
Fermentation Quality, In Vitro Digestibility, and Aerobic Stability of Ensiling Spent Mushroom Substrate with Microbial Additives
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Silage Production and Treatments
2.2. Chemical Composition Analysis
2.3. Fermentation Quality and Microbiological Analyses
2.4. Aerobic Stability Analysis
2.5. In Vitro Degradability Analysis
2.6. Statistical Analyses
3. Results
3.1. Fermentation Quality of Spent Mushroom Substrate Silage
3.2. Chemical Compositions of Spent Mushroom Substrate Silage
3.3. Energy and In Vitro Digestibility of Spent Mushroom Substrate Silage
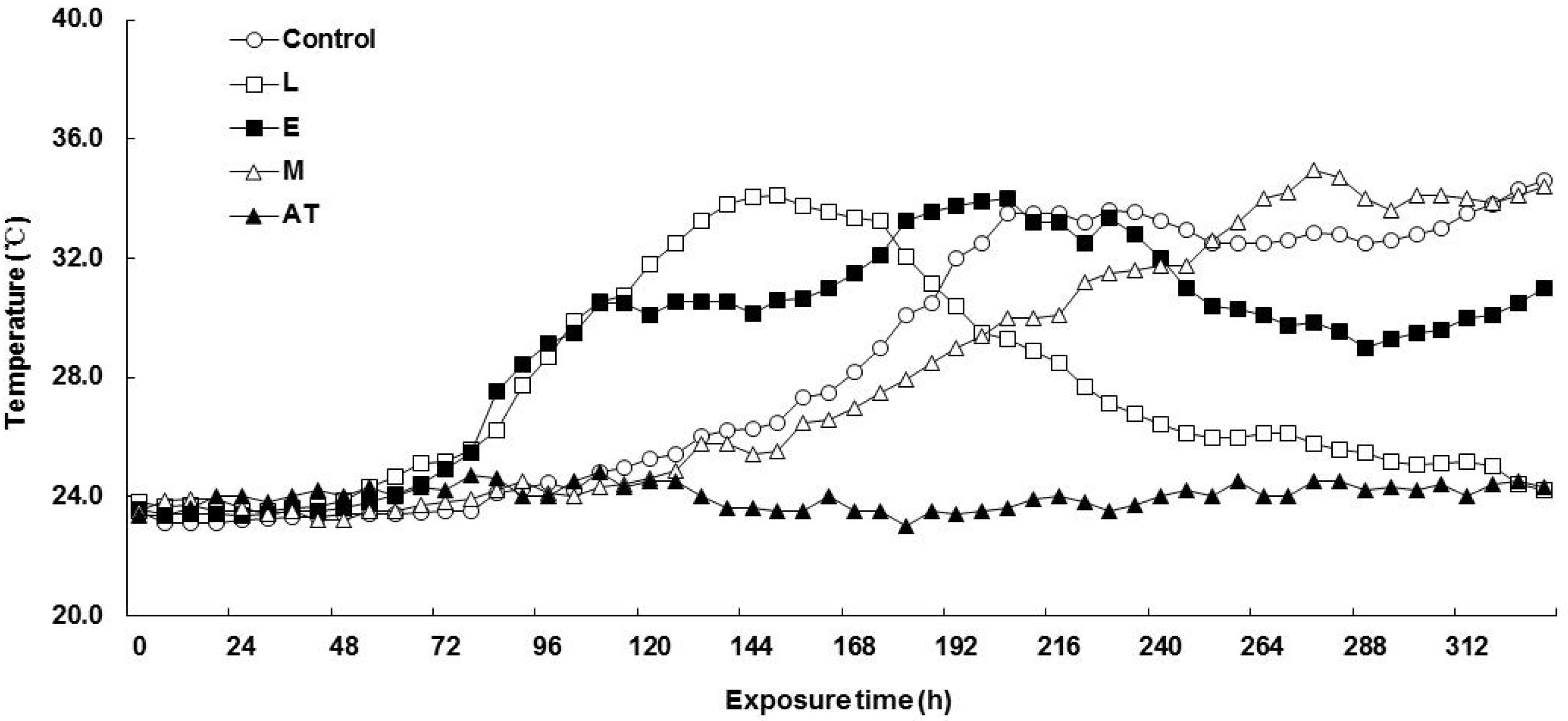
3.4. Aerobic Stability of Spent Mushroom Substrate Silage
4. Discussion
4.1. Effects of Additives on Fermentation Quality
4.2. Effects of Additives on Chemical Composition and In Vitro Digestibility
4.3. Effect of Additives on Aerobic Stability
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rinne, M.; Franco, M.; Jalava, T.; Järvenpää, E.; Kahala, M.; Blasco, L.; Siljander-Rasi, H.; Kuoppala, K. Carrot by-product fermentation quality and aerobic spoilage could be modified with silage additives. Agric. Food Sci. 2019, 28, 59–69. [Google Scholar] [CrossRef]
- Phan, C.-W.; Sabaratnam, V. Potential uses of spent mushroom substrate and its associated lignocellulosic enzymes. Appl. Microbiol. Biotechnol. 2012, 96, 863–873. [Google Scholar] [CrossRef]
- Xu, C.; Cai, Y.; Zhang, J.; Matsuyama, H. Feeding value of total mixed ration silage with spent mushroom substrate. Anim. Sci. J. 2010, 81, 194–198. [Google Scholar] [CrossRef]
- Lau, K.L.; Tsang, Y.Y.; Chiu, S.W. Use of spent mushroom compost to bioremediate PAH-contaminated samples. Chemosphere 2003, 52, 1539–1546. [Google Scholar] [CrossRef]
- Ren, H.; Feng, Y.; Pei, J.; Li, J.; Wang, Z.; Fu, S.; Zheng, Y.; Li, Z.; Peng, Z. Effects of Lactobacillus plantarum additive and temperature on the ensiling quality and microbial community dynamics of cauliflower leaf silages. Bioresour. Technol. 2020, 307, 123238. [Google Scholar] [CrossRef]
- Napasirth, V.; Napasirth, P.; Sulinthone, T.; Phommachanh, K.; Cai, Y. Microbial population, chemical composition and silage fermentation of cassava residues. Anim. Sci. J. 2015, 86, 842–848. [Google Scholar] [CrossRef]
- Kaewpila, C.; Khota, W.; Gunun, P.; Kesorn, P.; Cherdthong, A. Strategic Addition of Different Additives to Improve Silage Fermentation, Aerobic Stability and In Vitro Digestibility of Napier Grasses at Late Maturity Stage. Agriculture 2020, 10, 262. [Google Scholar] [CrossRef]
- Cai, Y.; Du, Z.; Yamasaki, S.; Nguluve, D.; Tinga, B.; Macome, F.; Oya, T. Community of natural lactic acid bacteria and silage fermentation of corn stover and sugarcane tops in Africa. Asian-Australas. J. Anim. Sci. 2020, 33, 1252–1264. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, L.; Ma, G.; Jiang, X.; Yang, J.; Lv, J.; Zhang, Y. Cellulase Interacts with Lactic Acid Bacteria to Affect Fermentation Quality, Microbial Community, and Ruminal Degradability in Mixed Silage of Soybean Residue and Corn Stover. Animals 2021, 11, 334. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 18th ed.; AOAC Int.: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Playne, M.J.; McDonald, P. The buffering constituents of herbage and of silage. J. Sci. Food Agric. 1966, 17, 264–268. [Google Scholar] [CrossRef]
- Deriaz, R.E. Routine analysis of carbohydrates and lignin in herbage. J. Sci. Food Agric. 1961, 12, 152–160. [Google Scholar] [CrossRef]
- Gao, J.L.; Wang, P.; Zhou, C.H.; Li, P.; Tang, H.Y.; Zhang, J.B.; Cai, Y. Chemical composition and in vitro digestibility of corn stover during field exposure and the fermentation characteristics of silage prepared with microbial additives. Asian-Australas. J. Anim. Sci. 2019, 32, 1854–1863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owens, V.N.; Albrecht, K.A.; Muck, R.E. Protein degradation and ensiling characteristics of red clover and alfalfa wilted under varying levels of shade. Can. J. Plant Sci. 1999, 79, 209–222. [Google Scholar] [CrossRef]
- Robinson, D. Compensatory Changes in the Partitioning of Dry Matter in Relation to Nitrogen Uptake and Optimal Variations in Growth. Ann. Bot. 1986, 58, 841–848. [Google Scholar] [CrossRef]
- Yuan, X.; Guo, G.; Wen, A.; Desta, S.T.; Wang, J.; Wang, Y.; Shao, T. The effect of different additives on the fermentation quality, in vitro digestibility and aerobic stability of a total mixed ration silage. Anim. Feed Sci. Technol. 2015, 207, 41–50. [Google Scholar] [CrossRef]
- Kung, L.J.; Sheperd, A.C.; Smagala, A.M.; Endres, K.M.; Bessett, C.A.; Ranjit, N.K.; Glancey, J. The Effect of Preservatives Based on Propionic Acid on the Fermentation and Aerobic Stability of Corn Silage and a Total Mixed Ration. J. Dairy Sci. 1998, 81, 1322–1330. [Google Scholar] [CrossRef]
- He, L.; Zhou, W.; Wang, Y.; Wang, C.; Chen, X.; Zhang, Q. Effect of applying lactic acid bacteria and cellulase on the fermentation quality, nutritive value, tannins profile and in vitro digestibility of Neolamarckia cadamba leaves silage. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1429–1436. [Google Scholar] [CrossRef]
- Kaewpila, C.; Thip-uten, S.; Cherdthong, A.; Khota, W. Impact of Cellulase and Lactic Acid Bacteria Inoculant to Modify Ensiling Characteristics and In Vitro Digestibility of Sweet Corn Stover and Cassava Pulp Silage. Agriculture 2021, 11, 66. [Google Scholar] [CrossRef]
- McDonald, P.; Henderson, A.R.; Heron, S.J.E. The Biochemistry of Silage, 2nd ed.; Chalcombe Publications: Marlow, UK, 1991. [Google Scholar]
- Silva, V.P.; Pereira, O.G.; Leandro, E.S.; Da Silva, T.C.; Ribeiro, K.G.; Mantovani, H.C.; Santos, S.A. Effects of lactic acid bacteria with bacteriocinogenic potential on the fermentation profile and chemical composition of alfalfa silage in tropical conditions. J. Dairy Sci. 2016, 99, 1895–1902. [Google Scholar] [CrossRef] [Green Version]
- Kwak, W.S.; Kim, Y.I.; Seok, J.S.; Oh, Y.K.; Lee, S.M. Molasses and microbial inoculants improve fermentability and silage quality of cotton waste-based spent mushroom substrate. Bioresour. Technol. 2009, 100, 1471–1473. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yu, Z.; Yang, H.; Na, R.S. The effects of stage of growth and additives with or without cellulase on fermentation and in vitro degradation characteristics of Leymus chinensis silage. Grass Forage Sci. 2016, 71, 595–606. [Google Scholar] [CrossRef]
- Davies, D.R.; Merry, R.J.; Williams, A.P.; Bakewell, E.; Leemans, D.; Tweed, J. Proteolysis during Ensilage of Forages Varying in Soluble Sugar Content. J. Dairy Sci. 1998, 81, 444–453. [Google Scholar] [CrossRef]
- Huo, W.; Wang, X.; Wei, Z.; Zhang, H.; Liu, Q.; Zhang, S.; Wang, C.; Chen, L.; Xu, Q.; Guo, G. Effect of lactic acid bacteria on the ensiling characteristics and in vitro ruminal fermentation parameters of alfalfa silage. Ital. J. Anim. Sci. 2021, 20, 623–631. [Google Scholar] [CrossRef]
- Selmer-Olsen, I.; Henderson, A.R.; Robertson, S.; McGinn, R. Cell wall degrading enzymes for silage. The fermentation of enzyme-treated ryegrass in laboratory silos. Grass Forage Sci. 1993, 48, 45–54. [Google Scholar] [CrossRef]
- Xing, L.; Chen, L.J.; Han, L.J. The effect of an inoculant and enzymes on fermentation and nutritive value of sorghum straw silages. Bioresour. Technol. 2009, 100, 488–491. [Google Scholar] [CrossRef]
- Wang, M.; Wang, L.; Yu, Z. Fermentation dynamics and bacterial diversity of mixed lucerne and sweet corn stalk silage ensiled at six ratios. Grass Forage Sci. 2019, 74, 264–273. [Google Scholar] [CrossRef]
- Filya, I. The Effect of Lactobacillus buchneri and Lactobacillus plantarum on the Fermentation, Aerobic Stability, and Ruminal Degradability of Low Dry Matter Corn and Sorghum Silages. J. Dairy Sci. 2003, 86, 3575–3581. [Google Scholar] [CrossRef] [Green Version]
- Huisden, C.M.; Adesogan, A.T.; Kim, S.C.; Ososanya, T. Effect of applying molasses or inoculants containing homofermentative or heterofermentative bacteria at two rates on the fermentation and aerobic stability of corn silage. J. Dairy Sci. 2009, 92, 690–697. [Google Scholar] [CrossRef] [Green Version]
- Pahlow, G.; Muck, R.E.; Driehuis, F.; Elferink, S.J.W.H.O.; Spoelstra, S.F. Silage Science and Technology; American Society of Agronomy, Crop Science Society of America, Soil Science Society of America: Madison, WI, USA, 2003; pp. 31–93. [Google Scholar]
- Fang, J.; Cao, Y.; Matsuzaki, M.; Suzuki, H. Effects of apple pomace proportion levels on the fermentation quality of total mixed ration silage and its digestibility, preference and ruminal fermentation in beef cows. Anim. Sci. J. 2016, 87, 217–223. [Google Scholar] [CrossRef]
- Contreras-Govea, F.E.; Muck, R.E.; Mertens, D.R.; Weimer, P.J. Microbial inoculant effects on silage and in vitro ruminal fermentation, and microbial biomass estimation for alfalfa, bmr corn, and corn silages. Anim. Feed Sci. Technol. 2011, 163, 2–10. [Google Scholar] [CrossRef]
- Dehghani, M.R.; Weisbjerg, M.R.; Hvelplund, T.; Kristensen, N.B. Effect of enzyme addition to forage at ensiling on silage chemical composition and NDF degradation characteristics. Livest. Sci. 2012, 150, 51–58. [Google Scholar] [CrossRef]
- Pholsen, S.; Khota, W.; Pang, H.; Higgs, D.; Cai, Y. Characterization and application of lactic acid bacteria for tropical silage preparation. Anim. Sci. J. 2016, 87, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Tabacco, E.; Righi, F.; Quarantelli, A.; Borreani, G. Dry matter and nutritional losses during aerobic deterioration of corn and sorghum silages as influenced by different lactic acid bacteria inocula. J. Dairy Sci. 2011, 94, 1409–1419. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Duniere, L.; Lynch, J.; McAllister, T.; Baah, J.; Wang, Y. Impact of ferulic acid esterase producing lactobacilli and fibrolytic enzymes on conservation characteristics, aerobic stability and fiber degradability of barley silage. Anim. Feed Sci. Technol. 2015, 207, 62–74. [Google Scholar] [CrossRef]
- Khota, W.; Pholsen, S.; Higgs, D.; Cai, Y. Natural lactic acid bacteria population of tropical grasses and their fermentation factor analysis of silage prepared with cellulase and inoculant. J. Dairy Sci. 2016, 99, 9768–9781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mertens, D.R. Creating a System for Meeting the Fiber Requirements of Dairy Cows. J. Dairy Sci. 1997, 80, 1463–1481. [Google Scholar] [CrossRef] [PubMed]
- Rinne, M.; Winquist, E.; Pihlajaniemi, V.; Niemi, P.; Seppälä, A.; Siika-Aho, M. Fibrolytic enzyme treatment prior to ensiling increased press-juice and crude protein yield from grass silage. Bioresour. Technol. 2020, 299, 122572. [Google Scholar] [CrossRef]
- Bala, P.; Malik, R.; Srinivas, B. Effect of fortifying concentrate supplement with fibrolytic enzymes on nutrient utilization, milk yield and composition in lactating goats. Anim. Sci. J. 2009, 80, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Cai, Y.; Takahashi, T.; Yoshida, N.; Tohno, M.; Uegaki, R.; Nonaka, K.; Terada, F. Effect of lactic acid bacteria inoculant and beet pulp addition on fermentation characteristics and in vitro ruminal digestion of vegetable residue silage. J. Dairy Sci. 2011, 94, 3902–3912. [Google Scholar] [CrossRef] [Green Version]
- Refat, B.; Christensen, D.A.; McKinnon, J.J.; Yang, W.; Beattie, A.D.; McAllister, T.A.; Eun, J.-S.; Abdel-Rahman, G.A.; Yu, P. Effect of fibrolytic enzymes on lactational performance, feeding behavior, and digestibility in high-producing dairy cows fed a barley silage–based diet. J. Dairy Sci. 2018, 101, 7971–7979. [Google Scholar] [CrossRef] [PubMed]
- Borreani, G.; Tabacco, E. The relationship of silage temperature with the microbiological status of the face of corn silage bunkers. J. Dairy Sci. 2010, 93, 2620–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danner, H.; Holzer, M.; Mayrhuber, E.; Braun, R. Acetic Acid Increases Stability of Silage under Aerobic Conditions. Appl. Environ. Microbiol. 2003, 69, 562–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, L.M.; Harrison, J.H.; Davidson, D.; Mahanna, W.C.; Shinners, K.; Linder, D. Corn Silage Management: Effects of Maturity, Inoculation, and Mechanical Processing on Pack Density and Aerobic Stability. J. Dairy Sci. 2002, 85, 434–444. [Google Scholar] [CrossRef]
- Wilkinson, J.M.; Davies, D.R. The aerobic stability of silage: Key findings and recent developments. Grass Forage Sci. 2013, 68, 1–19. [Google Scholar] [CrossRef]
- Kleinschmit, D.H.; Schmidt, R.J.; Kung, L., Jr. The Effects of Various Antifungal Additives on the Fermentation and Aerobic Stability of Corn Silage. J. Dairy Sci. 2005, 88, 2130–2139. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Benno, Y.; Ogawa, M.; Kumai, S. Effect of Applying Lactic Acid Bacteria Isolated from Forage Crops on Fermentation Characteristics and Aerobic Deterioration of Silage. J. Dairy Sci. 1999, 82, 520–526. [Google Scholar] [CrossRef]
- Gandra, J.R.; Oliveira, E.R.; Gandra, E.R.D.S.; Takiya, C.S.; de Goes, R.H.T.B.; Oliveira, K.M.P.; Silveira, K.A.; Araki, H.M.C.; Orbach, N.D.; Vasquez, D.N. Inoculation of Lactobacillus buchneri alone or with Bacillus subtilis and total losses, aerobic stability, and microbiological quality of sunflower silage. J. Appl. Anim. Res. 2016, 45, 609–614. [Google Scholar] [CrossRef]
- Chen, L.; Guo, G.; Yuan, X.; Shimojo, M.; Yu, C.; Shao, T. Effect of Applying Molasses and Propionic Acid on Fermentation Quality and Aerobic Stability of Total Mixed Ration Silage Prepared with Whole-plant Corn in Tibet. Asian-Australas. J. Anim. Sci. 2014, 27, 349–356. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flammulina velutipes | Pleurotus eryngii | ||
|---|---|---|---|
| Corncob meal (%) | 37 | Corncob meal (%) | 23 |
| Rice bran (%) | 38 | Corn straw powder (%) | 18 |
| Sawdust (%) | 8 | Sawdust (%) | 39 |
| Soybean hulls (%) | 5 | ||
| Cottonseed hull (%) | 5 | Soybean meal (%) | 6 |
| Brewer’s grain (%) | 5 | Corn meal (%) | 12 |
| Lime (%) | 1 | Lime (%) | 1 |
| Calcium carbonate (%) | 1 | Calcium carbonate (%) | 1 |
| Total (%) | 100 | Total (%) | 100 |
| Spent Mushroom Substrate † | ||||
|---|---|---|---|---|
| Item ‡ | F-SMS | P-SMS | SEM | p-Value |
| Chemical composition and buffering capacity | ||||
| Dry matter (g kg−1 FW) | 492 | 504 | 0.351 | 0.000 |
| Organic matter (g kg−1 DM) | 872 | 903 | 0.677 | 0.000 |
| Crude protein (g kg−1 DM) | 90.4 | 80.7 | 0.664 | 0.000 |
| Neutral detergent fibre (g kg−1 DM) | 729 | 727 | 4.112 | 0.684 |
| Acid detergent fibre (g kg−1 DM) | 428 | 497 | 4.141 | 0.000 |
| Acid detergent lignin (g kg−1 DM) | 111 | 176 | 3.164 | 0.000 |
| Water-soluble carbohydrate (g kg−1 DM) | 24.4 | 34.2 | 0.120 | 0.000 |
| Calcium (g kg−1 DM) | 17.0 | 16.5 | 0.896 | 0.609 |
| Phosphorus (g kg−1 DM) | 7.31 | 1.82 | 0.247 | 0.000 |
| Buffering capacity (mEq kg−1 DM) | 282 | 157 | 12.209 | 0.001 |
| Energy | ||||
| GE (MJ kg−1 DM) | 19.2 | 18.8 | 0.034 | 0.000 |
| DE (MJ kg−1 DM) | 10.3 | 7.69 | 0.090 | 0.000 |
| ME (MJ kg−1 DM) | 8.30 | 6.18 | 0.071 | 0.000 |
| NEm (MJ kg−1 DM) | 5.59 | 4.00 | 0.059 | 0.000 |
| NEl (MJ kg−1 DM) | 4.69 | 3.35 | 0.047 | 0.000 |
| NEg (MJ kg−1 DM) | 2.82 | 1.63 | 0.051 | 0.000 |
| Microbial counts | ||||
| Lactic acid bacteria (log10 cfu g−1 FW) | 4.68 | 4.30 | 0.032 | 0.000 |
| Yeasts (log10 cfu g−1 FW) | 3.10 | 4.20 | 0.040 | 0.000 |
| Moulds (log10 cfu g−1 FW) | 0.21 | 2.33 | 0.072 | 0.000 |
| Item ‡ | S-Silage § | Additives † | SEM | p-Value | Significance of Main Effects and Interactions | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | L | E | M | L | E | L × E | ||||
| pH value | F-silage | 4.40 b | 4.26 a | 4.24 a | 4.20 a | 0.033 | 0.002 | 0.005 | 0.002 | 0.072 |
| P-silage | 4.21 b | 3.96 a | 3.95 a | 3.93 a | 0.031 | 0.000 | 0.000 | 0.000 | 0.001 | |
| SEM | 0.041 | 0.027 | 0.032 | 0.025 | ||||||
| p-value | 0.041 | 0.000 | 0.001 | 0.000 | ||||||
| LA (g kg−1 DM) | F-silage | 56.9 a | 77.2 b | 67.4 b | 67.9 b | 0.318 | 0.002 | 0.002 | 0.802 | 0.002 |
| P-silage | 118 | 121 | 122 | 126 | 0.889 | 0.839 | 0.596 | 0.488 | 0.947 | |
| SEM | 0.624 | 0.198 | 0.861 | 0.784 | ||||||
| p-value | 0.001 | 0.000 | 0.020 | 0.002 | ||||||
| AA (g kg−1 DM) | F-silage | 144 | 151 | 141 | 151 | 1.554 | 0.887 | 0.462 | 0.925 | 0.886 |
| P-silage | 2.72 | 0.90 | 0.94 | 1.04 | 0.149 | 0.583 | 0.443 | 0.460 | 0.393 | |
| SEM | 1.004 | 0.273 | 1.635 | 1.059 | ||||||
| p-value | 0.000 | 0.000 | 0.013 | 0.000 | ||||||
| PA (g kg−1 DM) | F-silage | 19.5 | 20.6 | 18.3 | 20.4 | 0.295 | 0.850 | 0.456 | 0.757 | 0.793 |
| P-silage | 3.44 | 4.50 | 5.91 | 4.70 | 0.112 | 0.249 | 0.903 | 0.123 | 0.191 | |
| SEM | 0.134 | 0.101 | 0.382 | 0.158 | ||||||
| p-value | 0.000 | 0.000 | 0.032 | 0.001 | ||||||
| BA (g kg−1 DM) | F-silage | 5.01 | 5.00 | 11.2 | 9.76 | 0.317 | 0.184 | 0.757 | 0.040 | 0.757 |
| P-silage | 4.33 | 4.17 | 5.52 | 4.18 | 0.121 | 0.638 | 0.398 | 0.505 | 0.505 | |
| SEM | 0.110 | 0.062 | 0.346 | 0.308 | ||||||
| p-value | 0.577 | 0.250 | 0.177 | 0.210 | ||||||
| AN (g kg−1 TN) | F-silage | 12.8 b | 10.9 ab | 10.4 ab | 9.40 a | 0.085 | 0.023 | 0.048 | 0.011 | 0.490 |
| P-silage | 11.6 b | 8.06 a | 9.84 ab | 9.92 ab | 0.068 | 0.007 | 0.008 | 0.894 | 0.006 | |
| SEM | 0.107 | 0.103 | 0.028 | 0.029 | ||||||
| p-value | 0.324 | 0.050 | 0.130 | 0.140 | ||||||
| Item ‡ | S-Silage § | Additives † | SEM | p-Value | Significance of Main Effects and Interactions | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | L | E | M | L | E | L × E | ||||
| DM (g kg−1 FW) | F-silage | 414 | 414 | 416 | 412 | 0.312 | 0.705 | 0.398 | 0.965 | 0.446 |
| P-silage | 457 a | 462 b | 462 b | 460 b | 0.141 | 0.015 | 0.084 | 0.134 | 0.007 | |
| SEM | 0.322 | 0.122 | 0.324 | 0.100 | ||||||
| p-value | 0.000 | 0.000 | 0.000 | 0.000 | ||||||
| OM (g kg−1 DM) | F-silage | 885 | 887 | 880 | 876 | 1.184 | 0.803 | 0.891 | 0.382 | 0.740 |
| P-silage | 899 | 898 | 897 | 899 | 0.130 | 0.234 | 0.218 | 0.469 | 0.128 | |
| SEM | 1.476 | 0.358 | 0.595 | 0.423 | ||||||
| p-value | 0.457 | 0.033 | 0.054 | 0.006 | ||||||
| CP (g kg−1 DM) | F-silage | 115 | 113 | 120 | 119 | 0.555 | 0.612 | 0.737 | 0.219 | 0.928 |
| P-silage | 88.2 a | 88.8 ab | 92.9 ab | 95.3 b | 0.216 | 0.032 | 0.369 | 0.006 | 0.564 | |
| SEM | 0.381 | 0.510 | 0.500 | 0.230 | ||||||
| p-value | 0.002 | 0.009 | 0.006 | 0.001 | ||||||
| NDF (g kg−1 DM) | F-silage | 690 | 694 | 668 | 662 | 1.765 | 0.262 | 0.910 | 0.063 | 0.689 |
| P-silage | 682 ab | 650 a | 684 bc | 717 c | 1.035 | 0.002 | 0.944 | 0.002 | 0.002 | |
| SEM | 0.213 | 0.855 | 2.295 | 1.529 | ||||||
| p-value | 0.022 | 0.007 | 0.527 | 0.022 | ||||||
| ADF (g kg−1 DM) | F-silage | 402 | 406 | 393 | 389 | 0.886 | 0.260 | 0.965 | 0.067 | 0.566 |
| P-silage | 464 b | 445 a | 465 b | 478 b | 0.469 | 0.001 | 0.472 | 0.001 | 0.001 | |
| SEM | 0.281 | 0.461 | 1.134 | 0.656 | ||||||
| p-value | 0.000 | 0.001 | 0.003 | 0.000 | ||||||
| ADL (g kg−1 DM) | F-silage | 135 ab | 150 b | 95.3 a | 101 a | 1.372 | 0.011 | 0.327 | 0.002 | 0.658 |
| P-silage | 188 c | 183 bc | 159 ab | 156 a | 0.819 | 0.009 | 0.562 | 0.001 | 0.828 | |
| SEM | 1.760 | 0.783 | 0.930 | 0.727 | ||||||
| p-value | 0.040 | 0.013 | 0.002 | 0.002 | ||||||
| Item ‡ | S-Silage § | Additives † | SEM | p-Value | Significance of Main Effects and Interactions | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | L | E | M | L | E | L × E | ||||
| GE (MJ kg−1 DM) | F-silage | 19.3 | 19.3 | 19.3 | 19.1 | 0.251 | 0.813 | 0.468 | 0.607 | 0.778 |
| P-silage | 18.9 ab | 18.7 a | 18.9 ab | 19.0 b | 0.058 | 0.034 | 0.937 | 0.025 | 0.032 | |
| SEM | 0.343 | 0.094 | 0.060 | 0.048 | ||||||
| p-value | 0.288 | 0.006 | 0.002 | 0.040 | ||||||
| DE (MJ kg−1 DM) | F-silage | 11.1 | 10.9 | 11.3 | 11.2 | 0.316 | 0.664 | 0.698 | 0.272 | 0.773 |
| P-silage | 7.89 | 7.75 | 7.74 | 7.79 | 0.059 | 0.115 | 0.254 | 0.254 | 0.054 | |
| SEM | 0.297 | 0.063 | 0.310 | 0.134 | ||||||
| p-value | 0.007 | 0.000 | 0.000 | 0.001 | ||||||
| ME (MJ kg−1 DM) | F-silage | 8.82 | 8.69 | 8.95 | 8.98 | 0.262 | 0.668 | 0.787 | 0.279 | 0.671 |
| P-silage | 6.27 | 6.19 | 6.17 | 6.20 | 0.046 | 0.225 | 0.380 | 0.216 | 0.137 | |
| SEM | 0.247 | 0.039 | 0.262 | 0.100 | ||||||
| p-value | 0.008 | 0.000 | 0.008 | 0.001 | ||||||
| NEm (MJ kg−1 DM) | F-silage | 6.04 | 5.94 | 6.15 | 6.19 | 0.205 | 0.625 | 0.807 | 0.249 | 0.642 |
| P-silage | 4.07 | 4.01 | 4.00 | 4.01 | 0.037 | 0.277 | 0.461 | 0.193 | 0.193 | |
| SEM | 0.180 | 0.031 | 0.215 | 0.083 | ||||||
| p-value | 0.007 | 0.000 | 0.009 | 0.001 | ||||||
| NEl (MJ kg−1 DM) | F-silage | 5.05 | 4.96 | 5.14 | 5.17 | 0.171 | 0.624 | 0.810 | 0.250 | 0.633 |
| P-silage | 3.40 | 3.36 | 3.35 | 3.36 | 0.029 | 0.298 | 0.446 | 0.236 | 0.188 | |
| SEM | 0.150 | 0.024 | 0.180 | 0.069 | ||||||
| p-value | 0.007 | 0.000 | 0.009 | 0.001 | ||||||
| NEg (MJ kg−1 DM) | F-silage | 3.19 | 3.11 | 3.30 | 3.35 | 0.165 | 0.503 | 0.890 | 0.175 | 0.583 |
| P-silage | 1.66 | 1.63 | 1.61 | 1.62 | 0.026 | 0.265 | 0.356 | 0.129 | 0.356 | |
| SEM | 0.120 | 0.028 | 0.190 | 0.066 | ||||||
| p-value | 0.005 | 0.000 | 0.011 | 0.001 | ||||||
| Item ‡ | S-Silage § | Additives † | SEM | p-Value | Significance of Main Effects and Interactions | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | L | E | M | L | E | L × E | ||||
| IVDMD (g kg−1 DM) | F-silage | 516 ab | 512 a | 522 ab | 526 b | 4.383 | 0.034 | 0.968 | 0.066 | 0.568 |
| P-silage | 471 a | 487 b | 468 a | 461 a | 3.856 | 0.000 | 0.478 | 0.001 | 0.001 | |
| SEM | 0.218 | 0.358 | 0.870 | 0.504 | ||||||
| p-value | 0.000 | 0.001 | 0.003 | 0.000 | ||||||
| IVNDFD (g kg−1 DM) | F-silage | 445 a | 439 a | 483 b | 495 c | 0.437 | 0.000 | 0.345 | 0.000 | 0.020 |
| P-silage | 409 a | 416 ab | 430 b | 405 a | 0.500 | 0.006 | 0.040 | 0.225 | 0.002 | |
| SEM | 0.375 | 0.469 | 0.657 | 0.301 | ||||||
| p-value | 0.001 | 0.008 | 0.001 | 0.000 | ||||||
| IVADFD(g kg−1 DM) | F-silage | 353 a | 361 a | 431 b | 452 c | 0.479 | 0.000 | 0.003 | 0.000 | 0.085 |
| P-silage | 339 a | 341 a | 355 b | 329 a | 0.465 | 0.004 | 0.006 | 0.560 | 0.003 | |
| SEM | 0.352 | 0.340 | 0.556 | 0.587 | ||||||
| p-value | 0.017 | 0.004 | 0.000 | 0.000 | ||||||
| IVHCD(g kg−1 DM) | F-silage | 573 b | 549 a | 560 a | 557 a | 0.418 | 0.003 | 0.002 | 0.405 | 0.007 |
| P-silage | 560 a | 576 b | 589 c | 556 a | 0.382 | 0.000 | 0.012 | 0.138 | 0.000 | |
| SEM | 0.196 | 0.430 | 0.538 | 0.359 | ||||||
| p-value | 0.003 | 0.003 | 0.006 | 0.755 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yi, Q.; Wang, P.; Tang, H.; Yu, M.; Zhao, T.; Sheng, Z.; Luo, H. Fermentation Quality, In Vitro Digestibility, and Aerobic Stability of Ensiling Spent Mushroom Substrate with Microbial Additives. Animals 2023, 13, 920. https://0-doi-org.brum.beds.ac.uk/10.3390/ani13050920
Yi Q, Wang P, Tang H, Yu M, Zhao T, Sheng Z, Luo H. Fermentation Quality, In Vitro Digestibility, and Aerobic Stability of Ensiling Spent Mushroom Substrate with Microbial Additives. Animals. 2023; 13(5):920. https://0-doi-org.brum.beds.ac.uk/10.3390/ani13050920
Chicago/Turabian StyleYi, Qixuan, Peng Wang, Hongyu Tang, Meng Yu, Tianyue Zhao, Ziyang Sheng, and Hailing Luo. 2023. "Fermentation Quality, In Vitro Digestibility, and Aerobic Stability of Ensiling Spent Mushroom Substrate with Microbial Additives" Animals 13, no. 5: 920. https://0-doi-org.brum.beds.ac.uk/10.3390/ani13050920