


Pathogenic Bacteria in Free-Living Birds, and Its Public Health Significance

Department of Veterinary Public Health, Faculty of Veterinary Medicine, University of Warmia and Mazury in Olsztyn, Oczapowskiego 14, 10-710 Olsztyn, Poland
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Animals 2024, 14(6), 968; https://0-doi-org.brum.beds.ac.uk/10.3390/ani14060968
Submission received: 10 February 2024
/
Revised: 7 March 2024
/
Accepted: 16 March 2024
/
Published: 20 March 2024
(This article belongs to the Special Issue Diagnosis, Evaluation, and Management of Wildlife, Exotic and Zoo Animals Disease)
Abstract
:Simple Summary
The avian population, occupying a diverse range of ecological niches and urban environments, serves as a crucial reservoir and sentinel for various pathogens. The role of birds in the transmission of infectious diseases is well established, and their ability to harbor a wide range of pathogens has been documented by numerous studies. As such, their surveillance and monitoring can provide valuable insights into the emergence and spread of bacterial diseases. Therefore, understanding the complex interactions between birds and their pathogens is of utmost importance in the field of public health and requires continued research and investigation.
Abstract
Birds that roam freely, particularly those that migrate, have the potential to carry a range of diseases that can be passed on to humans. The vast movement of these birds across diverse environments and urban areas can contribute to the spread of bacteria over long distances, impacting both human and animal populations. Stress, overcrowding, and human interaction can also play a role in the transmission of infectious diseases among birds and humans. Therefore, it is crucial to comprehend the intricate connections between birds, vectors, zoonotic pathogens, and the environment, especially given the increasing urbanization and emergence of zoonotic illnesses. This review aims to provide a systematic overview of the significance of avian species in transmitting bacterial pathogens that pose a risk to public health.
{kind=link}
{kind=link}
1. Introduction
Free-living birds are perceived as important sources of viruses, bacteria, and fungi that can significantly impact other animal populations and human health. Given their large movement capacities, they might transfer numerous pathogens during migrations, which could result in intra- and interspecies transmission [1]. This creates the potential for the establishment of novel foci of emerging or re-emerging communicable diseases along bird migration routes [2]. Birds, in particular free-living birds, have been widely reported as reservoirs of pathogens and, due to their ability to occupy different ecological niches and to adapt to many urban, suburban, and livestock environments, represent true sentinels [3]. Numerous studies conducted worldwide have reported the presence of various bacteria in wild birds, many of which may have a high zoonotic potential [4,5,6]. Long-distance migrants, such as barn swallows, fly from Europe to Africa for wintering, covering a distance of over 10,000 km every year [7,8]. Researchers discovered that migrant swallows were considerably more likely than resident swallows to have Salmonella spp. in their microbiome [9], which might play a role in the national and international dissemination of this bacteria. Some factors may significantly determine the role of wild migratory birds in the spread of certain pathogens.
The impact of stress levels and crowding on wild birds can lead to the transmission of infectious diseases among them, resulting in a rapid spread of pathogens throughout the population [2]. In light of their high population densities, close social interactions, spatial mobility, and capacity to colonize anthropogenic environments, birds possess significant potential to serve as reservoirs for pathogens that can be transmitted to other vertebrates, including humans [10]. Simultaneously, when people enter the habitats of wild birds, including to feed them, they become exposed to pathogens. It is important to note that birds, due to their mobility and the deposition of droppings, can act as vectors for the environmental circulation of zoonotic agents. Additionally, they may also spread antimicrobial-resistant bacteria with various resistance gene profiles, making it crucial to monitor their role in the spread of infectious diseases [11].
When raising the subject of wildlife fowl, the microbiological risks from the production and consumption of game birds cannot be omitted. They can carry common foodborne bacteria (e.g., Salmonella, Campylobacter, Yersinia, and Listeria) in their intestines [12,13]. Because game birds are generally asymptomatic carriers of pathogenic bacteria, these factors can easily be disseminated to hunters and consumers unnoticed through the handling and consumption of infected game birds [13]. However, the microbial contamination of game bird carcasses may be affected by factors like shot location, slaughter hygiene, and the cold chain.
Understanding the connections between birds, vectors, zoonotic pathogens, and the environment is crucial, given rising urbanization and the rise of zoonotic illnesses [14], as they comprise over 75% of all emerging human illnesses [15]. The objective of the present review is to evaluate the possible transmission of pathogenic bacteria from wild birds to the surrounding environment. These bacteria can then be transmitted to humans, highlighting the interconnection between environmental, animal, and human health. It is worth noting that most birds infected with zoonotic bacteria do not display clinical symptoms and are asymptomatic carriers. However, they pose a significant risk to human health, making it crucial to understand the potential dangers.
2. Materials and Methods
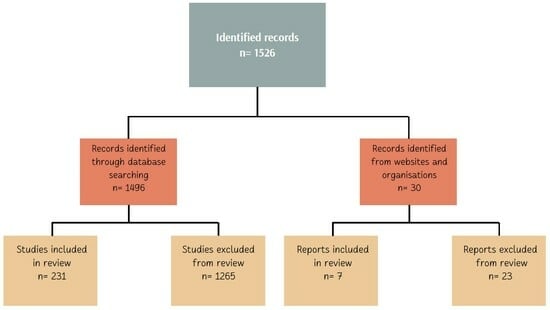
A systematic literature review was undertaken to elucidate the role of free-living birds as reservoirs and vectors of pathogenic bacteria, adhering to the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines (Figure 1).
The search strategy encompassed electronic databases, including PubMed and ScienceDirect, to identify pertinent articles published from September 2022 to January 2024. Search terms were meticulously chosen to cover a broad spectrum of relevant topics, including variations of “free-living birds”, “wild birds”, “migratory birds”, “avian”, “pathogenic bacteria”, “bacterial pathogens”, “resistance”, “virulence genes”, and related terminologies.
A total of 238 articles were meticulously selected for inclusion in this review, based on their relevance and contribution to understanding the role of free-living birds in the dissemination of pathogenic bacteria. Systematic data extraction was conducted, focusing on key elements such as bacterial species identified, host species involved, geographical locations, transmission routes, and public health implications.
Selected articles underwent critical evaluation for methodological rigor. Parameters assessed included study design, sample size, sampling methods, laboratory techniques utilized for bacterial identification and characterization, and statistical analyses employed to ensure robustness in the findings.
Any limitations or biases identified within the included studies were meticulously noted and discussed within the context of this review, thereby providing a systematic analysis of the synthesized evidence.
3. Bacterial Pathogens in Wild Birds
Campylobacteriosis is the most frequent zoonotic disease reported worldwide [16]. Campylobacter spp. can colonize the intestinal tract of most mammals and birds, both domestic and free-living. Although these bacteria are able to colonize the intestines of both domestic and free-living birds, are generally perceived as non-pathogenic. This assumption has been confirmed by different histopathological investigations, which showed no pathological lesions or marked alteration in crypt features in birds in a commensal state [17]. Usually, transmission of Campylobacter from birds to humans is due to consumption of poultry products, as poultry carcasses undergo cross-contamination during post-slaughter processing, mainly during defeathering and evisceration. Meta-analysis performed by Ansarifar et al. [18] reveals that the quail meat has the highest contamination (68.4%) with Campylobacter spp. followed by chicken (56.1%), turkey (27.4%) and ostrich meat (11.7%). The frequency of Campylobacter in wild birds may be affected by several variables, including geographic location, season, the health of the bird, the species of bird, the type of sample taken, the methodology, and ecological factors [19]. There have been reports of Campylobacter spp. in a variety of wildlife species, including squab [20], quail [21], guinea fowl [22], and duck [23,24,25]. Generally, Campylobacter isolates recovered from wild birds are described as genetically divergent; however, despite the high genetic divergence among environmental Campylobacter isolates, overlapping of sequence types of isolates derived from human outbreaks and wild birds is still observed [26,27]. An investigation performed in 2008 including the analysis of Campylobacter jejuni isolates from human, raw pea, and wild bird faecal samples confirmed the epidemiologic link between illness and the consumption of raw peas contaminated by sandhill cranes [27]. In a study performed by Abdollahpour et al. [28], wild bird faeces were identified as a source of Campylobacter jejuni infection in children’s playgrounds in Iran. Children’s behaviour, such as frequent hand–mouth contact, contributed to the ingestion of birds’ faeces that carried pathogenic bacteria. On the other hand, studies performed by Southern et al. [29] and Stuart et al. [30] showed the correlation between Campylobacter infection in humans and drinking pasteurized milk from bottles with damaged tops due to attacks by birds.
The wide distribution of Campylobacter spp. among wild birds also poses a risk to the farm environment. In the study conducted by Sanad et al. [31] in the USA, genotypically identical isolates of Campylobacter jejuni were recovered from the faeces of starlings and cattle on the same establishment. These bacteria were detected in 36.6% of dairy and 50.4% of starling faecal samples, and several genotypes overlapped between dairy cattle and starling isolates. These findings point to starlings as possible C. jejuni reservoirs and their potential impact on the epidemiology of clinically significant C. jejuni in the dairy community. The common distribution of Campylobacter among free-living birds poses a specific risk in the case of species that can reach the consumer’s table as a by-product of the shooting industry. The studies conducted by Kovanen et al. [32] in Finland showed that game birds may pose a risk for acquiring campylobacteriosis because they had C. jejuni genotypes highly similar to human isolates detected previously. Therefore, hygienic measures during slaughter and meat handling warrant special attention.
Listeria monocytogenes is the causative agent of human listeriosis, a severe illness that is particularly fatal for the elderly, pregnant women, and newborns [33]. It is a dangerous disease with a considerable fatality rate of 20–30% worldwide [34]. Listeriosis is a serious infection usually caused by ingestion of food contaminated with L. monocytogenes; however, birds may disseminate L. monocytogenes in nature and may also contaminate foods when entering food processing environments and outdoor marketplaces [35]. These bacteria are common in natural habitats like soil, surface water, sewage, or mammals’ and birds’ faeces [36,37]. However, many species of birds, including chickens, turkeys, pigeons, ducks, geese, pheasants, canaries, and cockatiels, are susceptible to natural infection, and clinical disease in birds is rare [38]. Birds can be infected via the airborne route by inhalation, uptake, or debeaking, and young birds appear to be more susceptible to Listeria infection than older birds, with listeriosis most commonly manifested as septicaemia [39]. The prevalence of this pathogen in wild birds varies from 3.4% in pigeons to 46% in urban rooks [35,40,41,42,43]. Hellstrom et al. [35] discovered that the incidence of L. monocytogenes in faeces is higher in birds at landfill sites than in metropolitan areas, and isolated strains are frequently similar to those detected in foods and food processing environments [35]. Widespread contamination of the environment with birds’ faeces can become a source of food- and feed-borne illnesses in humans and animals, with the chain of food production reflecting a continuum between the farm environment and human populations predisposed to listeriosis [44]. In 2013, an outbreak of listeriosis in a small pheasant breeder farm was described. Approximately 300 young birds and a few adult birds were found dead within a few days of the onset of clinical signs, which included diarrhoea and significant neurological symptoms. L. monocytogenes was observed in heart blood smears, liver, and brain impression smears [38]. This information is very alarming as the common pheasant is the most hunted, widespread, and economically important non-migratory game bird in Europe [45].
Clostridium perfringens is one of the major causative agents of enteric diseases both in humans and animals [46]. The main sources of this bacterium are healthy birds’ digestive contents, soil, dust, poultry litter, and animal faeces [47]. C. perfringens has been isolated from many bird species, such as ducks [48,49], wild crows [50], western bluebirds [51], ostriches [52], greater sage-grouses [53], pheasants [54], and capercaillies [55]. Moreover, once infected, the birds can transmit pathogens by dropping contaminated feces, which becomes a significant risk factor for illness among individuals and domestic animals. The study conducted by Craven et al. [56] revealed the prevalence of this enteropathogen in wild birds’ faeces near broiler chicken houses, which suggests that wild birds that gain entry to poultry grow-out houses have the potential to transmit these pathogens to poultry.
Necrotic enteritis (NE) caused by C. perfingens results in huge economic harm to chicken farms [57], as these bacteria that cause clinical or subclinical necrotic enteritis have the ability to synthesize toxins, bacteriocins, and enzymes of different natures, which modify the anatomical structure of the intestinal mucosa, enterocytes, and the cellular matrix, altering the physiological activities of the gastrointestinal tract, resulting in gastrointestinal disorders and diarrhea [47]. Seki et al. [58] examined the isolation and toxinotyping of C. perfringens from wild crows, starlings, and sparrows to clarify the relationships of these resident birds with the incursion of this bacteria to poultry farms. Among the resident birds examined, C. perfringens was isolated from 51.3% of large-billed crows, 43.4% of carrion crows, and 28.0% of white-cheeked starlings. As a result, it was suspected that among wild bird species subjected to testing, crows were likely to have a relationship with the incursion of C. perfringens to poultry farms.
Another significant multi-host pathogen is Staphylococcus aureus, a commensal organism that is able to effectively colonize a wide variety of host species, including many mammalian species and also birds [59]. This pathogen is part of the normal flora of the skin and other mucous membranes of poultry; however, in birds with reduced immunity, after crossing physical barriers, it gains access to tissues and the bloodstream and may cause local or systemic infection of poultry. S. aureus may cause septicaemia, fibrinous arthritis and tenosynovitis, chondronecrosis, and osteomyelitis in chickens, turkeys, ducks, and geese [60]. Staphylococcal food-borne disease (SFD) is one of the most common foodborne diseases worldwide, resulting from the contamination of meat and meat products, even though, when it comes to poultry consumption, Staphylococcal food-borne disease has mostly been linked to the ingestion of chicken meat [61,62,63,64]. Wide distribution of S. aureus among wild birds also was observed, this pathogen was isolated from birds of prey (33% on average) [65,66,67], lesser yellowlegs (0.87%) [68], geese (7.1%) [69], magpies, cinereous vultures (8.3%) [70], white storks (34.8%) [71], and white ibis (10.9%) [72]. As Sousa et al. [65] underlined, birds seem to be a natural reservoir of S. aureus and coagulase-negative staphylococci resistant to several antibiotics. Interaction between wildlife and people is becoming more frequent as habitats converge, implying an increasing chance of exchange of these microbes in the various ecosystems. Raptors such as owls that feed mostly on small mammals and insects and are in direct contact with many species of wild rodents operate as a significant vector for the transmission of pathogens between humans and animals [66]. Some researchers suggest that wild birds’ droppings may also contribute to the contamination of bathing water with S. aureus [73]. According to Cragg and Clayton [74], S. aureus is one of the most prevalent bacteria in seagulls’ faecal flora; hence, beaches with more seagulls may be considered as at a higher risk of S. aureus contamination. Generally, Staphylococci have been identified as an appropriate model for “One Health” investigations, as some species and clones have been demonstrated to be bound through the three ecosystems of interest (humans, animals, and the environment) [75].
Escherichia coli is a normal commensal in the guts of birds and animals; however, some wild birds may carry pathogenic strains of E. coli. Likewise, enteropathogenic E. coli (EPEC) and Shiga-toxin-producing E. coli (STEC) [76] belong to the category of diarrheagenic E. coli that can cause serious disease in humans [77]. The EPEC pathotype leads to a high child mortality rate in developing countries, and diarrhea caused by these strains is a consequence of loss of intestinal microvillus, while STEC infection causes haemorrhagic colitis following the injury of the intestinal epithelium [78]. In Italy, STEC was isolated from wild ducks and live common quail faecal matter, which indicates the potential role of migratory ducks and quails in the transmission of STEC [79]. A study performed in Brazil by Sanches et al. [80] proved that captive wild birds are reservoirs of enteropathogenic E. coli and Shiga-toxin-producing E. coli strains. In that study, faecal samples were analysed from 516 birds in two municipal zoos located in São Paulo State, Brazil. A total of 401 isolates were identified as E. coli. The pathotype classification of the isolates showed that typical EPEC was detected in 2.99%, atypical EPEC in 1.99%, and STEC in 0.74% of samples. Even though the frequency of these pathotypes was low and restricted to a few orders, the data suggest the potential public health risk that these birds represent as reservoirs of diarrheagenic E. coli. Borges et al. [77] isolated two STEC (0.8%) and five EPEC strains (2.0%) from wild birds, and four EPEC strains (2.0%) were from pigeons. The incidence of EPEC and STEC in wild birds was comparable to that reported in previous investigations of these free-living avian species, where rates varied from 0% to 1.8% for STEC strains [81,82,83,84,85] and from 2.8% to 4.9% for EPEC strains [83,86,87,88]. The authors underline that wild birds could act as carriers of STEC and EPEC and, therefore, may constitute a considerable hazard to human and animal health by transmission of these strains to the environment.
The intestines of wild birds are very often colonized by Salmonella spp., mostly with no signs of infection, which therefore can be shed in faeces. These bacteria have been found among various species, such as passerines (8.8%) [89,90,91,92], teals (3.3%) [13], pigeons (27.5%) [93,94], and guinea fowls (8.9%) [95]. According to Tizard [96], salmonellosis in wild birds occurs most commonly in those on a carnivorous or omnivorous diet, those that feed on the ground or on the food subject to faecal contamination, or those that live or feed in contaminated water. Although several serovars of Salmonella may be found in the intestines of wild birds, and they persist within bird populations through several mechanisms, the wild birds may be temporary or permanent Salmonella carriers or even suffer from clinical salmonellosis [96]. Several studies have confirmed mass die-offs of free-ranging birds of various taxa due to infection with Salmonella spp. [97], and it has been estimated that Salmonella spp. was responsible for 21.5% of passerine cases and 5.4% of total bird mortality in the United States in the years 1985–2004. The well-documented prevalence of Salmonella among free-living birds promotes the spread of this pathogen in the environment and poses a real threat to human health and life. The Centre for Disease Control and Prevention (CDC) confirmed an outbreak of salmonellosis in humans linked to songbirds, as they reported a total of 29 people infected, aged from 21 days to 89 years. Illnesses started on dates ranging from 26 December 2020 to 29 April 2021, in 12 states. Of the 21 people interviewed, 13 (62%) reported owning a bird feeder and 4 (19%) people reported contact with a sick or dead wild bird, while 15 people (71.4%) had pets that had access to or contact with wild birds [98]. Moreover, Lawson et al. [99] emphasize that garden birds are a source of human salmonellosis and highlight how important personal hygiene measures are when feeding wild birds. The majority of documented outbreaks of passerine salmonellosis happened at or around feeding stations, which are probably places where people might come into contact with ill or deceased garden birds and their droppings [99]. Additionally, Craven et al. [56] suggest that wild birds that gain entry to poultry grow-out houses have the potential to transmit these pathogens to poultry. On the other hand, wild birds can also pollute food that will be directly consumed by humans.
Free-living birds may also be noticed as a significant vector of pathogenic Vibrio spp. Around 12 of the roughly 100 species in the genus Vibrio are related to clinical illnesses, such as cholera and vibriosis [100]. Authors found that the burden of disease in birds was most associated with V. cholerae, followed by V. metschnikovii and V. parahaemolyticus. Prevalence of Vibrio spp. in birds varies from 43% in crows [101], and 48% in gulls, up to 65% in Magellanic penguins [102]. Researchers have discovered that migratory waterbirds can be a long-distance vector of pathogenic Vibrio species, which can be a potential threat to public health. These findings were confirmed by Fu et al. [103], who, using high-resolution genome sequencing, noticed V. parahaemolyticus strains belonging to the same clone in faeces samples derived from an area at a distance of over 1150 km. The role of migratory birds as Vibrio disease vectors was also described by Zheng et al. [104]. These authors found a significant level of variation across the strains tested, with numerous novel V. cholerae alleles among migratory birds that had not previously been identified in databases. In another study conducted in Romania by Pall et al. [101], a higher number of Vibrio strains in migratory (74.66%) than in sedentary birds (25.33%) were indicated, with increased risk of transmission to humans or the environment in places where they transit or nest.
The Yersinia genus is comprised of 28 distinct species, of which 3 have the potential to cause illness in humans. Yersinia pestis is the culprit behind the plague, while Yersinia pseudotuberculosis and Y. enterocolitica are enteropathogenic species that can lead to yersiniosis [105,106]. It has been discovered that both Y. pseudotuberculosis and Y. enterocolitica are capable of causing disease in both humans and animals [107]. Avian pseudotuberculosis is a prevalent disease found worldwide and caused by Yersinia pseudotuberculosis. It affects various species of poultry, with young turkeys being the most susceptible. Along with poultry, a diverse range of wild birds and rodents are also prone to this disease [108]. Infection caused by this pathogen is also commonly observed in canaries and wild finches during European winters. The clinical signs include non-specific symptoms such as ruffled feathers, debilitation, and high mortality [109]. This particular pathogen has also been isolated from captive toucans [110], captive pink pigeons [111], black grouse, willow ptarmigan [112], as well as amazon parrots [113], and Eurasian collared doves [114]. A study conducted by Cork et al. in 1999 [115] examined 14 cases of avian pseudotuberculosis. The research revealed that the passerine species demonstrated an acute clinical course of the disease, with predominant enteric bacterial lesions. On the other hand, the pigeons and the psittaciformes examined exhibited a more chronic clinical course of the disease. These findings contribute to the understanding of the diverse manifestations of avian pseudotuberculosis in different avian species.
Recently, a study revealed that a cat was found to have been infected with Y. pseudotuberculosis. This particular feline had been diagnosed with diabetes mellitus, acromegaly, and dysorexia. The leading theory is that the cat may have contracted the infection after having contact with natural reservoirs such as rodents or wild birds. This discovery highlights the importance of considering the possibility of transmission to humans, especially when cats share the same household as their owners [116]. In humans, Y. pseudotuberculosis not only causes normally self-limiting gastroenteritis but also pseudoappendicitis, arthritis, pharyngitis, and erythema nodosum [117,118,119,120].
Y. enterocolitica infection is primarily transmitted to humans through the consumption of contaminated food or water. The pathogenic event commences with bacterial colonization of the intestinal tract, where it causes most of the pathologic effects and clinical manifestations [121]. Y. enterocolitica is the most significant cause of yersiniosis in humans in Europe [122]. While this pathogen is widely distributed in nature in aquatic and animal reservoirs, swine are considered a major reservoir for human pathogenic strains [123]. However, some authors suggest that birds may serve as carriers, potential reservoirs, and sources of infection for humans. In 2020, the occurrence of Y. enterocolitica in the vast majority of migratory game species in Poland was studied. The prevalence of Y. enterocolitica was determined at 1.4% in green-winged teals, at 5.0% in Eurasian coots, and at 4.8% in capercaillie [124]. This pathogen has also been found in male blackcaps [125], as well as pheasants, crows, magpies, bulbuls, and starlings [126].
Fukushima et al. [127] found that the close relationship between the regional distributions of Y. pseudotuberculosis in wild animals and humans suggests that wild animals are an important source of infection, while Niskanen et al. [107] suggest that birds are unlikely to be a direct source of Yersinia infections in humans. However, the identification of Y. pseudotuberculosis and Y. enterocolitica in bird feces indicates that wild birds cannot be excluded from the epidemiological discussion of human yersiniosis.
4. Virulence Profiles and Genetic Diversity of Bacteria Isolated from Free-Living Birds
Virulence factors are bacteria-associated molecules that enhance their ability to evade the host defences and cause disease [128]. These factors are either secretory, membrane-associated, or cytosolic [129]. The prevalence of virulence profiles of bacteria is one of the most important issues in recent microbiological studies, and it is worth studying which virulence factors prevail among free-living birds.
Although birds are considered the main reservoir for Campylobacter spp., the major cause of gastroenteritis in humans, there is little knowledge about the virulence profiles of Campylobacter isolated from free-living birds. Despite the large quantity of research that has revealed the frequency of Campylobacter spp., only a few studies have conducted investigations into the virulence genes crucial to pathogenesis [18]. Although the pathogenesis of Campylobacter infection is not completely understood, multiple mechanisms are thought to be involved, and virulence factors including adhesion, bacterial invasion, and toxin generation are thought to play a role in its pathogenesis in humans [19,130]. Important virulence factors such as flaA, cadF, and ciaB are necessary for Campylobacter to adhere to and colonize epithelial cells in the host intestine [131]. The cadF, a gene encoding Campylobacter adhesion to fibronectin F [132], flaA encoding flagellin A protein, and ciaB encoding invasion antigen B have been found in many wild bird species. FlaA and cadF were present in 100% of C. jejuni isolates obtained from free-living birds in China, including crows, Daurian jackdaws, and silver pheasants, with the overall prevalence of this bacteria among wild birds at the level of 10.96% [133]. Similar results were achieved by Shyaka et al. [134], who found cadF, flaA, and ciaB in 100% of C. jejuni strains isolated from crows, pigeons, and Eurasian tree sparrows in Japan. Wei et al. [135], in epidemiological studies in South Korea, noticed among Campylobacter isolates from wild birds the common prevalence of genes flaA, cadF, racR, and dnaJ responsible for colonization and adhesion, genes ciaB and pldA, which are responsible for invasion, and genes cdtA, cdtB and cdtC encoding cytolethal distending toxin (CDT). Similar findings were confirmed by Shyaka et al. [134] in studies on the characterization of Campylobacter isolated from resident wild birds in Japan. Campylobacter, specifically C. jejuni, can trigger the polyneuropathic disorder denominated Guillain–Barré syndrome (GBS). The C. jejuni strains that can elicit GBS carry either wlaN or cgtB, coding both genes for a β-1,3-galactosyltransferase enzyme that is required to produce sialylated lipooligosaccharide (LOSSIAL). In a study by Guirado et al. [136] a similar percentage of strains were positive for LOSSIAL-related genes (wlaN and cgtB) among human (28%) and broiler chicken (22%) strains, while the percentage increased to 40% among wild bird strains. The distribution of the wlaN gene ratio shows significant differences ranging from 30% among wild birds in Ireland [137] up to 61.1% in Poland [138]. In comparison, the presence of the wlaN gene in poultry varies from 10.7% in Brazil [139] and 13.6% in Germany [140] up to 34.6% in Egypt [141]. The screening of the cgtB and the wlaN genes in Tunisian poultry revealed the presence of cgtB in 21.2% of C. jejuni strains, whereas none of them carried the wlaN gene [142]. This is concerning, as GBS may be fatal in the acute phase but also affects long-term prognosis due to irreversible sequelae and secondary medical complications [143].
While numerous pathogens are solely taken up by professional phagocytes, Listeria monocytogenes may invade nonphagocytic cells such as epithelial cells [144], hepatocytes [145], and endothelial cells [146]. As a result, L. monocytogenes may pass through three major barriers: the intestinal epithelial cell barrier, the blood–brain endothelial cell barrier, and the fetoplacental endothelial cell barrier [147]. One of the key virulence factors of Listeria monocytogenes is the ability to invade host cells and replicate intracellularly. The internalin (inl) gene family, which encodes proteins that interact with host cell receptors to mediate invasion, is one of the virulence genes implicated in this process. The most well-known members of this family, InlA and InlB, have been demonstrated to be necessary for the pathogenicity of L. monocytogenes in animal models [148]. Additionally, L. monocytogenes needs the actA gene, which encodes a protein involved in actin polymerization and motility, to move intracellularly and spread from one cell to another [149]. In a study performed by Chen [150] in the USA, the common prevalence of inlA, inlB, and actA genes was noted among L. monocytogenes strains isolated from wild birds; moreover, these strains were identified as putatively hypervirulent L. monocytogenes strains.
Listeriolysin O (LLO) is another major virulence factor in this bacterium, which is encoded by the hlyA gene. LLO is required by L. monocytogenes for virulence and is found only in virulent strains of the species [151]. Gu et al. [152] described an outbreak of listeriosis among pheasants in China and the hlyA gene sequence from the isolated strains showed 99% similarity with several L. monocytogenes strains in the GenBank database. According to Hellström et al. [35], L. monocytogenes strains are frequently similar to those detected in foods and food processing environments; thus, birds may disseminate L. monocytogenes in nature and may also contaminate foods when entering food processing environments and outdoor marketplaces.
Concerning Clostridium perfringens, its pathogenicity is primarily caused by the synthesis of various toxins, which are encoded by genes found on plasmids or inside the bacterial chromosome. Many of them have gastrointestinal tract activity, which determines the bacteria’s pathogenicity [153]. The alpha-toxin (cpa) gene, which codes for an extracellular phospholipase C, is a significant virulence factor of C. perfringens and has been linked to several illnesses, including gas gangrene and food poisoning [154]. Additionally, it has been demonstrated that the beta-toxin gene (cpb) encodes a powerful cytotoxin that generates holes in host cell membranes and causes cell death. Moreover, the epsilon-toxin gene (etx) gene encodes a necrotizing toxin that causes severe harm to host tissues and contributes to the virulence of C. perfringens [155]. In the study conducted in China by Cao et al. [156] among C. perfringens strains recovered from wild birds, the prevailing prevalence rate cpa, cpb, and etx genes were noted respectively in 100%, 81.5%, and 44.4% of the isolates. Such common prevalence of genes-affected pathogenic properties is disturbing, as C. perfringens strains producing enterotoxin (CPE) are the second most common cause of bacterial food-borne illness in the USA [153].
Staphylococcus bacteria are traditionally classified into two groups based on their capacity to coagulate blood plasma (the coagulase response); furthermore, coagulase-negative staphylococci are normally less virulent and express fewer virulence factors [157]. Bacteria in this genus can produce a variety of virulence factors, including surface proteins required for colonization and extracellular toxins responsible for tissue damage and the deactivation of host defence systems [158]. Sulikowska et al. [159] characterized Staphylococcus bacteria recovered from dead free-living birds and captive capercaillies kept in south-eastern Poland. The results of multiplex PCR for the five classical enterotoxins A–E showed that the genome of S. aureus derived from pheasants, owls, and thrushes contained the gene sea, responsible for the production of enterotoxin A. A study performed in Spain on genomic characterization of S. aureus derived from wildlife, including birds, revealed the common presence of resistance genes as well as superantigens (such as exfoliative toxins, enterotoxins, or toxic shock syndrome toxin 1) implicated in the pathogenicity of certain human S. aureus infections. These findings highlight the role of wild animals as a reservoir for some clinically relevant S. aureus strains and the importance of wildlife surveillance, because the environment–animals–humans interaction impacts the transmission and evolution dynamics of S. aureus [160].
In different studies, the prevalence of diarrhea-inducing virulence factors among E. coli strains derived from free-living birds has been noted. Research conducted in Italy by Bertelloni et al. [79] revealed the high incidence of the eae gene (encoding intimin, an outer membrane protein) of EPEC and the stx gene (encoding Shiga toxin) of STEC among E. coli strains originating from seagulls, waterfowl, and feral pigeons. These birds frequently reach and contaminate both rural and urban areas with their droppings, and may be important sources of E. coli infection for other animals and humans. Also, Chandran and Mazumder [161] confirmed the significant distribution of STEC and EPEC strains in a wide range of free-living birds in Canada, with the overall prevalence of the stx and eae virulence genes noted at mean levels of 23% and 15%, respectively. However, the distribution of these genes differs significantly among bird species; for the stx gene, the prevalence rate ranges from 91% in ravens up to 3% in songbirds, and for the eae gene, from 93% in hawks up to 4.6% in geese. Interestingly, the studies performed by Seleem et al. [162] covering the phylogenetic analysis of the obtained stx genes revealed high genetic relatedness to those isolated from human cases in countries where such birds either lived or which were in their migratory pathway. In conclusion, the study underlines the potential role of migratory birds in spreading STEC across their migratory path.
Salmonella is known to possess many virulence genes that play an important role in bacterial pathogenesis, including fimbriae, flagella, or Type III secretion systems (T3SS). These virulence factors allow bacteria to connect to host cells, penetrate tissues, and avoid detection by the host immune system [163,164,165,166]. According to a study by Iwahori et al. [167], Salmonella isolates from wild birds in Japan exhibited a comparable distribution of virulence genes to those isolated from other animal species and humans. The genes belonging to T3SS encoded by Salmonella pathogenicity island-1 (SPI-1) and -2 (SPI-2) were found in the majority of the Salmonella isolates, as were the fimbriae and flagella genes. Those findings followed studies performed by Matias et al. [168] in Brazil and by Thomas et al. [169] in Oklahoma, which emphasize that the common existence of these virulence genes shows that wild birds may serve as a reservoir for Salmonella, which might infect people and other animals.
Vibrio spp. recovered from free-living birds are known to possess various virulence traits, which contribute to their pathogenicity. The type III secretion system (T3SS) is one such virulence factor that is involved in the release of effector proteins directly into host cells [170]. It has been shown to contribute to the colonization of the host intestine and the induction of diarrhea [171]. The cholera toxin (CT), which causes severe diarrhea associated with cholera [172], is another key virulence component of Vibrio species. Although CT is most often linked with Vibrio cholerae, other Vibrio species, such as V. parahaemolyticus, have been demonstrated to generate a toxin comparable to CT, known as thermostable direct hemolysin (TDH) [173]. TDH has been discovered in certain V. alginolyticus isolates, indicating that this virulence factor may also play a role in this species’ pathogenicity [174]. In a study conducted by Zheng et al. [175], it was reported that positive V. parahaemolyticus strains were detected in mallard faeces in Japan during the month of February, while in brown-headed gull feces in Thailand, the strains were detected in the warmer months of March, September, and October. The researchers conjectured that the acquisition of virulence genes, specifically TDH, may be unrelated to factors such as season, temperature, bird species, or location. The genome analysis of V. parahaemolyticus from aquatic bird feces in Thailand proved that the organization of the T3SS genes in bird fecal isolates was almost identical to those of human clinical strains, posing public health concerns about pathogen dissemination in recreational areas [176]. According to the study, aquatic birds carry a variety of new strains of V. parahaemolyticus, which are potentially harmful and found in the marine environment. The genetic diversity of Vibrio spp. Strains makes the birds an alternative source of this bacteria.
Yersinia enterocolitica is a heterogeneous species divided into six biotypes and various serogroups. The different bioserovars exhibit distinctive virulence properties, hosts, and geographical distributions [177,178]. Pathogenic strains of Y. enterocolitica and Y. pseudotuberculosis carry chromosomal (iail, invA, and ystA) and plasmid-borne (plasmid of Yersinia virulence, pyV) genes, e.g., yadA and virF, which are necessary for full virulence [179].
In a study by Niskanen et al. [107], the authors managed to isolate three virF-positive Y. pseudotuberculosis strains and two virF-positive Y. enterocolitica strains. The strains of Y. pseudotuberculosis came from thrushes, two from song thrushes and one from a redwing. All Y. pseudotuberculosis strains were of bioserotype 1/O:2, which has been reported to be associated with human yersiniosis in different countries in Europe [180,181]. Ten of the Y. enterocolitica isolates, all isolated from barnacle geese, belonged to bioserotype 3/O:3, which is associated with human disease. Two of these strains were found to be virF positive; thus, they carried the virulence plasmid. While the other eight isolates were virF negative, they should still be considered to be potentially pathogenic to humans, since the accidental loss of the plasmid during isolation procedures is possible [107]. Odyniec et al. [124] found 1A/NI to be the predominant bioserotype of Y. enterolocolitica in wild birds. The presence of single Y. enterocolitica strains belonging to bioserotypes 1A/O:9 and 1B/NI was also demonstrated. It is noteworthy that in Poland, mallards have yielded five distinct serotyped strains of Y. enterocolitica. What piques curiosity about these findings is that three out of the five strains belonged to serotype O:8. Normally, strains that fall under the O:8 serotype are categorized as biotype 1B and are deemed the most harmful to humans [182].
5. Antimicrobial Resistance Profiles of Bacteria Isolated from Free-Living Birds
Antimicrobial resistance is a major cause of death worldwide, with the greatest burdens in low-resource settings [183]. This global health challenge involves the transfer of bacteria and genes between humans, animals, and the environment [184]. The acquisition of resistance genes is believed to increase the pathogenicity of microorganisms and the severity of infection with a large possibility of therapy failure [185].
Wild birds have been described as sentinels, reservoirs, and potential antibiotic resistance spreaders [11]. It is crucial to remember that transmission of bacteria, especially antimicrobial-resistant bacteria (ARB), between humans, farm animals, pets, and wild animals is not fully understood. Because of their long-distance mobility, wild birds may transmit ARB over long distances, contributing to the spread of these strains [66,186]. The scale of antimicrobial resistance in wildlife, despite several reports on this subject, continually seems to be incompletely known and underestimated [187,188,189,190]. Although wild birds are infrequently exposed to antimicrobial agents, their faecal waste might function as reservoirs and potential disseminators of resistant bacteria in the environment [4]. It is believed that birds contract resistant bacteria from humans and other sources by consuming infected food or water [3]. Antibiotic residues and bacteria expressing antibiotic resistance may enter the environment as a result of the spread of medicated animal manure and urban effluents into agricultural land [191]. Environmental pollution, the presence of livestock, and human density are all variables that also contribute to the incidence of resistant bacterial strains in wild birds [192,193].
One of the most significant zoonotic pathogens linked to multidrug resistance illnesses in people and birds is Campylobacter spp. [194]. Migratory wild birds may have a substantial impact on the spread of Campylobacter spp. to farm animals and their habitats, particularly poultry farms, as these bacteria reside in the digestive tracts of wild birds [195,196,197,198]. Campylobacter has developed several resistance mechanisms in response to the use of antibiotics in clinical settings and animal agriculture. As a result, antibiotic-resistant Campylobacter is becoming more common, endangering the efficacy of antibiotic treatments, and raising serious concerns for public health [199]. In the treatment of human campylobacteriosis, fluoroquinolones and macrolides are commonly considered the first line of defence [200]. The antibiotic resistance data observed in wild birds in connection with the aforementioned treatment approach are alarming. According to Casalino et al. [201], most Campylobacter isolates from wild birds show resistance to trimethoprim (52.1%), ciprofloxacin (43.7%), and enrofloxacin (31.2%), while Indykiewicz et al. [202] state that Campylobacter spp. are commonly susceptible to azithromycin (97.62%) and erythromycin (95.24%), and slightly less frequently to tetracycline (50.0%) and ciprofloxacin (47.62%). The multidrug resistance of Campylobacter strains varies from 21.56% in Italy [201] up to 72% in Egypt [198]. The findings of the study indicate that wild birds can serve as carriers of antimicrobial-resistant Campylobacter spp. strains, posing a significant threat to the well-being of both animals and humans. Extended and unwarranted utilization of antibiotics has led to the development of resistance in Campylobacter, thereby making the treatment of human infections caused by this bacterium increasingly challenging [203]. The discovery of innovative treatments is often a formidable task, particularly when contending with Gram-negative bacteria that have evolved an outer membrane as a defence mechanism against undesirable compounds. Consequently, finding new substances that can effectively combat these bacteria presents a significant challenge [204].
The capacity of Listeria species to quickly acquire resistance to any antimicrobial treatment, as has been shown with other infections important to humans, poses a new and growing hazard to human and animal health [205]. It mostly affects high-risk patient populations, where it can result in serious and fatal infections [206,207,208,209]. As a result, antibiotic therapy is frequently required to manage the illness caused by this bacterium [210]. Ampicillin, either alone or in combination with gentamicin, is the current therapy strategy for severe cases of listeriosis [211]. At present, our knowledge regarding the antibiotic resistance of Listeria spp. in wild birds is limited. Jagtap et al. [212] found the isolates gained from wild birds to be susceptible to all antibiotics when subjected to testing, while Fadel et al. [213] were not able to perform AMR testing, as L. monocytogenes isolates lost their viability during preservation. The data currently accessible pertain predominantly to farm-raised poultry. Based on the results published by Cokal et al. [214], it has been determined that 6.7% of the samples taken from both broiler and layer flocks exhibited resistance to both ampicillin and gentamicin, respectively. Additionally, findings indicated that the presence of multidrug-resistant (MDR) isolates was observed in 20% of the samples. It can be postulated that the prevalence of AMR in L. monocytogenes in the environment may lead to comparable findings in birds that are not under human care. Nonetheless, additional research is imperative to establish the extent of antibiotic resistance among these avian species.
At present, there exists a paucity of available data regarding the antimicrobial resistance exhibited by C. perfringens strains isolated from wild avian populations. Previous research has focused predominantly on assessing the susceptibility of livestock species to this pathogen. Antimicrobial susceptibility testing of ducks in China [48] showed that isolates demonstrated the highest resistance against gentamicin (95.72%), followed by bacitracin (71.05%) and lincomycin (65.79%). Despite the relatively high degree of antimicrobial resistance exhibited by these antibiotics, further data have revealed an even more concerning situation. In human medicine, penicillin and clindamycin are the preferred antibiotics for treating infections caused by C. perfringens [215], while cephalosporins are considered alternative drugs for penicillin-allergic patients [216]. It is worrisome to note that isolates showed resistance to penicillin (7.24%), cefotaxime (2.96%), and cefepime (19.08%). A significant proportion, 81.58%, of the isolates obtained have been identified as multi-drug resistant. This raises concerns about the effectiveness of current treatment regimens and underscores the need for urgent action.
It is pertinent to note that the mecA gene, responsible for encoding the penicillin-binding proteins that render beta-lactam antibiotics ineffective, is the principal cause of methicillin resistance in Staphylococcus aureus. This protein has enabled Staphylococcus aureus to evolve robust resistance mechanisms against clinically vital antibiotic classes, i.e., penicillins and glycopeptides, that are the treatment of choice in Staphylococcal infections [217]. A study by Kutkowska et al. [218] suggests that rooks from urban areas and passerine birds from the natural habitat carry antibiotic-resistant S. aureus strains, probably reflecting the presence of such isolates in environmental food sources. Their tests revealed that 19.5% of S. aureus isolates from rook samples and 37.5% from wild-living birds were resistant to methicillin. The strains of S. aureus isolated from free-living birds in southeastern Poland exhibited resistance to enrofloxacin (44.2%), tetracycline (36.4%), fusidic acid (36.4%), erythromycin (33.3%), and ampicillin (33.3%). In addition, more than 20% of staphylococcal strains were resistant to three additional antibiotics: penicillin G, clindamycin, and amoxicillin [159]. In a study conducted by Russo et al. in 2022 [3], it was found that all isolates were resistant to clindamycin and cefoxitin, while 66.6% were also resistant to erythromycin, doxycycline, tetracycline, and trimethoprim–sulfamethoxazole. The clinical community recognizes resistance to beta-lactam antibiotics (penicillins, cephalosporins, monobactams, and carbapenems) as the most perilous, due to their highly effective and wide-ranging mechanism of action against Staphylococcal infections. Furthermore, these drugs exhibit low toxicity in both humans and animals, further enhancing their therapeutic value [219].
Antimicrobial resistance in E. coli poses a public health concern worldwide [76]. This pathogen has been linked to systemic illness in birds and is the most common opportunistic enterobacteria in captive animals [80,220]. Shobrak et al. [221] captured six types of migrating wild birds and nine types of non-migrating birds from the Taif area for isolation of Escherichia spp. The prevalence range varied from 92% in non-migrating birds to 94% in migrating birds. All isolates recovered from non-migrating birds were found to be resistant to oxacillin, while all isolates recovered from migrating birds demonstrated resistance to oxacillin, chloramphenicol, oxytetracycline, and lincomycin. In the study, all examined isolates exhibited multidrug-resistant (MDR) phenotypes that were resistant to approximately 3–10 antimicrobial agents. The authors of the study posit that wild birds may have been exposed to antimicrobial residues and bacteria that had developed antibiotic resistance as a result of human waste in waterways. Additionally, certain antimicrobial agents employed in human and veterinary medicine may not be fully eliminated, resulting in their dispersion in the environment via wastewater and soil [222]. Hasan et al. [223] discovered that E. coli isolated from both domestic and wild birds was most commonly resistant to tetracycline, ampicillin, trimethoprim/sulfamethoxazole, and nalidixic acid, while Nowaczek et al. [224] found E. coli isolated from free-living birds to be resistant to gentamicin too. On average, MDR was found in 42.7% of the isolates obtained from avian samples [223,224,225,226]. In the treatment of infections caused by E. coli, such as those acquired in the community or a hospital setting, the typical approach has been to use first-line antibiotics. These include cephalosporins, fluoroquinolones, and trimethoprim–sulfamethoxazole. However, managing E. coli infections is becoming increasingly complex due to the emergence and spread of multiresistance to these commonly used antimicrobial agents [227]. The global prevalence of antibiotic resistance in E. coli over the years has been a cause for concern and emphasizes the importance of appropriate measures to prevent transmission [228].
Salmonella is widely distributed among birds, making them an excellent indication of the AMR issue affecting wildlife [229]. According to a recent study conducted in Spain, a substantial proportion of Salmonella strains isolated from wild birds demonstrated resistance to at least one of the twelve antimicrobials tested. Specifically, the study revealed that 40.5% of the strains analyzed exhibited such resistance to ciprofloxacin (29.7%), nalidixic acid (29.7%), ampicillin (10.8%), colistin (21.6%), or tetracycline (13.5%) [229]. Ciprofloxacin, nalidixic acid, and colistin are antimicrobial agents of last resort that are utilized in the treatment of human infectious diseases caused by multi-resistant bacteria. These drugs are typically employed in the context of severe, life-threatening infections [230,231]. Due to the elevated levels of antimicrobial resistance observed in these particular antibiotics, there must be ongoing and vigilant monitoring of AMR in wild bird populations. This highlights the necessity for continued surveillance of wild bird populations to identify and track AMR trends. Different antibiotic sensitivity patterns were observed by Spinu et al. [232]. They discovered that 100% of Salmonella spp. isolates from birds in the Danube Delta were resistant to penicillin, amikacin, and erythromycin. Wild birds in Turkey carried Salmonella serotypes that were all resistant to lincomycin, nalidixic acid, and penicillin [233], whereas in Italy, almost all the analyzed strains (97.7%) showed resistance to at least one class of antibiotics, with the highest resistance values observed for tetracycline (81.8%) [234]. The research findings underscore the importance of maintaining a vigilant approach to monitoring antimicrobial resistance in wild bird populations and implementing effective strategies to mitigate this issue.
The incidence of drug-resistant strains of Vibrio spp. has been observed to be on the rise [235,236]. Out of the 76 Vibrio spp. isolates obtained from migratory and sedentary birds in Romania, 81.57% showed a multidrug resistance phenotype. Additionally, 18.42% of the strains were resistant to at least three antimicrobials, which mainly included penicillin, aminoglycosides, and macrolides [101]. Aminoglycoside antibiotics (AGAs) represent a category of cationic antibiotics that exhibit broad-spectrum activity. They are widely employed for the treatment of Gram-negative bacterial infections. However, the increasing prevalence of bacterial resistance to AGAs is leading to suboptimal treatment outcomes. Consequently, the efficacy of AGAs as a treatment option for Gram-negative bacterial infections is being eroded [237]. Antibiotic sensitivity experiments performed in China revealed that 37.10% of isolates obtained from migratory birds were resistant to ampicillin [175]. The rise in the occurrence of multidrug-resistant strains of V. cholerae on a global scale is a matter of growing concern. This trend not only poses a significant challenge to public health but also underscores the need for enhanced surveillance and control measures. Given the severe consequences of cholera outbreaks, particularly in resource-limited settings, it is critical that we take proactive steps to address this emerging threat.
An area of particular concern within the field of antimicrobial resistance research pertains to the dearth of comprehensive data regarding the susceptibility patterns exhibited by Yersinia spp. strains isolated from avian hosts. Despite the critical importance of understanding the antimicrobial resistance profiles of these pathogens, the existing body of literature is notably sparse in this regard, underscoring a significant gap in our current understanding of the potential transmission dynamics and selective pressures contributing to antimicrobial resistance within avian populations. The analysis of antimicrobial susceptibility in Y. enterocollitica from wild bird strains revealed that all tested isolates were resistant to amoxicillin with clavulanic acid, ampicillin, and cefalexin. It is noteworthy that the strains isolated from migratory birds such as green-winged teals and Eurasian coots, and those obtained from birds living in a specific territory like capercaillie, showed different levels of resistance to chemotherapeutics. The strains isolated from migratory birds were also resistant to kanamycin and streptomycin, and they were resistant and intermediately resistant to cefotaxime, ceftazidime, chloramphenicol, gentamycin, and tetracycline, to which the strains isolated from the capercaillie were susceptible. Susceptibility to only one or two chemotherapeutics is a cause for concern, as migratory birds rely on public sources of drinking water [124].
6. Conclusions
The findings of this systematic literature review shed light on the intricate interplay between free-living birds and pathogenic bacteria, emphasizing the significant public health implications inherent in understanding and addressing this dynamic relationship. The movement of birds across vast geographical areas, coupled with environmental stressors and human interaction, amplifies the potential for the transmission of bacterial pathogens to both avian and human populations.
The review underscores the global nature of the transmission dynamics of bacterial pathogens carried by free-living birds. The migratory patterns of avian species facilitate the dissemination of infectious agents across continents, transcending geographical boundaries and posing challenges to traditional disease surveillance and control measures. In light of increasing urbanization and the encroachment of human settlements into avian habitats, the risks associated with zoonotic pathogens carried by birds are further compounded.
Multidisciplinary research efforts are essential to elucidate the epidemiological dynamics of pathogenic bacteria in free-living birds and their implications for public health. Collaboration among ornithologists, microbiologists, epidemiologists, and public health officials is imperative to enhance surveillance, early detection, and mitigation strategies for zoonotic disease transmission. By fostering interdisciplinary collaborations, researchers can gain a comprehensive understanding of the complex interactions between avian species and bacterial pathogens, thus informing evidence-based interventions to safeguard public health [238].
Public awareness initiatives play a pivotal role in promoting proactive measures to minimize the transmission of bacterial pathogens from free-living birds to humans. Educating the public about the potential risks associated with interactions between humans and birds can empower individuals to adopt preventive measures and mitigate the spread of zoonotic diseases. Moreover, targeted communication strategies can help dispel misconceptions and promote responsible behaviors concerning bird–human interactions.
In conclusion, by comprehensively understanding the epidemiological significance of avian species in transmitting bacterial pathogens, proactive measures can be implemented to mitigate the risks posed to both human and animal populations. Through sustained research efforts, collaborative partnerships, and public engagement, we can effectively address the challenges posed by the transmission of bacterial pathogens by free-living birds, thereby safeguarding public health in an era marked by increasing urbanization and the emergence of zoonotic illnesses.
Author Contributions
Conceptualization, A.K. and B.W.; methodology, A.K. and B.W.; formal analysis, B.W.; investigation, A.K.; Data curation, A.K.; writing—original draft preparation, A.K.; writing—reviewing and editing, B.W.; visualization, A.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Minister of Science under the Regional Initiative of Excellence Program.
Institutional Review Board Statement
Ethical review and approval were waived for this study. All data used were publicly available or pre-existing datasets, ensuring compliance with ethical standards. This decision was made considering the minimal risk and ethical implications associated with the study’s methodology, with no risks to participants’ privacy, confidentiality, or welfare.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors gratefully acknowledge the support received during the preparation of this manuscript. We appreciate the assistance of colleagues and peers for their insightful discussions and feedback.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gogu-Bogdan, M.; Cobzaru, I.; Páll, E.; Niculae, M.; Spinu, M. Wild birds as potential vectors for pathogen dissemination on migration routes in the Danube Delta Wetlands. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 890–897. [Google Scholar]
- Tsiodras, S.; Kelesidis, T.; Kelesidis, I.; Bauchinger, U.; Falagas, M.E. Human infections associated with wild birds. J. Infect. 2008, 56, 83–98. [Google Scholar] [CrossRef]
- Russo, T.P.; Minichino, A.; Gargiulo, A.; Varriale, L.; Borrelli, L.; Pace, A.; Santaniello, A.; Pompameo, M.; Fioretti, A.; Dipineto, L. Prevalence and Phenotypic Antimicrobial Resistance among ESKAPE Bacteria and Enterobacterales Strains in Wild Birds. J. Antibiot. 2022, 11, 1825. [Google Scholar] [CrossRef]
- Foti, M.; Mascetti, A.; Fisichella, V.; Fulco, E.; Orlandella, B.M.; Lo Piccolo, F. Antibiotic Resistance Assessment in Bacteria Isolated in Migratory Passeriformes Transiting through the Metaponto Territory (Basilicata, Italy). Avian Res. 2017, 8, 26. [Google Scholar] [CrossRef]
- Ahmed, Z.S.; Elshafiee, E.A.; Khalefa, H.S.; Kadry, M.; Hamza, D.A. Evidence of Colistin Resistance Genes (Mcr-1 and Mcr-2) in Wild Birds and Its Public Health Implication in Egypt. Antimicrob. Resist. Infect. Control 2019, 8, 197. [Google Scholar] [CrossRef]
- Giacopello, C.; Foti, M.; Mascetti, A. Antimicrobial Resistance Patterns of Enterobacteriaceae in European Wild Bird Species Admitted in a Wildlife Rescue. Centre. Vet. Ital. 2016, 52, 139–144. [Google Scholar] [PubMed]
- Pancerasa, M.; Ambrosini, R.; Saino, N.; Casagrandi, R. Barn swallows long-distance migration occurs between significantly temperature-correlated areas. Sci. Rep. 2018, 8, 12359. [Google Scholar] [CrossRef]
- Winkler, D.W.; Gandoy, F.A.; Areta, J.I.; Iliff, M.J.; Rakhimberdiev, E.; Kardynal, K.J.; Hobson, K.A. Long-Distance Range Expansion and Rapid Adjustment of Migration in a Newly Established Population of Barn Swallows Breeding in Argentina. Curr. Biol. 2017, 27, 1080–1084. [Google Scholar] [CrossRef]
- Choi, O.; Corl, A.; Wolfenden, A.; Lublin, A.; Ishaq, S.; Turjeman, S.; Getz, W.; Nathan, R.; Bowie, R.; Kamath, P. High-Throughput Sequencing for Examining Salmonella Prevalence and Pathogen—Microbiota Relationships in Barn Swallows. Front. Ecol. Evol. 2021, 9, 683183. [Google Scholar] [CrossRef]
- Nabi, G.; Wang, Y.; Lü, L.; Jiang, C.; Ahmad, S.; Wu, Y.; Li, D. Bats and birds as viral reservoirs: A physiological and ecological perspective. Sci. Total Environ. 2021, 754, 142372. [Google Scholar] [CrossRef] [PubMed]
- Bonnedahl, J.; Järhult, J.D. Antibiotic Resistance in Wild Birds. Ups. J. Med. Sci. 2014, 119, 113–116. [Google Scholar] [CrossRef]
- Najdenski, H.; Dimova, T.; Zaharieva, M.M.; Nikolov, B.; Petrova-Dinkova, G.; Dalakchieva, S.; Popov, K.; Hristova-Nikolova, I.; Zehtindjiev, P.; Peev, S.; et al. Migratory birds along the Mediterranean—Black Sea Flyway as carriers of zoonotic pathogens. Can. J. Microbiol. 2018, 64, 915–924. [Google Scholar] [CrossRef]
- Sauvala, M.; Woivalin, E.; Kivistö, R.; Laukkanen-Ninios, R.; Laaksonen, S.; Stephan, R.; Fredriksson-Ahomaa, M. Hunted game birds—Carriers of foodborne pathogens. Food Microbiol. 2021, 98, 103768. [Google Scholar] [CrossRef]
- Hamer, S.A.; Lehrer, E.; Magle, S.B. Wild birds as sentinels for multiple zoonotic pathogens along an urban to rural gradient in greater Chicago, Illinois. Zoonoses Public Health 2012, 59, 355–364. [Google Scholar] [CrossRef]
- Taylor, L.H.; Latham, S.M.; Woolhouse, M.E. Risk factors for human disease emergence. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2001, 29, 983–989. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). The European Union One Health 2021 Zoonoses Report. 2022. Available online: https://www.efsa.europa.eu/en/efsajournal/pub/7666 (accessed on 12 November 2023).
- Golz, J.C.; Epping, L.; Knüver, M.T.; Borowiak, M.; Hartkopf, F.; Deneke, C.; Malorny, B.; Semmler, T.; Stingl, K. Whole genome sequencing reveals extended natural transformation in Campylobacter impacting diagnostics and the pathogens adaptive potential. Sci. Rep. 2020, 10, 3686. [Google Scholar] [CrossRef] [PubMed]
- Ansarifar, E.; Riahi, S.M.; Tasara, T. Campylobacter prevalence from food, animals, human and environmental samples in Iran: A systematic review and meta-analysis. BMC Microbiol. 2023, 23, 126. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.A.; Gulhan, T. Campylobacter in Wild Birds: Is It an Animal and Public. Health Concern? Front. Microbiol. 2022, 12, 812591. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, J.S.; Atwill, E.R.; Hunter, A. Prevalence of Campylobacter and Salmonella at a squab (young pigeon) processing plant. Poult. Sci. 2001, 80, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Vashin, I.; Stoyanchev, T. Presence of Campylobacter spp. in meat and internal organs of Japanese quail (Coturnix coturnix). Trakia J. Sci. 2005, 3, 23–25. [Google Scholar]
- Kilonzo-Nthenge, A.; Nahashon, S.N.; Chen, F.; Adefope, N. Prevalence and antimicrobial resistance of pathogenic bacteria in chicken and guinea fowl. Poult. Sci. 2008, 87, 1841–1848. [Google Scholar] [CrossRef]
- Jamali, H.; Ghaderpour, A.; Radmehr, B.; Swee Chuan Wei, K.; Chai, L.C.; Ismail, S. Prevalence and antimicrobial resistance of Campylobacter species isolates in ducks and geese. Food Control. 2014, 50, 328–330. [Google Scholar] [CrossRef]
- Colles, F.M.; Ali, J.S.; Sheppard, S.K.; McCarthy, N.D.; Maiden, M.C. Campylobacter populations in wild and domesticated Mallard ducks (Anas platyrhynchos). Environ. Microbiol. Rep. 2011, 3, 574–580. [Google Scholar] [CrossRef]
- Kim, J.; Park, H.; Kim, J.; Kim, J.H.; Jung, J.I.; Cho, S.; Ryu, S.; Jeon, B. Comparative Analysis of Aerotolerance, Antibiotic Resistance, and Virulence Gene Prevalence in Campylobacter jejuni Isolates from Retail Raw Chicken and Duck Meat in South Korea. Microorganisms 2019, 7, 433. [Google Scholar] [CrossRef]
- Wysok, B.; Wojtacka, J.; Wiszniewska-Łaszczych, A.; Szteyn, J. Antimicrobial Resistance and Virulence Properties of Campylobacter Spp. Originating from Domestic Geese in Poland. Animals 2020, 10, 742. [Google Scholar] [CrossRef]
- Kwan, P.S.; Xavier, C.; Santovenia, M.; Pruckler, J.; Stroika, S.; Joyce, K.; Gardner, T.; Fields, P.I.; McLaughlin, J.; Tauxe, R.V.; et al. Multilocus sequence typing confirms wild birds as the source of a Campylobacter outbreak associated with the consumption of raw peas. Appl. Environ. Microbiol. 2014, 80, 4540–4546. [Google Scholar] [CrossRef] [PubMed]
- Abdollahpour, N.; Zendehbad, B.; Alipour, A.; Khayatzadeh, J. Wild-bird feces as a source of Campylobacter jejuni infection in children’s playgrounds in Iran. Food Control 2015, 50, 378–381. [Google Scholar] [CrossRef]
- Southern, J.P.; Smith, R.M.; Palmer, S.R. Bird attack on milk bottles: Possible mode of transmission of Campylobacter jejuni to man. Lancet 1990, 336, 1425–1427. [Google Scholar] [CrossRef] [PubMed]
- Stuart, J.; Sufi, F.; McNulty, C.; Park, P. Outbreak of campylobacter enteritis in a residential school associated with bird pecked bottle tops. Commun. Dis. Rep. CDR Rev. 1997, 7, 38–40. [Google Scholar]
- Sanad, Y.M.; Closs, G.J.; Kumar, A.; LeJeune, J.T.; Rajashekara, G. Molecular epidemiology and public health relevance of Campylobacter isolated from dairy cattle and European starlings in Ohio, USA. Foodborne Pathog. Dis. 2013, 10, 229–236. [Google Scholar] [CrossRef]
- Kovanen, S.; Rossi, M.; Pohja-Mykrä, M.; Nieminen, T.; Raunio-Saarnisto, M.; Sauvala, M.; Fredriksson-Ahomaa, M.; Hänninen, M.L.; Kivistö, R. Population Genetics and Characterization of Campylobacter jejuni Isolates from Western Jackdaws and Game Birds in Finland. Appl. Environ. Microbiol. 2019, 85, e02365-18. [Google Scholar] [CrossRef]
- Osek, J.; Lachtara, B.; Wieczorek, K. Listeria monocytogenes—How This Pathogen Survives in Food-Production Environments? Front. Microbiol. 2022, 13, 866462. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, R.; Gorris, L.; Hayman, M.J.; Whiting, R. A Review of Listeria monocytogenes: An Update on Outbreaks, Virulence, Dose-response, Ecology, and Risk Assessments. Food Control. 2017, 75, 1–13. [Google Scholar] [CrossRef]
- Hellström, S.; Kiviniemi, K.; Autio, T.; Korkeala, H. Listeria monocytogenes is common in wild birds in Helsinki region and genotypes are frequently similar with those found along the food chain. J. Appl. Microbiol. 2008, 104, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Sauders, B.D.; Overdevest, J.; Fortes, E.; Windham, K.; Schukken, Y.; Lembo, A.; Wiedmann, M. Diversity of Listeria species in urban and natural environments. Appl. Environ. Microbiol. 2012, 78, 4420–4433. [Google Scholar] [CrossRef] [PubMed]
- Milillo, S.R.; Friedly, E.C.; Saldivar, J.C.; Muthaiyan, A.; O’Bryan, C.; Crandall, P.G.; Johnson, M.G.; Ricke, S.C. A review of the ecology, genomics, and stress response of Listeria innocua and Listeria monocytogenes. Crit. Rev. Food Sci. Nutr. 2012, 52, 712–725. [Google Scholar] [CrossRef] [PubMed]
- Crespo, R.; Garner, M.M.; Hopkins, S.G.; Shah, D.H. Outbreak of Listeria monocytogenes in an urban poultry flock. BMC Vet. Res. 2013, 9, 204. [Google Scholar] [CrossRef] [PubMed]
- Saif, Y.M. Diseases of Poultry, 11th ed.; Iowa State Press: Ames, IA, USA, 2005; pp. 968–969. [Google Scholar]
- Yoshida, T.; Sugimoto, T.; Sato, M.; Hirai, K. Incidence of Listeria monocytogenes in Wild Animals in Japan. J. Vet. Med. Sci. 2000, 62, 673–675. [Google Scholar] [CrossRef] [PubMed]
- Bouttefroy, A.; Lemaître, J.P.; Rousset, A. Prevalence of Listeria sp. in droppings from urban rooks (Corvus frugilegus). J. Appl. Microbiol. 1997, 82, 641–647. [Google Scholar] [CrossRef]
- Weber, A.; Potel, J.; Schäfer-Schmidt, R.; Prell, A.; Datzmann, C. Untersuchungen zum Vorkommen von Listeria monocytogenes in Kotproben von Haus- und Heimtieren. Zentralbl Hyg. Umweltmed. 1995, 198, 117–123. [Google Scholar]
- Weis, J.; Seeliger, H.P. Incidence of Listeria monocytogenes in nature. Appl. Microbiol. 1975, 30, 29–32. [Google Scholar] [CrossRef]
- Schoder, D.; Guldimann, C.; Märtlbauer, E. Asymptomatic Carriage of Listeria monocytogenes by Animals and Humans and Its Impact on the Food Chain. Foods 2022, 11, 3472. [Google Scholar] [CrossRef] [PubMed]
- Committee against Birds Slaughter (CABS). Shooting Bag Figures for Waterfowl and Coots, Cranes, and Rails. In 533 Committee Against Bird Slaughter; 2010. Available online: https://laczanasptaki.pl/wp-content/uploads/2018/11/CABS-Hunting-in-Europe-2017.pdf (accessed on 12 November 2023).
- Kiu, R.; Hall, L.J. An update on the human and animal enteric pathogen Clostridium perfringens. Emerg. Microbes Infect. 2018, 7, 141. [Google Scholar] [CrossRef] [PubMed]
- Mora, Z.V.; Macías-Rodríguez, M.E.; Arratia-Quijada, J.; Gonzalez-Torres, Y.S.; Nuño, K.; Villarruel-López, A. Clostridium perfringens as Foodborne Pathogen in Broiler Production: Pathophysiology and Potential Strategies for Controlling Necrotic Enteritis. Animals 2020, 10, 1718. [Google Scholar] [CrossRef]
- Xiu, L.; Liu, Y.; Wu, W.; Chen, S.; Zhong, Z.; Wang, H. Prevalence and multilocus sequence typing of Clostridium perfringens isolated from 4 duck farms in Shandong province, China. Poult. Sci. 2020, 99, 5105–5117. [Google Scholar] [CrossRef] [PubMed]
- Naser, M.; Mohamed, E.; Awadallah, M.; Merwad, A.; Selim, M. Clostridium perfringens in duck farms, retail outlets and in contact workers in Egypt incidence and toxin genotyping. Egypt. J. Food Safety 2018, 5, 111–120. [Google Scholar] [CrossRef]
- Asaoka, Y.; Yanai, T.; Hirayama, H.; Une, Y.; Saito, E.; Sakai, H.; Goryo, M.; Fukushi, H.; Masegi, T. Fatal necrotic enteritis associated with Clostridium perfringens in wild crows (Corvus macrorhynchos). Avian Pathol. 2004, 33, 19–24. [Google Scholar] [CrossRef]
- Bildfell, R.J.; Eltzroth, E.K.; Songer, J.G. Enteritis as a Cause of Mortality in the Western Bluebird (Sialia mexicana). Avian Dis. 2001, 45, 760–763. [Google Scholar] [CrossRef]
- Razmyar, J.; Kalidari, G.A.; Tolooe, A.; Rad, M.; Movassaghi, A.R. Genotyping of Clostridium perfringens isolated from healthy and diseased ostriches (Struthio camelus). Iran. J. Microbiol. 2014, 6, 31–36. [Google Scholar]
- Hagen, C.A.; Bildfell, R.J. An observation of Clostridium perfringens in Greater Sage-Grouse. J. Wildl. Dis. 2007, 43, 545–547. [Google Scholar] [CrossRef]
- Chary, R.; Lucas, A. Entérotoxémie à Clostridium perfringens type A chez des faisans d’élevage. Commentaires étiologiques. Bull. Acad. Vet. Fr. 1970, 43, 351–356. [Google Scholar]
- Stuve, G.; Hofshagen, M.; Holt, G. Necrotizing lesions in the intestine, gizzard, and liver in captive capercaillies (Tetrao urogallus) associated with Clostridium perfringens. J. Wildl. Dis. 1992, 28, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Craven, S.E.; Stern, N.J.; Line, E.; Bailey, J.S.; Cox, N.A.; Fedorka-Cray, P. Determination of the incidence of Salmonella spp., Campylobacter jejuni, and Clostridium perfringens in wild birds near broiler chicken houses by sampling intestinal droppings. Avian Dis. 2000, 44, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Fathima, S.; Hakeem, W.G.A.; Shanmugasundaram, R.; Selvaraj, R.K. Necrotic Enteritis in Broiler Chickens: A Review on the Pathogen, Pathogenesis, and Prevention. Microorganisms 2022, 10, 1958. [Google Scholar] [CrossRef]
- Seki, K.; Suzuki, N.; Nagai, A.; Nakamoto, A.; Yamamoto, Y.; Kamatsuka, T.; Kimura, K.; Kikuchi, H.; Uetsuka, K. Prevalence of Clostridium perfringens in wild resident birds (Corvus spp., Sturnus cineraceus, Passer montanus) in Ibaraki prefecture. Jpn. J. Anim. Hyg. 2022, 48, 17–24. [Google Scholar]
- Chaguza, C.; Smith, J.T.; Bruce, S.A.; Gibson, R.; Martin, I.W.; Andam, C.P. Prophage-encoded immune evasion factors are critical for Staphylococcus aureus host infection, switching, and adaptation. Cell Genom. 2022, 2, 100194. [Google Scholar] [CrossRef]
- Andreasen, C.B. Staphylococcosis. Diseases of Poultry; John Wiley and Sons, Inc.: Iowa, IA, USA, 2013; pp. 971–976. [Google Scholar]
- Le, H.; Dalsgaard, A.; Skytt, A.P.; Nguyen, H.; Ta, Y.; Nguyen, T. Large-Scale Staphylococcus aureus Foodborne Disease Poisoning Outbreak among Primary School Children. Microbiol. Res. 2021, 12, 43–52. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Outbreak of staphylococcal food poisoning from a military unit lunch party—United States, 2012. Morb. Mortal. Wkly. Rep. 2013, 62, 1026–1028. [Google Scholar]
- Wang, X.; Tao, X.; Xia, X.; Yang, B.; Xi, M.; Meng, J.; Zhang, J.; Xu, B. Staphylococcus aureus and methicillin-resistant Staphylococcus aureus in retail raw chicken in China. Food Control. 2013, 29, 103–106. [Google Scholar] [CrossRef]
- Sallam, K.I.; Abd-Elghany, S.M.; Elhadidy, M.; Tamura, T. Molecular Characterization and Antimicrobial Resistance Profile of Methicillin-Resistant Staphylococcus aureus in Retail Chicken. J. Food Prot. 2015, 78, 1879–1984. [Google Scholar] [CrossRef]
- Sousa, M.; Silva, N.; Igrejas, G.; Silva, F.; Sargo, R.; Alegria, N.; Benito, D.; Gómez, P.; Lozano, C.; Gómez-Sanz, E.; et al. Antimicrobial resistance determinants in Staphylococcus spp. recovered from birds of prey in Portugal. Vet. Microbiol. 2014, 171, 436–440. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.; Lopes, A.F.; Soeiro, V.; Caniça, M.; Manageiro, V.; Pereira, J.E.; Maltez, L.; Capelo, J.L.; Igrejas, G.; Poeta, P. Nocturnal Birds of Prey as Carriers of Staphylococcus aureus and Other Staphylococci: Diversity, Antimicrobial Resistance and Clonal Lineages. Antibiotics 2022, 11, 240. [Google Scholar] [CrossRef] [PubMed]
- Vidal, A.; Baldomà, L.; Molina-López, R.A.; Martin, M.; Darwich, L. Microbiological diagnosis and antimicrobial sensitivity profiles in diseased free-living raptors. Avian Pathol. 2017, 46, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Wardyn, S.E.; Kauffman, L.K.; Smith, T.C. Methicillin-resistant Staphylococcus aureus in central Iowa wildlife. J. Wildl. Dis. 2012, 48, 1069–1073. [Google Scholar] [CrossRef]
- Thapaliya, D.; Dalman, M.; Kadariya, J.; Little, K.; Mansell, V.; Taha, M.Y.; Grenier, D.; Smith, T.C. Characterization of Staphylococcus aureus in Goose Feces from State Parks in Northeast Ohio. Ecohealth. 2017, 14, 303–309. [Google Scholar] [CrossRef]
- Ruiz-Ripa, L.; Gómez, P.; Alonso, C.A.; Camacho, M.C.; de la Puente, J.; Fernández-Fernández, R.; Ramiro, Y.; Quevedo, M.A.; Blanco, J.M.; Zarazaga, M.; et al. Detection of MRSA of Lineages CC130-mecC and CC398-mecA and Staphylococcus delphini-lnu(A) in Magpies and Cinereous Vultures in Spain. Microb. Ecol. 2019, 78, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Gómez, P.; Lozano, C.; Camacho, M.C.; Lima-Barbero, J.F.; Hernández, J.M.; Zarazaga, M.; Höfle, Ú.; Torres, C. Detection of MRSA ST3061-t843-mecC and ST398-t011-mecA in white stork nestlings exposed to human residues. J. Antimicrob. Chemother. 2016, 71, 53–57. [Google Scholar] [CrossRef]
- Awad-Alla, M.E.; Abdien, H.M.; Dessouki, A.A. Prevalence of bacteria and parasites in White Ibis in Egypt. Vet. Ital. 2010, 46, 277–286. [Google Scholar]
- Topić, N.; Cenov, A.; Jozić, S.; Glad, M.; Mance, D.; Lušić, D.; Kapetanović, D.; Mance, D.; Vukić Lušić, D. Staphylococcus aureus-An Additional Parameter of Bathing Water Quality for Crowded Urban Beaches. Int. J. Environ. Res. Public Health. 2021, 18, 5234. [Google Scholar] [CrossRef]
- Cragg, J.; Clayton, Y.M. Bacterial and fungal flora of seagull droppingsd in Jersey. J. Clin. Pathol. 1971, 24, 317–319. [Google Scholar] [CrossRef]
- Abdullahi, I.N.; Fernández-Fernández, R.; Juárez-Fernández, G.; Martínez-Álvarez, S.; Eguizábal, P.; Zarazaga, M.; Lozano, C.; Torres, C. Wild Animals Are Reservoirs and Sentinels of Staphylococcus aureus and MRSA Clones: A Problem with “One Health” Concern. Antibiotics 2021, 10, 1556. [Google Scholar] [CrossRef]
- Ong, K.H.; Khor, W.C.; Quek, J.Y. Occurrence and antimicrobial resistance traits of Escherichia coli from wild birds and rodents in Singapore. Int. J. Environ. Res. Public Health. 2020, 17, 5605. [Google Scholar] [CrossRef]
- Borges, C.A.; Cardozo, M.V.; Beraldo, L.G.; Oliveira, E.S.; Maluta, R.P.; Barboza, K.B.; Werther, K.; Ávila, F.A. Wild birds and urban pigeons as reservoirs for diarrheagenic Escherichia coli with zoonotic potential. J. Microbiol. 2017, 55, 344–348. [Google Scholar] [CrossRef]
- Navarro, A.; Cauich-Sánchez, P.I.; Trejo, A.; Gutiérrez, A.; Díaz, S.P.; Díaz, C.M.; Cravioto, A.; Eslava, C. Characterization of Diarrheagenic Strains of Escherichia coli Isolated from Cattle Raised in Three Regions of Mexico. Front. Microbiol. 2018, 9, 2373. [Google Scholar] [CrossRef] [PubMed]
- Bertelloni, F.; Lunardo, E.; Rocchigiani, G. Occurrence of Escherichia coli virulence genes in Faeces of wild birds from Central Italy. Asian Pac. J. Trop. Med. 2019, 12, 142. [Google Scholar]
- Sanches, L.A.; Gomes, M.D.S.; Teixeira, R.H.F.; Cunha, M.P.V.; Oliveira, M.G.X.; Vieira, M.A.M.; Gomes, T.A.T.; Knobl, T. Captive wild birds as reservoirs of enteropathogenic E. coli (EPEC) and Shiga-toxin producing E. coli (STEC). Braz. J. Microbiol. 2017, 48, 760–763. [Google Scholar] [CrossRef]
- Kobayashi, H.; Pohjanvirta, T.; Pelkonen, S. Prevalence and characteristics of intimin-and shiga toxin-producing Escherichia coli from gulls, pigeons and broilers in Finland. J. Vet. Med. Sci. 2002, 64, 1071–1073. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.A.; Samanta, I.; Bhat, M.A.; Nishikawa, Y. Investigation of shiga toxin-producing Escherichia coli in avian species in India. Lett. Appl. Microbiol. 2004, 39, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.; Nicoli, J.; Nascimento, T.; Diniz, C. Diarrheagenic Escherichia coli strains recovered from urban pigeons (Columba livia) in Brazil and their antimicrobial susceptibility patterns. Curr. Microbiol. 2009, 59, 302–308. [Google Scholar] [CrossRef]
- Caballero, M.; Rivera, I.; Jara, L.M.; Ulloa-Stanojlovic, F.M.; Shiva, C. 2 Isolation and molecular identification of potentially pathogenic Escherichia coli and Campylobacter jejuni in feral pigeons from an urban area in the city of Lima, Peru. Rev. Inst. Med. Trop. São Paulo 2015, 57, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Koochakzadeh, A.; Askari Badouei, M.; Zahraei Salehi, T.; Aghasharif, S.; Soltani, M.; Ehsan, M. Prevalence of shiga toxin-producing and enteropathogenic Escherichia coli in wild and pet birds in Iran. Rev. Bras. Cienc. Avic. 2015, 17, 445–450. [Google Scholar] [CrossRef]
- Hughes, L.; Bennett, M.; Coffey, P.; Elliott, J.; Jones, T.; Jones, R.; Lahuerta-Marin, A.; McNiffe, K.; Norman, D.; Williams, N. Risk factors for the occurrence of Escherichia coli virulence genes eae, stx1 and stx2 in wild bird populations. Epidemiol. Infect. 2009, 137, 1574. [Google Scholar] [CrossRef]
- Oh, J.Y.; Kang, M.S.; Hwang, H.T.; An, B.K.; Kwon, J.H.; Kwon, Y.K. Epidemiological investigation of eaeA-positive Escherichia coli and Escherichia albertii strains isolated from healthy wild birds. J. Microbiol. 2011, 49, 747–752. [Google Scholar] [CrossRef]
- Sacristán, C.; Esperón, F.; Herrera-León, S.; Iglesias, I.; Neves, E.; Nogal, V.; Muñoz, M.; de la Torre, A. Virulence genes, antibiotic resistance and integrons in Escherichia coli strains isolated from synanthropic birds from Spain. Avian Pathol. 2014, 43, 172–175. [Google Scholar] [CrossRef]
- Giovannini, S.; Pewsner, M.; Hüssy, D.; Hächler, H.; Ryser Degiorgis, M.P.; von Hirschheydt, J.; Origgi, F.C. Epidemic of salmonellosis in passerine birds in Switzerland with spillover to domestic cats. Vet. Pathol. 2013, 50, 597–606. [Google Scholar] [CrossRef]
- Cohen, E.; Azriel, S.; Auster, O.; Gal, A.; Zitronblat, C.; Mikhlin, S.; Scharte, F.; Hensel, M.; Rahav, G.; Gal-Mor, O. Pathoadaptation of the passerine-associated Salmonella enterica serovar Typhimurium lineage to the avian host. PLoS Pathog. 2021, 17, e1009451. [Google Scholar] [CrossRef]
- Mather, A.E.; Lawson, B.; de Pinna, E.; Wigley, P.; Parkhill, J.; Thomson, N.R.; Page, A.J.; Holmes, M.A.; Paterson, G.K. Genomic Analysis of Salmonella enterica Serovar Typhimurium from Wild Passerines in England and Wales. Appl. Environ. Microbiol. 2016, 82, 6728–6735. [Google Scholar] [CrossRef] [PubMed]
- Söderlund, R.; Jernberg, C.; Trönnberg, L.; Pääjärvi, A.; Ågren, E.; Lahti, E. Linked seasonal outbreaks of Salmonella Typhimurium among passerine birds, domestic cats and humans, Sweden, 2009 to 2016. Euro Surveill. 2019, 24, 1900074. [Google Scholar] [CrossRef]
- Karim, S.J.I.; Islam, M.; Sikder, T.; Rubaya, R.; Halder, J.; Alam, J. Multidrug-resistant Escherichia coli and Salmonella spp. isolated from pigeons. Vet. World. 2020, 13, 2156–2165. [Google Scholar] [CrossRef] [PubMed]
- Pasmans, F.; Van Immerseel, F.; Hermans, K.; Heyndrickx, M.; Collard, J.M.; Ducatelle, R.; Haesebrouck, F. Assessment of virulence of pigeon isolates of Salmonella enterica subsp. enterica serovar typhimurium variant copenhagen for humans. J. Clin. Microbiol. 2004, 42, 2000–2002. [Google Scholar] [CrossRef] [PubMed]
- Bhogoju, S.; Nahashon, S.; Wang, X.; Darris, C.; Kilonzo-Nthenge, A. A comparative analysis of the microbial profile of Guinea fowl and chicken using a metagenomic approach. PLoS ONE. 2018, 13, e0191029. [Google Scholar] [CrossRef]
- Tizard, I. Salmonellosis in wild birds Sem. Avian Exot. Pet. Med. 2004, 13, 50–64. [Google Scholar] [CrossRef]
- Hall, A.J.; Saito, E.K. Avian wildlife mortality events due to Salmonellosis in the United States, 1985–2004. J. Wild. Dis. 2008, 44, 585–593. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Salmonella Outbreak Linked to Wild Songbirds. Available online: https://www.cdc.gov/salmonella/typhimurium-04-21/index.html (accessed on 24 November 2023).
- Lawson, B.; de Pinna, E.; Horton, R.A.; Macgregor, S.K.; John, S.K. Epidemiological Evidence That Garden Birds Are a Source of Human Salmonellosis in England and Wales. PLoS ONE 2014, 9, e88968. [Google Scholar] [CrossRef]
- Ayala, A.J.; Ogbunugafor, C.B. When Vibrios Take Flight: A Meta-Analysis of Pathogenic Vibrio Species in Wild and Domestic Birds. Adv. Exp. Med. Biol. 2023, 1404, 295–336. [Google Scholar] [PubMed]
- Páll, E.; Niculae, M.; Brudașcă, G.F.; Ravilov, R.K.; Șandru, C.D.; Cerbu, C.; Olah, D.; Zăblău, S.; Potârniche, A.V.; Spinu, M.; et al. Assessment and Antibiotic Resistance Profiling in Vibrio Species Isolated from Wild Birds Captured in Danube Delta Biosphere Reserve, Romania. Antibiotics 2021, 10, 333. [Google Scholar] [CrossRef]
- Cardoso, M.D.; Lemos, L.S.; Roges, E.M.; de Moura, J.F.; Tavares, D.C.; Matias, C.A.R.; Rodrigues, D.P.; Siciliano, S. A comprehensive survey of Aeromonas sp. and Vibrio sp. in seabirds from southeastern Brazil: Outcomes for public health. J. Appl. Microbiol. 2018, 124, 1283–1293. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Hao, J.; Yang, Q. Long-distance transmission of pathogenic Vibrio species by migratory waterbirds: A potential threat to the public health. Sci. Rep. 2019, 9, 16303. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Zhu, L.W.; Jing, J.; Guan, J.Y.; Lu, G.J.; Xie, L.H.; Ji, X.; Chu, D.; Sun, Y.; Chen, P.; et al. Pan-Genome Analysis of Vibrio cholerae and Vibrio metschnikovii Strains Isolated from Migratory Birds at Dali Nouer Lake in Chifeng, China. Front. Vet. Sci. 2021, 8, 638820. [Google Scholar] [CrossRef] [PubMed]
- Carniel, E. The Yersinia high-pathogenicity island: An iron-uptake island. Microbes Infect. 2001, 3, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Hammerl, J.A.; Barac, A.; Bienert, A.; Demir, A.; Drüke, N.; Jäckel, C.; Matthies, N.; Jun, J.W.; Skurnik, M.; Ulrich, J.; et al. Birds Kept in the German Zoo “Tierpark Berlin” Are a Common Source for Polyvalent Yersinia pseudotuberculosis Phages. Front. Microbiol. 2022, 12, 634289. [Google Scholar] [CrossRef]
- Niskanen, T.; Waldenström, J.; Fredriksson-Ahomaa, M.; Olsen, B.; Korkeala, H. virF-positive Yersinia pseudotuberculosis and Yersinia enterocolitica found in migratory birds in Sweden. Appl. Environ. Microbiol. 2003, 69, 4670–4675. [Google Scholar] [CrossRef]
- Domanska-Blicharz, K.; Opolska, J.; Lisowska, A.; Szczotka-Bochniarz, A. Bacterial and Viral Rodent-borne Infections on Poultry Farms. An. Attempt at a Systematic Review. J. Vet. Res. 2023, 67, 1–10. [Google Scholar]
- Dorrestein, G.M. Passerines. In Handbook of Avian Medicine; Saunders Ltd.: Philadelphia, PA, USA, 2009; pp. 169–208. [Google Scholar]
- Nakamura, S.; Hayashidani, H.; Sotohira, Y.; Une, Y. Yersiniosis caused by Yersinia pseudotuberculosis in captive toucans (Ramphastidae) and a Japanese squirrel (Sciurus lis) in zoological gardens in Japan. J. Vet. Med. Sci. 2016, 78, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Shopland, S.; Barbon, A.R.; Cotton, S.; Whitford, H.; Barrows, M. Retrospective review of mortality in captive pink pigeons (nesoenas mayeri) housed in european collections: 1977–2018. J. Zoo. Wildl. Med. 2020, 51, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Hanssen, I. Yersinia pseudotuberculosis infection in captive black grouse (Tetrao lyrurus) and willow ptarmigan (Lagopus l. lagopus). Acta Vet. Scand. 1982, 23, 622–623. [Google Scholar] [CrossRef] [PubMed]
- Galosi, L.; Farneti, S.; Rossi, G.; Cork, S.C.; Ferraro, S.; Magi, G.E.; Petrini, S.; Valiani, A.; Cuteri, V.; Attili, A.R. Yersinia pseudotuberculosis, serogroup o:1a, infection in two amazon parrots (Amazona aestiva and Amazona oratrix) with hepatic hemosiderosis. J. Zoo. Wildl. Med. 2015, 46, 588–591. [Google Scholar] [CrossRef] [PubMed]
- Stoute, S.T.; Cooper, G.L.; Bickford, A.A.; Carnaccini, S.; Shivaprasad, H.L.; Sentíes-Cué, C.G. Yersinia pseudotuberculosis in Eurasian Collared Doves (Streptopelia decaocto) and Retrospective Study of Avian Yersiniosis at the California Animal Health and Food Safety Laboratory System (1990–2015). Avian Dis. 2016, 60, 82–86. [Google Scholar] [CrossRef]
- Cork, S.C.; Collins-Emerson, J.M.; Alley, M.R.; Fenwick, S.G. Visceral lesions caused by Yersinia pseudotuberculosis, serotype II, in different species of bird. Avian Pathol. 1999, 28, 393–399. [Google Scholar] [CrossRef]
- Scarpellini, R.; Giunti, M.; Bulgarelli, C.; Mondo, E.; Esposito, E.; Assirelli, G.; Piva, S. Case report: First isolation of Yersinia pseudotuberculosis from the blood of a cat. Front. Vet. Sci. 2024, 10, 1261925. [Google Scholar] [CrossRef]
- Kaasch, A.J.; Dinter, J.; Goeser, T.; Plum, G.; Seifert, H. Yersinia pseudotuberculosis bloodstream infection and septic arthritis: Case report and review of the literature. Infection 2012, 40, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M. The third international consensus definitions for Sepsis and septic shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Fukushima, H.; Matsuda, Y.; Seki, R.; Tsubokura, M.; Takeda, N.; Shubin, F.N. Geographical heterogeneity between far eastern and Western countries in prevalence of the virulence plasmid, the Superantigen Yersinia pseudotuberculosis-derived mitogen, and the high-Pathogenicity Island among Yersinia pseudotuberculosis strains. J. Clin. Microbiol. 2001, 39, 3541–3547. [Google Scholar] [CrossRef]
- Amphlett, A. Far East Scarlet-Like Fever: A Review of the Epidemiology, Symptomatology, and Role of Superantigenic Toxin: Yersinia pseudotuberculosis-Derived Mitogen, A. Open Forum Infect. Dis. 2015, 3, ofv202. [Google Scholar] [CrossRef]
- Fàbrega, A.; Vila, J. Yersinia enterocolitica: Pathogenesis, virulence and antimicrobial resistance. Enferm. Infecc. Microbiol. Clin. 2012, 30, 24–32. [Google Scholar] [CrossRef]
- European Food Safety Authority and European Centre for Disease Prevention and Control (EFSA). EU summary report on zoonoses, zoonotic agents and food-borne outbreaks 2015. EFSA J. 2016, 14, 4634–4865. [Google Scholar]
- Bottone, E.J. Yersinia enterocolitica: The charisma continues. Clin. Microbiol. Rev. 1997, 10, 257–276. [Google Scholar] [CrossRef]
- Odyniec, M.; Stenzel, T.; Ławreszuk, D.; Bancerz-Kisiel, A. Bioserotypes, Virulence Markers, and Antimicrobial Susceptibility of Yersinia enterocolitica Strains Isolated from Free-Living Birds. Biomed. Res. Int. 2020, 2020, 8936591. [Google Scholar] [CrossRef]
- Foti, M.; Rinaldo, D.; Guercio, A. Pathogenic microorganisms carried by migratory birds passing through the territory of the island of Ustica, Sicily (Italy). Avian Pathol. 2011, 40, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Ito, K.; Kubokura, Y.; Maruyama, T.; Kaneko, K.; Ogawa, M. Occurrence of Yersinia enterocolitica in wild-living birds and Japanese serows. Appl. Environ. Microbiol. 1985, 49, 198–200. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, H.; Gomyoda, M. Intestinal carriage of Yersinia pseudotuberculosis by wild birds and mammals in Japan. Appl. Environ. Microbiol. 1991, 57, 1152–1155. [Google Scholar] [CrossRef]
- Leitão, J.H. Microbial Virulence Factors. Int. J. Mol. Sci. 2020, 21, 5320. [Google Scholar] [CrossRef]
- Sharma, A.K.; Dhasmana, N.; Dubey, N.; Kumar, N.; Gangwal, A.; Gupta, M.; Singh, Y. Bacterial Virulence Factors: Secreted for Survival. Indian J. Microbiol. 2017, 57, 1–10. [Google Scholar] [CrossRef]
- Kreling, V.; Falcone, F.H.; Kehrenberg, C.; Hensel, A. Campylobacter sp.: Pathogenicity factors and prevention methods—New molecular targets for innovative antivirulence drugs? Appl. Microbiol. Biotechnol. 2020, 104, 10409–10436. [Google Scholar] [CrossRef]
- Ghorbanalizadgan, M.; Bakhshi, B.; Kazemnejad Lili, A.; Najar-Peerayeh, S.; Nikmanesh, B. A molecular survey of Campylobacter jejuni and Campylobacter coli virulence and diversity. Iran. Biomed. J. 2014, 18, 158–164. [Google Scholar] [PubMed]
- Krause-Gruszczynska, M.; Rohde, M.; Hartig, R.; Genth, H.; Schmidt, G.; Keo, T.; König, W.; Miller, W.G.; Konkel, M.E.; Backert, S. Role of the small Rho GTPases Rac1 and Cdc42 in host cell invasion of Campylobacter jejuni. Cell Microbiol. 2007, 9, 2431–2444. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Luo, J.; Huang, J.; Wang, C.; Li, M.; Wang, B.; Wang, B.; Chang, H.; Ji, J.; Sen, K.; et al. Emergence of Genetic Diversity and Multi-Drug Resistant Campylobacter jejuni from Wild Birds in Beijing, China. Front. Microbiol. 2019, 10, 2433. [Google Scholar] [CrossRef] [PubMed]
- Shyaka, A.; Kusumoto, A.; Chaisowwong, W.; Okouchi, Y.; Fukumoto, S.; Yoshimura, A.; Kawamoto, K. Virulence characterization of Campylobacter jejuni isolated from resident wild birds in Tokachi area, Japan. J. Vet. Med. Sci. 2015, 77, 967–972. [Google Scholar] [CrossRef]
- Wei, B.; Kang, M.; Jang, H.K. Genetic characterization and epidemiological implications of Campylobacter isolates from wild birds in South Korea. Transbound. Emerg. Dis. 2019, 66, 56–65. [Google Scholar] [CrossRef]
- Guirado, P.; Paytubi, S.; Miró, E.; Iglesias-Torrens, Y.; Navarro, F.; Cerdà-Cuéllar, M.; Stephan-Otto Attolini, C.; Balsalobre, C.; Madrid, C. Differential Distribution of the wlaN and cgtB Genes, Associated with Guillain-Barré Syndrome, in Campylobacter jejuni Isolates from Humans, Broiler Chickens, and Wild Birds. Microorganisms 2020, 8, 325. [Google Scholar] [CrossRef]
- Deirdre, M. Prendergast and others, Genomic diversity, virulence and source of Campylobacter jejuni contamination in Irish poultry slaughterhouses by whole genome sequencing. J. Appl. Microbiol. 2022, 133, 3150–3160. [Google Scholar]
- Andrzejewska, M.; Śpica, D.; Skowron, K.; Indykiewicz, P.; Klawe, J.J. Genetic relatedness, virulence, and drug susceptibility of Campylobacter isolated from water and wild birds. Front. Cell. Infect. Microbiol. 2022, 12, 1005085. [Google Scholar] [CrossRef]
- Sierra-Arguello, Y.M.; Perdoncini, G.; Rodrigues, L.B.; Ruschel Dos Santos, L.; Apellanis Borges, K.; Quedi Furian, T.; Pippi Salle, C.T.; de Souza Moraes, H.L.; Pereira Gomes, M.J.; Pinheiro do Nascimento, V. Identification of pathogenic genes in Campylobacter jejuni isolated from broiler carcasses and broiler slaughterhouses. Sci. Rep. 2021, 11, 4588. [Google Scholar] [CrossRef] [PubMed]
- El-Adawy, H.; Hotzel, H.; García-Soto, S.; Tomaso, H.; Hafez, H.M.; Schwarz, S.; Neubauer, H.; Linde, J. Genomic insight into Campylobacter jejuni isolated from commercial turkey flocks in Germany using whole-genome sequencing analysis. Front. Vet. Sci. 2023, 10, 1092179. [Google Scholar] [CrossRef] [PubMed]
- Aljazzar, A.; Abd El-Hamid, M.I.; El-Malt, R.M.S.; El-Gharreb, W.R.; Abdel-Raheem, S.M.; Ibrahim, A.M.; Abdelaziz, A.M.; Ibrahim, D. Prevalence and Antimicrobial Susceptibility of Campylobacter Species with Particular Focus on the Growth Promoting, Immunostimulant and Anti-Campylobacter jejuni Activities of Eugenol and Trans-Cinnamaldehyde Mixture in Broiler Chickens. Animals 2022, 12, 905. [Google Scholar] [CrossRef] [PubMed]
- Béjaoui, A.; Gharbi, M.; Bitri, S.; Nasraoui, D.; Ben Aziza, W.; Ghedira, K.; Rfaik, M.; Marzougui, L.; Ghram, A.; Maaroufi, A. Virulence Profiling, Multidrug Resistance and Molecular Mechanisms of Campylobacter Strains from Chicken Carcasses in Tunisia. Antibiotics 2022, 11, 830. [Google Scholar] [CrossRef] [PubMed]
- Levison, L.S.; Thomsen, R.W.; Andersen, H. Increased mortality following Guillain–Barré syndrome: A population-based cohort study. Eur. J. Neurol. 2022, 29, 1145–1154. [Google Scholar] [CrossRef]
- Gaillard, J.L.; Berche, P.; Mounier, J.; Richard, S.; Sansonetti, P. In vitro model of penetration and intracellular growth of Listeria monocytogenes in the human enterocyte-like cell line Caco-2. Infect. Immun. 1987, 55, 2822–2829. [Google Scholar] [CrossRef]
- Wood, S.; Maroushek, N.; Czuprynski, C.J. Multiplication of Listeria monocytogenes in a murine hepatocyte cell line. Infect. Immun. 1993, 61, 3068–3072. [Google Scholar] [CrossRef]
- Ireton, K.; Payrastre, B.; Cossart, P. The Listeria monocytogenes protein InlB is an agonist of mammalian phosphoinositide 3-kinase. J. Biol. Chem. 1999, 274, 17025–17032. [Google Scholar] [CrossRef]
- Werbrouck, H.; Grijspeerdt, K.; Botteldoorn, N.; Van Pamel, E.; Rijpens, N.; Van Damme, J.; Uyttendaele, M.; Herman, L.; Van Coillie, E. Differential inlA and inlB expression and interaction with human intestinal and liver cells by Listeria monocytogenes strains of different origins. Appl. Environ. Microbiol. 2006, 72, 3862–3871. [Google Scholar] [CrossRef]
- Pizarro-Cerda, J.; Cossart, P. Listeria monocytogenes membrane trafficking and lifestyle: The exception or the rule? Annu. Rev. Cell Dev. Biol. 2006, 22, 657–676. [Google Scholar]
- Kocks, C.; Gouin, E.; Tabouret, M.; Berche, P.; Ohayon, H.; Cossart, P. Listeria monocytogenes-induced actin assembly requires the actA gene product, a surface protein. Cell 1992, 68, 521–531. [Google Scholar] [CrossRef]
- Chen, T.; Orsi, R.H.; Chen, R.; Gunderson, M.; Roof, S.; Wiedmann, M.; Childs-Sanford, S.E.; Cummings, K.J. Characterization of Listeria monocytogenes isolated from wildlife in central New York. Vet. Med. Sci. 2022, 8, 1319–1329. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Liu, Q.; Wu, Z.; Lu, Z. Detection of hlyA Gene of Listeria Monocytogenes with Electrochemical DNA Biosensor. In Proceedings of the 2nd International Conference on Bioinformatics and Biomedical Engineering, Shanghai, China, 16–18 May 2008; pp. 375–378. [Google Scholar]
- Gu, Y.; Liang, X.; Huang, Z.; Yang, Y. Outbreak of Listeria Monocytogenes in Pheasants. Poult. Sci. 2015, 94, 2905–2908. [Google Scholar] [CrossRef] [PubMed]
- Uzal, F.A.; Freedman, J.C.; Shrestha, A.; Theoret, J.R.; Garcia, J.; Awad, M.M.; Adams, V.; Moore, R.J.; Rood, J.I.; McClane, B.A. Towards an understanding of the role of Clostridium perfringens toxins in human and animal disease. Future Microbiol. 2014, 9, 361–377. [Google Scholar] [CrossRef] [PubMed]
- Rood, J.I.; Adams, V.; Lacey, J. Clostridium perfringens and Clostridial Diseases. Cold Spring Harb. Perspect. 2018, 8. [Google Scholar]
- Li, J.; Adams, V.; Bannam, T.L.; Miyamoto, K.; Garcia, J.P.; Uzal, F.A.; Rood, J.I. Toxin plasmids of Clostridium perfringens. Microbiol. Mol. Biol. 2018, 82, 208–233. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Wang, Y.; Wang, Y.; Zhang, L.; Yan, S. Prevalence and molecular characteristics of Clostridium perfringens isolated from wild birds in China. J. Wildl. Dis. 2020, 56, 928–931. [Google Scholar]
- Foster, T. Staphylococcus. Medical Microbiology, 4th ed.; University of Texas Medical Branch. at Galveston: Galveston, TX, USA, 1996; Chapter 12. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK8448/ (accessed on 24 November 2023).
- Cunha, M.L.R.S.; Calsolari, R.A.O.; Araújo, J.P., Jr. Detection of enterotoxin and toxic shock syndrome toxin 1 genes in Staphylococcus, with emphasis on coagulase-negative staphylococci. Microbiol. Immunol. 2007, 51, 381–390. [Google Scholar] [CrossRef]
- Sulikowska, M.; Marek, A.; Jarosz, Ł.; Stępień-Pyśniak, D.; Urban-Chmiel, R. Antimicrobial Resistance and Virulence Genes in Staphylococci Isolated from Aviary Capercaillies and Free-living Birds in South-eastern Poland. J. Vet. Res. 2022, 15, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Seijas, C.; Mascarós, P.; Lizana, V.; Martí-Marco, A.; Arnau-Bonachera, A.; Chillida-Martínez, E.; Cardells, J.; Selva, L.; Viana, D.; Corpa, J.M. Genomic Characterization of Staphylococcus aureus in Wildlife. Animals 2023, 13, 1064. [Google Scholar] [CrossRef]
- Chandran, A.; Mazumder, A. Occurrence of diarrheagenic virulence genes and genetic diversity in Escherichia coli isolates from fecal material of various avian hosts in British Columbia, Canada. Appl. Environ. Microbiol. 2014, 80, 1933–1940. [Google Scholar] [CrossRef]
- Seleem, A.; Sabry, M.A.; Abdel-Moein, K.A. Migratory birds as a potential overseas transmitter of Shiga toxin-producing Escherichia coli. Int. J. Vet. Sci. Med. 2021, 9, 52–58. [Google Scholar] [CrossRef]
- Chaudhary, J.H.; Nayak, J.B.; Brahmbhatt, M.N.; Makwana, P.P. Virulence genes detection of Salmonella serovars isolated from pork and slaughterhouse environment in Ahmedabad, Gujarat. Vet. World. 2015, 8, 121–124. [Google Scholar] [CrossRef]
- Kombade, S.; Navneet, K. Pathogenicity Island in Salmonella. In Salmonella Spp.—A Global Challenge; IntechOpen: London, UK, 2021; Available online: https://www.intechopen.com/chapters/75674 (accessed on 18 October 2023).
- Nikiema, M.E.M.; Kakou-ngazoa, S.; Ky/Ba, A. Characterization of virulence factors of Salmonella isolated from human stools and street food in urban areas of Burkina Faso. BMC Microbiol. 2021, 21, 338. [Google Scholar] [CrossRef]
- Ibarra, J.A.; Steele-Mortimer, O. Salmonella--the ultimate insider. Salmonella virulence factors that modulate intracellular survival. Cell Microbiol. 2009, 11, 1579–1586. [Google Scholar] [CrossRef]
- Iwahori, A.; Umeda, K.; Ohnishi, T.; Kawanishi, M. Molecular characteristics of Salmonella isolates from wild birds in Japan. J. Vet. Med. Sci. 2021, 83, 1287–1292. [Google Scholar]
- Matias, C.A.; Pereira, I.A.; de Araújo Mdos, S.; Santos, A.F.; Lopes, R.P.; Christakis, S.; Rodrigues Ddos, P.; Siciliano, S. Characteristics of Salmonella spp. Isolated from Wild Birds Confiscated in Illegal Trade Markets, Rio de Janeiro, Brazil. Biomed. Res. Int. 2016, 2016, 3416864. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Fenske, G.J.; Antony, L. Whole genome sequencing-based detection of antimicrobial resistance and virulence in non-typhoidal Salmonella enterica isolated from wildlife. Gut Pathog. 2017, 9, 66. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Orth, K. Virulence determinants for Vibrio parahaemolyticus infection. Curr. Opin. Microbiol. 2013, 16, 70–77. [Google Scholar] [CrossRef]
- Park, K.S.; Ono, T.; Rokuda, M.; Jang, M.H.; Okada, K.; Iida, T.; Honda, T. Functional characterization of two type III secretion systems of Vibrio parahaemolyticus. Infect. Immun. 2004, 72, 6659–6665. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.B.; Morris, J.G., Jr.; Levine, M.M. Cholera. Clin. Microbiol. Rev. 1995, 8, 48–86. [Google Scholar] [CrossRef] [PubMed]
- Honda, T.; Iida, T. The pathogenicity of Vibrio parahaemolyticus and the role of the thermostable direct hemolysin and related hemolysins. Rev. Med. Microbiol. 1993, 4, 106–113. [Google Scholar] [CrossRef]
- Xu, X.; Wu, Q.; Zhang, J.; Cheng, J.; Zhang, S. Prevalence, molecular characterization, and antibiotic susceptibility of Vibrio parahaemolyticus and Vibrio alginolyticus strains isolated from cultured silver pomfret (Pampus argenteus) in China. BioMed. Res. Int. 2013. [Google Scholar]
- Zheng, L.; Yang, C.; Chen, P.; Zhu, L.; Wen, H.; Liu, M.; Guan, J.; Lu, G.; Jing, J.; Sun, S.; et al. Vibrio parahaemolyticus from Migratory Birds in China Carries an Extra Copy of tRNA-Gly and Plasmid-Mediated Quinolone Resistance Gene qnrD. Microbiol. Spectr. 2023, 11, e02170-22. [Google Scholar] [CrossRef]
- Muangnapoh, C.; Tamboon, E.; Supha, N.; Toyting, J.; Chitrak, A.; Kitkumthorn, N.; Ekchariyawat, P.; Iida, T.; Suthienkul, O. Multilocus Sequence Typing and Virulence Potential of Vibrio parahaemolyticus Strains Isolated from Aquatic Bird Feces. Microbiol. Spectr. 2022, 10, e00886-22. [Google Scholar] [CrossRef]
- Bottone, E.J. Yersinia enterocolitica: Revisitation of enduring human pathogen. Clin. Microbiol. Newsl. 2015, 37, 1–8. [Google Scholar] [CrossRef]
- Terentjeva, M.; Ķibilds, J.; Meistere, I.; Gradovska, S.; Alksne, L.; Streikiša, M.; Ošmjana, J.; Valciņa, O. Virulence Determinants and Genetic Diversity of Yersinia Species Isolated from Retail Meat. Pathogens 2022, 11, 37. [Google Scholar] [CrossRef]
- Bancerz-Kisiel, A.; Pieczywek, M.; Łada, P.; Szweda, W. The most important virulence markers of Yersinia enterocolitica and their role during infection. Genes 2018, 9, 235. [Google Scholar] [CrossRef]
- Aleksic, S.; Bockenmühl, J.; Wuthe, H.H. Epidemiology of Yersinia pseudotuberculosis in Germany, 1983–1993. Contrib. Microbiol. Immunol. 1995, 13, 55–58. [Google Scholar] [PubMed]
- Press, N.; Fyfe, M.; Bowie, W.; Kelly, M. Clinical and microbiological follow-up of an outbreak of Yersinia pseudotuberculosis serotype 1b. Scand. J. Infect. Dis. 2001, 33, 523–526. [Google Scholar] [PubMed]
- Bancerz-Kisiel, A.; Szczerba-Turek, A.; Platt-Samoraj, A.; Szweda, W. Distribution of the ymoA and ystA genes and enterotoxins Yst production by Yersinia enterocolitica strains isolated from humans and pigs. Pol. J. Vet. Sci. 2012, 15, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Larsson, D.G.J.; Flach, C.F. Antibiotic resistance in the environment. Nat. Rev. Microbiol. 2022, 20, 257–269. [Google Scholar] [PubMed]
- Abd El-Baky, R.M.; Ibrahim, R.A.; Mohamed, D.S.; Ahmed, E.F.; Hashem, Z.S. Prevalence of Virulence Genes and Their Association with Antimicrobial Resistance Among Pathogenic E. coli Isolated from Egyptian Patients with Different Clinical Infections. Infect. Drug Resist. 2020, 13, 1221–1236. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Rodríguez, C.; Alt, K.; Grobbel, M.; Hammerl, J.A.; Irrgang, A.; Szabo, I.; Stingl, K.; Schuh, E.; Wiehle, L.; Pfefferkorn, B. Wildlife as Sentinels of Antimicrobial Resistance in Germany? Front. Vet. Sci. 2021, 7, 627821. [Google Scholar] [PubMed]
- Guenther, S.; Grobbel, M.; Heidemanns, K.; Schlegel, M.; Ulrich, R.G.; Ewers, C.; Wieler, L.H. First insights into antimicrobial resistance among faecal Escherichia coli isolates from small wild mammals in rural areas. Sci. Total Environ. 2010, 408, 3519–3522. [Google Scholar] [CrossRef]
- Arnold, K.E.; Williams, N.J.; Bennett, M. Disperse abroad in the land: The role of wildlife in the dissemination of antimicrobial resistance. Biol. Lett. 2016, 12, 20160137. [Google Scholar] [CrossRef]
- Mo, S.S.; Urdahl, A.M.; Madslien, K.; Sunde, M.; Nesse, L.L.; Slettemeås, J.S.; Norström, M. What does the fox say? Monitoring antimicrobial resistance in the environment using wild red foxes as an indicator. PLoS ONE 2018, 13, e0198019. [Google Scholar] [CrossRef]
- Wasyl, D.; Zając, M.; Lalak, A.; Skarżyńska, M.; Samcik, I.; Kwit, R.; Jabłoński, A.; Bocian, Ł.; Woźniakowski, G.; Hoszowski, A.; et al. Antimicrobial Resistance in Escherichia coli Isolated from Wild Animals in Poland. Microb. Drug Resist. 2018, 24, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Blanco, G.; Lemus, J.A.; Grande, J. Microbial pollution in wildlife: Linking agricultural manuring and bacterial antibiotic resistance in red-billed choughs. Environ. Res. 2009, 109, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Skurnik, D.; Ruimy, R.; Andremont, A.; Amorin, C.; Rouquet, P.; Picard, B.; Denamur, E. Effect of human vicinity on antimicrobial resistance and integrons in animal faecal Escherichia coli. J. Antimicrob. Chemother. 2006, 57, 1215–1219. [Google Scholar] [CrossRef]
- Allen, H.K.; Donato, J.; Wang, H.H.; Cloud-Hansen, K.A.; Davies, J.; Handelsman, J. Call of the wild: Antibiotic resistance genes in natural environments. Nat. Rev. Microbiol. 2010, 8, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Neogi, S.B.; Islam, M.M.; Islam, S.K.S. Risk of multi-drug resistant Campylobacter spp. and residual antimicrobials at poultry farms and live bird markets in Bangladesh. BMC Infect. Dis. 2020, 20, 278. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.; Oh, J.; Jeong, O.; Moon, O.; Kang, M.; Jung, B.; An, B.; Youn, S.; Kim, H.; Jang, I.; et al. Prevalence of Campylobacter species in wild birds of South Korea. Avian Pathol. 2017, 46, 474–480. [Google Scholar] [CrossRef]
- Sensale, M.; Cuomo, A.; Dipineto, L.; Santaniello, A.; Calabria, M.; Menna, L.F.; Fioretti, A. Survey of Campylobacter jejuni and Campylobacter coli in different taxa and ecological guilds of migratory birds. Ital. J. Anim. Sci. 2006, 5, 291–294. [Google Scholar] [CrossRef]
- Elnohi, M.Y.; Saleha, A.A.; Jalila, A.; Khairani-Bejo, S.; Puan, C.L.; Jalo, M.I.; Fauziah, N. Wild Birds as A Probable Source of Colonization of Campylobacter in Poultry. In Proceedings of the WPSA [Malaysia Branch] and WVPA [Malaysia Branch] Scientific Conference, Serdang, Malaysia, 30 November–1 December 2013; pp. 130–131. [Google Scholar]
- Tawakol, M.M.; Nabil, N.M.; Samir, A. The potential role of migratory birds in the transmission of pathogenic Campylobacter species to broiler chickens in broiler poultry farms and live bird markets. BMC Microbiol. 2023, 23, 66. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Sahin, O.; Grover, M.; Zhang, Q. New and alternative strategies for the prevention, control, and treatment of antibiotic-resistant Campylobacter. Transl. Res. 2020, 223, 76–88. [Google Scholar] [CrossRef]
- Casalino, G.; D’Amico, F.; Dinardo, F.R.; Bozzo, G.; Napoletano, V.; Camarda, A.; Bove, A.; Lombardi, R.; D’Onghia, F.P.; Circella, E. Prevalence and Antimicrobial Resistance of Campylobacter jejuni and Campylobacter coli in Wild Birds from a Wildlife Rescue Centre. Animals 2022, 12, 2889. [Google Scholar] [CrossRef] [PubMed]
- Indykiewicz, P.; Andrzejewska, M.; Minias, P.; Śpica, D.; Kowalski, J. Prevalence and Antibiotic Resistance of Campylobacter spp. in Urban. and Rural. Black-Headed Gulls Chroicocephalus ridibundus. Ecohealth 2021, 18, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Serwecińska, L. Antimicrobials and Antibiotic-Resistant Bacteria: A Risk to the Environment and to Public Health. Water 2020, 12, 3313. [Google Scholar] [CrossRef]
- Huemer, M.; Mairpady Shambat, S.; Brugger, S.D.; Zinkernagel, A.S. Antibiotic resistance and persistence-Implications for human health and treatment perspectives. EMBO Rep. 2020, 21, e51034. [Google Scholar] [CrossRef]
- Luque-Sastre, L.; Arroyo, C.; Fox, E.M.; McMahon, B.J.; Bai, L.; Li, F.; Fanning, S. Antimicrobial Resistance in Listeria Species. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Morvan, A.; Moubareck, C.; Leclercq, A.; Hervé-Bazin, M.; Bremont, S.; Lecuit, M.; Courvalin, P.; Le Monnier, A. Antimicrobial resistance of Listeria monocytogenes strains isolated from humans in France. Antimicrob. Agents Chemother. 2010, 54, 2728–2731. [Google Scholar] [CrossRef]
- Goulet, V.; Marchetti, P. Listeriosis in 225 non-pregnant patients in 1992: Clinical aspects and outcome in relation to predisposing conditions. Scand. J. Infect. Dis. 1996, 28, 367–374. [Google Scholar] [CrossRef]
- Farber, J.M.; Peterkin, P.I. Listeria monocytogenes, a food-borne pathogen. Microbiol. Rev. 1991, 55, 476–511. [Google Scholar] [CrossRef] [PubMed]
- Mylonakis, E.; Hohmann, E.L.; Calderwood, S.B. Central nervous system infection with Listeria monocytogenes. 33 years’ experience at a general hospital and review of 776 episodes from the literature. Medicine 1998, 77, 313–336. [Google Scholar] [CrossRef]
- Olaimat, A.N.; Al-Holy, M.A.; Shahbaz, H.M.; Al-Nabulsi, A.A.; Abu Ghoush, M.H.; Osaili, T.M.; Ayyash, M.M.; Holley, R.A. Emergence of Antibiotic Resistance in Listeria monocytogenes Isolated from Food Products: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1277–1292. [Google Scholar] [CrossRef] [PubMed]
- Temple, M.E.; Nahata, M.C. Treatment of listeriosis. Ann. Pharmacother. 2000, 34, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Jagtap, U.V.; Kolhe, R.; Deshpande, P.D.; Kurkure, N.; Dhandore, C.V.; Muglikar, D.; Jadhav, S.; Nighot, N.K.; Barbuddhe, S. Isolation of Listeria monocytogenes from Peridomestic Birds and Captive Wild Animals. Curr. Sci. 2017, 113, 1783–1787. [Google Scholar] [CrossRef]
- Fadel, H.M.; Ahmed, A. Screening of crows and waterfowls for Salmonella and Listeria monocytogenes infection. Int. J. One Health 2019, 5, 9–19. [Google Scholar] [CrossRef]
- Çokal, Y.; Günaydın, E.; Goncagül, G. The investigation of the presence of Listeria species in poultry farms and antimicrobial resistance profiles of Listeria monocytogenes strains. J. İstanbul Vet. Sci. 2022, 6, 26–34. [Google Scholar] [CrossRef]
- Zhong, J.X.; Zheng, H.R.; Wang, Y.Y.; Bai, L.L.; Du, X.L.; Wu, Y.; Lu, J.X. Molecular characteristics and phylogenetic analysis of Clostridium perfringens from different regions in China, from 2013 to 2021. Front Microbiol. 2023, 14, 1195083. [Google Scholar] [CrossRef] [PubMed]
- Schwartzman, J.D.; Reller, L.B.; Wang, W.L. Susceptibility of Clostridium perfringens isolated from human infections to twenty antibiotics. Antimicrob. Agents Chemother. 1977, 11, 695–697. [Google Scholar] [CrossRef]
- Tareen, A.R.; Zahra, R. Community acquired methicillin resistant Staphylococci (CA-MRS) in fecal matter of wild birds—A ‘one health’ point of concern. J. Infect. Public. Health. 2023, 16, 877–883. [Google Scholar] [CrossRef] [PubMed]
- Kutkowska, J.; Turska-Szewczuk, A.; Kucharczyk, M. Methicillin-resistant Staphylococcus aureus and glycopeptide-resistant enterococci in fecal samples of birds from South-Eastern Poland. BMC Vet. Res. 2019, 15, 472. [Google Scholar] [CrossRef]
- Pandey, N.; Cascella, M. Beta-Lactam Antibiotics; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Mattes, B.R.; Consiglio, S.; de Almeida, B.Z.; Guido, M.C.; Orsi, R.B.; da Silva, R.M.; Costa, A.; Ferreira, A.J.P.; Knöbl, T. Influência da biossegurança na colonização instestinal por Escherichia coli em psitacídeos. Arq. Inst. Biol. 2005, 72, 13–16. [Google Scholar] [CrossRef]
- Shobrak, M.Y.; Abo-Amer, A.E. Role of wild birds as carriers of multi-drug resistant Escherichia coli and Escherichia vulneris. Braz. J. Microbiol. 2015, 45, 1199–1209. [Google Scholar] [CrossRef]
- Sarmah, A.K.; Meyer, M.T.; Boxall, A.B. A global perspective on the use, sales, exposure pathways, occurrence, fate and effects of veterinary antibiotics (VAs) in the environment. Chemosphere 2006, 65, 725–759. [Google Scholar] [CrossRef] [PubMed]
- Hasan, B.; Sandegren, L.; Melhus, A.; Drobni, M.; Hernandez, J.; Waldenström, J.; Alam, M.; Olsen, B. Antimicrobial drug-resistant Escherichia coli in wild birds and free-range poultry, Bangladesh. Emerg. Infect. Dis. 2012, 18, 2055–2058. [Google Scholar] [CrossRef] [PubMed]
- Nowaczek, A.; Dec, M.; Stępień-Pyśniak, D.; Urban-Chmiel, R.; Marek, A.; Różański, P. Antibiotic Resistance and Virulence Profiles of Escherichia coli Strains Isolated from Wild Birds in Poland. Pathogens 2021, 10, 1059. [Google Scholar] [CrossRef]
- Yuan, Y.; Liang, B.; Jiang, B.W.; Zhu, L.W.; Wang, T.C.; Li, Y.G.; Liu, J.; Guo, X.J.; Ji, X.; Sun, Y. Migratory wild birds carrying multidrug-resistant Escherichia coli as potential transmitters of antimicrobial resistance in China. PLoS ONE 2021, 16, e0261444. [Google Scholar] [CrossRef]
- Mohamed, M.I.; Abu, J.; Zakaria, Z.; Khan, A.R.; Abdul Aziz, S.; Bitrus, A.A.; Habib, I. Multi-Drug Resistant Pathogenic Escherichia coli Isolated from Wild Birds, Chicken, and the Environment in Malaysia. Antibiotics 2022, 11, 1275. [Google Scholar] [CrossRef]
- Arbab, S.; Ullah, H.; Wang, W.; Zhang, J. Antimicrobial drug resistance against Escherichia coli and its harmful effect on animal health. Vet. Med. Sci. 2022, 8, 1780–1786. [Google Scholar] [CrossRef] [PubMed]
- Salleh, M.Z.; Nik Zuraina, N.M.N.; Hajissa, K.; Ilias, M.I.; Deris, Z.Z. Prevalence of Multidrug-Resistant Diarrheagenic Escherichia coli in Asia: A Systematic Review and Meta-Analysis. Antibiotics 2022, 11, 1333. [Google Scholar] [CrossRef] [PubMed]
- Martín-Maldonado, B.; Vega, S.; Mencía-Gutiérrez, A.; Lorenzo-Rebenaque, L.; de Frutos, C.; González, F.; Revuelta, L.; Marin, C. Urban birds: An important source of antimicrobial resistant Salmonella strains in Central Spain. Comp. Immunol. Microbiol. Infect. Dis. 2020, 72, 101519. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2017. EFSA J. 2019, 17, 5598. [Google Scholar]
- Kern, W.V. Multiresistente Bakterien: Antibiotikaverordnung und Reserveantibiotika. Dtsch. Med. Wochenschr. 2018, 143, 643–650. [Google Scholar] [CrossRef]
- Spinu, M.; Gurzau, A.E.; Niculae, M.; Brudasca, G.F.; Sandru, C.D.; Krieb, C.; Páll, E. Reciprocal relationships in antibiotic resistance of Salmonella spp. carried by wild birds and fish in the danube delta. Int. J. Infect. Dis. 2019, 79, 43. [Google Scholar] [CrossRef]
- Şahan Yapicier, O.; Hesna Kandir, E.; Öztürk, D. Antimicrobial Resistance of E. coli and Salmonella Isolated from Wild Birds in a Rehabilitation Center in Turkey. Arch. Razi Inst. 2022, 77, 257–267. [Google Scholar] [PubMed]
- Botti, V.; Navillod, F.V.; Domenis, L.; Orusa, R.; Pepe, E.; Robetto, S.; Guidetti, C. Salmonella spp. and antibiotic-resistant strains in wild mammals and birds in north-western Italy from 2002 to 2010. Vet. Ital. 2013, 49, 195–202. [Google Scholar] [PubMed]
- Mukhopadhyay, A.K.; Garg, S.; Mitra, R.; Basu, A.; Rajendran, K.; Dutta, D.; Bhattacharya, S.K.; Shimada, T.; Takeda, T.; Takeda, Y.; et al. Temporal shifts in traits of Vibrio cholerae strains isolated from hospitalized patients in Calcutta: A 3-year (1993 to 1995) analysis. J Clin Microbiol. 1996, 34, 2537–2543. [Google Scholar] [CrossRef]
- Das, B.; Verma, J.; Kumar, P.; Ghosh, A.; Ramamurthy, T. Antibiotic resistance in Vibrio cholerae: Understanding the ecology of resistance genes and mechanisms. Vaccin. 2020, 38, 83–92. [Google Scholar] [CrossRef]
- Tian, C.; Yuan, M.; Tao, Q.; Xu, T.; Liu, J.; Huang, Z.; Wu, Q.; Pan, Y.; Zhao, Y.; Zhang, Z. Discovery of Novel Resistance Mechanisms of Vibrio parahaemolyticus Biofilm against Aminoglycoside Antibiotics. Antibiotics 2023, 12, 638. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, J.S.; Jeggo, M. The One Health Approach-Why Is. It So Important? Trop. Med. Infect. Dis. 2019, 4, 88. [Google Scholar]
Figure 1.
PRISMA flow diagram showing article selection, documenting included and excluded articles at each stage.
Figure 1.
PRISMA flow diagram showing article selection, documenting included and excluded articles at each stage.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kobuszewska, A.; Wysok, B. Pathogenic Bacteria in Free-Living Birds, and Its Public Health Significance. Animals 2024, 14, 968. https://0-doi-org.brum.beds.ac.uk/10.3390/ani14060968
AMA Style
Kobuszewska A, Wysok B. Pathogenic Bacteria in Free-Living Birds, and Its Public Health Significance. Animals. 2024; 14(6):968. https://0-doi-org.brum.beds.ac.uk/10.3390/ani14060968
Chicago/Turabian StyleKobuszewska, Aleksandra, and Beata Wysok. 2024. "Pathogenic Bacteria in Free-Living Birds, and Its Public Health Significance" Animals 14, no. 6: 968. https://0-doi-org.brum.beds.ac.uk/10.3390/ani14060968
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.