The Effects of Sea and Road Transport on Physiological and Electroencephalographic Responses in Brahman Crossbred Heifers


 , , , ,
, , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Transport and Treatment
2.2. Housing and Management
2.3. Physiological and Haematological Indicators
2.4. Electroencephalogram Recording
2.5. Statistical Analysis
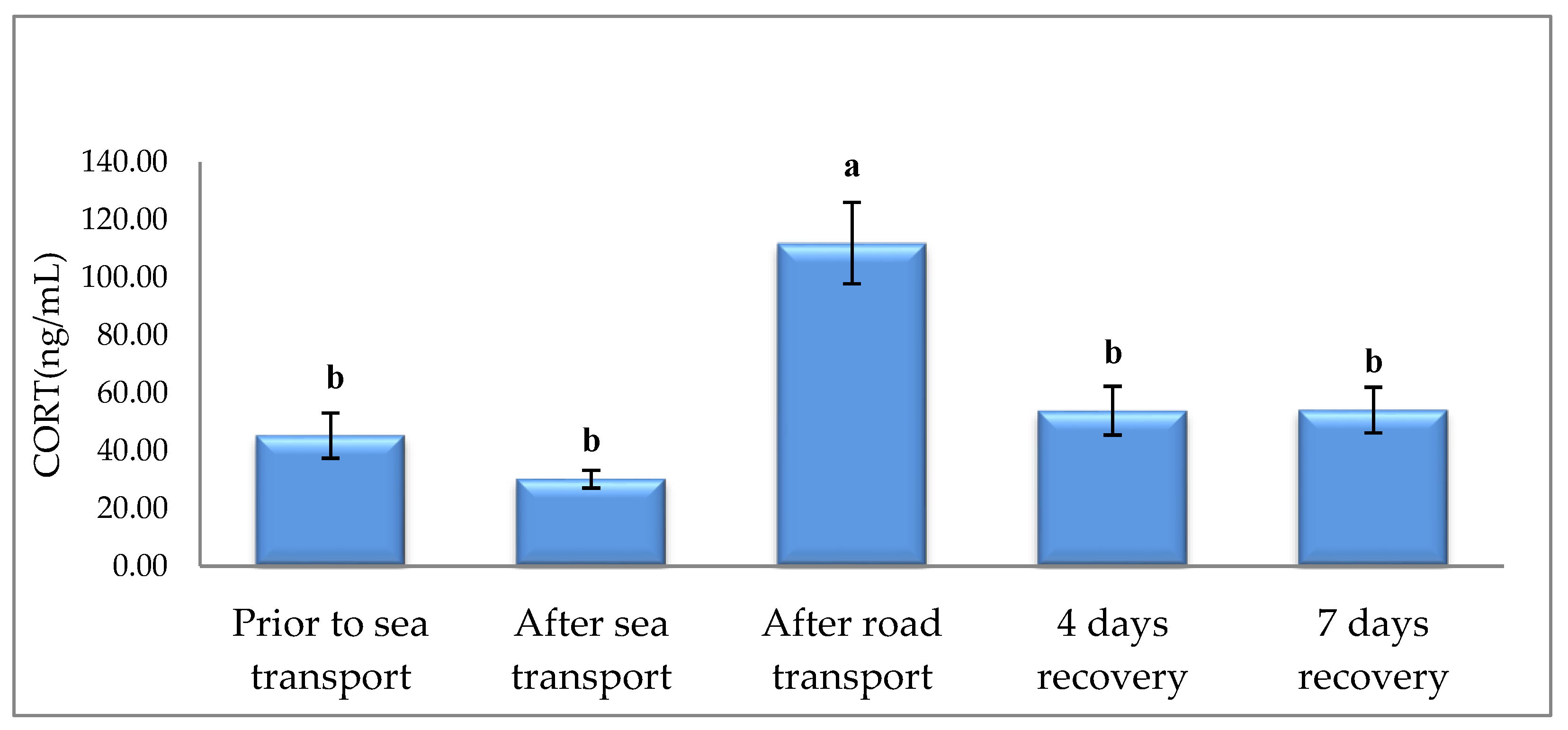
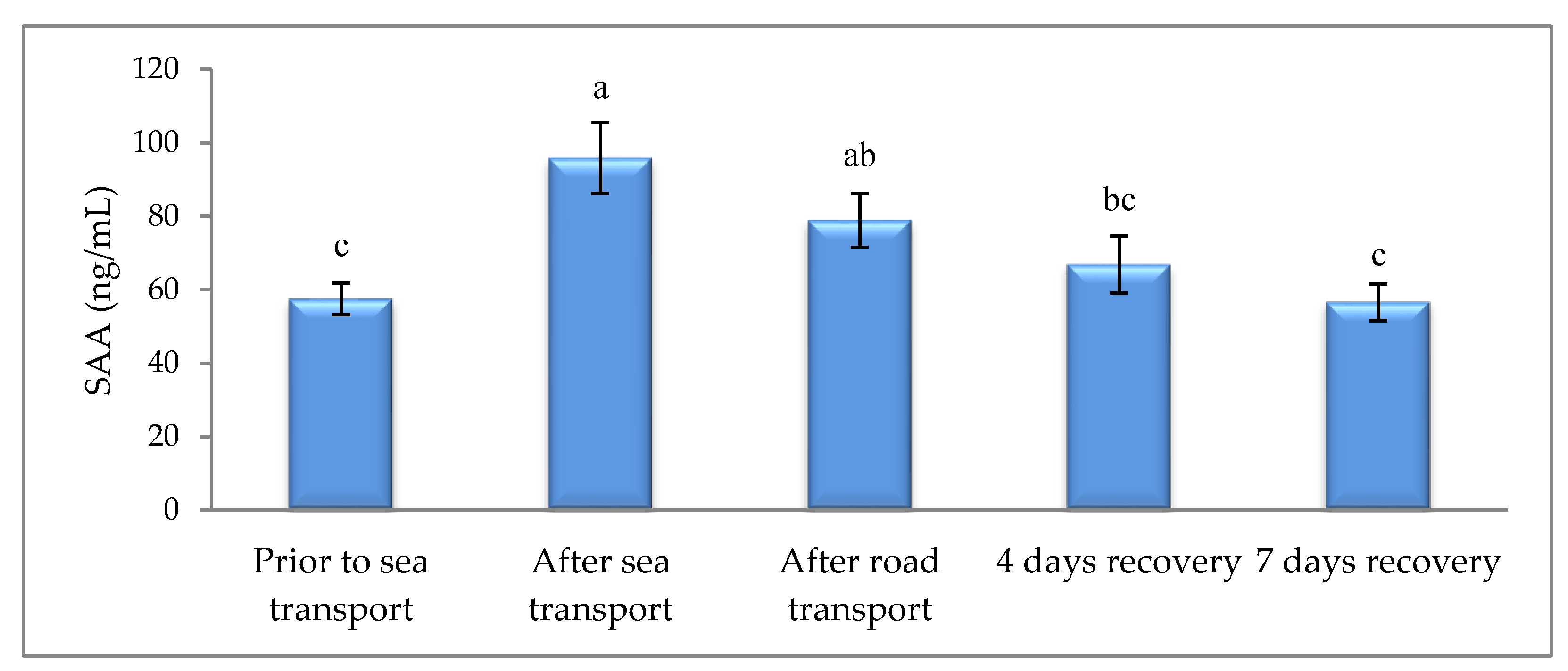
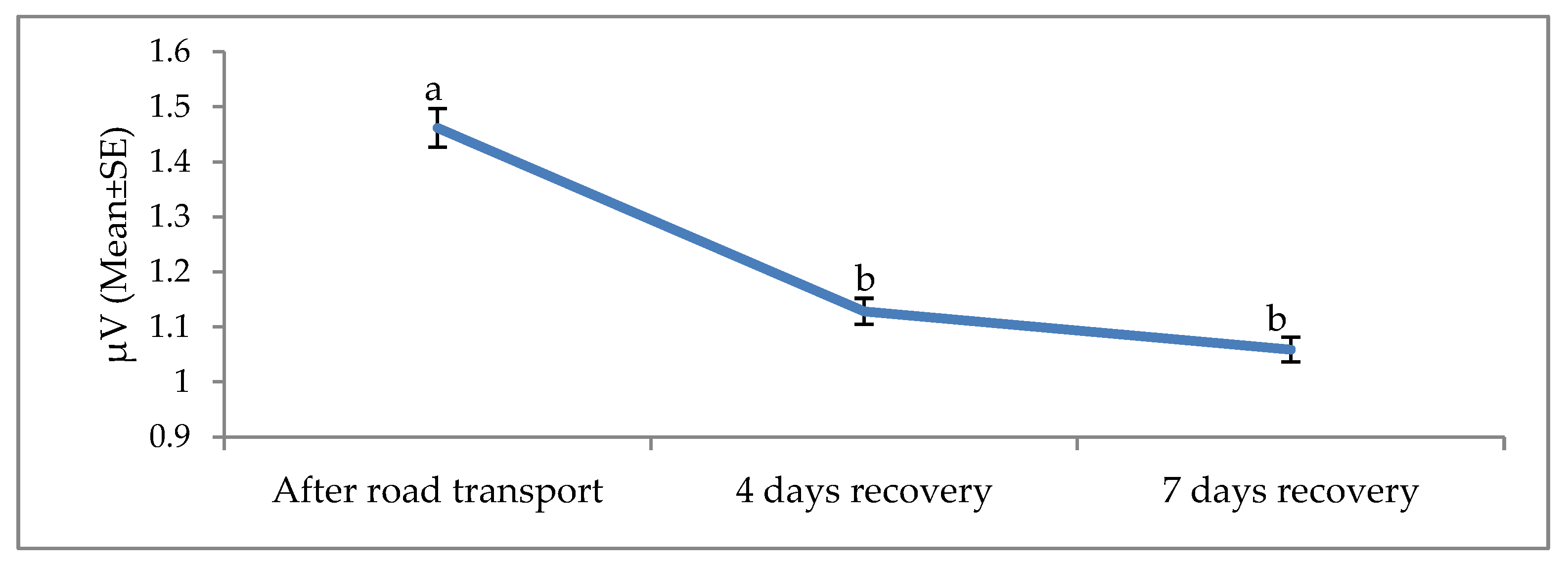
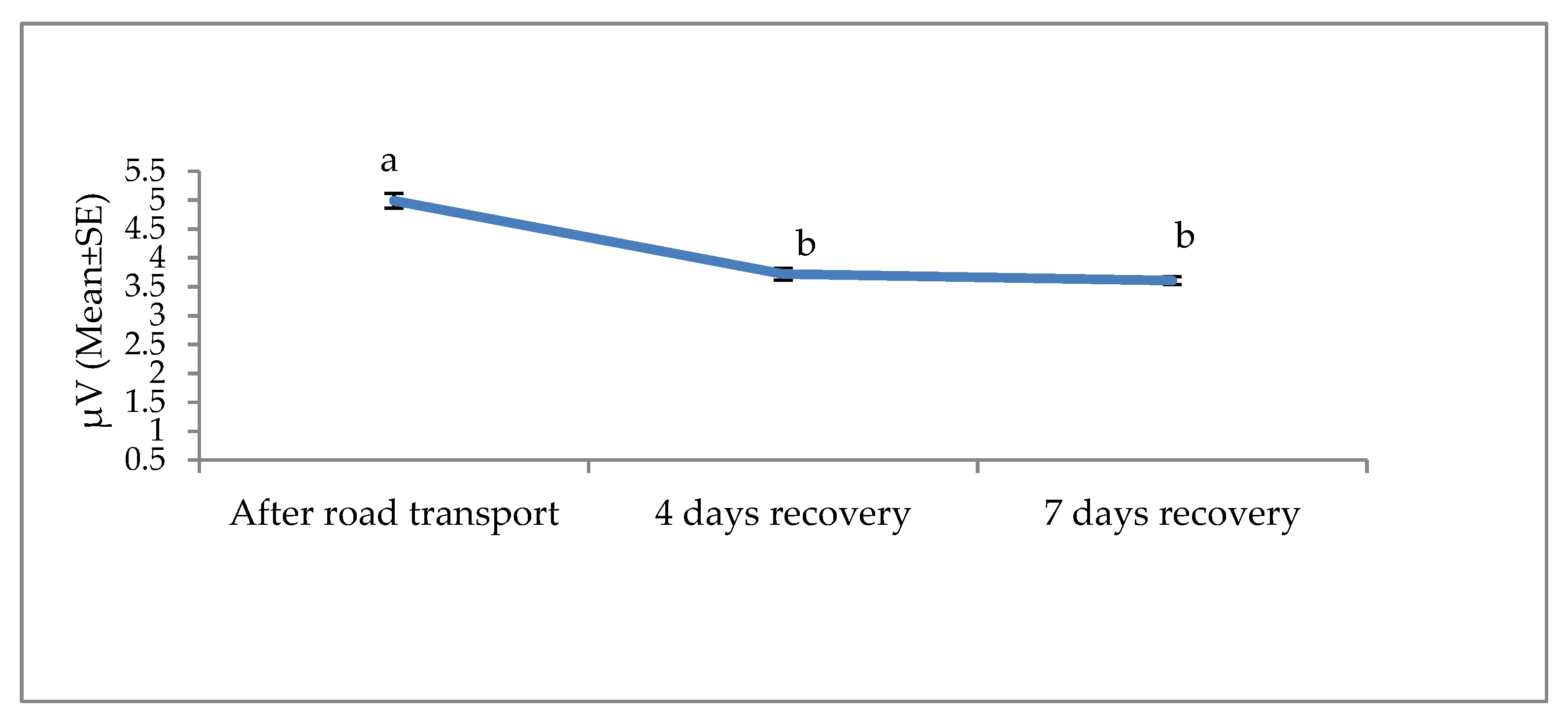
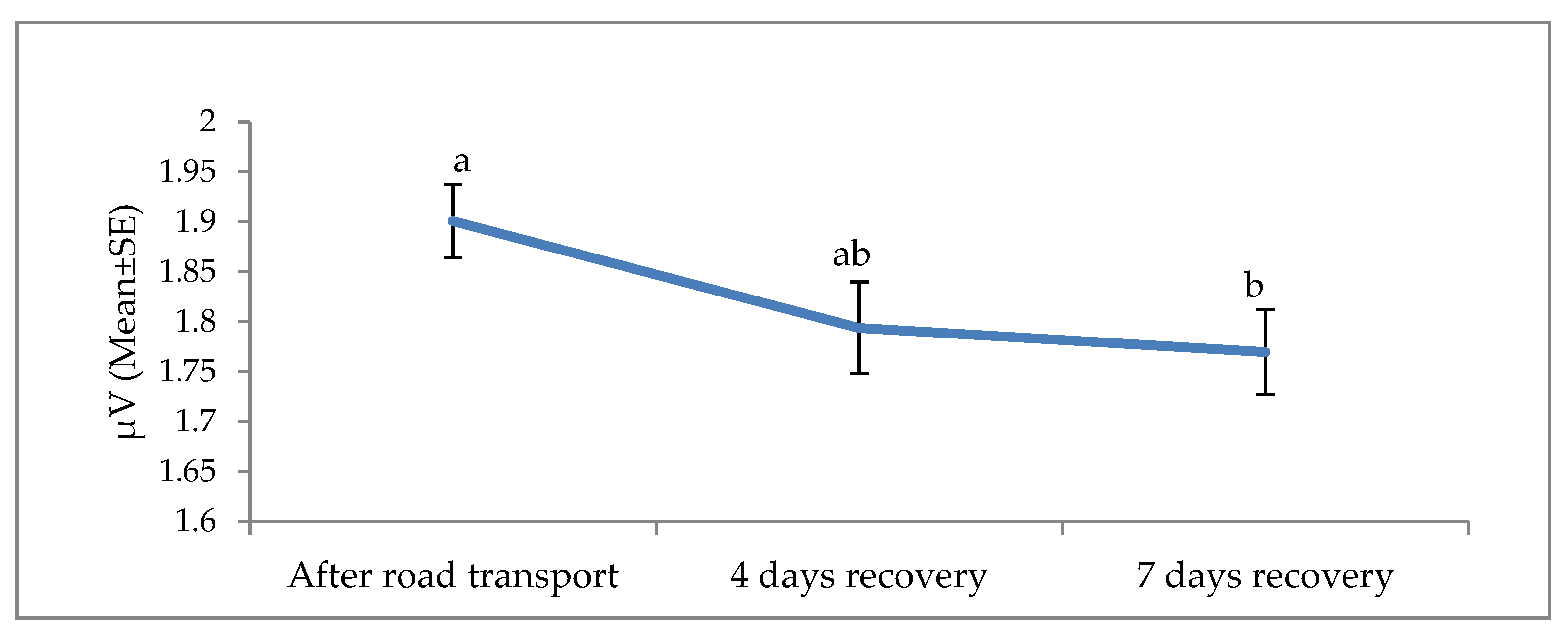
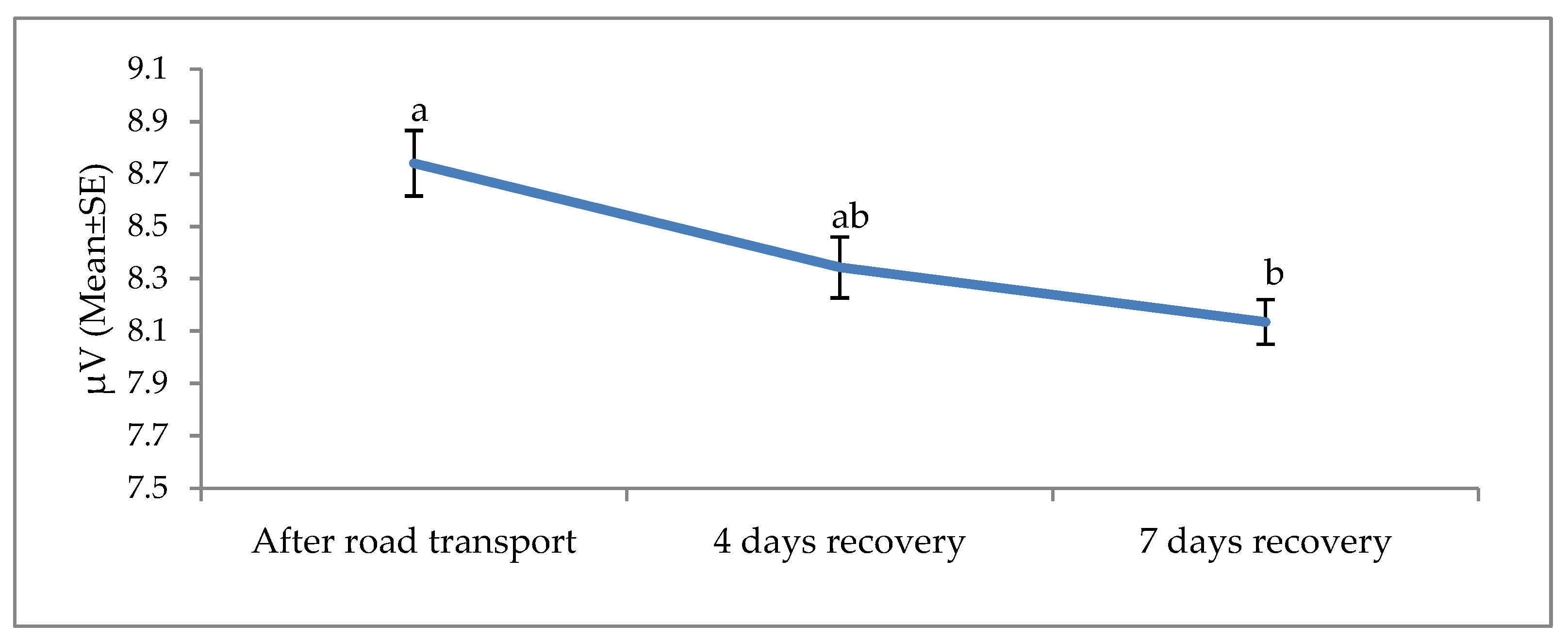
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Phillips, C.J.C. The Animal Trade; CAB International: Wallingford, UK, 2015. [Google Scholar]
- Collins, T.; Hampton, J.; Barnes, A. A systematic review of heat load in Australian livestock transported by sea. Animals 2018, 8, 164. [Google Scholar] [CrossRef]
- Nicol, C.J.; Scott, G.B. Pre-slaughter handling and transport of broiler chickens. Appl. Anim. Behav. Sci. 1990, 28, 57–73. [Google Scholar] [CrossRef]
- Von Holleben, K.; Henke, S.; Schmidt, T.; Bostelmann, N.; Hartung, J. Handling of slaughter cattle in pre and post transport situations including loading and unloading on journeys up to 8 hours in Germany. Dtw. Dtsch. Tierarztl. Wochenschr. 2003, 110, 93–99. [Google Scholar]
- Minka, N.S.; Ayo, J.O. Physiological responses of transported goats treated with ascorbic acid during the hot-dry season. Anim. Sci. J. 2007, 78, 164–172. [Google Scholar] [CrossRef]
- Padalino, B.; Tullio, D.; Cannone, S.; Bozzo, G. Road transport of farm animals: Mortality, morbidity, species and country of origin at a Southern Italian control post. Animals 2018, 8, 155. [Google Scholar] [CrossRef] [PubMed]
- Zulkifli, I. Effects of Early Age Feed Restriction and Dietary Ascorbic Acid on Heterophil/ Lymphocyte and Tonic Immobility Reactions of Transported Broiler Chickens. Asian-Australas. J. Anim. Sci. 2003, 16, 1545–1549. [Google Scholar] [CrossRef]
- Al-Aqil, A.; Zulkifli, I. Changes in heat shock protein 70 expression and blood characteristics in transported broiler chickens as affected by housing and early age feed restriction. Poult. Sci. 2009, 88, 1358–1364. [Google Scholar] [CrossRef] [Green Version]
- Zulkifli, I.; Che Norma, M.T.; Chong, C.H.; Loh, T.C. The effects of crating and road transportation on stress and fear responses of broiler chickens treated with ascorbic acid. Arch. Geflugelkd. 2000, 65, 33–37. [Google Scholar]
- Zulkifli, I.; Al-Aqil, A.; Sazili, A.Q. Fear-Related Behaviour, Muscle Glycogen Stores and Serum Creatine Kinase Activity in Transported Broiler Chickens as Affected by Housing and Early Age Feed Restriction. J. Anim. Vet. Adv. 2012, 11, 364–369. [Google Scholar] [CrossRef]
- Zulkifli, I.; Azalea Han, O.; Soleimani, A.F.; Babjee, S.A. Transportation and Translocation Effects on Leucocytic and Behavioural Responses: A Comparison Between the Red Jungle Fowl and Broiler. J. Anim. Vet. Adv. 2010, 9, 1389–1394. [Google Scholar] [CrossRef]
- Rajion, M.A.; Saat, I.M.; Zulkifli, I.; Goh, Y.M. The effects of road transportation on some physiological stress measures in goats. Asian-Australas. J. Anim. Sci. 2001, 14, 1250–1252. [Google Scholar] [CrossRef]
- Zulkifli, I.; Bahyuddin, N.; Wai, C.Y.; Farjam, A.S.; Sazili, A.Q.; Rajion, M.A.; Goh, Y.M. Physiological responses in goats subjected to road transportation under the hot, humid tropical conditions. Int. J. Agric. Biol. 2010, 12, 840–844. [Google Scholar]
- Nakyinsige, K.; Sazili, A.Q.; Aghwan, Z.A.; Zulkifli, I.; Goh, Y.M.; Fatimah, A.B. Changes in blood constituents of rabbits subjected to transportation under hot, humid tropical conditions. Asian-Australas. J. Anim. Sci. 2013, 26, 874. [Google Scholar] [CrossRef] [PubMed]
- Australian Bureau of Statistics. The Total Export Numbers for Cattle Exported by Air and Sea in 2017. Available online: http://www.livecorp.com.au/LC/files/c1/c1690e97-e20a-45ec-867b-3427eb572a3d.pdf/ (accessed on 25 December 2018).
- Norris, R.T.; Richards, R.B.; Creeper, J.H.; Jubb, T.F.; Madin, B.; Kerr, J.W. Cattle deaths during sea transport from Australia. Aust. Vet. J. 2003, 81, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Broom, D.M. Welfare assessment and welfare problem areas during handling and transport. In Livestock Handling and Transport; Grandin, T., Ed.; CABI: Wallingford, UK, 2000; Volume 2, pp. 43–61. [Google Scholar]
- Lomborg, S.R.; Nielsen, L.R.; Heegaard, P.M.; Jacobsen, S. Acute phase proteins in cattle after exposure to complex stress. Vet. Res. Commun. 2008, 32, 575–582. [Google Scholar] [CrossRef]
- Cooke, R.F.; Guarnieri Filho, T.A.; Cappellozza, B.I.; Bohnert, D.W. Rest stops during road transport: Impacts on performance and acute-phase protein responses of feeder cattle. J. Anim. Sci. 2013, 91, 5448–5454. [Google Scholar] [CrossRef] [PubMed]
- Knowles, T.G.; Warriss, P.D.; Vogel, K. Stress physiology of animals during transport. In Livestock Handling and Transport: Theories and Applications; Grandin, T., Ed.; CABI Publishing: Wallingford, UK, 2014; pp. 399–420. [Google Scholar]
- Werner, M.; Hepp, C.; Soto, C.; Gallardo, P.; Bustamante, H.; Gallo, C. Effects of a long distance transport and subsequent recovery in recently weaned crossbred beef calves in Southern Chile. Livest. Sci. 2013, 152, 42–46. [Google Scholar] [CrossRef]
- Chang, P.F.; Arendt-Nielsen, L.; Chen, A.C. Comparative cerebral responses to non-painful warm vs. cold stimuli in man: EEG power spectra and coherence. Int. J. Psychophysiol. 2005, 55, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Tops, M.; van Peer, J.M.; Wester, A.E.; Wijers, A.A.; Korf, J. State-dependent regulation of cortical activity by cortisol: An EEG study. Neurosci. Lett. 2006, 404, 39–43. [Google Scholar] [CrossRef] [Green Version]
- McAllister-Williams, R.H.; Massey, A.E.; Fairchild, G. Repeated cortisol administration attenuates the EEG response to buspirone in healthy volunteers: Evidence for desensitization of the 5-HT1 A autoreceptor. J. Psychopharmacol. 2007, 21, 826–832. [Google Scholar] [CrossRef]
- Murrell, J.C.; Johnson, C.B. Neurophysiological techniques to assess pain in animals. J. Vet. Pharmacol. Ther. 2006, 29, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Gibson, T.J.; Johnson, C.B.; Murrell, J.C.; Hulls, C.M.; Mitchinson, S.L.; Stafford, K.J.; Johnstone, A.C.; Mellor, D.J. Electroencephalographic responses of halothane-anaesthetised calves to slaughter by ventral-neck incision without prior stunning. N. Z. Vet. J. 2009, 57, 77–83. [Google Scholar] [CrossRef]
- Zulkifli, I.; Goh, Y.M.; Norbaiyah, B.; Sazili, A.Q.; Lotfi, M.; Soleimani, A.F.; Small, A.H. Changes in blood parameters and electroencephalogram of cattle as affected by different stunning and slaughter methods in cattle. Anim. Prod. Sci. 2014, 54, 187. [Google Scholar] [CrossRef]
- SAS. SAS/STAT Software, version 9.4; SAS Inst. Inc.: Cary, NC, USA, 2005. [Google Scholar]
- Earley, B.; Murray, M.; Prendiville, D.J.; Pintado, B.; Borque, C.; Canali, E. The effect of transport by road and sea on physiology, immunity and behaviour of beef cattle. Res. Vet. Sci. 2012, 92, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Blum, J.W.; Jans, F.; Moses, W.; Fröhli, D.; Zemp, M.; Wanner, M.; Hart, I.C.; Thun, R.; Keller, U. Twentyfour-Hour Pattern of Blood Hormone and Metabolite Concentrations in High-Yielding Dairy Cows: Effects of Feeding Low or High Amounts of Starch, or Crystalline Fat 6. Zent. Vet. Reihe A 1985, 32, 401–418. [Google Scholar] [CrossRef]
- Matteri, R.L.; Carroll, J.A.; Dyer, C.J. Neuroendocrine responses to stress. In The Biology of Animal Stress; Moberg, G.P., Mench, J.A., Eds.; CABI Publishing: Wallingford, UK, 2000; pp. 43–76. [Google Scholar]
- Davis, A.K.; Maney, D.L.; Maerz, J.C. The use of leukocyte profiles to measure stress in vertebrates: A review for ecologists. Funct. Ecol. 2008, 22, 760–772. [Google Scholar] [CrossRef]
- Yagi, Y.; Shiono, H.; Chikayama, Y.; Ohnuma, A.; Nakamura, I.; Yayou, K.-I. Transport stress increases somatic cell counts in milk, and enhances the migration capacity of peripheral blood neutrophils of dairy cows. J. Vet. Med. Sci. 2004, 66, 381–387. [Google Scholar] [CrossRef]
- Fazio, E.; Medica, P.; Alberghina, D.; Cavaleri, S.; Ferlazzo, A. Effect of long-distance road transport on thyroid and adrenal function and haematocrit values in Limousin cattle: Influence of body weight decrease. Vet. Res. Commun. 2005, 29, 713–719. [Google Scholar] [CrossRef]
- Gupta, S.; Earley, B.; Crowe, M.A. Effect of 12-hour road transportation on physiological, immunological and haematological parameters in bulls housed at different space allowances. Vet. J. 2007, 173, 605–616. [Google Scholar] [CrossRef]
- Buckham Sporer, K.R.; Weber, P.S.; Burton, J.L.; Earley, B.; Crowe, M.A. Transportation of young beef bulls alters circulating physiological parameters that may be effective biomarkers of stress. J. Anim. Sci. 2008, 86, 1325–1334. [Google Scholar] [CrossRef]
- Sporer, K.R.; Xiao, L.; Tempelman, R.J.; Burton, J.L.; Earley, B.; Crowe, M.A. Transportation stress alters the circulating steroid environment and neutrophil gene expression in beef bulls. Vet. Immunol. Immunopathol. 2008, 121, 300–320. [Google Scholar] [CrossRef] [PubMed]
- Earley, B.; McDonnell, B.; Murray, M.; Prendiville, D.J.; Crowe, M.A. The effect of sea transport from Ireland to the Lebanon on inflammatory, adrenocortical, metabolic and behavioural responses of bulls. Res. Vet. Sci. 2011, 91, 454–464. [Google Scholar] [CrossRef]
- Broom, D.M. The welfare of livestock during road transport. In Long Distance Transport and the Welfare of Farm Animals; Appleby, M.C., Cussen, V.A., Garces, L., Lambert, L.A., Turner, J., Eds.; CABI: Wallingford, UK, 2008; pp. 157–181. [Google Scholar]
- Navarro, G.; Col, R.; Phillips, C.J.C. Effects of space allowance and simulated sea transport motion on behavioural and physiological responses of sheep. Appl. Anim. Behav. Sci. 2018, 2018, 40–48. [Google Scholar] [CrossRef]
- Murata, H.; Shimada, N.; Yoshioka, M. Current research on acute phase proteins in veterinary diagnosis: An overview. Vet. J. 2004, 168, 28–40. [Google Scholar] [CrossRef]
- O’reilly, E.L.; Eckersall, P.D. Acute phase proteins: A review of their function, behaviour and measurement in chickens. Worlds Poult. Sci. J. 2014, 70, 27–44. [Google Scholar] [CrossRef]
- Alsemgeest, S.P.; Lambooy, I.E.; Wierenga, H.K.; Dieleman, S.J.; Meerkerk, B.; van Ederen, A.M.; Niewold, T.A. Influence of physical stress on the plasma concentration of serum amyloid-A (SAA) and haptoglobin (Hp) in calves. Vet. Q. 1995, 17, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Deak, T.; Meriwether, J.L.; Fleshner, M.; Spencer, R.L.; Abouhamze, A.; Moldawer, L.L.; Grahn, R.E.; Watkins, L.R.; Maier, S.F. Evidence that brief stress may induce the acute phase response in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1997, 273, R1998–R2004. [Google Scholar] [CrossRef]
- Hicks, T.A.; McGlone, J.J.; Whisnant, C.S.; Kattesh, H.G.; Norman, R.L. Behavioral, endocrine, immune, and performance measures for pigs exposed to acute stress. J. Anim. Sci. 1998, 76, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Arthington, J.D.; Eichert, S.D.; Kunkle, W.E.; Martin, F.G. Effect of transportation and commingling on the acute-phase protein response, growth, and feed intake of newly weaned beef calves. J. Anim. Sci. 2003, 81, 1120–1125. [Google Scholar] [CrossRef]
- Hickey, M.C.; Drennan, M.; Earley, B. The effect of abrupt weaning of suckler calves on the plasma concentrations of cortisol, catecholamines, leukocytes, acute-phase proteins and in vitro interferon-gamma production. J. Anim. Sci. 2003, 81, 2847–2855. [Google Scholar] [CrossRef]
- Zulkifli, I.; Najafi, P.; Nurfarahin, A.J.; Soleimani, A.F.; Kumari, S.; Aryani, A.A.; O’Reilly, E.L.; Eckersall, P.D. Acute phase proteins, interleukin 6, and heat shock protein 70 in broiler chickens administered with corticosterone. Poult. Sci. 2014, 93, 3112–3118. [Google Scholar] [CrossRef] [Green Version]
- Najafi, P.; Zulkifli, I.; Soleimani, A.F.; Goh, Y.M. Acute phase proteins response to feed deprivation in broiler chickens. Poult. Sci. 2016, 95, 760–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murata, H.; Miyamoto, T. Bovine haptoglobin as a possible immunomodulator in the sera of transported calves. Br. Vet. J. 1993, 149, 277–283. [Google Scholar] [CrossRef]
- Piccione, G.; Casella, S.; Giannetto, C.; Giudice, E.; Fazio, F. Utility of acute phase proteins as biomarkers of transport stress in ewes. Small Rumin. Res. 2012, 107, 167–171. [Google Scholar] [CrossRef]
- Fazio, F.; Ferrantelli, V.; Cicero, A.; Casella, S.; Piccione, G. Utility of acute phase proteins as biomarkers of transport stress in ewes and beef cattle. Ital. J. Food Saf. 2015, 4, 4210. [Google Scholar] [CrossRef]
- Warriss, P.D. The handling of cattle pre-slaughter and its effects on carcass and meat quality. Appl. Anim. Behav. Sci. 1990, 28, 171–186. [Google Scholar] [CrossRef]
- Knowles, T.G. A review of the road transport of cattle. Vet. Rec. 1999, 144, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Tarrant, P.V.; Kenny, F.J.; Harrington, D.; Murphy, M. Long distance transportation of steers to slaughter: Effect of stocking density on physiology, behaviour and carcass quality. Livest. Prod. Sci. 1992, 30, 223–238. [Google Scholar] [CrossRef]
- Schaefer, A.L.; Jones, S.D.; Stanley, R.W. The use of electrolyte solutions for reducing transport stress. J. Anim. Sci. 1997, 75, 258–265. [Google Scholar] [CrossRef]
- Knowles, T.G.; Brown, S.N.; Warriss, P.D.; Phillips, A.J.; Dolan, S.K.; Hunt, P.; Ford, J.E.; Edwards, J.E.; Watkins, P.E. Effects on sheep of transport by road for up to 24 hours. Vet. Rec. 1995, 136, 431–438. [Google Scholar] [CrossRef]
- Blecha, F.; Boyles, S.L.; Riley, J.G. Shipping suppresses lymphocyte blastogenic responses in Angus and Brahman× Angus feeder calves. J. Anim. Sci. 1984, 59, 576–583. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, P.J. Investigation of the effects of transport and lairage on hydration state and resting behaviour of calves for export. Vet. Rec. 1992, 130, 413–416. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, T.; Schaefer, A.L.; Tong, A.; Stanley, R.; Jones, S.; Robertson, W.; Dyck, R. The effects of transportation on carcass yield, meat qualityb and hematology values in electrolyte treated cattle. In Proceedings of the 39th International Congress Meat Science Technology, Calgary, AB, Canada, 1–6 August 1993; pp. 1–6. [Google Scholar]
- Jackson, P.G.G.; Cockcroft, P.D. Clinical Examination of Farm Animals, 1st ed.; Blackwell Science: Oxford, UK, 2002. [Google Scholar]
- Earley, B.; O’Riordan, E.G. Effects of transporting bulls at different space allowances on physiological, haematological and immunological responses to a 12-h journey by road. Ir. J. Agric. Food Res. 2006, 45, 39–50. [Google Scholar]
- Buckham Sporer, K.R.; Burton, J.L.; Earley, B.; Crowe, M.A. Transportation stress in young bulls alters expression of neutrophil genes important for the regulation of apoptosis, tissue remodeling, margination, and anti-bacterial function. Vet. Immunol. Immunopathol. 2007, 118, 19–29. [Google Scholar] [CrossRef]
- Pettiford, S.G.; Ferguson, D.M.; Lea, J.M.; Lee, C.; Paull, D.R.; Reed, M.T.; Hinch, G.N.; Fisher, A.D. Effect of loading practices and 6-hour road transport on the physiological responses of yearling cattle. Aust. J. Exp. Agric. 2008, 48, 1028–1033. [Google Scholar] [CrossRef]
- Ottaway, C.A.; Husband, A.J. The influence of neuroendocrine pathways on lymphocyte migration. Immunol. Today 1994, 15, 511–517. [Google Scholar] [CrossRef]
- Brenner, I.; Shek, P.N.; Zamecnik, J.; Shephard, R.J. Stress hormones and the immunological responses to heat and exercise. Int. J. Sports Med. 1998, 19, 130–143. [Google Scholar] [CrossRef]
- Fisher, D.D.; Wilson, L.L.; Scholz, R.W. Environmental and genetic effects on hematologic characteristics of beef cows. Am. J. Vet. Res 1980, 41, 1533–1536. [Google Scholar]
- Etim, N.N.; Oguike, M.A. Environmental and management stressors: Implications for reproductive and productive performances of farm animals in the tropics. J. Agric. Sustain. 2014, 5, 153–170. [Google Scholar]
- Payne, J.M.; Payne, S. The Metabolic Profile Test; Oxford University Press: Oxford, UK, 1987; pp. 1–179. [Google Scholar]
- Warriss, P.D.; Brown, S.N.; Knowles, T.G.; Kestin, S.C.; Edwards, J.E.; Dolan, S.K.; Phillips, A.J. Effects on cattle of transport by road for up to 15 hours. Vet. Rec. 1995, 136, 319–323. [Google Scholar] [CrossRef]
- Tadich, N.; Gallo, C.; Bustamante, H.; Schwerter, M.; Van Schaik, G. Effects of transport and lairage time on some blood constituents of Friesian-cross steers in Chile. Livest. Prod. Sci. 2005, 93, 223–233. [Google Scholar] [CrossRef]
- Manjoo, M.; Burger, F.J.; Kielblock, A.J. A relationship between heat load and plasma enzyme concentration. J. Therm. Biol. 1985, 10, 221–225. [Google Scholar] [CrossRef]
- Kannan, G.; Kouakou, B.; Terrill, T.H.; Gelaye, S. Endocrine, blood metabolite, and meat quality changes in goats as influenced by short-term, preslaughter stress. J. Anim. Sci. 2003, 81, 1499–1507. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.B. New approaches to identifying and measuring pain. In Proceedings of the Australian Animal Welfare Strategy Science Summer on Pain and Pain Management, Melbourne Convention Centre, Victoria, Australia, 18 May 2007. [Google Scholar]
- Bergamasco, L.; Coetzee, J.F.; Gehring, R.; Murray, L.; Song, T.; Mosher, R.A. Effect of intravenous sodium salicylate administration prior to castration on plasma cortisol and electroencephalography parameters in calves. J. Vet. Pharmacol. Ther. 2011, 34, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.-H.; Lee, J.-T. Stress and EEG. Convergence and Hybrid Information Technologies. 2010. Available online: https://www.intechopen.com/books/convergence-and-hybrid-information-technologies/stress-and-eeg/ (accessed on 5 February 2018).
- Kaka, U.; Hui Cheng, C.; Meng, G.Y.; Fakurazi, S.; Kaka, A.; Behan, A.A.; Ebrahimi, M. Electroencephalographic changes associated with antinociceptive actions of lidocaine, ketamine, meloxicam, and morphine administration in minimally anaesthetized dogs. Biomed. Res. Int. 2015, 2015, 305367. [Google Scholar] [CrossRef] [PubMed]
- Kaka, U.; Goh, Y.M.; Chean, L.W.; Chen, H.C. Electroencephalographic changes associated with non-invasive nociceptive stimulus in minimally anaesthetised dogs. Pol. J. Vet. Sci. 2016, 19, 675–683. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Prior to Sea Transport | After Sea Transport | After Road Transport | After 4 d Recovery | After 7 d Recovery | p-Value |
|---|---|---|---|---|---|---|
| ALT(U/L) | 35.00 c ± 2.47 | 40.29 b ± 1.34 | 28.65 c ± 1.11 | 48.90 a ± 2.09 | 42.06 b ± 1.57 | <0.0001 |
| ALP(U/L) | 138.37 b ± 2.25 | 173.30 ab ± 11.29 | 175.70 a ± 11.24 | 139.65 b ± 10.89 | 155.62b a ± 13.31 | 0.0219 |
| AST(U/L) | 42.00 d ± 1.10 | 110.98 bc ± 3.23 | 103.36 c ± 2.92 | 159.78 a ± 4.94 | 119.18 b ± 6.41 | <0.0001 |
| CK(U/L) | 155.00 c ± 5.94 | 200.45 b ± 22.96 | 257.40 a ± 4.68 | 160.05 c ± 3.75 | 162.31 c ± 10.03 | <0.0001 |
| LDH(U/L) | 2137.30 a ± 81.60 | 1914.10 b ± 76.67 | 2151.35 a ± 20.42 | 2272.55 a ± 98.14 | 2210.31 a ± 83.18 | 0.0150 |
| Parameter | After Sea Transport | After Road Transport | 4 Days Recovery | 7 Days Recovery | p-Value |
|---|---|---|---|---|---|
| WBC (×109 /L) | 6.09 c ± 0.37 | 11.06 a ± 0.51 | 6.58 c ± 0.27 | 9.08 b ± 0.44 | <0.0001 |
| PCV (%) | 39.90 ± 0.70 | 40.58 ± 0.80 | 40.25 ± 0.66 | 39.20 ± 0.50 | 0.5212 |
| RBC (×1012 /L) | 9.53 bc ± 0.32 | 9.24 c ± 0.12 | 10.11b a ± 0.20 | 10.26 a ± 0.22 | <0.0001 |
| LYM (×109 /L) | 5.52 ± 0.24 | 5.08 ± 0.28 | 5.69 ± 0.21 | 5.44 ± 0.23 | 0.3391 |
| HGB (g/L) | 135.55 a ± 2.56 | 124.75 b ± 1.17 | 137.7 a ± 2.35 | 137.85 a ± 2.90 | <0.0001 |
| HCT (L/L) | 0.38 a ± 0.05 | 0.35 b ± 0.04 | 0.39 a ± 0.06 | 0.38 a ± 0.08 | <0.0001 |
| PLT (×109 /L) | 333.50 a ± 22.15 | 308.87 a ± 35.25 | 357.00 a ± 26.95 | 371.85 a ± 29.71 | 0.4400 |
| NEU (×109 /L) | 3.11 b ± 0.16 | 4.15 a ± 0.34 | 3.24 b ± 0.14 | 3.38 b ± 0.17 | <0.0001 |
| N:L ratio | 0.56 b ± 0.07 | 0.82 a ± 0.12 | 0.57 b ± 0.05 | 0.62 b ± 0.07 | 0.0141 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zulkifli, I.; Abubakar, A.A.; Sazili, A.Q.; Goh, Y.M.; Imlan, J.C.; Kaka, U.; Sabow, A.B.; Awad, E.A.; Othman, A.H.; Raghazali, R.; et al. The Effects of Sea and Road Transport on Physiological and Electroencephalographic Responses in Brahman Crossbred Heifers. Animals 2019, 9, 199. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9050199
Zulkifli I, Abubakar AA, Sazili AQ, Goh YM, Imlan JC, Kaka U, Sabow AB, Awad EA, Othman AH, Raghazali R, et al. The Effects of Sea and Road Transport on Physiological and Electroencephalographic Responses in Brahman Crossbred Heifers. Animals. 2019; 9(5):199. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9050199
Chicago/Turabian StyleZulkifli, Idrus, Ahmed A. Abubakar, Awis Q. Sazili, Yong M. Goh, Jurhamid C. Imlan, Ubedullah Kaka, Azad B. Sabow, Elmutaz A. Awad, Azalea H. Othman, Razlina Raghazali, and et al. 2019. "The Effects of Sea and Road Transport on Physiological and Electroencephalographic Responses in Brahman Crossbred Heifers" Animals 9, no. 5: 199. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9050199