
Morphological and Transcriptomic Analyses Reveal the Toxicological Mechanism and Risk of Nitrate Exposure in Bufo gargarizans Embryos

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Solution
2.2. Animal Husbandry and Exposure Experiment
2.3. Embryonic Developmental Stage Determination and Morphologic Measurements
2.4. Transcriptomic Analysis
2.5. Statistical Analysis
3. Results
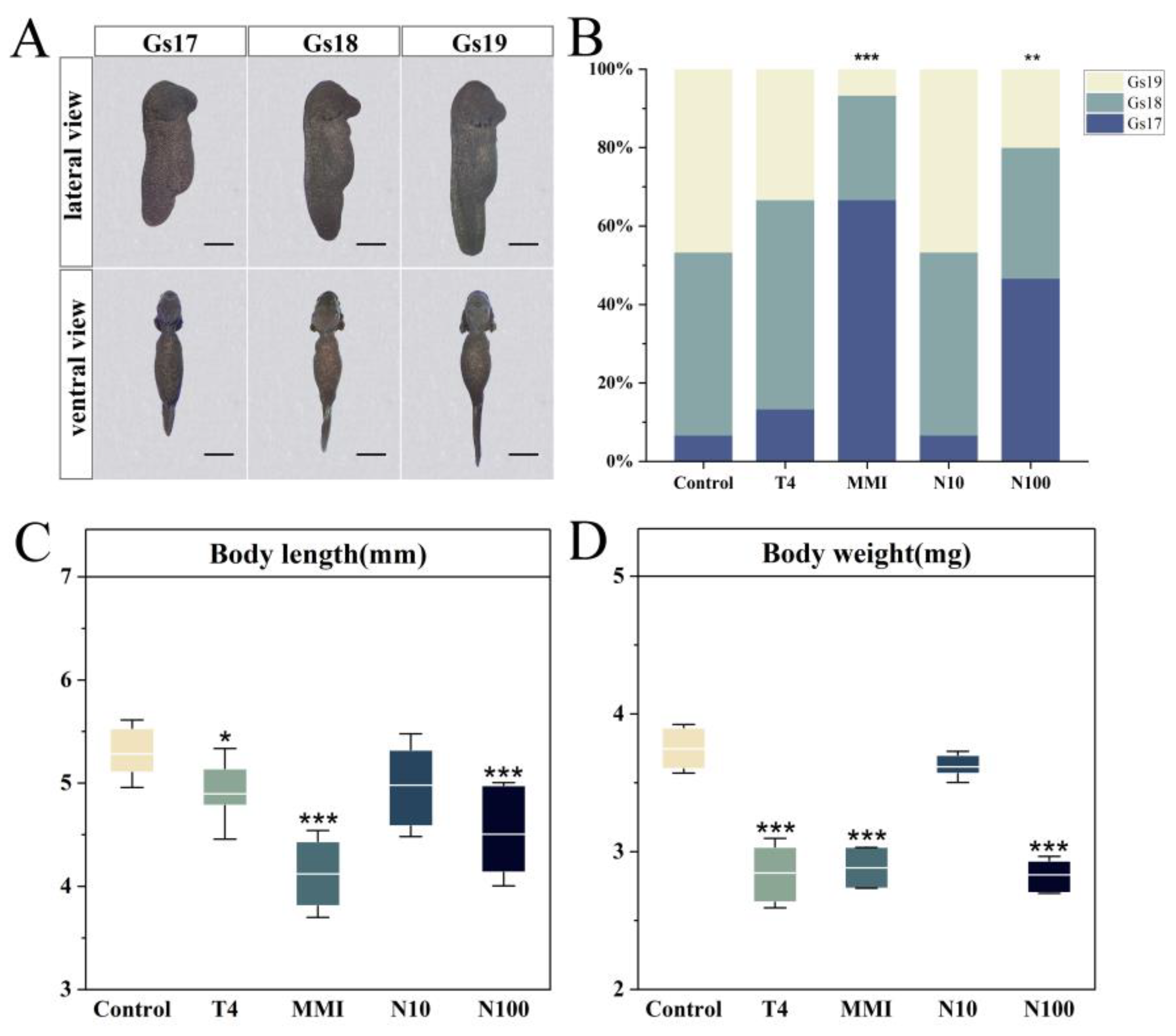
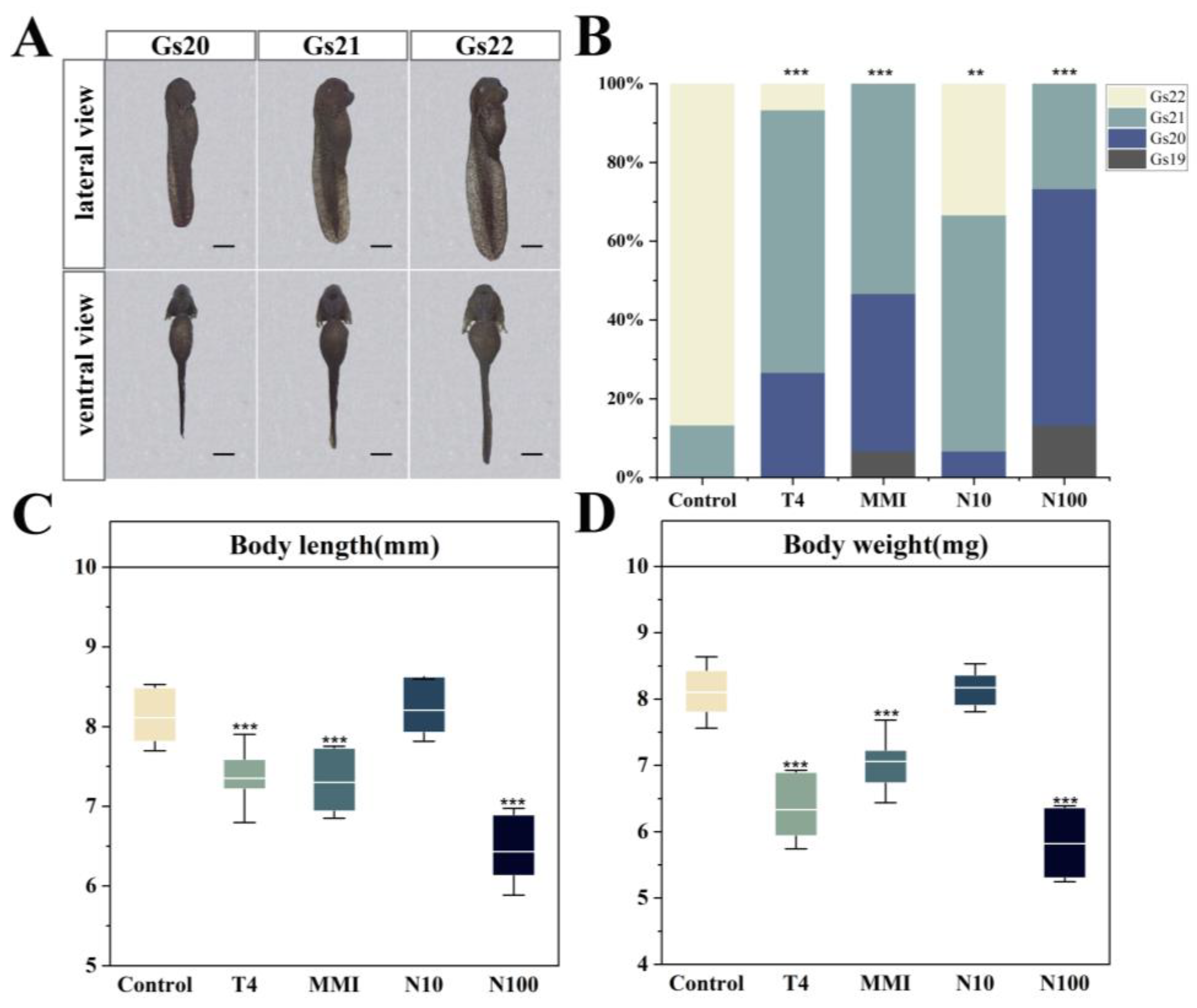
3.1. Embryonic Growth and Development
3.2. Identification of Differentially Expressed Genes (DEGs)
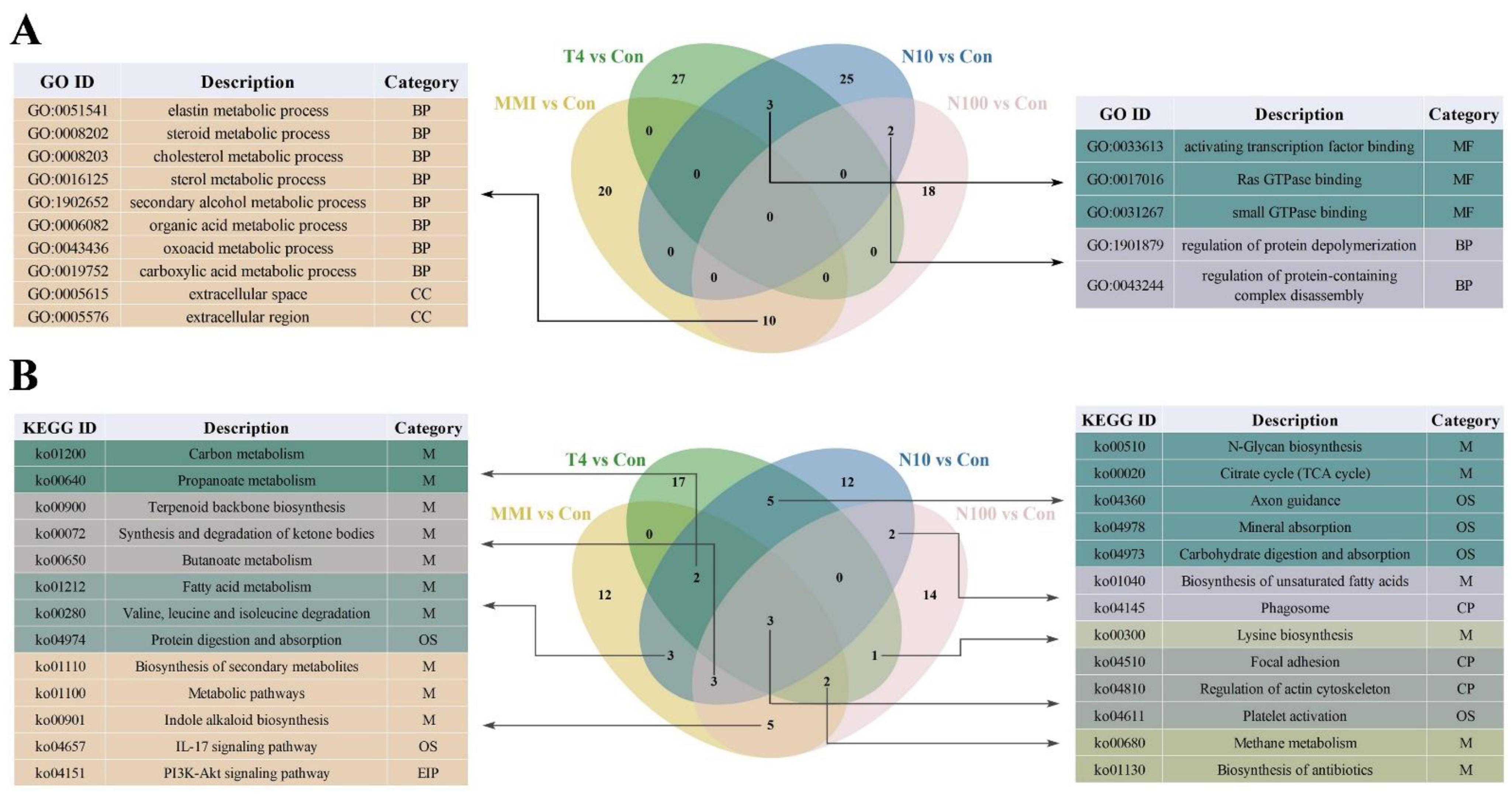
3.3. GO and KEGG Enrichment
3.4. Transcriptional Expression Profiles of the Genes Related to Enrichment Pathways
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kazakis, N.; Matiatos, I.; Ntona, M.-M.; Bannenberg, M.; Kalaitzidou, K.; Kaprara, E.; Mitrakas, M.; Ioannidou, A.; Vargemezis, G.; Voudouris, K. Origin, implications and management strategies for nitrate pollution in surface and ground waters of Anthemountas basin based on a δ15N-NO3− and δ18O-NO3− isotope approach. Sci. Total Environ. 2020, 724, 138211. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Wang, X.; Qian, S.; Liu, P.; Li, X.; Li, J. Exposure to nitrate induces alterations in blood parameter responses, liver immunity, and lipid metabolism in juvenile turbot (Scophthalmus maximus). Aquat. Toxicol. 2022, 251, 106280. [Google Scholar] [CrossRef]
- Ahmed, M.; Rauf, M.; Mukhtar, Z.; Saeed, N.A. Excessive use of nitrogenous fertilizers: An unawareness causing serious threats to environment and human health. Environ. Sci. Pollut. Res. Int. 2017, 24, 26983–26987. [Google Scholar] [CrossRef] [PubMed]
- Velusamy, K.; Periyasamy, S.; Kumar, P.S.; Vo, D.-V.N.; Sindhu, J.; Sneka, D.; Subhashini, B. Advanced techniques to remove phosphates and nitrates from waters: A review. Environ. Chem. Lett. 2021, 19, 3165–3180. [Google Scholar] [CrossRef]
- Cao, Y.; Tang, C.; Song, X.; Liu, C.; Zhang, Y. Characteristics of nitrate in major rivers and aquifers of the Sanjiang Plain, China. J. Environ. Monit. 2012, 14, 2624–2633. [Google Scholar] [CrossRef] [PubMed]
- Re, V.; Sacchi, E.; Kammoun, S.; Tringali, C.; Trabelsi, R.; Zouari, K.; Daniele, S. Integrated socio-hydrogeological approach to tackle nitrate contamination in groundwater resources. The case of Grombalia Basin (Tunisia). Sci. Total Environ. 2017, 593–594, 664–676. [Google Scholar] [CrossRef] [PubMed]
- Camargo, J.A.; Alonso, A.; Salamanca, A. Nitrate toxicity to aquatic animals: A review with new data for freshwater invertebrates. Chemosphere 2005, 58, 1255–1267. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Wang, Y.; Xiao, Y.; Li, X.; Zhou, L.; Wang, Y.; Du, T.; Ma, X.; Li, J. Investigating the effect of nitrate on juvenile turbot (Scophthalmus maximus) growth performance, health status, and endocrine function in marine recirculation aquaculture systems. Ecotoxicol. Environ. Saf. 2021, 208, 111617. [Google Scholar] [CrossRef]
- Edwards, T.M.; Guillette, L.J. Reproductive characteristics of male mosquitofish (Gambusia holbrooki) from nitrate-contaminated springs in Florida. Aquat. Toxicol. 2007, 85, 40–47. [Google Scholar] [CrossRef]
- Kellock, K.A.; Moore, A.P.; Bringolf, R.B. Chronic nitrate exposure alters reproductive physiology in fathead minnows. Environ. Pollut. 2018, 232, 322–328. [Google Scholar] [CrossRef]
- Schram, E.; Roques, J.A.C.; Abbink, W.; Yokohama, Y.; Spanings, T.; de Vries, P.; Bierman, S.; van de Vis, H.; Flik, G. The impact of elevated water nitrate concentration on physiology, growth and feed intake of African catfish Clarias gariepinus (Burchell 1822). Aquac. Res. 2014, 45, 1499–1511. [Google Scholar] [CrossRef]
- Li, J.; Ye, Z. The potential role and regulatory mechanisms of MUC5AC in chronic obstructive pulmonary disease. Molecules 2020, 25, 4437. [Google Scholar] [CrossRef] [PubMed]
- Ruthsatz, K.; Bartels, F.; Stützer, D.; Eterovick, P.C. Timing of parental breeding shapes sensitivity to nitrate pollution in the common frog Rana temporaria. J. Therm. Biol. 2022, 108, 103296. [Google Scholar] [CrossRef] [PubMed]
- Licht, L.E. Shedding light on ultraviolet radiation and amphibian embryos. BioScience 2003, 53, 551–561. [Google Scholar] [CrossRef]
- Pechenik, J.A. Larval experience and latent effects—Metamorphosis is not a new beginning. Integr. Comp. Biol. 2006, 46, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Urbina, J.; Bredeweg, E.M.; Blaustein, A.R.; Garcia, T.S. Direct and latent effects of pathogen exposure across native and invasive amphibian life stages. Front. Vet. Sci. 2021, 8, 732993. [Google Scholar] [CrossRef] [PubMed]
- Hinther, A.; Edwards, T.M.; Guillette, L.J., Jr.; Helbing, C.C. Influence of nitrate and nitrite on thyroid hormone responsive and stress-associated gene expression in cultured Rana catesbeiana tadpole tail fin tissue. Front. Genet. 2012, 3, 51. [Google Scholar] [CrossRef]
- Ortiz-Santaliestra, M.E.; Sparling, D.W. Alteration of larval development and metamorphosis by nitrate and perchlorate in southern leopard frogs (Rana sphenocephala). Arch. Environ. Contam. Toxicol. 2007, 53, 639–646. [Google Scholar] [CrossRef]
- Wang, M.; Chai, L.; Zhao, H.; Wu, M.; Wang, H. Effects of nitrate on metamorphosis, thyroid and iodothyronine deiodinases expression in Bufo gargarizans larvae. Chemosphere 2015, 139, 402–409. [Google Scholar] [CrossRef]
- Nieuwkoop, P.D.; Faber, J. Normal Table of Xenopus laevis (Daudin); Elsevier/North-Holland: New York, NY, USA, 1956. [Google Scholar]
- Gosner, K.L. A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica 1960, 16, 183–190. [Google Scholar]
- Honda, J.; Ogawa, K.; Taniguchi, K. Immunohistochemical and morphometric studies on the development of the thyroid, parathyroid and ultimobranchial body in Xenopus laevis Daudin. Exp. Anim. 1993, 42, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Morvan Dubois, G.; Sebillot, A.; Kuiper, G.G.; Verhoelst, C.H.; Darras, V.M.; Visser, T.J.; Demeneix, B.A. Deiodinase activity is present in Xenopus laevis during early embryogenesis. Endocrinology 2006, 147, 4941–4949. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Fong, J.J.; Jiang, J.-P.; Li, P.-P.; Waldman, B.; Chong, J.R.; Lee, H.; Min, M.-S. Phylogeographic study of the Bufo gargarizans species complex, with emphasis on Northeast Asia. Anim. Cells Syst. 2021, 25, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Li, C.; Wang, H.; Zhao, M.; Gu, Q.; Gu, Z.; Liao, C.; Wu, H. Mutual mate choice in the Asiatic toad, Bufo gargarizans, exerts stabilizing selection on body size. Chin. Sci. Bull. 2014, 59, 38–45. [Google Scholar] [CrossRef]
- Othman, S.N.; Litvinchuk, S.N.; Maslova, I.; Dahn, H.; Messenger, K.R.; Andersen, D.; Jowers, M.J.; Kojima, Y.; Skorinov, D.V.; Yasumiba, K.; et al. From Gondwana to the Yellow Sea, evolutionary diversifications of true toads Bufo sp. in the Eastern Palearctic and a revisit of species boundaries for Asian lineages. Elife 2022, 11, e70494. [Google Scholar] [CrossRef]
- Niu, Z.; Liu, Y.; Wang, Y.; Liu, Y.; Chai, L.; Wang, H. Impairment of bile acid metabolism and altered composition by lead and copper in Bufo gargarizans tadpoles. Sci. Total Environ. 2023, 900, 165901. [Google Scholar] [CrossRef] [PubMed]
- Crane, H.M.; Pickford, D.B.; Hutchinson, T.H.; Brown, J.A. The effects of methimazole on development of the fathead minnow, Pimephales promelas, from embryo to adult. Toxicol. Sci. 2006, 93, 278–285. [Google Scholar] [CrossRef]
- Fabrezi, M.; Cruz, J.C. Phenotypic variation through ontogeny: Thyroid axis disruption during larval development in the frog Pleurodema borellii. Front. Ecol. Evol. 2021, 9, 715988. [Google Scholar] [CrossRef]
- Brunelli, E.; Bernabò, I.; Berg, C.; Lundstedt-Enkel, K.; Bonacci, A.; Tripepi, S. Environmentally relevant concentrations of endosulfan impair development, metamorphosis and behaviour in Bufo bufo tadpoles. Aquat. Toxicol. 2009, 91, 135–142. [Google Scholar] [CrossRef]
- Eriyamremu, G.E.; Osagie, V.E.; Omoregie, S.E.; Omofoma, C.O. Alterations in glutathione reductase, superoxide dismutase, and lipid peroxidation of tadpoles (Xenopus laevis) exposed to Bonny Light crude oil and its fractions. Ecotoxicol. Environ. Saf. 2008, 71, 284–290. [Google Scholar] [CrossRef]
- Godfrey, A.; Hooser, B.; Abdelmoneim, A.; Horzmann, K.A.; Freemanc, J.L.; Sepúlveda, M.S. Thyroid disrupting effects of halogenated and next generation chemicals on the swim bladder development of zebrafish. Aquat. Toxicol. 2017, 193, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Santos, T.P.; da Silva Bastos, P.E.; da Silva, J.F.; de Medeiros Vieira, S.M.; da Silva, M.C.G.; de Andrade, A.L.C.; Padilha, R.M.O.; dos Santos Magnabosco, A.R.; Cadena, M.R.S.; Cadena, P.G. Single and joint toxic effects of thyroid hormone, levothyroxine, and amiodarone on embryo-larval stages of zebrafish (Danio rerio). Ecotoxicology 2023, 32, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Wu, B.; Xiong, X.; Wang, J. Short-term toxicity of ammonia, nitrite, and nitrate to early life stages of the rare minnow (Gobiocypris rarus). Environ. Toxicol. Chem. 2016, 35, 1422–1427. [Google Scholar] [CrossRef]
- Allran, J.; Karasov, W. Effects of atrazine and nitrate on northern leopard frog (Rana pipiens) larvae exposed in the laboratory from posthatch through metamorphosis. Environ. Toxicol. Chem. 2000, 19, 2850–2855. [Google Scholar] [CrossRef]
- Berven, K.A. Factors affecting population fluctuations in larval and adult stages of the wood frog (Rana sylvatica). Ecology 1990, 71, 1599–1608. [Google Scholar] [CrossRef]
- Smith, D.C. Adult recruitment in chorus frogs: Effects of size and date at metamorphosis. Ecology 1987, 68, 344–350. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, H.; Chai, L.; Li, X.; Wu, M.; Wang, H. Effects of perchlorate and exogenous T4 exposures on development, metamorphosis and endochondral ossification in Bufo gargarizans larvae. Aquat. Toxicol. 2022, 242, 106036. [Google Scholar] [CrossRef] [PubMed]
- Rowe, C.L.; Kinney, O.M.; Nagle, R.D.; Congdon, J.D. Elevated maintenance costs in an anuran (Rana catesbeiana) exposed to a mixture of trace elements during the embryonic and early larval periods. Physiol. Zool. 1998, 71, 27–35. [Google Scholar] [CrossRef]
- Fernández, L.P.; López-Márquez, A.; Santisteban, P. Thyroid transcription factors in development, differentiation and disease. Nat. Rev. Endocrinol. 2015, 11, 29–42. [Google Scholar] [CrossRef]
- Gandarilla-Esparza, D.D.; Calleros-Rincón, E.Y.; Macias, H.M.; González-Delgado, M.F.; Vargas, G.G.; Sustaita, J.D.; González-Zamora, A.; Ríos-Sánchez, E.; Pérez-Morales, R. FOXE1 polymorphisms and chronic exposure to nitrates in drinking water cause metabolic dysfunction, thyroid abnormalities, and genotoxic damage in women. Genet. Mol. Biol. 2021, 44, e20210020. [Google Scholar] [CrossRef]
- De Felice, M.; Di Lauro, R. Thyroid development and its disorders: Genetics and molecular mechanisms. Endocr. Rev. 2004, 25, 722–746. [Google Scholar] [CrossRef]
- Cerqueira, N.M.F.S.A.; Oliveira, E.F.; Gesto, D.S.; Santos-Martins, D.; Moreira, C.; Moorthy, H.N.; Ramos, M.J.; Fernandes, P.A. Cholesterol Biosynthesis: A Mechanistic Overview. Biochemistry 2016, 55, 5483–5506. [Google Scholar] [CrossRef]
- Kovacs, W.J.; Olivier, L.M.; Krisans, S.K. Central role of peroxisomes in isoprenoid biosynthesis. Prog. Lipid Res. 2002, 41, 369–391. [Google Scholar] [CrossRef]
- Olżyńska, A.; Kulig, W.; Mikkolainen, H.; Czerniak, T.; Jurkiewicz, P.; Cwiklik, L.; Rog, T.; Hof, M.; Jungwirth, P.; Vattulainen, I. Tail-oxidized cholesterol enhances membrane permeability for small solutes. Langmuir 2020, 36, 10438–10447. [Google Scholar] [CrossRef]
- Simons, K.; Ikonen, E. How cells handle cholesterol. Science 2000, 290, 1721–1726. [Google Scholar] [CrossRef]
- Fernández-Pérez, E.J.; Sepúlveda, F.J.; Peters, C.; Bascuñán, D.; Riffo-Lepe, N.O.; González-Sanmiguel, J.; Sánchez, S.A.; Peoples, R.W.; Vicente, B.; Aguayo, L.G. Effect of cholesterol on membrane fluidity and association of Aβ oligomers and subsequent neuronal damage: A double-edged sword. Front. Aging Neurosci. 2018, 10, 226. [Google Scholar] [CrossRef]
- Lee, A.G. How lipids affect the activities of integral membrane proteins. BBA—Biomembr. 2004, 1666, 62–87. [Google Scholar] [CrossRef]
- Paukner, K.; Králová Lesná, I.; Poledne, R. Cholesterol in the cell membrane-An emerging player in atherogenesis. Int. J. Mol. Sci. 2022, 23, 533. [Google Scholar] [CrossRef]
- Ammendolia, D.A.; Bement, W.M.; Brumell, J.H. Plasma membrane integrity: Implications for health and disease. BMC Biol. 2021, 19, 71. [Google Scholar] [CrossRef]
- Papalazarou, V.; Maddocks, O.D.K. Supply and demand: Cellular nutrient uptake and exchange in cancer. Mol. Cell 2021, 81, 3731–3748. [Google Scholar] [CrossRef]
- Sunshine, H.; Iruela-Arispe, M.L. Membrane lipids and cell signaling. Curr. Opin. Lipidol. 2017, 28, 408–413. [Google Scholar] [CrossRef]
- Rone, M.B.; Fan, J.; Papadopoulos, V. Cholesterol transport in steroid biosynthesis: Role of protein–protein interactions and implications in disease states. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2009, 1791, 646–658. [Google Scholar] [CrossRef]
- Bablok, M.; Gellisch, M.; Scharf, M.; Brand-Saberi, B.; Morosan-Puopolo, G. Spatiotemporal expression pattern of the chicken glucocorticoid receptor during early embryonic development. Ann. Anat. 2023, 247, 152056. [Google Scholar] [CrossRef]
- Fowden, A.L.; Forhead, A.J. Glucocorticoids as regulatory signals during intrauterine development. Exp. Physiol. 2015, 100, 1477–1487. [Google Scholar] [CrossRef]
- Kuo, T.; McQueen, A.; Chen, T.-C.; Wang, J.-C. Regulation of glucose homeostasis by glucocorticoids. In Glucocorticoid Signaling: From Molecules to Mice to Man; Wang, J.-C., Harris, C., Eds.; Springer New York: New York, NY, USA, 2015; pp. 99–126. [Google Scholar]
- Vegiopoulos, A.; Herzig, S. Glucocorticoids, metabolism and metabolic diseases. Mol. Cell. Endocrinol. 2007, 275, 43–61. [Google Scholar] [CrossRef]
- Seckl, J.R.; Holmes, M.C. Mechanisms of Disease: Glucocorticoids, their placental metabolism and fetal ‘programming’ of adult pathophysiology. Nat. Clin. Pract. Endocrinol. Metab. 2007, 3, 479–488. [Google Scholar] [CrossRef]
- Stubbe, J.; Madsen, K.; Nielsen, F.T.; Skøtt, O.; Jensen, B.L. Glucocorticoid impairs growth of kidney outer medulla and accelerates loop of Henle differentiation and urinary concentrating capacity in rat kidney development. Am. J. Physiol.-Ren. Physiol. 2006, 291, F812–F822. [Google Scholar] [CrossRef]
- Sarapultsev, A.; Sarapultsev, P.; Dremencov, E.; Komelkova, M.; Tseilikman, O.; Tseilikman, V. Low glucocorticoids in stress-related disorders: The role of inflammation. Stress 2020, 23, 651–661. [Google Scholar] [CrossRef]
- NandyMazumdar, M. Chapter 2—Airway mucus, infection, and therapeutic strategies. In Viral, Parasitic, Bacterial, and Fungal Infections; Bagchi, D., Das, A., Downs, B.W., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 19–28. [Google Scholar]
- Takami, S.; Mizuno, T.; Oyanagi, T.; Tadaki, H.; Suzuki, T.; Muramatsu, K.; Takizawa, T.; Arakawa, H. Glucocorticoids inhibit MUC5AC production induced by transforming growth factor-α in human respiratory cells. Allergol. Int. 2012, 61, 451–459. [Google Scholar] [CrossRef]
- Evans, C.M.; Kim, K.; Tuvim, M.J.; Dickey, B.F. Mucus hypersecretion in asthma: Causes and effects. Curr. Opin. Pulm. Med. 2009, 15, 4–11. [Google Scholar] [CrossRef]
- Cornick, S.; Tawiah, A.; Chadee, K. Roles and regulation of the mucus barrier in the gut. Tissue Barriers 2015, 3, e982426. [Google Scholar] [CrossRef] [PubMed]
- Fahy, J.V.; Dickey, B.F. Airway mucus function and dysfunction. N. Engl. J. Med. 2010, 363, 2233–2247. [Google Scholar] [CrossRef] [PubMed]
- Rogers, D. Airway mucus hypersecretion in asthma and COPD. In Asthma and COPD: Basic Mechanisms and Clinical Management; Elsevier/Academic Press: Amsterdam, The Netherlands, 2009; pp. 211–223. [Google Scholar]
- Liang, S.C.; Nickerson-Nutter, C.; Pittman, D.D.; Carrier, Y.; Goodwin, D.G.; Shields, K.M.; Lambert, A.J.; Schelling, S.H.; Medley, Q.G.; Ma, H.L.; et al. IL-22 induces an acute-phase response. J. Immunol. 2010, 185, 5531–5538. [Google Scholar] [CrossRef] [PubMed]
- Sabat, R.; Ouyang, W.; Wolk, K. Therapeutic opportunities of the IL-22–IL-22R1 system. Nat. Rev. Drug Discov. 2014, 13, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Aujla, S.J.; Chan, Y.R.; Zheng, M.; Fei, M.; Askew, D.J.; Pociask, D.A.; Reinhart, T.A.; McAllister, F.; Edeal, J.; Gaus, K.; et al. IL-22 mediates mucosal host defense against Gram-negative bacterial pneumonia. Nat. Med. 2008, 14, 275–281. [Google Scholar] [CrossRef]
- Yan, J.; Yu, J.; Liu, K.; Liu, Y.; Mao, C.; Gao, W. The pathogenic roles of IL-22 in colitis: Its transcription regulation by musculin in T helper subsets and innate lymphoid cells. Front. Immunol. 2021, 12, 758730. [Google Scholar] [CrossRef] [PubMed]
- Cinelli, M.A.; Do, H.T.; Miley, G.P.; Silverman, R.B. Inducible nitric oxide synthase: Regulation, structure, and inhibition. Med. Res. Rev. 2020, 40, 158–189. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Hierholzer, C.; Harbrecht, B.; Menezes, J.M.; Kane, J.; MacMicking, J.; Nathan, C.F.; Peitzman, A.B.; Billiar, T.R.; Tweardy, D.J. Essential role of induced nitric oxide in the initiation of the inflammatory response after hemorrhagic shock. J. Exp. Med. 1998, 187, 917–928. [Google Scholar] [CrossRef]
- Greer, J. Pathophysiology of cardiovascular dysfunction in sepsis. BJA Educ. 2015, 15, 316–321. [Google Scholar] [CrossRef]
- Sato, K.; Ozaki, K.; Oh, I.; Meguro, A.; Hatanaka, K.; Nagai, T.; Muroi, K.; Ozawa, K. Nitric oxide plays a critical role in suppression of T-cell proliferation by mesenchymal stem cells. Blood 2007, 109, 228–234. [Google Scholar] [CrossRef]
- Zhang, J.; Ney, P.A. Role of BNIP3 and NIX in cell death, autophagy, and mitophagy. Cell Death Differ. 2009, 16, 939–946. [Google Scholar] [CrossRef]
- Gao, A.; Jiang, J.; Xie, F.; Chen, L. Bnip3 in mitophagy: Novel insights and potential therapeutic target for diseases of secondary mitochondrial dysfunction. Clin. Chim. Acta 2020, 506, 72–83. [Google Scholar] [CrossRef]
- Regula, K.M.; Ens, K.; Kirshenbaum, L.A. Inducible expression of BNIP3 provokes mitochondrial defects and hypoxia-mediated cell death of ventricular myocytes. Circ. Res. 2002, 91, 226–231. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Kim, Y.-J.; Lee, S.; Park, J.-H. BNip3 is a mediator of TNF-induced necrotic cell death. Apoptosis 2011, 16, 114–126. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, L.; Niu, Z.; Xiao, S.; Wang, H.; Zhang, Y. Morphological and Transcriptomic Analyses Reveal the Toxicological Mechanism and Risk of Nitrate Exposure in Bufo gargarizans Embryos. Animals 2024, 14, 961. https://0-doi-org.brum.beds.ac.uk/10.3390/ani14060961
Xie L, Niu Z, Xiao S, Wang H, Zhang Y. Morphological and Transcriptomic Analyses Reveal the Toxicological Mechanism and Risk of Nitrate Exposure in Bufo gargarizans Embryos. Animals. 2024; 14(6):961. https://0-doi-org.brum.beds.ac.uk/10.3390/ani14060961
Chicago/Turabian StyleXie, Lei, Ziyi Niu, Shimin Xiao, Hongyuan Wang, and Yongpu Zhang. 2024. "Morphological and Transcriptomic Analyses Reveal the Toxicological Mechanism and Risk of Nitrate Exposure in Bufo gargarizans Embryos" Animals 14, no. 6: 961. https://0-doi-org.brum.beds.ac.uk/10.3390/ani14060961