
Evidence for Beneficial Physiological Responses of the Land Snail Cornu aspersum to Probiotics’ (Lactobacillus plantarum) Dietary Intervention

 , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Maintenance
2.1.1. Snail Collection
2.1.2. Snail Feeding
2.1.3. Tissue Sampling
2.2. Analytical Procedures
2.2.1. RNA/DNA Ratio
2.2.2. Complementary DNA Synthesis
2.2.3. Real-Time Polymerase Chain Reaction
2.2.4. Immunoblotting/SDS-PAGE
2.3. Statistical Analysis
3. Results
3.1. Snail Growth and Mortality
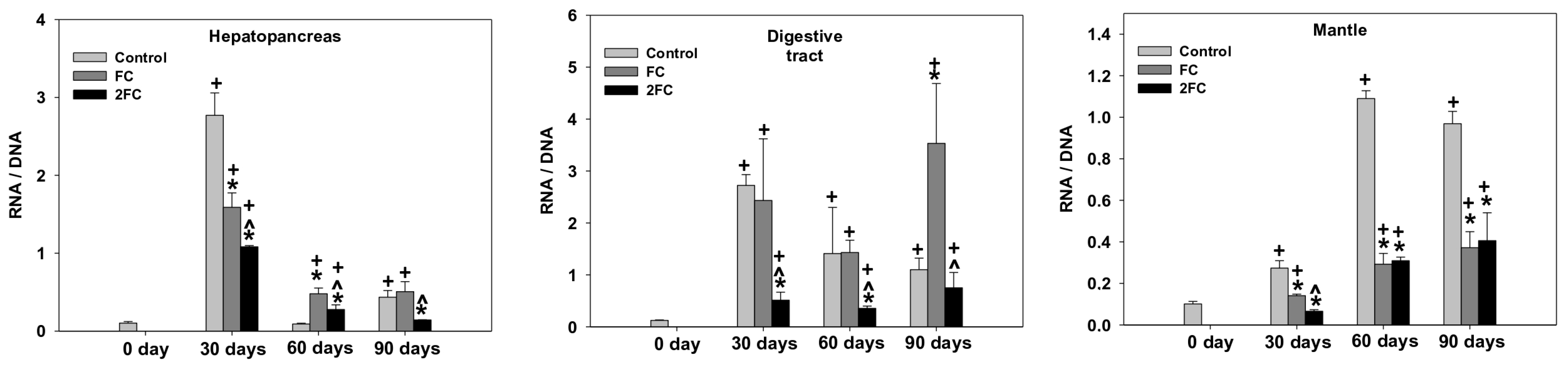
3.2. RNA/DNA Ratio
3.3. Expression of the ND2 Dehydrogenase Gene
3.4. HSP70 Relative Gene Expression
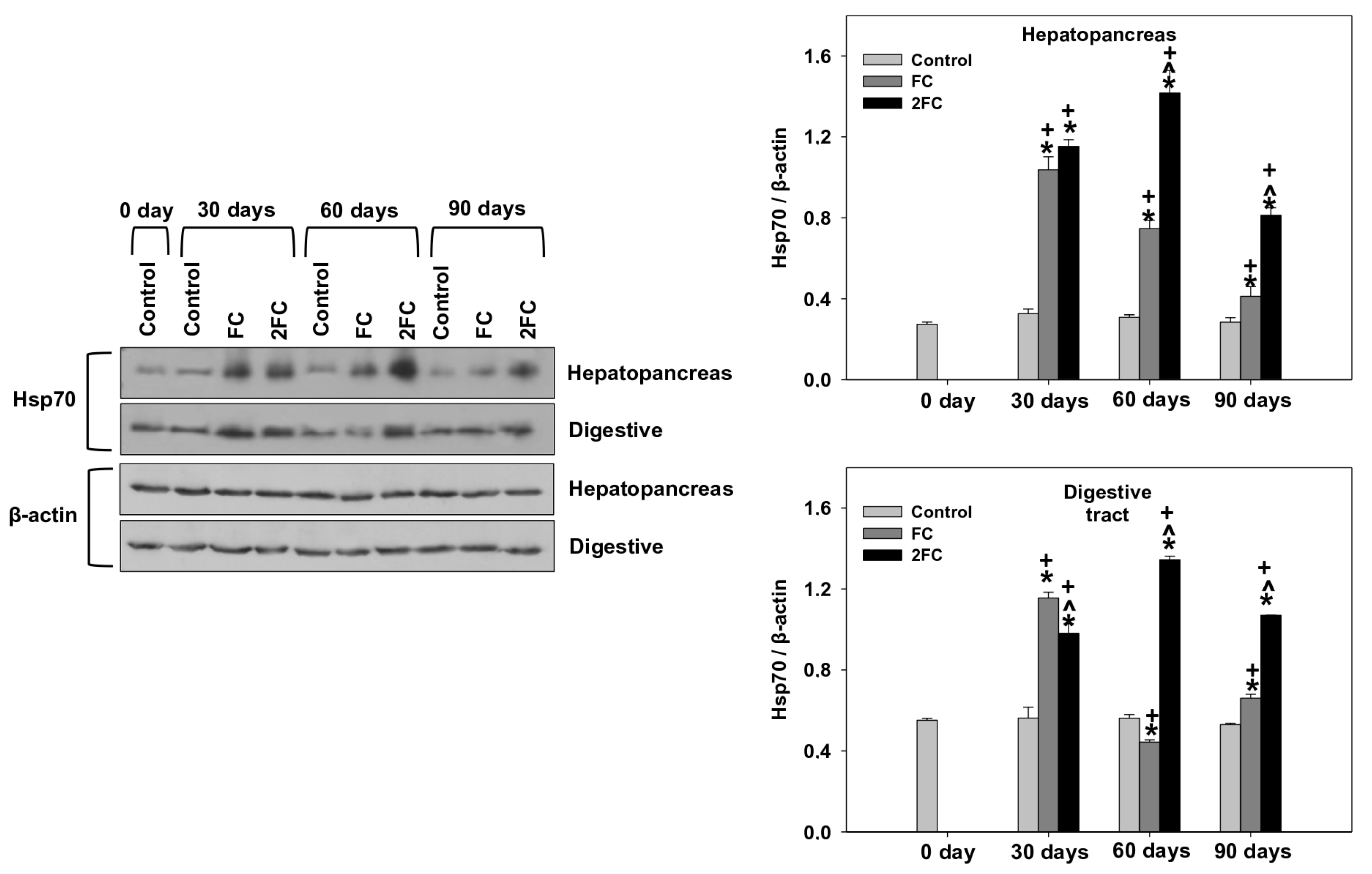
3.5. HSP70 and HSP90 Levels
3.6. Bax/Bcl-2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hatziioannou, M.; Issari, A.; Neofitou, C.; Aifadi, S.; Matsiori, S. Economic analysis and production techniques of snail farms in southern Greece. World J. Agric. Res. 2014, 2, 276–279. [Google Scholar] [CrossRef]
- Hatziioannou, M.; Exadaktilos, A.; Panagiotaki, P.; Lazaridou, M.; Neofitou, C. Setting Quality Standards for Farmed Snails Helix aspersa; Final Report, Pythagoras II; Ministry of Education and Religious Affairs: Volos, Greece, 2008; 168p. [Google Scholar]
- Staikou, A.; Lazaridou-Dimitriadou, M. Feeding experiments on and energy flux in a natural population of the edible snail Helix lucorum L. (Gastropoda: Pulmonata: Stylommatophora) in Greece. Malacologia 1989, 31, 217–227. [Google Scholar]
- Flari, V.; Charrier, M. Contribution to the study of carbohydrases in the digestive tract of the edible snail Helix lucorum L. (Gastropoda: Pulmonata: Stylommatophora) in relation to its age and its physiological state. Comp. Biochem. Physiol. 1992, 102, 363–372. [Google Scholar] [CrossRef]
- García, A.; Perea, J.; Martín, R.; Acero, R.; Mayoral, A.; Peña, F.; Luque, M. Effect of two diets on the growth of the Helix aspersa Müller during the juvenile stage. In Proceedings of the 56th Annual Meeting EAAP, Uppsala, Sweden, 5–8 June 2005; pp. 1–9. [Google Scholar]
- Gaggìa, F.; Mattarelli, P.; Biavati, B. Probiotics and prebiotics in animal feeding for safe food production. Int. J. Food Microbiol. 2010, 141, 15–28. [Google Scholar] [CrossRef]
- Yirga, H. The use of probiotics in animal nutrition. J. Probiotics Health 2015, 3, 132. [Google Scholar] [CrossRef]
- Schillinger, U.; Lücke, F.K. Antibacterial activity of Lactobacillus sake isolated from meat. Appl. Environ. Microbiol. 1989, 55, 1901–1906. [Google Scholar] [CrossRef]
- Silva, M.; Jacobus, N.V.; Deneke, C.; Gorbach, S.L. Antimicrobial substance from a human Lactobacillus strain. Antimicrob. Agents Chemother. 1987, 31, 1231–1233. [Google Scholar] [CrossRef] [PubMed]
- Fernández, M.F.; Boris, S.; Barbes, C. Probiotic properties of human lactobacilli strains to be used in the gastrointestinal tract. J. Appl. Microbiol. 2003, 94, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Ljungh, A.; Wadstrom, T. (Eds.) Lactobacillus Molecular Biology: From Genomics to Probiotics; Caister Academic Press: Poole, UK, 2009; pp. 3–58. [Google Scholar]
- Chaucheyras-Durand, F.; Durand, H. Probiotics in animal nutrition and health. Benef. Microbes 2010, 1, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Corcionivoschi, N.; Drinceanu, D.; Pop, I.M.; Stack, D.; Stef, L.; Julean, C.; Bourke, B. The effect of probiotics on animal health. Anim. Sci. Biotechnol. 2010, 43, 35–41. [Google Scholar]
- Fuller, R. Probiotics in human medicine. Gut 1991, 32, 439. [Google Scholar] [CrossRef]
- Nemati, M.; Omrani, G.R.; Ebrahimi, B.; Montazeri-Najafabady, N. The beneficial effects of probiotics via autophagy: A systematic review. BioMed Res. Int. 2021, 2021, 2931580. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, S.L. Between genotype and phenotype: Protein chaperones and evolvability. Nat. Rev. Genet. 2003, 4, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, S.L.; Lindquist, S. Hsp90 as a capacitor for morphological evolution. Nature 1998, 396, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, J.G.; Loeschcke, V. Larval crowding in Drosophila melanogaster induces Hsp70 expression, and leads to increased adult longevity and adult thermal stress resistance. J. Insect Physiol. 2001, 47, 1301–1307. [Google Scholar] [CrossRef] [PubMed]
- Hoseinifar, S.H.; Sun, Y.Z.; Zhou, Z.; Van Doan, H.; Davies, S.J.; Harikrishnan, R. Boosting immune function and disease bio-control through environment-friendly and sustainable approaches in finfish aquaculture: Herbal therapy scenarios. Rev. Fish Sci. Aquac. 2020, 28, 303–321. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Sun, Y.Z.; Wang, A.; Zhou, Z. Probiotics as means of diseases control in aquaculture, a review of current knowledge and future perspectives. Front. Microbiol. 2018, 9, 2429. [Google Scholar] [CrossRef] [PubMed]
- Wuertz, S.; Beça, F.; Kreuz, E.; Wanka, K.M.; Azeredo, R.; Machado, M.; Costas, B. Two probiotic candidates of the genus Psychrobacter modulate the immune response and disease resistance after experimental infection in Turbot (Scophthalmus maximus, Linnaeus 1758). Fishes 2023, 8, 144. [Google Scholar] [CrossRef]
- Kos, B.; Šušković, J.; Vuković, S.; Šimpraga, M.; Frece, J.; Matošić, S. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef]
- Kotzamanidis, C.; Kourelis, A.; Litopoulou-Tzanetaki, E.; Tzanetakis, N.; Yiangou, M. Evaluation of adhesion capacity, cell surface traits and immunomodulatory activity of presumptive probiotic Lactobacillus strains. Int. J. Food Microbiol. 2010, 140, 154–163. [Google Scholar] [CrossRef]
- Jones, S.E.; Versalovic, J. Probiotic Lactobacillus reuteri biofilms produce antimicrobial and anti-inflammatory factors. BMC Microbiol. 2009, 9, 1–9. [Google Scholar] [CrossRef]
- Dushku, E.; Ioannou, A.; Staikou, A.; Yiangou, M. Probiotic properties and immunomodulatory activity of gastrointestinal tract commensal bacterial strains isolated from the edible farmed snail Cornu aspersum maxima. Fish Shellfish Immunol. 2019, 92, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Dushku, E.; Kotzamanidis, C.; Avgousti, K.; Zdragas, A.; Vafeas, G.; Giantzi, V.; Staikou, A.; Yiangou, M. Listeria monocytogenes induced dysbiosis in snails and rebiosis achieved by administration of the gut commensal Lactobacillus plantarum Sgs14 strain. Fish Shellfish Immunol. 2020, 104, 337–346. [Google Scholar] [CrossRef]
- Lucas, A.; Beninger, P.G. The use of physiological condition indices in marine bivalve aquaculture. Aquaculture 1985, 44, 187–200. [Google Scholar] [CrossRef]
- Zhou, B.S.; Wu, R.S.S.; Randall, D.J.; Lam, P.K.S. Bioenergetics and RNA/DNA ratios in the common carp (Cyprinus carpio) under hypoxia. J. Comp. Physiol. B 2001, 171, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Foley, C.J.; Bradley, D.L.; Höök, T.O. A review and assessment of the potential use of RNA: DNA ratios to assess the condition of entrained fish larvae. Ecol. Indic. 2016, 60, 346–357. [Google Scholar] [CrossRef]
- Hoter, A.; El-Sabban, M.E.; Naim, H.Y. The HSP90 Family: Structure, Regulation, Function, and Implications in Health and Disease. Int. J. Mol. Sci. 2018, 19, 2560. [Google Scholar] [CrossRef] [PubMed]
- Khodapasand, E.; Jafarzadeh, N.; Farrokhi, F.; Kamalidehghan, B.; Houshmand, M. Is Bax/Bcl-2 ratio considered as a prognostic marker with age and tumor location in colorectal cancer? Iran. Biomed. J. 2015, 19, 69. [Google Scholar] [CrossRef]
- He, T.; Long, S.; Mahfuz, S.; Wu, D.; Wang, X.; Wei, X.; Piao, X. Effects of probiotics as antibiotics substitutes on growth performance, serum biochemical parameters, intestinal morphology, and barrier function of broilers. Animals 2019, 9, 985. [Google Scholar] [CrossRef]
- Bouétard, A.; Besnard, A.L.; Vassaux, D.; Lagadic, L.; Coutellec, M.A. Impact of the redox-cycling herbicide diquat on transcript expression and antioxidant enzymatic activities of the freshwater snail Lymnaeastagnalis. Aquat. Toxicol. 2013, 126, 256–265. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Moura, C.S.; Lollo, P.C.B.; Morato, P.N.; Amaya-Farfan, J. Dietary nutrients and bioactive substances modulate heat shock protein (HSP) expression: A review. Nutrients 2018, 10, 683. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. The role of probiotics, prebiotics and synbiotics in animal nutrition. Gut Pathog. 2018, 10, 21. [Google Scholar] [CrossRef] [PubMed]
- Price, R.; Jayeola, V.; Niedermeyer, J.; Parsons, C.; Kathariou, S. The Listeria monocytogenes key virulence determinants hly and prfA are involved in biofilm formation and aggregation but not colonization of fresh produce. Pathogens 2018, 7, 18. [Google Scholar] [CrossRef]
- Al-Shawi, S.G.; Dang, D.S.; Yousif, A.Y.; Al-Younis, Z.K.; Najm, T.A.; Matarneh, S.K. The potential use of probiotics to improve animal health, efficiency, and meat quality: A Review. Agriculture 2020, 10, 452. [Google Scholar] [CrossRef]
- Lambo, M.T.; Chang, X.; Liu, D. The recent trend in the use of multistrain probiotics in livestock production: An overview. Animals 2021, 11, 2805. [Google Scholar] [CrossRef] [PubMed]
- Bhogoju, S.; Nahashon, S. Recent advances in probiotic application in animal health and nutrition: A review. Agriculture 2022, 12, 304. [Google Scholar] [CrossRef]
- Nalla, K.; Manda, N.K.; Dhillon, H.S.; Kanade, S.R.; Rokana, N.; Hess, M.; Puniya, A.K. Impact of probiotics on dairy production efficiency. Front. Microbiol. 2022, 13, 805963. [Google Scholar] [CrossRef]
- Mansilla, F.I.; Miranda, M.H.; Uezen, J.D.; Maldonado, N.C.; Villar, M.A.U.; Merino, L.A.; Vingolo, G.M.; Nader-Macias, M.E.F. Effect of probiotic lactobacilli supplementation on growth parameters, blood profile, productive performance, and fecal microbiology in feedlot cattle. Res. Vet. Sci. 2023, 155, 76–87. [Google Scholar] [CrossRef]
- Kotzamanidis, C.; Malousi, A.; Dushku, E.; Dobly, A.; De Keersmaecker, S.C.; Roosens, N.H.; Karathodorou, A.; Staikou, A.; Zdragas, A.; Yiangou, M. Listeria monocytogenes isolates from Cornu aspersum snails: Whole genome-based characterization and host-pathogen interactions in a snail infection model. Fish Shellfish Immunol. 2022, 123, 469–478. [Google Scholar] [CrossRef]
- Parlapani, F.F.; Neofitou, C.; Boziaris, I.S. Microbiological quality of raw and processed wild and cultured edible snails. J. Sci. Food Agric. 2014, 94, 768–772. [Google Scholar] [CrossRef]
- Clemmesen, C. The effect of food availability, age or size on the RNA/DNA ratio of individually measured herring larvae: Laboratory calibration. Mar. Biol. 1994, 118, 377–382. [Google Scholar] [CrossRef]
- Bulow, F.J. RNA-DNA ratios as indicators of growth rates in fish: A review. In The Age and Growth of Fish; Summerfelt, R.C., Hall, G.E., Eds.; The Iowa State University Press: Ames, IA, USA, 1987; pp. 45–64. [Google Scholar]
- Kim, D.H.; Baek, J.M.; Lee, J.H.; Kim, B.R.; Yoon, S.J.; Kim, J.H. Food effect on the diel variations and starvation of the melania snail Semisulcospiragottschei using RNA/DNA ratios. Fish Aquat. Sci. 2011, 14, 411–416. [Google Scholar] [CrossRef]
- Ringø, E.; Harikrishnan, R.; Soltani, M.; Ghosh, K. The Effect of Gut Microbiota and Probiotics on Metabolism in Fish and Shrimp. Animals 2022, 12, 3016. [Google Scholar] [CrossRef]
- Imdad, S.; Lim, W.; Kim, J.H.; Kang, C. Intertwined Relationship of Mitochondrial Metabolism, Gut Microbiome and Exercise Potential. Int. J. Mol. Sci. 2022, 23, 2679. [Google Scholar] [CrossRef]
- Li, Y.; Yang, S.; Jin, X.; Li, D.; Lu, J.; Wang, X.; Wu, M. Mitochondria as novel mediators linking gut microbiota to atherosclerosis that is ameliorated by herbal medicine: A review. Front. Pharmacol. 2023, 14, 1082817. [Google Scholar] [CrossRef]
- Xu, Y.; Zheng, G.; Dong, S.; Liu, G.; Yu, X. Molecular cloning, characterization and expression analysis of HSP60, HSP70 and HSP90 in the golden apple snail, Pomaceacanaliculata. Fish Shellfish Immunol. 2014, 41, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, E.; Fajdiga, S.; Malago, J.; Koninkx, J.; Tooten, P.; Van Dijk, J. Inhibition of salmonella-induced IL-8 synthesis and expression of Hsp70 in enterocyte-like Caco-2 cells after exposure to non-starter lactobacilli. Int. J. Food Microbiol. 2006, 112, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef]
- Arvans, D.L.; Vavricka, S.R.; Ren, H.; Musch, M.W.; Kang, L.; Rocha, F.G.; Lucioni, A.; Turner, J.R.; Alverdy, J.; Chang, E.B. Luminal Bacterial Flora Determines Physiological Expression of Intestinal Epithelial Cytoprotective Heat Shock Proteins 25 and 72. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G696–G704. [Google Scholar] [CrossRef]
- Sarkar, G.; Mondal, S.; Bhattacharya, D.; Ponraj, P.; Sawhney, S.; Bala, P.; Chakraborty, D.; Sunder, J.; De, A.K. Effect of a multi-strain probiotic on growth performance, lipid panel, antioxidant profile, and immune response in andaman local piglets at weaning. Fermentation 2023, 9, 970. [Google Scholar] [CrossRef]
- Panteli, N.; Feidantsis, K.; Demertzioglou, M.; Paralika, V.; Karapanagiotis, S.; Mylonas, C.C.; Kormas, K.A.; Mente, E.; Makridis, P.; Antonopoulou, E. The Probiotic Phaeobacter inhibens Provokes Hypertrophic Growth via Activation of the IGF-1/Akt Pathway during the Process of Metamorphosis of Greater Amberjack (Seriola dumerili, Risso 1810). Animals 2023, 13, 2154. [Google Scholar] [CrossRef]
- Gracey, A.Y.; Cossins, A.R. Application of microarray technology in environmental and comparative physiology. Annu. Rev. Physiol. 2003, 65, 231–259. [Google Scholar] [CrossRef]
- Taniguchi, Y.; Choi, P.J.; Li, G.W.; Chen, H.; Babu, M.; Hearn, J.; Emili, A.; Xie, X.S. Quantifying E. coli proteome and transcriptome with single-molecule sensitivity in single cells. Science 2010, 329, 533–538. [Google Scholar] [CrossRef]
- Buckley, D.B.; Klaassen, C.D. Tissue-and gender-specific mRNA expression of UDP-glucuronosyltransferases (UGTs) in mice. Drug Metab. Dispos. 2007, 35, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Fournier, M.L.; Paulson, A.; Pavelka, N.; Mosley, A.L.; Gaudenz, K.; Bradford, W.D.; Glynn, E.; Li, H.; Sardiu, M.E.; Fieharty, B.; et al. Delayed correlation of mRNA and protein expression in rapamycin-treated cells and a role for Ggc1 in cellular sensitivity to rapamycin. Mol. Cell. Proteom. 2010, 9, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Vogel, C.; Marcotte, E.M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nature 2012, 13, 227–232. [Google Scholar] [CrossRef]
- Liu, J.C.; Ming, H.; Li, W.; Cheng, X.S. Heat shock protein 70 gene transfection protects rat myocardium cell against anoxiareoxygeneration injury. Chin. Med. J. 2007, 120, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Gan, F.; Ren, F.; Chen, X.; Lv, C.; Pan, C.; Ye, G.; Shi, J.; Shi, X.; Zhou, H.; Shituleni, A.S.; et al. Effects of selenium-enriched probiotics on heat shock protein mRNA levels in piglet under heat stress conditions. J. Agric. Food Chem. 2013, 61, 2385–2391. [Google Scholar] [CrossRef] [PubMed]
- Alvanou, M.V.; Feidantsis, K.; Staikou, A.; Apostolidis, A.P.; Michaelidis, B.; Giantsis, I.A. Probiotics, prebiotics, and synbiotics utilization in crayfish aquaculture and factors affecting gut microbiota. Microorganisms 2023, 11, 1232. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.Z.; Kumbhar, S.; Hamid, M.; Afzal, S.; Parveen, F.; Liu, Y.; Shu, H.; Mengistu, M.B.; Huang, K. Effects of selenium-enriched probiotics on heart lesions by influencing the mRNA expressions of selenoproteins and heat shock proteins in heat stressed broiler chickens. Pak. Vet. J. 2016, 36, 460–464. [Google Scholar]
- Salakou, S.; Kardamakis, D.; Tsamandas, A.C.; Zolota, V.; Apostolakis, E.; Tzelepi, V.; Papathanasopoulos, P.; Bonikos, D.S.; Papapetropoulos, T.; Petsas, T.; et al. Increased Bax/Bcl-2 ratio up-regulates caspase-3 and increases apoptosis in the thymus of patients with myasthenia gravis. In Vivo 2007, 21, 123–132. [Google Scholar] [PubMed]
- Wu, Y.; Wang, B.; Zeng, Z.; Liu, R.; Tang, L.; Gong, L.; Li, W. Effects of probiotics Lactobacillus plantarum 16 and Paenibacilluspolymyxa 10 on intestinal barrier function, antioxidative capacity, apoptosis, immune response, and biochemical parameters in broilers. Poult. Sci. 2019, 98, 5028–5039. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.F.; Lin, P.P.; Chen, C.H.; Yeh, Y.L.; Huang, C.C.; Huang, C.Y.; Tsai, C.C. Effects of lactic acid bacteria on cardiac apoptosis are mediated by activation of the phosphatidylinositol-3 kinase/AKT survival-signalling pathway in rats fed a high-fat diet. Int. J. Mol. Med. 2015, 35, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, S.; Chakraborty, T.; Prusty, A.K.; Paniprasad, K.; Mohanta, K.N. Beneficial effects of dietary probiotics mixture on hemato-immunology and cell apoptosis of Labeorohita fingerlings reared at higher water temperatures. PLoS ONE 2014, 9, e100929. [Google Scholar] [CrossRef]
- Yenari, M.A.; Liu, J.; Zheng, Z.; Vexler, Z.S.; Lee, J.E.; Giffard, R.G. Antiapoptotic and anti-inflammatory mechanisms of heat-shock protein protection. Ann. N. Y. Acad. Sci. 2005, 1053, 74–83. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Composition (in %) | |||
| Total nitrogenous substances | 20% | Calcium (Ca) | 1.00% |
| Total fats | 3.40% | Phosphorus (P) | 0.80% |
| Total fibrous substances | 3.30% | Sodium (Na) | 0.20% |
| Total ash | 5.70% | Lysine | 1.20% |
| Humidity | 13.00% | Methionine | 0.50% |
| Additives (per Kg of forage) | |||
| ΒΙΤ.A: Retinol (Ε672) | 12,000 mg | BIT.H: Biotin | 0.20 mg |
| ΒΙΤ.D3:Cholecalciferol (Ε671) | 3000 mg | Cholinechloride | 500 mg |
| ΒΙΤ.Ε: Tocopherol acetate (Ε307) | 50 mg | Co: Cobalt (CoCO3)(E3) | 0.25 mg |
| ΒΙΤ.Κ3: Menadione | 2.50 mg | I: Iodine [Ca(IO3)2](E2) | 1.50 mg |
| ΒΙΤ.Β1: Thiamine | 2 mg | Se: Selenium (Na2SeO3)(E8) | 0.20 mg |
| ΒΙΤ.Β2: Riboflavin (Ε101) | 4 mg | Fe: Iron (FeSO4)(E1) | 120 mg |
| ΒΙΤ.Β6: Pyridoxine(3α831) | 3 mg | Mn: Manganese(MnO)(E5) | 50 mg |
| ΒΙΤ.Β12: Cyanocobalamin | 0.02 mg | Cu: Copper (CuSO4·5H2O)(E4) | 20 mg |
| Niacin | 20 mg | Zn: Zinc (ZnO)(E6) | 120 mg |
| Pantothenic acid | 11 mg | Antioxidant: Ethoxycine(Ε324) | 2.50 mg |
| ΒΙΤ.C: Folic acid | 2 mg | ||
| Composition | |||
| Corn, soybean flour *, soybean oil, calcium carbonate, monocalcium phosphate, premix of vitamins and trace elements, sodium chloride (salt), sodium bicarbonate (soda). | |||
| Target Gene | Forward Primer (5′-3′) Reverse Primer (5′-3′) | Primer Efficiency |
|---|---|---|
| HSP70 | 5′-CAGCTTGAGGGCTACGTCTT-3′ 5′-GTTGCTGTGTTACTTTACCG-3′ | 1.96 |
| ND2 | 5′-TGCAGAGGGAGAATCTGAGT-3′ 5′-TGGATCGAAATTATCCCGACG-3′ | 1.98 |
| 30 Days | 60 Days | 90 Days | |
| Control | 0.26 | 0.33 | 0.3 |
| FC | 0.26 | 0.3 | 0.3 |
| 2FC | 0.3 | 0.33 | 0.36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Efstratiou, E.; Feidantsis, K.; Makri, V.; Staikou, A.; Giantsis, I.A. Evidence for Beneficial Physiological Responses of the Land Snail Cornu aspersum to Probiotics’ (Lactobacillus plantarum) Dietary Intervention. Animals 2024, 14, 857. https://0-doi-org.brum.beds.ac.uk/10.3390/ani14060857
Efstratiou E, Feidantsis K, Makri V, Staikou A, Giantsis IA. Evidence for Beneficial Physiological Responses of the Land Snail Cornu aspersum to Probiotics’ (Lactobacillus plantarum) Dietary Intervention. Animals. 2024; 14(6):857. https://0-doi-org.brum.beds.ac.uk/10.3390/ani14060857
Chicago/Turabian StyleEfstratiou, Efstratios, Konstantinos Feidantsis, Vasiliki Makri, Alexandra Staikou, and Ioannis A. Giantsis. 2024. "Evidence for Beneficial Physiological Responses of the Land Snail Cornu aspersum to Probiotics’ (Lactobacillus plantarum) Dietary Intervention" Animals 14, no. 6: 857. https://0-doi-org.brum.beds.ac.uk/10.3390/ani14060857