
Transcriptome Revealed Exposure to the Environmental Ammonia Induced Oxidative Stress and Inflammatory Injury in Spleen of Fattening Pigs

 , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animal Management and Experimental Design
2.2. Ammonia Exposure
2.3. Histological Observation
2.4. Determination of Antioxidant Indexes by ELISA
2.5. RNA-seq and Interpretation of Results
2.6. Verification of Differentially Expressed Genes
2.7. Statistical Analysis
3. Results
3.1. Splenic Ultrastructural Damage due to Ammonia Exposure
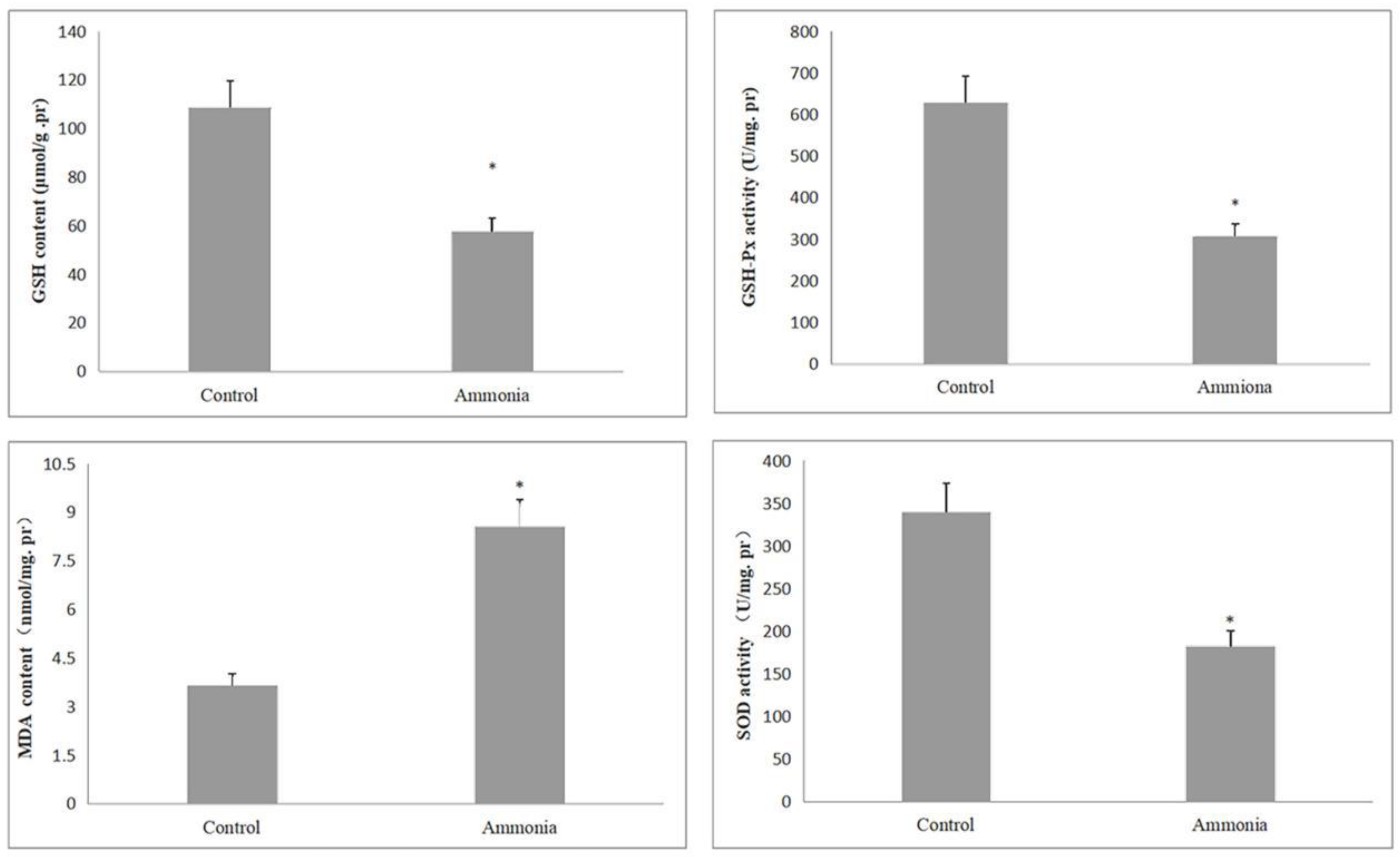
3.2. Effect of Excessive Ammonia on Oxidative Stress Indexes
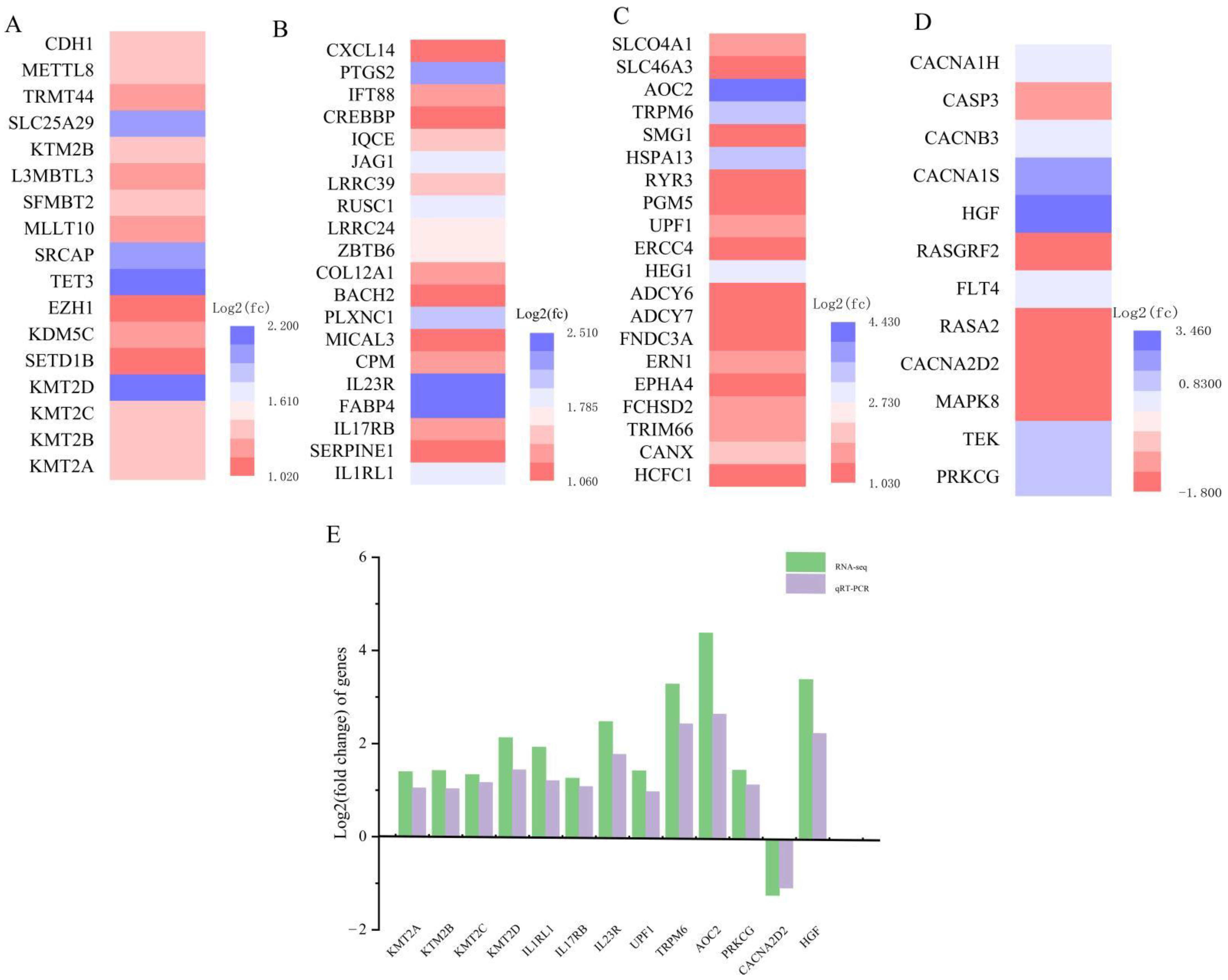
3.3. Differentially Expressed Gene Screening Analysis
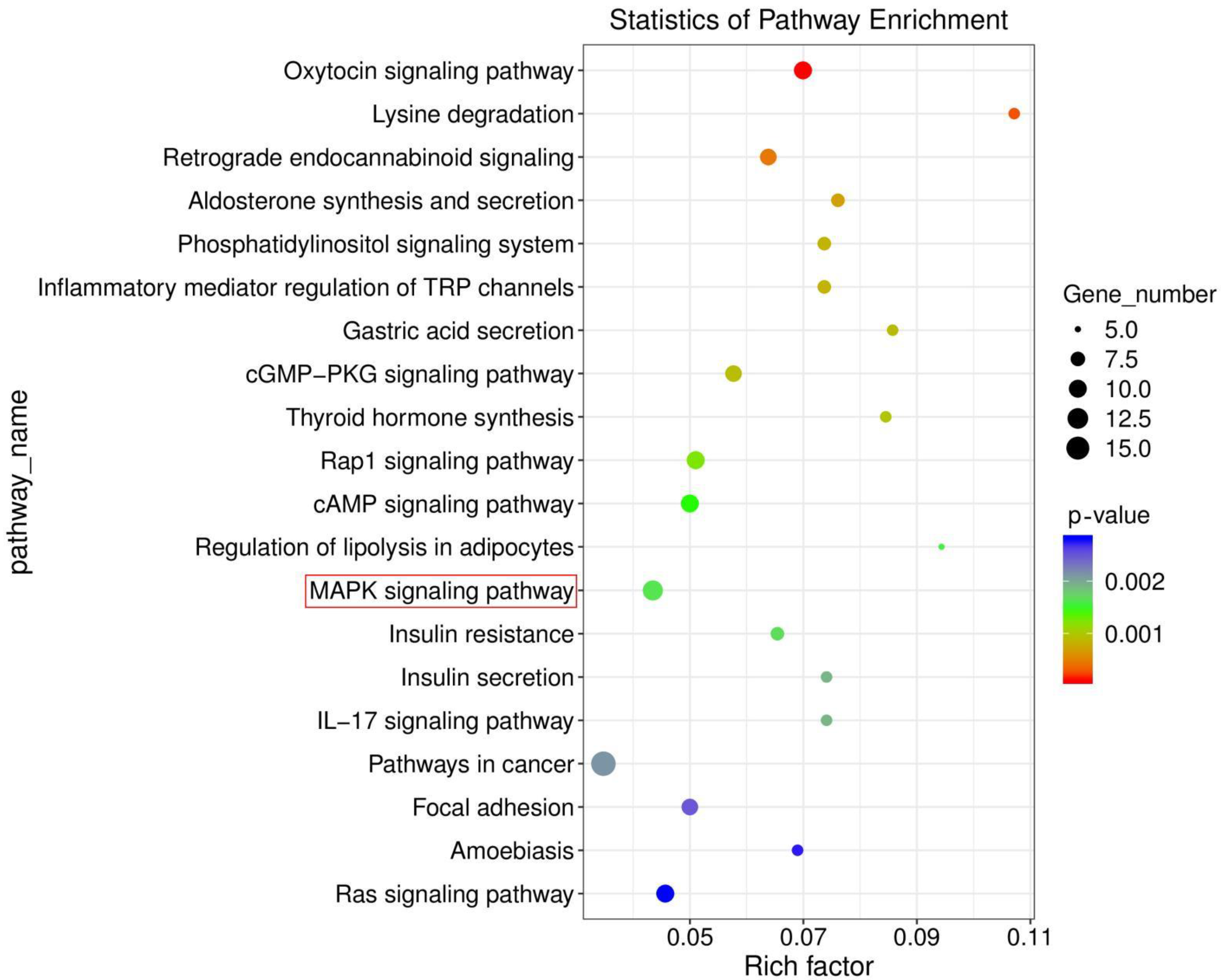
3.4. Ammonia Exposure Activated MAPK Signaling Pathway by KEGG Enrichment Analysis
3.5. GO Enrichment of DEGs
3.6. qRT-PCR Confirmed the Availability of Transcriptome Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shah, S.W.A.; Ishfaq, M.; Nasrullah, M.; Qayum, A.; Akhtar, M.U.; Jo, H.; Hussain, M.; Teng, X. Ammonia inhala-tion-induced inflammation and structural impairment in the bursa of fabricius and thymus of broilers through NF-κB signaling pathway. Environ. Sci. Pollut. Res. 2020, 27, 596–607. [Google Scholar] [CrossRef] [PubMed]
- Behera, S.N.; Sharma, M.; Aneja, V.P.; Balasubramanian, R. Ammonia in the atmosphere: A review on emission sources, atmospheric chemistry and deposition on terrestrial bodies. Environ. Sci. Pollut. Res. 2013, 20, 8092–8131. [Google Scholar] [CrossRef] [PubMed]
- Jdrejek, A. Regional differentiation and possibilities of reducing ammonia emissions from agriculture in poland. Rocz. Nauk. Stowarzyszenia Ekon. Rol. I Agrobiz. 2019, 21, 145–151. [Google Scholar] [CrossRef]
- Saeed, M.; Arain, M.A.; Naveed, M.; Alagawany, M.; Abd El-Hack, M.E.; Bhutto, Z.A.; Bednarczyk, M.; Kakar, M.U.; Abdel-Latif, M.; Chao, S. Yucca schidigera can mitigate ammonia emissions from manure and promote poultry health and production. Environ. Sci. Pollut. Res. 2018, 25, 027–033. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, R.; Li, X.; Li, J.; Ji, W.; Zeng, X.; Bao, J. Exposure to the environmental pollutant ammonia causes changes in gut microbiota and inflammatory markers in fattening pigs. Ecotoxicol. Environ. Saf. 2021, 62, 431–435. [Google Scholar] [CrossRef]
- Mahdinia, M.; Adeli, S.H.; Mohammadbeigi, A.; Heidari, H.; Ghamari, F.; Soltanzadeh, A. Respiratory disorders resulting from exposure to low concentrations of ammonia: A 5-Year Historical Cohort Study. J. Occup. Environ. Med. 2020, 62, 431–435. [Google Scholar] [CrossRef]
- Lewis, S.M.; Williams, A.; Eisenbarth, S.C. Structure and function of the immune system in the spleen. Sci. Immunol. 2019, 4, eaau6085. [Google Scholar] [CrossRef]
- Cheng, C.H.; Guo, Z.X.; Wang, A.L. Growth performance and protective effect of vitamin E on oxidative stress pufferfish (Takifugu obscurus) following by ammonia stress. Fish Physiol. Biochem. 2018, 44, 735–745. [Google Scholar] [CrossRef]
- Bonasio, R.; Tu, S.; Reinberg, D. Molecular signals of epigenetic states. Science 2010, 330, 612–616. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, M.C.; Jivan, A.; Shao, C.; Duan, L.; Goad, D.; Zaganjor, E.; Osborne, J.; McGlynn, K.; Stippec, S.; Earnest, S.; et al. The roles of MAPKs in disease. Cell Res. 2008, 18, 36–42. [Google Scholar] [CrossRef]
- Liu, B.; Wu, S.D.; Shen, L.J.; Zhao, T.X.; Wei, Y.; Tang, X.L.; Long, C.L.; Zhou, Y.; He, D.W.; Lin, T.; et al. Spermatogenesis dysfunction induced by PM2.5 from automobile exhaust via the ROS-mediated MAPK signaling pathway. Ecotoxicol. Environ. Saf. 2019, 167, 161–168. [Google Scholar] [CrossRef]
- Lecoeur, H.; Prina, E.; Rosazza, T.; Kokou, K.; N’Diaye, P.; Aulner, N.; Varet, H.; Bussotti, G.; Xing, Y.; Milon, G.; et al. Targeting macrophage histone H3 modification as a leishmania strategy to dampen the NF-κB/NLRP3-mediated inflammatory response. Cell Rep. 2020, 11, 1870–1882.e4. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, H.A.; Bhattacharyya, D.K.; Kalita, J.K. Differential expression analysis of RNA-seq reads: Overview, Taxonomy, and Tools. IEEE/ACM Trans. Comput. Biol. Bioinform. 2020, 17, 566–586. [Google Scholar] [CrossRef]
- Dhondt, L.; Croubels, S.; De, P.P.; Wallis, S.C.; Pandey, S.; Roberts, J.A.; Lipman, J.; De, C.P.; Devreese, M. Conventional pig as animal model for human renal drug excretion processes: Unravelling the porcine renal function by use of a cocktail of exogenous markers. Front. Pharmacol. 2020, 12, 883. [Google Scholar] [CrossRef]
- Vodicka, P. The miniature pig as an animal model in biomedical research. Ann. N. Y. Acad. Sci. 2010, 1049, 161–171. [Google Scholar] [CrossRef]
- Mebius, R.E.; Kraal, G. Structure and function of the spleen. Nat. Rev. Immunol. 2005, 5, 606–616. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, M.; Liu, C.; Huang, L.; Gao, Y.; Yu, M.; Zhao, S.; Li, X. Ammonia exposure induced cilia dysfunction of nasal mucosa in the piglets. Biomed. Res. Int. 2020, 25, 387. [Google Scholar] [CrossRef]
- Xia, C.; Zhang, X.; Zhang, Y.; Li, J.; Xing, H. Ammonia exposure causes the disruption of the solute carrier family gene network in pigs. Ecotoxicol. Environ. Saf. 2021, 210, 111870. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef]
- Thézénas, M.L.; De, L.B.; Laux-Biehlmann, A.; Bafligil, C.; Elger, B.; Tapmeier, T.; Morten, K.; Rahmioglu, N.; Dakin, S.G.; Charles, P.; et al. Amine oxidase 3 is a novel pro-inflammatory marker of oxidative stress in peritoneal endometriosis lesions. Sci. Rep. 2020, 10, 1495. [Google Scholar] [CrossRef] [Green Version]
- Nazarenus, T.; Cedarberg, R.; Bell, R.; Cheatle, J.; Forch, A.; Haifley, A.; Hou, A.; Wanja, K.B.; Shields, C.; Stoysich, K.; et al. Upf1p, a highly conserved protein required for nonsense-mediated mRNA decay, interacts with the nuclear pore proteins Nup100p and Nup116p. Gene 2005, 345, 199–212. [Google Scholar] [CrossRef] [Green Version]
- Saul, M.J.; Stein, S.; Grez, M.; Jakobsson, P.J.; Steinhilber, D.; Suess, B. UPF1 regulates myeloid cell functions and S100A9 expression by the hnRNP E2/miRNA-328 balance. Sci. Rep. 2016, 6, 31995. [Google Scholar] [CrossRef]
- Cao, G.; Lee, K.P.; van der Wijst, J.; de Graaf, M.; van der Kemp, A.; Bindels, R.J.; Hoenderop, J.G. Methionine sulfoxide reductase B1 (MsrB1) recovers TRPM6 channel activity during oxidative stress. J. Biol. Chem. 2010, 285, 26081–26087. [Google Scholar] [CrossRef] [Green Version]
- Sisinta, T.; Wispriyono, B.; Kusnoputranto, H. Malondialdehyde (mda) as biomarkers of oxidative stress to pm_(2.5) exposure at junior high school students in bandung city, west java, indonesia. Pollut. Res. 2018, 37, 287–294. [Google Scholar]
- Fokam, D.; Hoskin, D. Instrumental role for reactive oxygen species in the inflammatory response. Front. Biosci. 2020, 25, 1110–1119. [Google Scholar] [CrossRef]
- Wang, H.; Zeng, X.; Zhang, X.; Liu, H.; Xing, H. Ammonia exposure induces oxidative stress and inflammation by destroying the microtubule structures and the balance of solute carriers in the trachea of pigs. Ecotoxicol. Environ. Saf. 2021, 212, 111974. [Google Scholar] [CrossRef]
- Wang, K.S.; Li, J.; Wang, Z.; Mi, C.; Ma, J.; Piao, L.X.; Xu, G.H.; Li, X.; Jin, X. Artemisinin inhibits inflammatory response via regulating NF-κB and MAPK signaling pathways. Immunopharmacol. Immunotoxicol. 2017, 39, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Y.; Chi, H.Q.; Zhu, W.; Yang, G.Y.; Song, J.; Mo, L.J.; Zhang, Y.T.; Deng, Y.D.; Xu, F.F.; Yang, J.N.; et al. Cadmium induces renal inflammation by activating the NLRP3 inflammasome through ROS/MAPK/NF-κB pathway in vitro and in vivo. Arch. Toxicol. 2021, 95, 3497–3513. [Google Scholar] [CrossRef] [PubMed]
- Karalis, D.T.; Karalis, T.; Karalis, S.; Kleisiari, A.S. L-Carnitine as a diet supplement in patients with type II diabetes. Cureus 2020, 12, e7982. [Google Scholar] [CrossRef] [PubMed]
- Moosmang, S.; Haider, N.; Klugbauer, N.; Adelsberger, H.; Langwieser, N.; Müller, J.; Stiess, M.; Marais, E.; Schulla, V.; Lacinova, L.; et al. Role of hippocampal Cav1.2 Ca2+ channels in NMDA receptor-independent synaptic plasticity and spatial memory. J. Neurosci. 2005, 25, 9883–9892. [Google Scholar] [CrossRef] [Green Version]
- Piyush, C.; Saryu, M.S.; Sunil, B.G.; Chander, S.R.; Kumar, G.S. Hgf promotes htr-8/svneo cell migration through activation of mapk/pka signaling leading to up-regulation of wnt ligands and integrins that target β-catenin. Mol. Cell Biochem. 2018, 453, 11–32. [Google Scholar]
- Jiayong, Z.; Shengchen, W.; Xiaofang, H.; Gang, S.; Shiwen, X. The antagonistic effect of selenium on lead-induced necroptosis via MAPK/NF-κB pathway and HSPs activation in the chicken spleen. Ecotoxicol. Environ. Saf. 2020, 204, 111049. [Google Scholar] [CrossRef]
- Han, Y.H.; Chen, D.Q.; Jin, M.H.; Jin, Y.H.; Sun, H.N. Anti-inflammatory effect of hispidin on lps induced macrophage inflammation through mapk and jak1/stat3 signaling pathways. Appl. Biol. Chem. 2020, 63, 21. [Google Scholar] [CrossRef]
- Schaunaman, N.; Sanchez, A.; Dimasuay, K.G.; Pavelka, N.; Numata, M.; Alam, R.; Martin, R.J.; Chu, H.W. Interleukin 1 Receptor-Like 1 (IL1RL1) Promotes Airway Bacterial and Viral Infection and Inflammation. Infect. Immun. 2019, 87, e00340-19. [Google Scholar] [CrossRef] [Green Version]
- Iwakura, Y.; Ishigame, H. The IL-23/IL-17 axis in inflammation. J. Clin. Investig. 2006, 116, 1218–1222. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Hedl, M.; Abraham, C. IL23 induces IL23R recycling and amplifies innate receptor-induced signalling and cytokines in human macrophages, and the IBD-protective IL23R R381Q variant modulates these outcomes. Gut 2020, 69, 264–273. [Google Scholar] [CrossRef]
- Greer, E.L.; Shi, Y. Histone methylation: A dynamic mark in health, disease and inheritance. Nat. Rev. Genet. 2012, 13, 343–357. [Google Scholar] [CrossRef] [Green Version]
- Suganuma, T.; Workman, J.L. MAP kinases and histone modification. J. Mol. Cell Biol. 2012, 4, 348–350. [Google Scholar] [CrossRef] [Green Version]
- Whongsiri, P.; Pimratana, C.; Wijitsettakul, U.; Sanpavat, A.; Jindatip, D.; Hoffmann, M.J.; Goering, W.; Schulz, W.A.; Boonla, C. Oxidative stress and LINE-1 reactivation in bladder cancer are epigenetically linked through active chromatin formation. Free. Radic. Biol. Med. 2019, 134, 419–428. [Google Scholar] [CrossRef]
- Husmann, D.; Gozani, O. Histone lysine methyltransferases in biology and disease. Nat. Struct. Mol. Biol. 2019, 26, 880–889. [Google Scholar] [CrossRef]
- Lambhate, S.; tacharjee, D.; Jain, N. APC/C CDH1 ubiquitinates IDH2 contributing to ROS increase in mitosis. Cell. Signal. 2021, 86, 110087. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession No. | Primer Sequence (5′-3′) |
|---|---|---|
| KMT2A | XM_021062911.1 | F: CCTCTCGCTGCTTCACTTCA |
| R: TGAGTTTCGGTCAGAGCCAC | ||
| KMT2B | XM_003127067.5 | F: GAGCAAGATGATGCAGTGCG |
| R: CACGGACTTGTAGTGGCCTT | ||
| KMT2C | XM_021079123.1 | F: TTCGGATATAACTGCCCCGC |
| R: GAGCAGAGAGAGCTGCTGTT | ||
| KMT2D | XM_021091593.1 | F: CGATAGCTCTCCCAGCAAGG |
| R: GTACGGGGCGTGACAGATAG | ||
| IL1RL1 | XM_013995915.2 | F: CAGGGAAGAAGCCACATCGT |
| R: CAAAGCAAGCAGAGCACGTT | ||
| IL17RB | XM_005669645.3 | F: ATCTGTGTGACGGGCAAGAG |
| R: TTCTTTCATGCCTCCGGGTC | ||
| IL23R | NM_001137621.1 | F: GGAAATCATCGGCCTTGCAG |
| R: TTTGTGCTTTGCAATGAGGGA | ||
| UPF1 | XM_021083471.1 | F: GCCAGTTGTTGGCTGAGTTG |
| R: GAGTCGCATGTCAGAGTCAGT | ||
| TRPM6 | XM_021064975.1 | F: CCAGCCACATAGGGCTTTGA |
| R: GGATGACTGACCTCCCCTCT | ||
| AOC2 | XM_003131393.4 | F: AATGTTGGGGGTAGTGCCTG |
| R: CACATCTGGGCGGACTCATT | ||
| PRKCG | XM_021094903.1 | F: CCATTGGATCCCAGCACGAAT |
| R: CTGCAGTTGTCAGCATCAGC | ||
| CACNA2D2 | XM_021068821.1 | F: ATGGACCAACGTGTACGAGG |
| R: AGCAGGAACTCAAAATACTTGACC | ||
| HGF | XM_005667687.3 | F: GCTGCTTCCCCTTCCTCTTT |
| R: GCAAGAATTTGTGCCGGTGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Zhang, R.; Ding, S.; Nian, H.; Zeng, X.; Liu, H.; Xing, H.; Li, J.; Bao, J.; Li, X. Transcriptome Revealed Exposure to the Environmental Ammonia Induced Oxidative Stress and Inflammatory Injury in Spleen of Fattening Pigs. Animals 2022, 12, 1204. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12091204
Chen Y, Zhang R, Ding S, Nian H, Zeng X, Liu H, Xing H, Li J, Bao J, Li X. Transcriptome Revealed Exposure to the Environmental Ammonia Induced Oxidative Stress and Inflammatory Injury in Spleen of Fattening Pigs. Animals. 2022; 12(9):1204. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12091204
Chicago/Turabian StyleChen, Yongjie, Runxiang Zhang, Susu Ding, Haoyang Nian, Xiangyin Zeng, Honggui Liu, Houjuan Xing, Jianhong Li, Jun Bao, and Xiang Li. 2022. "Transcriptome Revealed Exposure to the Environmental Ammonia Induced Oxidative Stress and Inflammatory Injury in Spleen of Fattening Pigs" Animals 12, no. 9: 1204. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12091204