
How Pendular Is Human Brachiation? When Form Does Not Follow Function
 ,
,
Abstract
:Simple Summary
Abstract
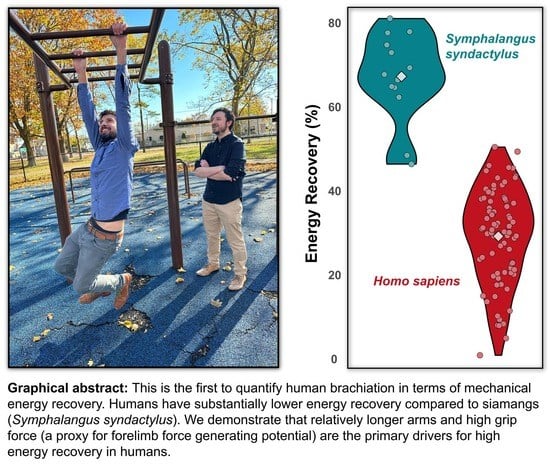
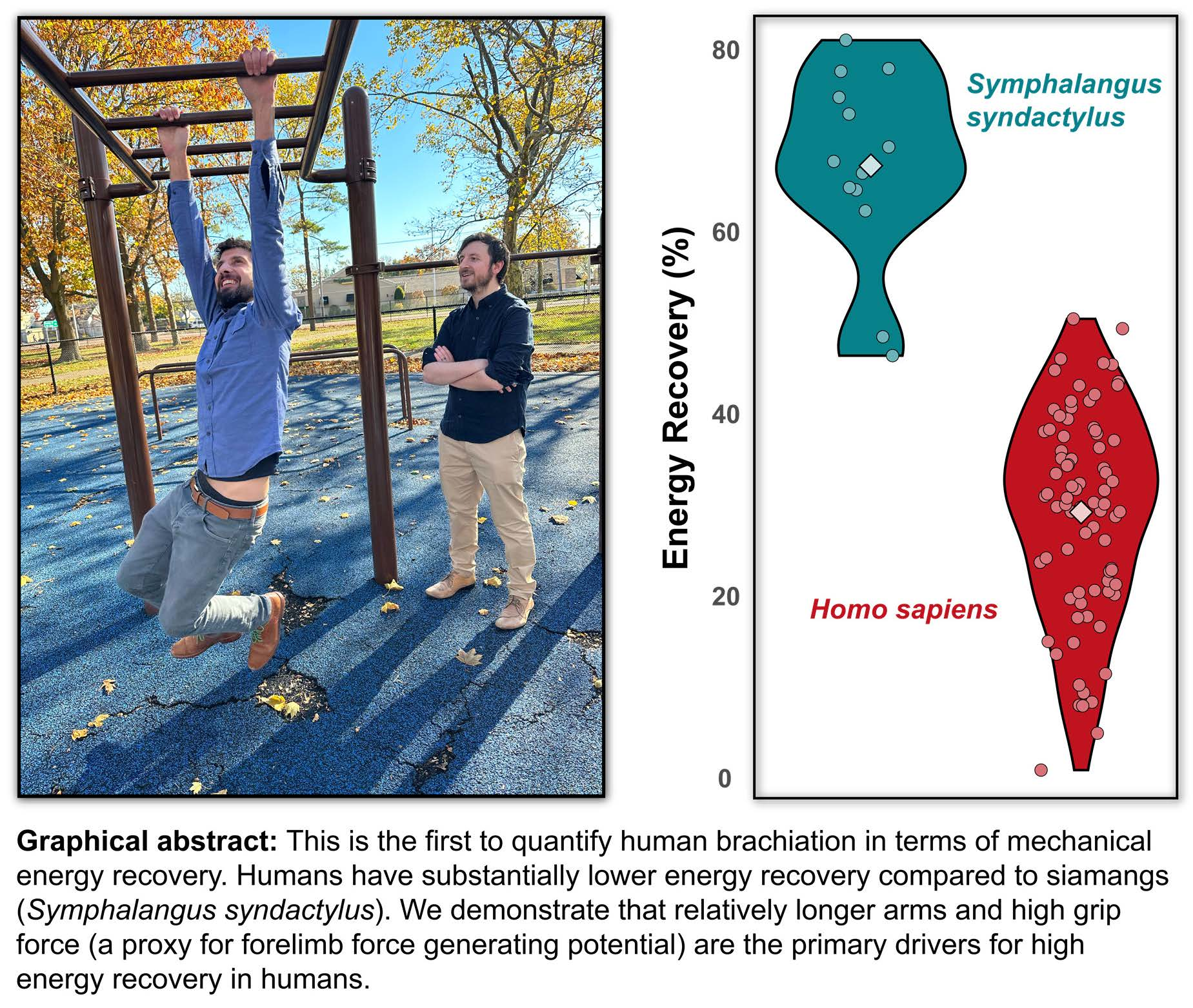
1. Introduction
2. Methods
2.1. Subjects and Experimental Procedure
2.2. Data Processing
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Byron, C.D.; Granatosky, M.C.; Covert, H.H. An Anatomical and Mechanical Analysis of the Douc Monkey (Genus Pygathrix), and Its Role in Understanding the Evolution osanof Brachiation. Am. J. Phys. Anthropol. 2017, 164, 801–820. [Google Scholar] [CrossRef] [PubMed]
- Avis, V. Brachiation: The Crucial Issue for Man’s Ancestry. Southwest J. Anthropol. 1962, 18, 119–148. [Google Scholar] [CrossRef]
- Dickinson, E.; Young, M.W.; Granatosky, M.C. Testing Mechanisms for Weight Support Distribution during Inverted Quadrupedalism in Primates. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2022, 337, 699–708. [Google Scholar] [CrossRef]
- Bertram, J.; Chang, Y.H. Mechanical Energy Oscillations of Two Brachiation Gaits: Measurement and Simulation. Am. J. Phys. Anthropol. 2001, 113, 201–216. [Google Scholar] [CrossRef]
- Granatosky, M.C.; Schmitt, D. The Mechanical Origins of Arm-Swinging. J. Hum. Evol. 2019, 130, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Granatosky, M.C. A Review of Locomotor Diversity in Mammals with Analyses Exploring the Influence of Substrate-Use, Body Mass, and Intermembral Index in Primates. J. Zool. 2018, 306, 207–216. [Google Scholar] [CrossRef]
- Larson, S.G. Parallel Evolution in the Hominoid Trunk and Forelimb. Evol. Anthropol. Issues News Rev. 1998, 6, 87–99. [Google Scholar] [CrossRef]
- Andrews, P.; Groves, C. Gibbons and Brachiation. In Gibbon and Siamang, Vol 4: Suspensory Behavior, Locomotion, and Other Behaviors of Captive Gibbons: Cognition; University of Michigan: Ann Arbor, MI, USA, 1976; pp. 167–218. [Google Scholar]
- Erikson, G. Brachiation in New World Monkeys and in Anthropoid Apes. Symp. Zool. Soc. Lond. 1963, 10, 135–164. [Google Scholar]
- Swartz, S. Pendular Mechanics and the Kinematics and Energetics of Brachiating Locomotion. Int. J. Primatol. 1989, 10, 387–418. [Google Scholar] [CrossRef]
- Swartz, S.M. Curvature of the Forelimb Bones of Anthropoid Primates: Overall Allometric Patterns and Specializations in Suspensory Species. Am. J. Phys. Anthropol. 1990, 83, 477–498. [Google Scholar] [CrossRef]
- Swartz, S.M.; Bertram, J.E.; Biewener, A.A. Telemetered in Vivo Strain Analysis of Locomotor Mechanics of Brachiating Gibbons. Nature 1989, 342, 270–272. [Google Scholar] [CrossRef]
- Hallgrimsson, B.; Swartz, S. Biomechanical Adaptation of Ulnar Cross-Sectional Morphology in Brachiating Primates. J. Morphol. 1995, 224, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Turnquist, J.E.; Schmitt, D.; Rose, M.D.; Cant, J.G. Pendular Motion in the Brachiation of Captive Lagothrix and Ateles. Am. J. Primatol. 1999, 48, 263–281. [Google Scholar] [CrossRef]
- Rein, T.R.; Harvati, K.; Harrison, T. Inferring the Use of Forelimb Suspensory Locomotion by Extinct Primate Species via Shape Exploration of the Ulna. J. Hum. Evol. 2015, 78, 70–79. [Google Scholar] [CrossRef]
- Migneault, D.; Chang, A.; Choi, E.; Doan, Q. Pediatric Falls: Are Monkey Bars Bad News? Cureus 2018, 10, e3548. [Google Scholar] [CrossRef] [PubMed]
- Churchill, S.E.; Holliday, T.W.; Carlson, K.J.; Jashashvili, T.; Macias, M.E.; Mathews, S.; Sparling, T.L.; Schmid, P.; de Ruiter, D.J.; Berger, L.R. The Upper Limb of Australopithecus Sediba. Science 2013, 340, 1233477. [Google Scholar] [CrossRef] [PubMed]
- Alba, D.M.; Almécija, S.; DeMiguel, D.; Fortuny, J.; de los Ríos, M.P.; Pina, M.; Robles, J.M.; Moyà-Solà, S. Miocene Small-Bodied Ape from Eurasia Sheds Light on Hominoid Evolution. Science 2015, 350, aab2625. [Google Scholar] [CrossRef]
- Alba, D.M.; Almécija, S.; Casanovas-Vilar, I.; Méndez, J.M.; Moyà-Solà, S. A Partial Skeleton of the Fossil Great Ape Hispanopithecus Laietanus from Can Feu and the Mosaic Evolution of Crown-Hominoid Positional Behaviors. PLoS ONE 2012, 7, e39617. [Google Scholar] [CrossRef]
- Preuschoft, H.; Demes, B. Biomechanics of Brachiation. In The Lesser Apes: Evolutionary and Behavioral Biology; Preuschoft, H., Chivers, D.J., Brockelman, W.Y., Creel, N., Eds.; Edinburgh Press Publishing: Edinburgh, UK, 1984; pp. 96–118. [Google Scholar]
- Preuschoft, H.; Demes, B. Biomechanic Determinants of Arm Length and Body Mass in Brachiators. Fortschr. Zool. 1985, 30, 39–43. [Google Scholar]
- Chang, Y.; Bertram, J.; Lee, D. External Forces and Torques Generated by the Brachiating White-Handed Gibbon (Hylobates Lar). Am. J. Phys. Anthropol. 2000, 113, 201–216. [Google Scholar] [CrossRef]
- Michilsens, F.; D’Août, K.; Aerts, P. How Pendulum-like Are Siamangs? Energy Exchange during Brachiation. Am. J. Phys. Anthropol. 2011, 145, 581–591. [Google Scholar] [CrossRef]
- Halliday, D.; Resnick, R.; Walker, J. Fundamentals of Physics; John Wiley & Sons: Hoboken, NJ, USA, 2013; ISBN 978-1-118-23071-8. [Google Scholar]
- Baker, G.L.; Blackburn, J.A. The Pendulum: A Case Study in Physics; OUP: Oxford, UK, 2008; ISBN 978-0-19-156530-4. [Google Scholar]
- Bertram, J.E.; Ruina, A.; Cannon, C.E.; Chang, Y.H.; Coleman, M.J. A Point-Mass Model of Gibbon Locomotion. J. Exp. Biol. 1999, 202, 2609–2617. [Google Scholar] [CrossRef] [PubMed]
- Fleagle, J.G. Dynamics of a Brachiating Siamang [Hylobates (Symphalangus) Syndactylus]. Nature 1974, 248, 259–260. [Google Scholar] [CrossRef] [PubMed]
- Usherwood, J.R. Understanding Brachiation: Insight from a Collisional Perspective. J. Exp. Biol. 2003, 206, 1631–1642. [Google Scholar] [CrossRef]
- Michilsens, F.; D’Août, K.; Vereecke, E.E.; Aerts, P. One Step beyond: Different Step-to-Step Transitions Exist during Continuous Contact Brachiation in Siamangs. Biol. Open 2012, 1, 411–421. [Google Scholar] [CrossRef]
- Granatosky, M.C. Kinetic and Kinematic Patterns of Arm-Swinging in the Red-Shanked Douc Langur (Pygathrix Nemaeus). J. Vietnam. Primatol. 2015, 2, 33–40. [Google Scholar]
- Abe, T.; Loenneke, J.P. Handgrip Strength Dominance Is Associated with Difference in Forearm Muscle Size. J. Phys. Ther. Sci. 2015, 27, 2147–2149. [Google Scholar] [CrossRef]
- Bosco, C.; Belli, A.; Astrua, M.; Tihanyi, J.; Pozzo, R.; Kellis, S.; Tsarpela, O.; Foti, C.; Manno, R.; Tranquilli, C. A Dynamometer for Evaluation of Dynamic Muscle Work. Europ. J. Appl. Physiol. 1995, 70, 379–386. [Google Scholar] [CrossRef]
- Bohannon, R.W. Dynamometer Measurements of Hand-Grip Strength Predict Multiple Outcomes. Percept. Mot. Skills 2001, 93, 323–328. [Google Scholar] [CrossRef]
- Bohannon, R.W. Hand-Grip Dynamometry Provides a Valid Indication of Upper Extremity Strength Impairment in Home Care Patients. J. Hand Ther. 1998, 11, 258–260. [Google Scholar] [CrossRef]
- Bardo, A.; Kivell, T.L.; Town, K.; Donati, G.; Ballieux, H.; Stamate, C.; Edginton, T.; Forrester, G.S. Get a Grip: Variation in Human Hand Grip Strength and Implications for Human Evolution. Symmetry 2021, 13, 1142. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Autumn, K.; Hsieh, S.T.; Dudek, D.M.; Chen, J.; Chitaphan, C.; Full, R.J. Dynamics of Geckos Running Vertically. J. Exp. Biol. 2006, 209, 260–272. [Google Scholar] [CrossRef]
- Blickhan, R.; Full, R. Locomotion Energetics of the Ghost Crab: II. Mechanics of the Centre of Mass During Walking and Running. J. Exp. Biol. 1987, 130, 155–174. [Google Scholar] [CrossRef]
- Cavagna, G.A.; Heglund, N.C.; Taylor, C.R. Mechanical Work in Terrestrial Locomotion: Two Basic Mechanisms for Minimizing Energy Expenditure. Am. J. Physiol. 1977, 233, R243–R261. [Google Scholar] [CrossRef] [PubMed]
- Heglund, N.C.; Fedak, M.A.; Taylor, C.R.; Cavagna, G.A. Energetics and Mechanics of Terrestrial Locomotion. IV. Total Mechanical Energy Changes as a Function of Speed and Body Size in Birds and Mammals. J. Exp. Biol. 1982, 97, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Reilly, S.M.; McElroy, E.J.; Odum, R.A.; Hornyak, V.A. Tuataras and Salamanders Show That Walking and Running Mechanics Are Ancient Features of Tetrapod Locomotion. Proc. R. Soc. Lond. B Biol. Sci. 2006, 273, 1563–1568. [Google Scholar] [CrossRef] [PubMed]
- Riskin, D.K.; Kendall, C.J.; Hermanson, J.W. The Crouching of the Shrew: Mechanical Consequences of Limb Posture in Small Mammals. PeerJ 2016, 4, e2131. [Google Scholar] [CrossRef]
- Young, M.W.; Flaim, N.D.; Yarbro, J.; Ragupathi, A.; Guru, N.; Dickinson, E.; Granatosky, M.C. Dynamics of Horizontal Walking and Vertical Climbing in the Australian Green Tree Frog (Ranoidea Caerulea). J. Exp. Biol. 2023, 226, jeb244833. [Google Scholar] [CrossRef]
- Cole, M.H.; van den Hoorn, W.; Kavanagh, J.K.; Morrison, S.; Hodges, P.W.; Smeathers, J.E.; Kerr, G.K. Concurrent Validity of Accelerations Measured Using a Tri-Axial Inertial Measurement Unit While Walking on Firm, Compliant and Uneven Surfaces. PLoS ONE 2014, 9, e98395. [Google Scholar] [CrossRef]
- Sosnoff, J.J.; Goldman, M.D.; Motl, R.W. Real-Life Walking Impairment in Multiple Sclerosis: Preliminary Comparison of Four Methods for Processing Accelerometry Data. Mult. Scler. 2010, 16, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Willey, J.S.; Biknevicius, A.R.; Reilly, S.M.; Earls, K.D. The Tale of the Tail: Limb Function and Locomotor Mechanics in Alligator Mississippiensis. J. Exp. Biol. 2004, 207, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. LmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. (Eds.) Formal Inference from More than One Model: Multimodel Inference (MMI). In Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002; pp. 149–205. ISBN 978-0-387-22456-5. [Google Scholar]
- R: Calculation of Akaike Weights/Relative Likelihoods/Delta-AICs. Available online: https://search.r-project.org/CRAN/refmans/qpcR/html/akaike.weights.html (accessed on 14 February 2023).
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 6th ed.; Freeman & Company: New York, NY, USA, 2012. [Google Scholar]
- Winter, B. Linear Models and Linear Mixed Effects Models in R with Linguistic Applications. arXiv 2013, arXiv:1308.5499. [Google Scholar]
- Parsons, P.; Taylor, C. Energetics of Brachiation versus Walking: A Comparison of a Suspended and an Inverted Pendulum Mechanism. Physiol. Zool. 1977, 50, 182–188. [Google Scholar] [CrossRef]
- Matthis, J.S.; Fajen, B.R. Humans Exploit the Biomechanics of Bipedal Gait during Visually Guided Walking over Complex Terrain. Proc. R. Soc. B Biol. Sci. 2013, 280, 20130700. [Google Scholar] [CrossRef] [PubMed]
- Nyakatura, J.A.; Andrada, E. A Mechanical Link Model of Two-Toed Sloths: No Pendular Mechanics during Suspensory Locomotion. Acta Theriol. 2013, 58, 83–93. [Google Scholar] [CrossRef]
- Brandwood, A.; Jayes, A.S.; Alexander, R.M. Incidence of Healed Fracture in the Skeletons of Birds, Molluscs and Primates. J. Zool. 1986, 208, 55–62. [Google Scholar] [CrossRef]
- Granatosky, M.C.; Schmitt, D.; Hanna, J. Comparison of Spatiotemporal Gait Characteristics between Vertical Climbing and Horizontal Walking in Primates. J. Exp. Biol. 2019, 222, jeb185702. [Google Scholar] [CrossRef]
- Lammers, A.R.; Zurcher, U. Stability during Arboreal Locomotion. In Theoretical Biomechanics; Klika, V., Ed.; InTech: Vienna, Austria, 2011. [Google Scholar]
- Payne, R.C.; Crompton, R.H.; Isler, K.; Savage, R.; Vereecke, E.E.; Günther, M.M.; Thorpe, S.K.S.; D’Août, K. Morphological Analysis of the Hindlimb in Apes and Humans. II. Moment Arms. J. Anat. 2006, 208, 725–742. [Google Scholar] [CrossRef] [PubMed]
- Myatt, J.P.; Crompton, R.H.; Payne-Davis, R.C.; Vereecke, E.E.; Isler, K.; Savage, R.; D’Août, K.; Günther, M.M.; Thorpe, S.K.S. Functional Adaptations in the Forelimb Muscles of Non-Human Great Apes. J. Anat. 2012, 220, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Jungers, W.L.; Stern, J.T. Preliminary Electromyographical Analysis of Brachiation in Gibbon and Spider Monkey. Int. J. Primatol. 1981, 2, 19–33. [Google Scholar] [CrossRef]
- Heglund, N.C.; Taylor, C.R. Speed, Stride Frequency and Energy Cost per Stride: How Do They Change with Body Size and Gait? J. Exp. Biol. 1988, 138, 301–318. [Google Scholar] [CrossRef] [PubMed]
- Griffin, T.M.; Roberts, T.J.; Kram, R. Metabolic Cost of Generating Muscular Force in Human Walking: Insights from Load-Carrying and Speed Experiments. J. Appl. Physiol. 2003, 95, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Gordon, K.E.; Ferris, D.P.; Kuo, A.D. Metabolic and Mechanical Energy Costs of Reducing Vertical Center of Mass Movement During Gait. Arch. Phys. Med. Rehabil. 2009, 90, 136–144. [Google Scholar] [CrossRef]
- Minetti, A.E.; Ardigo, L.P.; Reinach, E.; Saibene, F. The Relationship between Mechanical Work and Energy Expenditure of Locomotion in Horses. J. Exp. Biol. 1999, 202, 2329–2338. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Suleski, M.; Craig, J.M.; Kasprowicz, A.E.; Sanderford, M.; Li, M.; Stecher, G.; Hedges, S.B. TimeTree 5: An Expanded Resource for Species Divergence Times. Mol. Biol. Evol. 2022, 39, msac174. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Individual | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
|---|---|---|---|---|---|---|---|---|---|
| n | 19 | 10 | 5 | 10 | 6 | 5 | 7 | 8 | 10 |
| Height [m] | 1.7 | 1.75 | 1.68 | 1.85 | 1.83 | 1.75 | 1.75 | 1.93 | 1.79 |
| Weight [kg] | 65.77 | 85.73 | 66.22 | 79.38 | 102.15 | 74.39 | 80.2 | 100.24 | 97.52 |
| Forelimb Length [m] | 0.7 | 0.77 | 0.66 | 0.75 | 0.76 | 0.74 | 0.71 | 0.85 | 0.76 |
| Forelimb Length/Height | 0.41 | 0.44 | 0.39 | 0.4 | 0.42 | 0.42 | 0.41 | 0.44 | 0.43 |
| Grip Force/Body Weight | 0.77 | 0.53 | 0.47 | 0.57 | - | 0.42 | 0.64 | 0.36 | 0.61 |
| Avg Foreaft Velocity [m/s] | 0.59 ± 0.11 | 0.54 ± 0.04 | 0.49 ± 0.05 | 0.66 ± 0.06 | 0.44 ± 0.05 | 0.50 ± 0.04 | 0.68 ± 0.06 | 0.49 ± 0.08 | 0.70 ± 0.06 |
| Duty Factor [%] | 62.02 ± 4.61 | 71.95 ± 3.71 | 77.54 ± 2.59 | 73.87 ± 1.58 | 81.72 ± 2.19 | 79.72 ± 2.27 | 72.10 ± 2.5 | 76.36 ± 3.64 | 66.10 ± 3.13 |
| Stance Time [s] | 0.85 ± 0.17 | 0.84 ± 0.11 | 1.10 ± 0.14 | 0.78 ± 0.07 | 1.22 ± 0.14 | 1.10 ± 0.1 | 0.74 ± 0.09 | 0.97 ± 0.11 | 0.62 ± 0.06 |
| Swing Time [s] | 0.53 ± 0.16 | 0.32 ± 0.03 | 0.32 ± 0.03 | 0.27 ± 0.03 | 0.27 ± 0.02 | 0.28 ± 0.01 | 0.28 ± 0.01 | 0.30 ± 0.05 | 0.32 ± 0.03 |
| Stride Frequency [Hz] | 0.75 ± 0.15 | 0.87 ± 0.07 | 0.71 ± 0.07 | 0.96 ± 0.09 | 0.67 ± 0.06 | 0.73 ± 0.05 | 0.98 ± 0.09 | 0.79 ± 0.07 | 1.07 ± 0.07 |
| Stride Length [m] | 0.80 ± 0.19 | 0.62 ± 0.06 | 0.69 ± 0.00 | 0.69 ± 0.00 | 0.66 ± 0.05 | 0.69 ± 0.00 | 0.69 ± 0.00 | 0.62 ± 0.08 | 0.65 ± 0.03 |
| Energy Recovery [%] | 33.99 ± 10.42 | 36.06 ± 7.84 | 19.75 ± 5.45 | 15.63 ± 10.29 | 29.21 ± 10.74 | 28.39 ± 5.31 | 30.07 ± 6.55 | 28.08 ± 9.43 | 37.07 ± 11.08 |
| Predicted Pendulum Period [s] | 2.10 | 2.10 | 2.12 | 2.24 | 2.30 | 2.16 | 2.22 | 2.40 | 2.24 |
| Pendulum Period [s] | 1.39 ± 0.31 | 1.16 ± 0.10 | 1.41 ± 0.14 | 1.05 ± 0.10 | 1.49 ± 0.13 | 1.38 ± 0.10 | 1.03 ± 0.09 | 1.27 ± 0.12 | 0.94 ± 0.06 |
| Mediolateral Excursion [m] | 0.45 ± 0.17 | 0.25 ± 0.15 | 0.39 ± 0.17 | 0.40 ± 0.14 | 0.39 ± 0.18 | 0.37 ± 0.12 | 0.10 ± 0.08 | 0.40 ± 0.13 | 0.13 ± 0.05 |
| Vertical Excursion [m] | 0.24 ± 0.11 | 0.10 ± 0.06 | 0.31 ± 0.10 | 0.14 ± 0.06 | 0.38 ± 0.22 | 0.23 ± 0.15 | 0.17 ± 0.09 | 0.27 ± 0.12 | 0.13 ± 0.07 |
| Response Variable | Fixed Effect | Estimate | Standard Error | df | t Value | p-Value |
|---|---|---|---|---|---|---|
| Energy Recovery % | Average Fore-aft Velocity [m/s] | 10.86 | 31.32 | 74.00 | 0.35 | 0.730 |
| Forelimb/Height Ratio | 895.76 | 231.88 | 74.00 | 3.86 | <0.001 | |
| Weight [kg] | −0.03 | 0.34 | 74.00 | −0.10 | 0.920 | |
| Mediolateral Excursion [m] | −7.20 | 14.35 | 74.00 | −0.50 | 0.617 | |
| Vertical Excursion [m] | 31.88 | 21.27 | 74.00 | 1.50 | 0.138 | |
| Body weight adjusted grip force | 82.29 | 24.96 | 74.00 | 3.30 | 0.002 |
| ΔAIC | Forelimb Length | Grip Force | Vertical Excursion | Mediolateral Excursion | Fore-Aft Velocity | Body Mass |
|---|---|---|---|---|---|---|
| 0 | + | + | ||||
| 0.2274 | + | + | + | |||
| 1.7685 | + | + | + | + | ||
| 1.8837 | + | + | + | |||
| 1.8954 | + | + | + | + | ||
| 1.9957 | + | + | + | |||
| 2 | + | + | + | |||
| 2.1012 | + | + | + | + | ||
| Importance | 1.00 | 1.00 | 0.49 | 0.19 | 0.18 | 0.17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Young, M.W.; Virga, J.Q.; Kantounis, S.J.; Lynch, S.K.; Chernik, N.D.; Gustafson, J.A.; Cannata, M.J.; Flaim, N.D.; Granatosky, M.C. How Pendular Is Human Brachiation? When Form Does Not Follow Function. Animals 2023, 13, 1438. https://0-doi-org.brum.beds.ac.uk/10.3390/ani13091438
Young MW, Virga JQ, Kantounis SJ, Lynch SK, Chernik ND, Gustafson JA, Cannata MJ, Flaim ND, Granatosky MC. How Pendular Is Human Brachiation? When Form Does Not Follow Function. Animals. 2023; 13(9):1438. https://0-doi-org.brum.beds.ac.uk/10.3390/ani13091438
Chicago/Turabian StyleYoung, Melody W., James Q. Virga, Stratos J. Kantounis, Samantha K. Lynch, Noah D. Chernik, Jon A. Gustafson, Matthew J. Cannata, Nicholas D. Flaim, and Michael C. Granatosky. 2023. "How Pendular Is Human Brachiation? When Form Does Not Follow Function" Animals 13, no. 9: 1438. https://0-doi-org.brum.beds.ac.uk/10.3390/ani13091438