
Phosphoproteomic Analysis of the Jejunum Tissue Response to Colostrum and Milk Feeding in Dairy Calves during the Passive Immunity Period
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sample Collection
2.2. Protein Extraction and Digestion
2.3. Phosphopeptide Enrichment Using Ti4+-IMAC
2.4. Phosphopeptide Identification and Phosphorylation Site Localization Using LC-MS/MS
2.5. Phosphoprotein Identification, Site Localization, Peptides Analysis, and Quantification
2.6. GO and KEGG Pathway Analysis
3. Results
3.1. Phosphoproteome Profiles of Jejunum Tissue Response to Colostrum and Mature Milk Feeding
3.2. Characterization of the Phosphopeptides and Phosphorylation Sites of Phosphoproteins
3.3. Quantitation of Protein Phosphorylation Changes
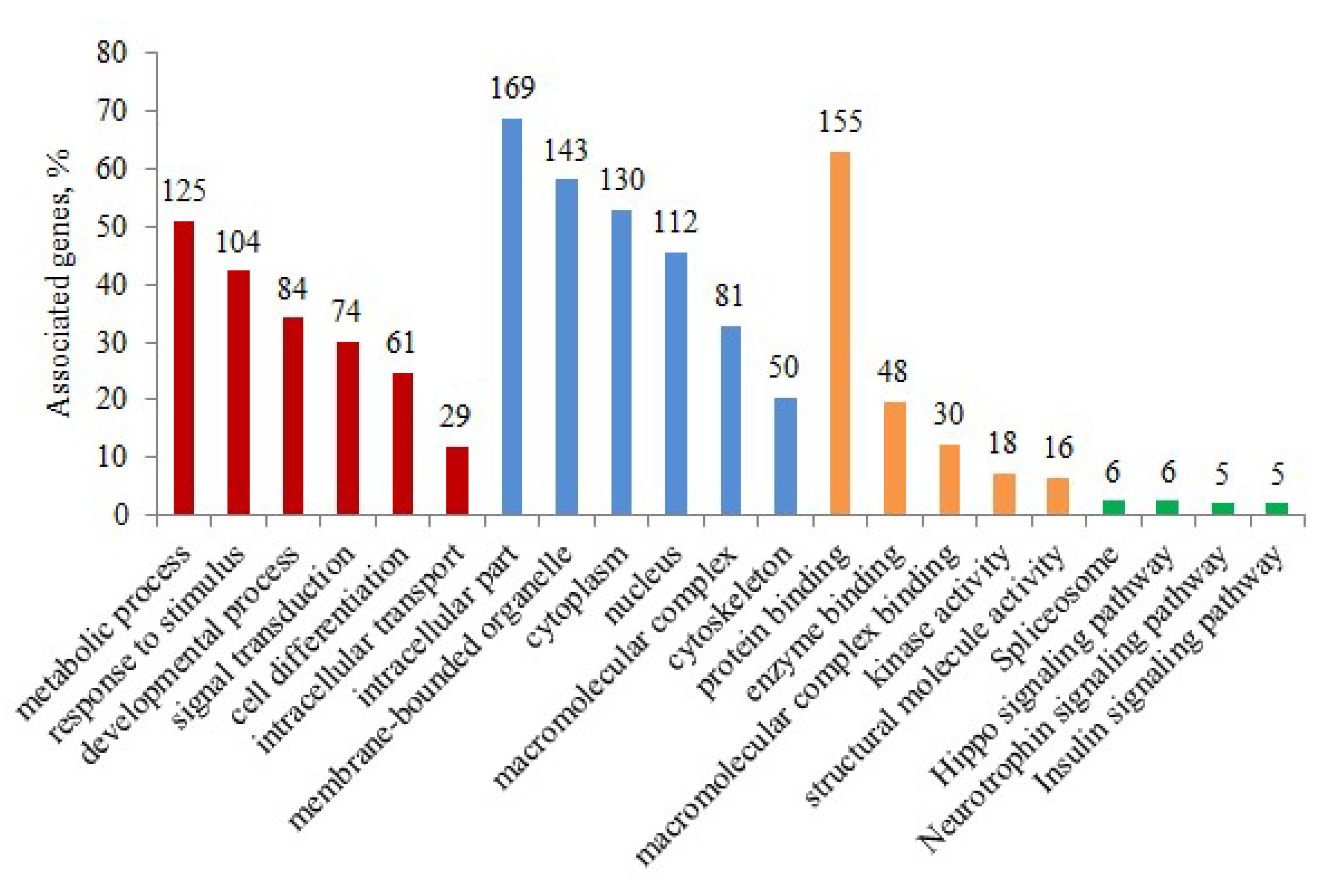
3.4. Functional Categorization of Phosphoproteins from Gene Ontology (GO) Terms
3.5. Phosphoprotein–Phosphoprotein Interaction Networks
4. Discussion
4.1. Phosphoproteins Changes Are Associated with Growth and Development
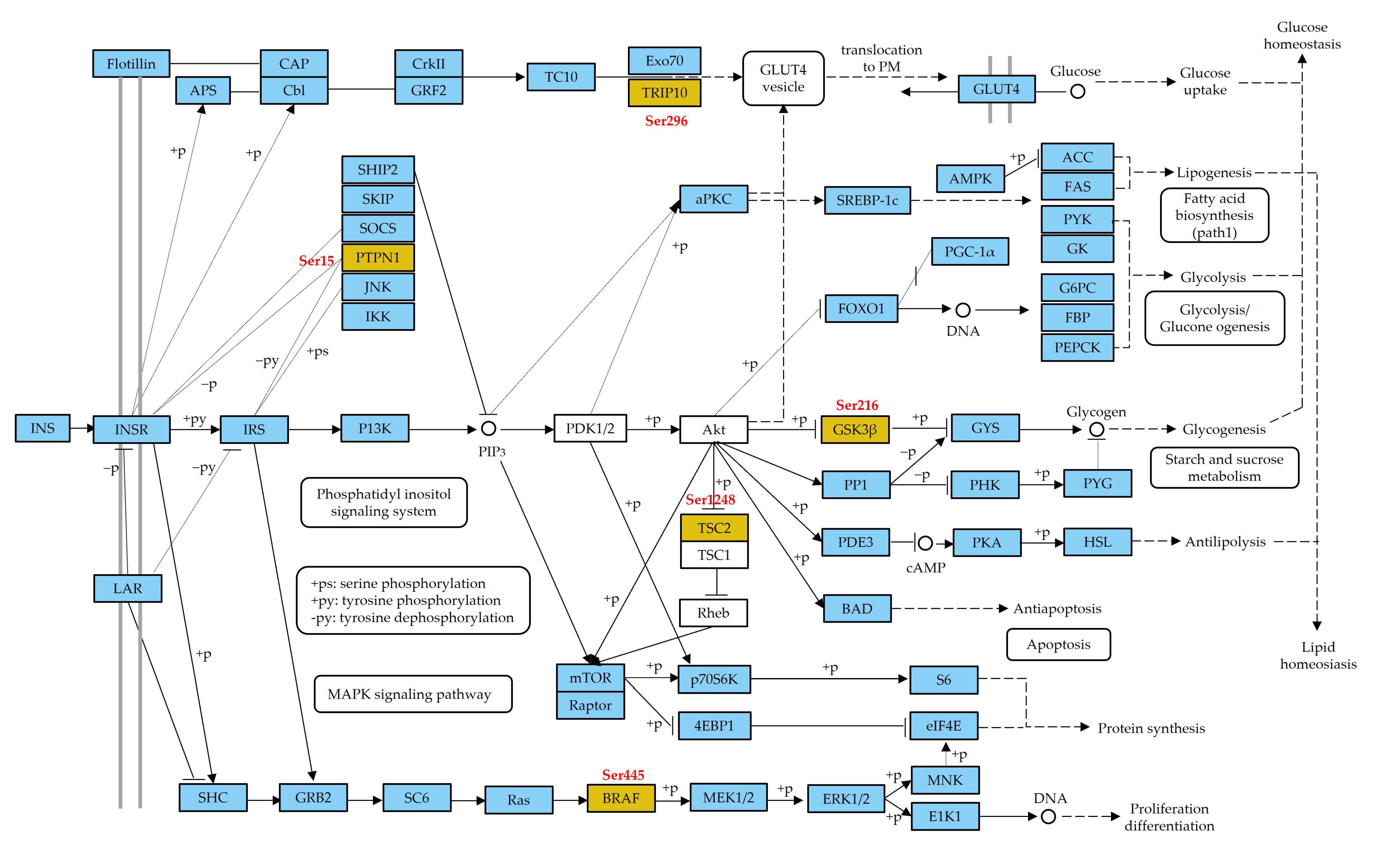
4.2. Phosphoproteins Associated with Insulin Signaling Pathway Play an Important Role in Glycometabolism
4.3. Phosphorylated Proteins Induced by Colostrum Are Involved in the Innate Immunity Response
4.4. GSK3β-TSC2 Interaction Regulates Biological Process through the mTOR Signaling Pathway
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Ti4+-IMAC | Ti4+-immobilized metal ion affinity chromatography |
| ACN | Acetonitrile |
| TFA | Trifluoroacetic acid |
| LC-MS/MS | Liquid chromatography-tandem mass spectrometry |
References
- Roffler, B.; Fäh, A.; Sauter, S.N.; Hammon, H.M.; Gallmann, P.; Brem, G.; Blum, J.W. Intestinal morphology, epithelial cell proliferation, and absorptive capacity in neonatal calves fed milk-born insulin-like growth factor-I or a colostrum extract. J. Dairy Sci. 2003, 86, 1797–1806. [Google Scholar] [CrossRef] [Green Version]
- Fischer–Tlustos, A.J.; Pyo, J.; Song, Y.; Renaud, D.L.; Guan, L.L.; Steele, M.A. Short communication: Effect of delaying the first colostrum feeding on small intestinal histomorphology and serum insulin–like growth factor–1 concentrations in neonatal male Holstein calves. J. Dairy Sci. 2020, 103, 12109–12116. [Google Scholar] [CrossRef] [PubMed]
- Ghaffari, M.H.; Sadri, H.; Steinhoff–Wagner, J.; Hammon, H.M.; Sauerwein, H. Effects of colostrum feeding on the mRNA abundance of genes related to toll–like receptors, key antimicrobial defense molecules, and tight junctions in the small intestine of neonatal dairy calves. J. Dairy Sci. 2021, 104, 10363–10373. [Google Scholar] [CrossRef]
- McGuirk, S.M.; Collins, M. Managing the production, storage, and delivery of colostrum. Vet. Clin. N. Am. Food Anim. Pract. 2004, 20, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Godden, S.M. Colostrum management for dairy calves. Vet. Clin. N. Am. Food Anim. Pract. 2008, 24, 19–39. [Google Scholar] [CrossRef] [PubMed]
- Urie, N.J.; Lombard, J.E.; Shivley, C.B.; Kopral, C.A.; Adams, A.E.; Earleywine, T.J.; Olson, J.D.; Garry, F.B. Preweaned heifer management on US dairy operations: Part V. Factors associated with morbidity and mortality in preweaned dairy heifer calves. J. Dairy Sci. 2018, 101, 9229–9244. [Google Scholar] [CrossRef] [Green Version]
- Godden, S.M.; Lombard, J.E.; Woolums, A.R. Colostrum management for dairy calves. Vet. Clin. N. Am. Food Anim. Pract. 2019, 35, 535–556. [Google Scholar] [CrossRef]
- Uetake, K. Newborn calf welfare: A review focusing on mortality rates. Anim. Sci. J. 2013, 84, 101–105. [Google Scholar] [CrossRef]
- Renaud, D.L.; Duffield, T.F.; LeBlanc, S.J.; Haley, D.B.; Kelton, D.F. Clinical and metabolic indicators associated with early mortality at a milk-fed veal facility: A prospective case-control study. J. Dairy Sci. 2018, 101, 2669–2678. [Google Scholar] [CrossRef] [Green Version]
- Jochims, K.; Kaup, F.J.; Drommer, W.; Pickel, M. An immunoelectron microscopic investigation of colostral IgG absorption across the intestine of newborn calves. Res. Vet. Sci. 1994, 57, 75–80. [Google Scholar] [CrossRef]
- Santaolalla, R.; Abreu, M.T. Innate immunity in the small intestine. Curr. Opin. Gastroenterol. 2012, 28, 124–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martens, E.C.; Neumann, M.; Desai, M.S. Interactions of commensal and pathogenic microorganisms with the intestinal mucosal barrier. Nat. Rev. Microbiol. 2018, 16, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhao, X.; Huang, D.; Wang, J.; Qi, Y.; Jiang, L.; Zhao, H.; Cheng, G. Changes in intestinal proteins induced by colostrum uptake in neonatal calves: Analysis by two–dimensional gel electrophoresis–based proteomics analysis. Anim. Prod. Sci. 2019, 59, 1483–1490. [Google Scholar] [CrossRef] [Green Version]
- Blum, J.W.; Hammon, H.M. Colostrum effects on the gastrointestinal tract, and on nutritional, endocrine and metabolic parameters in neonatal calves. Livest. Prod. Sci. 2000, 66, 151–159. [Google Scholar] [CrossRef]
- Qi, Y.; Zhao, X.; Huang, D.; Pan, X.; Yang, Y.; Zhao, H.; Hu, H.; Cheng, G. Exploration of the relationship between intestinal colostrum or milk, and serum metabolites in neonatal calves by metabolomics analysis. J. Agric. Food Chem. 2018, 66, 7200–7208. [Google Scholar] [CrossRef]
- Zhao, X.W.; Qi, Y.X.; Huang, D.W.; Pan, X.C.; Cheng, G.L.; Zhao, H.L.; Yang, Y.X. Changes in serum metabolites in response to ingested colostrum and milk in neonatal calves, measured by nuclear magnetic resonance-based metabolomics analysis. J. Dairy Sci. 2018, 101, 7168–7181. [Google Scholar] [CrossRef] [Green Version]
- Liang, G.; Malmuthuge, N.; Bao, H.; Stothard, P.; Griebel, P.J. Transcriptome analysis reveals regional and temporal differences in mucosal immune system development in the small intestine of neonatal calves. BMC Genom. 2016, 17, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Liang, G.; Malmuthuge, N.; McFadden, T.B.; Bao, H.; Griebel, P.J.; Stothard, P.; Guan, L.L. Potential regulatory role of microRNAs in the development of bovine gastrointestinal tract during early life. PLoS ONE 2014, 9, e92592. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.W.; Huang, D.W.; Zhu, H.L.; Pan, X.C.; Wang, X.X.; Qi, Y.X.; Cheng, G.L.; Zhao, H.L.; Yang, Y.X. Alterations of the circular RNA profile in the jejunum of neonatal calves in response to colostrum and milk feeding. J. Dairy Sci. 2019, 102, 7038–7048. [Google Scholar] [CrossRef]
- Zhao, X.W.; Zhu, H.L.; Qi, Y.X.; Wu, T.; Huang, D.W.; Ding, H.S.; Chen, S.; Li, M.; Cheng, G.L.; Zhao, H.L.; et al. Quantitative comparative phosphoproteomic analysis of the effects of colostrum and milk feeding on liver tissue of neonatal calves. J. Dairy Sci. 2021, 104, 8265–8275. [Google Scholar] [CrossRef]
- Zhao, X.W.; Zhu, H.L.; Qi, Y.X.; Wu, T.; Huang, D.W.; Cheng, G.L.; Yang, Y.X.; Bu, D.P.; Hu, H.; Meng, L.F. Regulatory role of phosphoproteins in the development of bovine small intestine during early life. J. Dairy Sci. 2022, 105, 9240–9252. [Google Scholar] [CrossRef] [PubMed]
- Villén, J.; Beausoleil, S.A.; Gerber, S.A.; Gygi, S.P. Large-scale phosphorylation analysis of mouse liver. Proc. Natl. Acad. Sci. USA 2007, 104, 1488–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Rauprich, A.B.; Hammon, H.M.; Blum, J.W. Influence of feeding different amounts of first colostrum on metabolic, endocrine, and health status and on growth performance in neonatal calves. J. Anim. Sci. 2000, 78, 896–908. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Zou, Y.; Wu, Z.H.; Li, S.L.; Cao, Z.J. Colostrum quality affects immune system establishment and intestinal development of neonatal calves. J. Dairy Sci. 2015, 98, 7153–7163. [Google Scholar] [CrossRef] [Green Version]
- Woodgett, J.R. Molecular cloning and expression of glycogen synthase kinase-3/factor A. EMBO J. 1990, 9, 2431–2438. [Google Scholar] [CrossRef]
- Aparicio, I.M.; Garcia-Herreros, M.; Fair, T.; Lonergan, P. Identification and regulation of glycogen synthase kinase-3 during bovine embryo development. Reproduction 2010, 140, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Forde, J.E.; Dale, T.C. Glycogen synthase kinase 3, a key regulator of cellular fate. Cell. Mol. Life Sci. 2007, 64, 1930–1944. [Google Scholar] [CrossRef]
- Bali, S.K.; Bryce, D.; Prein, C.; Woodgett, J.R.; Beier, F. Glycogen synthase kinase 3 alpha/beta deletion induces precocious growth plate remodeling in mice. J. Mol. Med. 2021, 99, 831–844. [Google Scholar] [CrossRef]
- Shahbazian, D.; Parsyan, A.; Petroulakis, E.; Topisirovic, I.; Martineau, Y.; Gibbs, B.F. Control of cell survival and proliferation by mammalian eukaryotic initiation factor 4B. Mol. Cell Biol. 2010, 30, 1478–1485. [Google Scholar] [CrossRef]
- Shahbazian, D.; Roux, P.P.; Mieulet, V.; Cohen, M.S.; Raught, B.; Taunton, J.; Hershey, J.W.; Blenis, J.; Pende, M.; Sonenberg, N. The mTOR/PI3K and MAPK pathways converge on eIF4B to control its phosphorylation and activity. EMBO J. 2006, 25, 2781–2791. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, G.; Vazquez-Pianzola, P.; Zurbriggen, A.; Altmann, M.; Sierra, J.M.; Rivera-Pomar, R. Two functionally redundant isoforms of Drosophila melanogaster eukaryotic initiation factor 4B are involved in cap-dependent translation, cell survival and proliferation. Eur. J. Biochem. 2004, 271, 2923–2936. [Google Scholar] [CrossRef]
- Shahbazian, D.; Parsyan, A.; Petroulakis, E.; Hershey, J.W.B.; Sonenberg, N. Eif4b controls survival and proliferation and is regulated by proto-oncogenic signaling pathways. Cell Cycle 2010, 9, 4106–4109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwiatkowski, D.J. Tuberous sclerosis: From tubers to mTOR. Ann. Hum. Genet. 2003, 67, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Inoki, K.; Li, Y.; Xu, T.; Guan, K.L. Rheb GTPase is a direct target of TSC2 GAP activity and regulates mTOR signaling. Genes Dev. 2003, 17, 1829–1834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammon, H.M.; Steinhoff-Wagner, J.; Flor, J.; Schonhusen, U.; Metges, C.C. Lactation Biology Symposium: Role of colostrum and colostrum components on glucose metabolism in neonatal calves. J. Anim. Sci. 2013, 91, 685–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girard, J. Metabolic adaptations to changes of nutrition at birth. Biol. Neonate 1990, 58, 3–15. [Google Scholar] [CrossRef]
- Hsu, C.C.; Leu, Y.W.; Tseng, M.J.; Lee, K.D.; Kuo, T.Y.; Yen, J.Y.; Lai, Y.L.; Hung, Y.C.; Sun, W.S.; Chen, C.M.; et al. Functional characterization of Trip10 in cancer cell growth and survival. J. Biomed. Sci. 2011, 18, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, L.; Adams, R.D.; Saltiel, A.R. The TC10-interacting protein CIP4/2 is required for insulin-stimulated Glut4 translocation in 3T3L1 adipocytes. Proc. Natl. Acad. Sci. USA 2002, 99, 12835–12840. [Google Scholar] [CrossRef] [Green Version]
- Lodhi, I.J.; Chiang, S.H.; Chang, L.; Vollenweider, D.; Watson, R.T.; Inoue, M.; Pessin, J.E.; Saltiel, A.R. Gapex-5, a Rab31 guanine nucleotide exchange factor that regulates Glut4 trafficking in adipocytes. Cell Metab. 2007, 5, 59–72. [Google Scholar] [CrossRef]
- Hartig, S.M.; Ishikura, S.; Hicklen, R.S.; Feng, Y.; Blanchard, E.G.; Voelker, K.A.; Pichot, C.S.; Grange, R.W.; Raphael, R.M.; Klip, A.; et al. The F-BAR protein CIP4 promotes GLUT4 endocytosis through bidirectional interactions with N-WASp and Dynamin-2. J. Cell Sci. 2009, 122, 2283–2291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Hartig, S.M.; Bechill, J.E.; Blanchard, E.G.; Caudell, E.; Corey, S.J. The Cdc42-interacting protein-4 (CIP4) gene knock-out mouse reveals delayed and decreased endocytosis. J. Biol. Chem. 2010, 285, 4348–4354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tegtmeyer, L.C.; Rust, S.; van Scherpenzeel, M. Multiple phenotypes in phosphoglucomutase 1 deficiency. N. Engl. J. Med. 2014, 370, 533–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, W.H.S.; Maclaurin, J.C.; Prineas, J.W. Skeletal muscle glycogenosis: An investigation of two similar cases. J. Neurol. Neurosurg Psychiatry 1963, 26, 60–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelletier, J.; Bellot, G.; Gounon, P.; Lacas-Gervais, S.; Pouysségur, J.; Mazure, N.M. Glycogen synthesis is induced in hypoxia by the hypoxia-inducible factor and promotes cancer cell survival. Front Oncol. 2012, 2, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preisler, N.; Haller, R.G.; Vissing, J. Exercise in muscle glycogen storage diseases. J. Inherit. Metab. Dis. 2015, 38, 551–563. [Google Scholar] [CrossRef]
- Preisler, N.; Laforêt, P.; Echaniz-Laguna, A.; Ørngreen, M.C.; Lonsdorfer-Wolf, E.; Doutreleau, S.; Geny, B.; Stojkovic, T.; Piraud, M.; Petit, F.M.; et al. Fat and carbohydrate metabolism during exercise in phosphoglucomutase type 1 deficiency. J. Clin. Endocrinol. Metab. 2013, 98, 1235–1240. [Google Scholar] [CrossRef] [Green Version]
- Maa, M.C.; Hsieh, C.Y.; Leu, T.H. Overexpression of p97Eps8 leads to cellular transformation. Implication of pleckstrin homology domain in p97Eps8-mediated ERK activation. Oncogene 2001, 20, 106–112. [Google Scholar] [CrossRef] [Green Version]
- Di Fiore, P.P.; Scita, G. Eps8 in the midst of GTPases. Int. J. Biochem. Cell Biol. 2002, 34, 1178–1183. [Google Scholar] [CrossRef]
- Chen, Y.J.; Hsieh, M.Y.; Chang, M.Y.; Chen, H.C.; Leu, T.H. Eps8 facilitates phagocytosis by increasing tlr4-myd88 interaction in lps-stimulated macrophages. J. Biol. Chem. 2012, 26, 18806–18819. [Google Scholar] [CrossRef]
- Kobayashi, K.; Hernandez, L.D.; Galán, J.E.; Janeway, C.A., Jr.; Medzhitov, R.; Flavell, R.A. IRAK-M is a negative regulator of toll-like receptor signaling. Cell 2002, 110, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Balaci, L.; Spada, M.C.; Olla, N.; Sole, G.; Loddo, L.; Anedda, F.; Naitza, S.; Zuncheddu, M.A.; Maschio, A.; Altea, D. IRAK-M is involved in the pathogenesis of early-onset persistent asthma. Am. J. Hum. Genet. 2007, 80, 1103–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saenger, Y.; Magidson, J.; Liaw, B.; de Moll, E.; Harcharik, S.; Fu, Y.; Wassmann, K.; Fisher, D.; Kirkwood, J.; Oh, W.K.; et al. Blood mRNA expression profiling predicts survival in patients treated with tremelimumab. Clin. Cancer Res. 2014, 20, 3310–3318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kesselring, R.; Glaesner, J.; Hiergeist, A.; Naschberger, E.; Neumann, H.; Brunner, S.M.; Wege, A.K.; Seebauer, C.; Köhl, G.; Merkl, S. IRAK-M expression in tumor cells supports colorectal cancer progression through reduction of antimicrobial defense and stabilization of STAT3. Cancer Cell 2016, 29, 684–696. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.M.; Blenis, J. Molecular mechanisms of mTOR-mediated translational control. Nat. Rev. Mol. Cell Biol. 2009, 10, 307–318. [Google Scholar] [CrossRef]
- Inoki, K.; Ouyang, H.; Zhu, T.; Lindvall, C.; Wang, Y.; Zhang, X.; Yang, Q.; Bennett, C.; Harada, Y.; Stankunas, K.; et al. TSC2 integrates Wnt and energy signals via a coordinated phosphorylation by AMPK and GSK3 to regulate cell growth. Cell 2006, 126, 955–968. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Qi, Y.; Wu, T.; Cheng, G. Phosphoproteomic Analysis of the Jejunum Tissue Response to Colostrum and Milk Feeding in Dairy Calves during the Passive Immunity Period. Animals 2023, 13, 145. https://0-doi-org.brum.beds.ac.uk/10.3390/ani13010145
Zhao X, Qi Y, Wu T, Cheng G. Phosphoproteomic Analysis of the Jejunum Tissue Response to Colostrum and Milk Feeding in Dairy Calves during the Passive Immunity Period. Animals. 2023; 13(1):145. https://0-doi-org.brum.beds.ac.uk/10.3390/ani13010145
Chicago/Turabian StyleZhao, Xiaowei, Yunxia Qi, Tao Wu, and Guanglong Cheng. 2023. "Phosphoproteomic Analysis of the Jejunum Tissue Response to Colostrum and Milk Feeding in Dairy Calves during the Passive Immunity Period" Animals 13, no. 1: 145. https://0-doi-org.brum.beds.ac.uk/10.3390/ani13010145