrointestinal tract of weaned pigs and dynamics linked to dietary calcium-phosphorus. PLoS ONE 2014, 9, e86950. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Xin, Y.; Ma, Y.; Xu, X.; Zhao, S.; Cao, J. Screening of Microbes Associated With Swine Growth and Fat Deposition Traits Across the Intestinal Tract. Front. Microbiol. 2020, 11, 586776. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Hu, H.; Zijlstra, R.T.; Zheng, J.; Gänzle, M.G. Metagenomic reconstructions of gut microbial metabolism in weanling pigs. Microbiome 2019, 7, 48. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Wei, S.; Chen, N.; Xiang, Y.; Wang, Y.; Jin, M. Characteristics of gut microbiota in pigs with different breeds, growth periods and genders. Microb. Biotechnol. 2022, 15, 793–804. [Google Scholar] [CrossRef]
- Wylensek, D.; Hitch, T.; Riedel, T.; Afrizal, A.; Kumar, N.; Wortmann, E.; Liu, T.; Devendran, S.; Lesker, T.R.; Hernández, S.B.; et al. A collection of bacterial isolates from the pig intestine reveals functional and taxonomic diversity. Nat. Commun. 2020, 11, 6389. [Google Scholar] [CrossRef]
- Qu, A.; Brulc, J.M.; Wilson, M.K.; Law, B.F.; Theoret, J.R.; Joens, L.A.; Konkel, M.E.; Angly, F.; Dinsdale, E.A.; Edwards, R.A.; et al. Comparative metagenomics reveals host specific metavirulomes and horizontal gene transfer elements in the chicken cecum microbiome. PLoS ONE 2008, 3, e2945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danzeisen, J.L.; Kim, H.B.; Isaacson, R.E.; Tu, Z.J.; Johnson, T.J. Modulations of the chicken cecal microbiome and metagenome in response to anticoccidial and growth promoter treatment. PLoS ONE 2011, 6, e27949. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.Y.; Lee, T.K.; Sul, W.J. Metagenomic Analysis of Chicken Gut Microbiota for Improving Metabolism and Health of Chickens—A Review. Asian-Australas. J. Anim. Sci. 2015, 28, 1217–1225. [Google Scholar] [CrossRef] [Green Version]
- Durazzi, F.; Sala, C.; Castellani, G.; Manfreda, G.; Remondini, D.; De Cesare, A. Comparison between 16S rRNA and shotgun sequencing data for the taxonomic characterization of the gut microbiota. Sci. Rep. 2021, 11, 3030. [Google Scholar] [CrossRef]
- De Cesare, A.; Sirri, F.; Manfreda, G.; Moniaci, P.; Giardini, A.; Zampiga, M.; Meluzzi, A. Effect of dietary supplementation with Lactobacillus acidophilus D2/CSL (CECT 4529) on caecum microbioma and productive performance in broiler chickens. PLoS ONE 2017, 12, e0176309. [Google Scholar] [CrossRef] [Green Version]
- Gilroy, R.; Ravi, A.; Getino, M.; Pursley, I.; Horton, D.L.; Alikhan, N.F.; Baker, D.; Gharbi, K.; Hall, N.; Watson, M.; et al. Extensive microbial diversity within the chicken gut microbiome revealed by metagenomics and culture. PeerJ 2021, 9, e10941. [Google Scholar] [CrossRef] [PubMed]
- Medvecky, M.; Cejkova, D.; Polansky, O.; Karasova, D.; Kubasova, T.; Cizek, A.; Rychlik, I. Whole genome sequencing and function prediction of 133 gut anaerobes isolated from chicken caecum in pure cultures. BMC Genom. 2018, 19, 561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.; Shi, Y.; Liu, P.; Yang, Y.; Zhou, C.; Li, G.; Luo, J.; Zhang, C.; Cao, H.; Hu, G.; et al. 16S rRNA gene sequencing reveals an altered composition of the gut microbiota in chickens infected with a nephropathogenic infectious bronchitis virus. Sci. Rep. 2020, 10, 3556. [Google Scholar] [CrossRef] [Green Version]
- Clavijo, V.; Florez, M.J.V. The gastrointestinal microbiome and its association with the control of pathogens in broiler chicken production: A review. Poult. Sci. 2018, 97, 1006–1021. [Google Scholar] [CrossRef]
- Such, N.; Farkas, V.; Csitari, G.; Pal, L.; Marton, A.; Menyhart, L.; Dublecz, K. Relative Effects of Dietary Administration of a Competitive Exclusion Culture and a Synbiotic Product, Age and Sampling Site on Intestinal Microbiota Maturation in Broiler Chickens. Vet. Sci. 2021, 8, 187. [Google Scholar] [CrossRef]
- Adenaike, A.S.; Akpan, U.; Awopejo, O.O.; Oloye, O.S.; Alli-Balogun, A.O.; Agbaje, M.; Ikeobi, C.O.N. Characterization of the cecal microbiome composition of Nigerian indigenous chickens. Trop. Anim. Health Prod. 2022, 54, 211. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Cho, S.; La, T.M.; Lee, H.J.; Lee, J.B.; Park, S.Y.; Song, C.S.; Choi, I.S.; Lee, S.W. Comparison of microbiota in the cloaca, colon, and magnum of layer chicken. PLoS ONE 2020, 15, e0237108. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xiang, H.; Zhang, H.; Zhu, X.; Wang, D.; Wang, J.; Yin, T.; Liu, L.; Kong, M.; Li, H.; et al. Rearing system causes changes of behavior, microbiome, and gene expression of chickens. Poult. Sci. 2019, 98, 3365–3376. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liao, X.P.; D’Souza, A.W.; Boolchandani, M.; Li, S.H.; Cheng, K.; Luis Martinez, J.; Li, L.; Feng, Y.J.; Fang, L.X.; et al. Environmental remodeling of human gut microbiota and antibiotic resistome in livestock farms. Nat. Commun. 2020, 11, 1427. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, G.A.; Richardson, E.; Clark, J.; Keshri, J.; Drechsler, Y.; Berrang, M.E.; Meinersmann, R.J.; Cox, N.A.; Oakley, B.B. Broiler chickens and early life programming: Microbiome transplant-induced cecal community dynamics and phenotypic effects. PLoS ONE 2020, 15, e0242108. [Google Scholar] [CrossRef]
- Kabir, M.H.B.; Han, Y.; Lee, S.H.; Nugraha, A.B.; Recuenco, F.; Murakoshi, F.; Xuan, X.; Kato, K. Prevalence and molecular characterization of Cryptosporidium species in poultry in Bangladesh. One Health 2020, 9, 100122. [Google Scholar] [CrossRef]
- Wei, S.; Morrison, M.; Yu, Z. Bacterial census of poultry intestinal microbiome. Poult. Sci. 2013, 92, 671–683. [Google Scholar] [CrossRef]
- Saxena, S.; Saxena, V.; Tomar, S.; Sapcota, D.; Gonmei, G. Characterisation of caecum and crop microbiota of Indian indigenous chicken targeting multiple hypervariable regions within 16S rRNA gene. Br. Poult. Sci. 2016, 57, 381–389. [Google Scholar] [CrossRef]
- Biasato, I.; Ferrocino, I.; Grego, E.; Dabbou, S.; Gai, F.; Gasco, L.; Cocolin, L.; Capucchio, M.T.; Schiavone, A. Gut Microbiota and Mucin Composition in Female Broiler Chickens Fed Diets including Yellow Mealworm (Tenebrio molitor, L.). Animals 2019, 9, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Diarra, M.S.; Choi, J.; Rodas-Gonzalez, A.; Lepp, D.; Liu, S.; Lu, P.; Mogire, M.; Gong, J.; Wang, Q.; et al. Effects of encapsulated cinnamaldehyde on growth performance, intestinal digestive and absorptive functions, meat quality and gut microbiota in broiler chickens. Transl. Anim. Sci. 2021, 5, txab099. [Google Scholar] [CrossRef]
- Videnska, P.; Faldynova, M.; Juricova, H.; Babak, V.; Sisak, F.; Havlickova, H.; Rychlik, I. Chicken faecal microbiota and disturbances induced by single or repeated therapy with tetracycline and streptomycin. BMC Vet. Res. 2013, 9, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Costa, V.M.; McGrann, K.M.; Hughes, D.W.; Wright, G.D. Sampling the antibiotic resistome. Science 2006, 311, 374–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skandalis, N.; Maeusli, M.; Papafotis, D.; Miller, S.; Lee, B.; Theologidis, I.; Luna, B. Environmental Spread of Antibiotic Resistance. Antibiotics 2021, 10, 640. [Google Scholar] [CrossRef]
- Kim, D.W.; Cha, C.J. Antibiotic resistome from the One-Health perspective: Understanding and controlling antimicrobial resistance transmission. Exp. Mol. Med. 2021, 53, 301–309. [Google Scholar] [CrossRef]
- Liu, J.; Taft, D.H.; Maldonado-Gomez, M.X.; Johnson, D.; Treiber, M.L.; Lemay, D.G.; DePeters, E.J.; Mills, D.A. The fecal resistome of dairy cattle is associated with diet during nursing. Nat. Commun. 2019, 10, 4406. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Catalina, A.; Atxaerandio, R.; Garcia-Rodriguez, A.; Goiri, I.; Gutierrez-Rivas, M.; Jimenez-Montero, J.A.; Gonzalez-Recio, O. Characterisation of the rumen resistome in Spanish dairy cattle. Anim. Microbiome 2021, 3, 63. [Google Scholar] [CrossRef]
- Ma, T.; McAllister, T.A.; Guan, L.L. A review of the resistome within the digestive tract of livestock. J. Anim. Sci. Biotechnol. 2021, 12, 121. [Google Scholar] [CrossRef]
- Hitch, T.C.A.; Thomas, B.J.; Friedersdorff, J.C.A.; Ougham, H.; Creevey, C.J. Deep sequence analysis reveals the ovine rumen as a reservoir of antibiotic resistance genes. Environ. Pollut. 2018, 235, 571–575. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, Y.; Liu, F.; Cao, J.; Lv, N.; Zhu, B.; Zhang, G.; Gao, G.F. Integrated metagenomic and metatranscriptomic profiling reveals differentially expressed resistomes in human, chicken, and pig gut microbiomes. Environ. Int. 2020, 138, 105649. [Google Scholar] [CrossRef]
- Koorakula, R.; Schiavinato, M.; Ghanbari, M.; Wegl, G.; Grabner, N.; Koestelbauer, A.; Klose, V.; Dohm, J.C.; Domig, K.J. Metatranscriptomic Analysis of the Chicken Gut Resistome Response to In-Feed Antibiotics and Natural Feed Additives. Front. Microbiol. 2022, 13, 833790. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.; Salvatierra, G.; Davila-Barclay, A.; Ayzanoa, B.; Castillo-Vilcahuaman, C.; Huang, M.; Pajuelo, M.J.; Lescano, A.G.; Cabrera, L.; Calderon, M.; et al. Market Chickens as a Source of Antibiotic-Resistant Escherichia coli in a Peri-Urban Community in Lima, Peru. Front. Microbiol. 2021, 12, 635871. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotech. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G. PICRUSt2: An improved and extensible approach for metagenome inference. BioRxiv 2019, 672295. [Google Scholar] [CrossRef] [Green Version]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Wemheuer, F.; Taylor, J.A.; Daniel, R.; Johnston, E.; Meinicke, P.; Thomas, T.; Wemheuer, B. Tax4Fun2: A R-based tool for the rapid prediction of habitat-specific functional profiles and functional redundancy based on 16S rRNA gene marker gene sequences. Environ. Microbiome 2020, 15, 11. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Jones, R.B.; Fodor, A.A. Inference-based accuracy of metagenome prediction tools varies across sample types and functional categories. Microbiome 2020, 8, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toole, D.R.; Zhao, J.; Martens-Habbena, W.; Strauss, S.L. Bacterial functional prediction tools detect but underestimate metabolic diversity compared to shotgun metagenomics in southwest Florida soils. Appl. Soil Ecol. 2021, 168, 104129. [Google Scholar] [CrossRef]
- Sun, B.; Hou, L.; Yang, Y. The Development of the Gut Microbiota and Short-Chain Fatty Acids of Layer Chickens in Different Growth Periods. Front. Vet. Sci. 2021, 8, 666535. [Google Scholar] [CrossRef]
- Kogut, M.H.; Arsenault, R.J. Editorial: Gut Health: The New Paradigm in Food Animal Production. Front. Vet. Sci. 2016, 3, 71. [Google Scholar] [CrossRef] [Green Version]
- Kraimi, N.; Dawkins, M.; Gebhardt-Henrich, S.G.; Velge, P.; Rychlik, I.; Volf, J.; Creach, P.; Smith, A.; Colles, F.; Leterrier, C. Influence of the microbiota-gut-brain axis on behavior and welfare in farm animals: A review. Physiol. Behav. 2019, 210, 112658. [Google Scholar] [CrossRef]
- O’Hara, E.; Neves, A.; Song, Y.; Guan, L.L. The Role of the Gut Microbiome in Cattle Production and Health: Driver or Passenger? Annu. Rev. Anim. Biosci. 2020, 8, 199–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, J.; Reese, A.T. Possibilities and limits for using the gut microbiome to improve captive animal health. Anim. Microbiome 2021, 3, 89. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Luo, S.; Yan, C. Gut Microbiota Implications for Health and Welfare in Farm Animals: A Review. Animals 2022, 12, 93. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Li, D.; Leng, D.; Kui, H.; Bai, X.; Wang, T. Gut microbiota and meat quality. Front. Microbiol. 2022, 13, 951726. [Google Scholar] [CrossRef] [PubMed]
- Lourenco, J.M.; Welch, C.B. Using microbiome information to understand and improve animal performance. Ital. J. Anim. Sci. 2022, 21, 899–913. [Google Scholar] [CrossRef]
- Khalil, A.; Batool, A.; Arif, S. Healthy Cattle Microbiome and Dysbiosis in Diseased Phenotypes. Ruminants 2022, 2, 134–156. [Google Scholar] [CrossRef]
- Clemmons, B.A.; Voy, B.H.; Myer, P.R. Altering the Gut Microbiome of Cattle: Considerations of Host-Microbiome Interactions for Persistent Microbiome Manipulation. Microb. Ecol. 2019, 77, 523–536. [Google Scholar] [CrossRef]
- Malmuthuge, N.; Guan, L.L. Understanding the gut microbiome of dairy calves: Opportunities to improve early-life gut health. J. Dairy Sci. 2017, 100, 5996–6005. [Google Scholar] [CrossRef] [Green Version]
- Campanaro, S.; Treu, L.; Kougias, P.G.; Zhu, X.; Angelidaki, I. Taxonomy of anaerobic digestion microbiome reveals biases associated with the applied high throughput sequencing strategies. Sci. Rep. 2018, 8, 1926. [Google Scholar] [CrossRef]
Figure 1.
List of the most abundant bacterial orders found across the GITs of different ruminant livestock species (see Table S1 for further details). For the sake of clarity, the intestine designation may refer to both small and large intestines.
Figure 1.
List of the most abundant bacterial orders found across the GITs of different ruminant livestock species (see Table S1 for further details). For the sake of clarity, the intestine designation may refer to both small and large intestines.

Figure 2.
List of the most abundant bacterial orders found across the GITs of the two monogastric livestock species presented in this study (see Table S2 for further details).
Figure 2.
List of the most abundant bacterial orders found across the GITs of the two monogastric livestock species presented in this study (see Table S2 for further details).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Forcina, G.; Pérez-Pardal, L.; Carvalheira, J.; Beja-Pereira, A. Gut Microbiome Studies in Livestock: Achievements, Challenges, and Perspectives. Animals 2022, 12, 3375. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12233375
AMA Style
Forcina G, Pérez-Pardal L, Carvalheira J, Beja-Pereira A. Gut Microbiome Studies in Livestock: Achievements, Challenges, and Perspectives. Animals. 2022; 12(23):3375. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12233375
Chicago/Turabian StyleForcina, Giovanni, Lucía Pérez-Pardal, Júlio Carvalheira, and Albano Beja-Pereira. 2022. "Gut Microbiome Studies in Livestock: Achievements, Challenges, and Perspectives" Animals 12, no. 23: 3375. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12233375
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.