Reaction Time Improvements by Neural Bistability
by
 , ,
, ,
Henk Koppelaar
1,* ,
,
Parastou Kordestani Moghadam
2,
Kamruzzaman Khan
3,
Sareh Kouhkani
4,
Gijs Segers
5 and
Martin van Warmerdam
6 1
Faculty of Electrical Engineering, Mathematics and Computer Science, University of Delft, 2628 XE Delft, The Netherlands
2
Social Determinants of Health Research Center, Lorestan University of Medical Sciences, Korramabad 6813833946, Iran
3
Department of Mathematics, Pabna University of Science and Technology, Pabna 6600, Bangladesh
4
Department of Mathematics, Islamic Azad University Branch of Shabestar, Shabestar 5381637181, Iran
5
Sports Trainer, Physical Education Teacher, Visual Performance Trainer, Gymi Sports, 4907 BC Oosterhout, The Netherlands
6
Van Warmerdam Optometry’s, 5234 KA Hertogenbosch, The Netherlands
*
Author to whom correspondence should be addressed.
Behav. Sci. 2019, 9(3), 28; https://0-doi-org.brum.beds.ac.uk/10.3390/bs9030028
Submission received: 9 December 2018
/
Revised: 22 January 2019
/
Accepted: 12 March 2019
/
Published: 18 March 2019
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The often reported reduction of Reaction Time (RT) by Vision Training) is successfully replicated by 81 athletes across sports. This enabled us to achieve a mean reduction of RTs for athletes eye-hand coordination of more than 10%, with high statistical significance. We explain how such an observed effect of Sensorimotor systems’ plasticity causing reduced RT can last in practice for multiple days and even weeks in subjects, via a proof of principle. Its mathematical neural model can be forced outside a previous stable (but long) RT into a state leading to reduced eye-hand coordination RT, which is, again, in a stable neural state.
1. Introduction
Exercise physiology and psychology, biochemistry, and biomechanics in sports have caused long lasting performance improvements in sports. However, the science of motor control in sport practices has not come to full maturity until today even though it was proposed in the IOC Congress on Sport Sciences [1] as early as 1996. This paper explains sensorimotor Reaction Time (RT) improvements of more than 10% of fully trained athletes even at the top of their abilities. The explanation purports the ability of the human neural system to become stabilized in more efficient control of the motor system, simply by vision training.
The societal impact of this research is related by Burris et al. [2], by way of Ted Williams’ observation “I think, without question, the hardest single thing to do in sports is to hit a baseball.” Burris and colleagues [2] explain “Major League Baseball (MLB) pitches move at speeds near the processing limits of the vestibular-ocular tracking, which leaves the batter with mere milliseconds to decipher the pitch, project its trajectory, decide to swing, and coordinate the timing and trajectory of a 2.25-inch diameter bat. The immense difficulty of this task is underscored by the fact that players who hit successfully on less than a third of their at-bats can receive one-hundred million dollar contracts in today’s free-agent market.”
Room for improvement of RTs is from Rathelot et al. [3]. They proved that neural fields competing in visual perception versus dexterous command may converge into a single command mode, which will induce extremely short reaction times, which are shorter than commonly achieved. Their finding is clearly task dependent [4,5,6,7] and sustained by Weiler et al. [8]. We exploit by proof of principle in Section 2 and Section 4 (via a mathematical FitzHugh-Nagumo model of the sensorimotor system, for short FHN) that a reduction of RTs is stable in a new states of mind outside their previously obtained RT, which was a comfort behavioral zone of athletes. The model explains how improved eye-hand reaction times after a tailored “More Input Eye vision is the Key” (MIEK) training occur using LCD shutter glasses, see Section 3. The reduced RTs observed after the training are of high statistical significance relative to the RTs before the training, as reported in Section 3 and Section 4.4.
Athletes, like all humans, miss over 9% of their visual inputs because of spontaneous eyeblinks. In the tailored MIEK Visual Performance training, we enforce eyeblinks up to much higher rates using LCD shutter glasses (operating similar to eye-blinks and called ‘strobospectacles’). The strobospectacles train athletes’ eyes to intermittently enforce taking a break, reducing visual input, and, thereby, decreasing redundant feedback [9]. The reduced RT by strobotraining has been found to last beyond the training period but is, currently, largely unexplained. This novel explanation is the goal of this paper. We concentrate on the motor control by neural fields. The sensor part of the human system deserves further scrutiny, because speeding up RTs has also been found by invisible stimuli.
Strobotraining naturally speeds-up RTs by involving the phenomenon of invisible stimuli [10] and by establishing stable memory effects [9,11,12,13,14,15,16]. We conjecture that strobotraining as a side effect invokes the human ability speeding up RTs by invisible stimuli, because of the fast shutting-off light. For example, experienced ball athletes do not follow the ball’s trajectory, but rather observe the initial movement of the ball, remember it, and, with a rapid eye saccade, go to a projected final position of the ball’s path. Memory effects are perceivable via EEG patterns [17,18], which are emanating from oscillating neural fields. Figure 1 depicts how to move a sensorimotor FHN 1 ability to a new field of improved ability FHN 2. The neural oscillations were correctly hypothesized by some authors as memory effects [9,13,14,16]. For more extensive information about FHN oscillations, see References [19].
By hypothesizing an idea complimentary to Schmidt’s team [20] for cognitive learning, we show that strobotraining results in a speed of motor response moves from a habitual zone in athletes’ sensorimotor system, i.e., by physical training achieved shortest RT, to a new stable neural state with about 10% faster RT than the old achieved habitual one. Note that the issue here is: without physical muscle training, a sensorimotor system change occurs. This is comprehensively reviewed by Ahissar [21]: perceptual training is aimed at modifying sensorimotor abilities. His report stems from auditorily perceptional prior work [22], followed up by Ho and coworkers [23], who also show oscillating neural correlates, which are different for left and right sides of the body. Later research applies the modelling to developing brains of adolescents [24].
The main proof by principle in this paper, by using a mathematical FitzHugh-Nagumo model, is that such old and new stabilities, in short ‘bistabilities,’ occur through two interacting models: from a habitual zone, via a meta-stable intermediate state, to a new stable zone. This type of modelling for research is common in physics and is reviewed for broader application by Kriegeskorte et al. [25].
The findings enforce Güllich’s research among top talents in sports [26] who revealed that world champions do train in many disciplines. As such, they learn more than one neural comfort zone.
2. Sensorimotor Tasks Induce Reaction Delays
Rathelot, Dum, and Strick [3] discovered that the posterior parietal cortex contains a common command apparatus for hand movements. If employed, this command reduces RTs to an optimum. Of course, the central nervous system has many sources of command to the motor apparatus of the human body. Such plasticity [3] of the sensorimotor system has been studied for the aural system [21,22] and has a central role for the visual system. The brains’ plasticity is extreme, even noise in neurons is sensible to signals [27]. Because the many sources of command [28,29] researchers advocate a dual task paradigm: two separate neural fields with one for the oculomotor system and one for the motor behavior of the limbs. Haak et al. [30] defend three systems, which are simulated by Ambrosio et al. [31]. Three systems may occur if proprioscepsis is considered. For instance, Ebsch and Rosenbaum [32] show that cortical circuits can respond to natural and artificial stimuli in the context of locally gained imbalance, which is this local behavior (i.e., the neural field) that is relocatable in the brain and, hence, can be untrained.
To make our analysis more tractable, we simplify the neural fields in the sensorimotor system, according to Graham [12,33], which is commonly done in neural modelling. We split it in an oculomotor system and a motor part, such that, if the first is sufficiently stimulated, then the subsequent motor response starts [32]. These two systems can alter their modus operandi to operate independently, or dependently but separately [6], or even under a common command [3]. Examples of various stabilities are: hand or foot movement does not change much if subjects either see or do not see their hand or foot, so this suggests independence of the two effector systems. Reaching for a glass of water requires executing hand-eye movements in fine temporal coupling between the two effectors. In sum, a large repertoire of eye-hand movements in our daily lives might use such different ‘circuitries’, namely neural fields. For instance, in smooth pursuit tasks, such as throttling, pilots in the mean have slightly shorter delays [34] than ball sports athletes, which corresponds to Reference [3].
Plasticity brings us from one state to the other. Jana et al. [4,6] hypothesized an executive controller assessing behavioral context, which allows switching between modes or merging of modes. To paraphrase Schmidt et al. [35] in response to a transient input from strobospectacles, the sensorimotor system relaxes to a single new state. Cooperation between different neural fields is a necessary reduction of computational complexity for large networks, according to Kaminski [36] and Stefanescu and Jirsa [37]. From our model (see Appendix A, Equation (A1)), propagating waves in detail are elicitable. Figure 1 shows a gradually spreading traveling wave [38] from FHN 1 reaching another field FHN 2. This enables mapping cortical wave patterns to the original point of FHN spikes trains. Such research has been conducted for smooth pursuit tasks [39].
Wave speed such as propagation between neural fields has been studied by Hafed et al. [40]. The strobotraining experiments, carried out in our analysis (Section 3), enforces athletes an uncommon way of processing visual input, i.e., enforced blinking. Previous research has proved the neural impact of blinking [41,42,43]. Thereafter, stable oscillations of the newly achieved FHN 2 state exist without continued input.
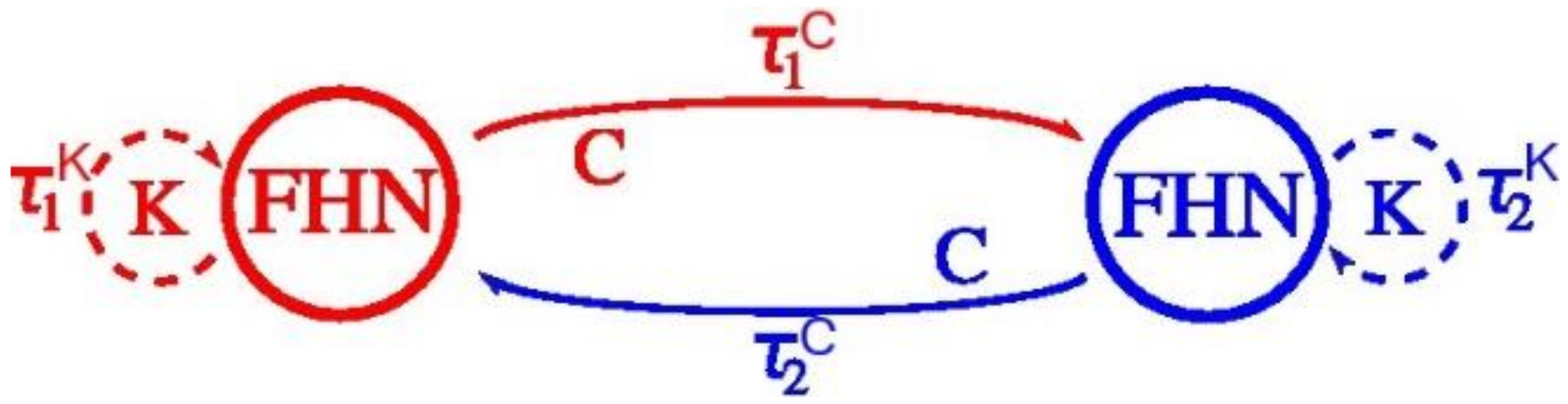
The principle we seek out to prove is that: speed-up of the sensorimotor system may happen because at least two stable states for the two effectors exist including each with different time delays, as depicted in Figure 2. The time delay of the coupling C between the two groups of sensorimotor neurons FHN1 and FHN2 is . Internal feedback K within the sensorimotor neurons group is . The simplified model neglects the internal feedback K to an apparatus of the body (enclosed by the dotted line) because of the complexity of the visual effector. Why to exclude? Because it includes feedback from one group of sensorimotor neurons to the eyes. For example, feedback to the eyes to express saccades after the hand moves a ball [44].
3. Materials and Methods
Basic comparison material was established using measurements from training elsewhere in the world. All publications on strobotraining were collected. If such training is done adequately, assessments [2,46] show that improvements may occur in nine tested abilities of students and stamina of the visual system. Positive results also appeared for toss-to-wall [46], baseball [2,13,47,48], basketball [49,50], cricket, football [9,51,52], fencing, frisbee [9], hockey [15,53], rugby [54], softball [55,56], tennis, table tennis [57], volleyball [49], and water-polo [49]. Outside of sports, improvements are known for motion sickness [58], older adults [59], and visually impaired youth [60]. The opposite of enforcing eye-blinks, i.e., stroboscopic lightning, prevented visual sharpening in fish [61]. A full overview including RT measurement equipment up to 2007, is from Erickson [62] and is extended later by Ellison [53] in his dissertation. There are differences in results between measurement techniques via virtual or real ball grasping [63].
To prove stability of shorter RTs, we use a bistable Fitzhugh-Nagumo model for the eye-hand coordination. The main idea of this explanation is the plasticity of the neural system, whose properties enable the un-training of old and learning novel behaviors. History of sports shows the relentless shift to shorter RTs. Otherwise, records would not be broken. Most of RT improvements comes from eye-hand coordination, i.e., the sensorimotor system. The RT improvement is achieved by shifting the control of the motor system to a new -albeit stable- neural field (otherwise, the RT would not be shorter and the new and faster behavior could not be repeated). The ultimate speed is achieved if the shift of the neural control is made into the cerebellum [3]. The learning example is a novel behavior that starts in a neural field in the right hemisphere. After training, it moves to the left hemisphere and, after full mastery, it resides in the cerebellum. Un-training a habit or behavior neurologically is mostly the same as training, i.e., shifting a neural field. An example of un-training (in perception) is: ignoring task irrelevant stimuli [64]; or, un-training the checkup of ambiguous features [65]. Both examples are replicated by our MIEK Visual Performance Training and reduce RTs of the sensorimotor system as is displayed clearly by the green versus red lines in Figures 4–6 below.
3.1. Participants of the Experiment
The current study was conducted with 81 athletes, men and women aged 19–40, from eight sport areas: the (Dutch) National Hockey League, Tennis, Squash, Taekwondo, Fencing, Snowboarding, and the top Umpires of two hockey leagues (KNHB and FIH). Testing and training were completed at the teams’ competition sites between 2014 and 2019. One of the authors (GS) and the team’s visual performance trainer conducted all trainings. Participation was voluntary, and no benefits or penalties were offered to encourage participation. The potential effects of the training were explained to participants. All participants had informed consent to the training.
3.2. Measurements
All athletes participating in the experiment underwent baseline measurements. Baseline measurements were conducted by authors MW and by GS. Followed MIEK Visual Performance Training once per week over the course of 12 weeks, which was conducted by GS. Each training totaled about 90 min, of which 30 min was completed with the stroboglasses. Lastly, the athletes underwent posterior measurements to determine progress or deterioration relative to the baseline measurement, which was also conducted by MW and by GS.
3.3. Computational Equipment
The FitzhughNagumo model was run on a Quadcore ASUS laptop, using Maple software, version 18 from MapleSoft, Waterloo, ON, Canada.
4. Results
4.1. Models with at Least Two Stable States Depict Learning
Graham [33] reviews the success of the simple decision rule model, which accounted for so many experimental results and represented most areas of visual cortex and the rest of the brain. In response to near threshold patterns, only a small proportion of neural neurons (‘analyzers’ in Reference [33]) are being stimulated above their threshold, as built in Equations (1). Retention or memory of an improved reaction time (RT) in this mathematical model is proved by its second stable state, which is different from its first stable state, i.e., its ‘habitual zone’. It started in 1961 (FitzHugh [67]) and in 1962. When Nagumo et al. [68] proposed a model for emulating the current signal observed in a living organism’s excitable cells. This is coined by the name ‘FitzHugh-Nagumo’ (FHN) model of mathematical neuroscience and is a simpler version of the Hodgkin-Huxley (HH) model of spiking neurons. Graham’s [33] hypothesized decision variable (DV) models the decision to move after the signal from a sensor system. The DV is an abstract construct that has very successfully predicted RTs distribution [69]. An independent source of insight for this is are experiments to study neural adaptation in conceptual processing [70]. Downstream activation from the perception to action in cognition even shows the RT assessment and even the decline of training effects we discuss here. Therefore, the DV mediates between eyes and hand, i.e., two mean fields of neurons. This leads, in our model, to a bifurcation of states or, said otherwise, the training brings the model to a critical value at which a system is unstable and, by further training, state variables bifurcate become stable again. The DV then starts the motor system to move earlier than before, in a previous state. Subsequent training then stabilizes this newly attained achievement as depicted by green lines in Figures 4–6 below.
4.2. Complimentary Work
Bistability is a well-known phenomenon such as the sleep-wake rhythm. This is possible because the brain has both stable states, but we cannot be in both states simultaneously. Therefore, it is bi-stable. Bistable perception is also well-known [71,72,73,74,75]. Exogenous measures such as disturbing sleep or control can bring or force a neural system from one to another stable state and back for some period [20,72,74,75]. Input from perception gradually build-up transitions of motor populations, until the competition between alternative representations is resolved by a neural threshold mechanism. Bistable neurons reflect neural collectives rather than properties of individual populations or neurons. Such bistability may occur even at a single neuron level [76] and at the level of lattices and fields. The single neuron approach to modelling complies with multiple neuron models, as explained by Graham [12,33]. We complement the pioneering work done by Schmidt et al. [20] to explain learning cognitive tasks by employing the plasticity of the brain to model shifting neural fields. A curiosity of difference between Schmidt et al. [20] and our work is the difference between the employed mathematical models. Prior work by Montbrio and Roxin [77] developed models to bridge the gap between large networks of spiking neurons and their observed firing rate. They need observations from EEGs to gauge cognitive learning tasks, which we do not pursue. They introduced a second order differential equation. With this, we depart from the FHN model [78], as inspired by Hodgkin and Huxley [79] with a higher order nonlinearity. The difference between the approaches is reminiscent of statistical physics and the ensemble theory for many neurons by Schmidt et al. [20] versus our FHN full causal differential equations (see also Rankin and Chavane) [80]. Our choice is supported by Chen and Majda [81] who invoke dimension reduction techniques for FHN ensemble models.
4.3. The FitzHugh-Nagumo (FHN) Model
The neural system with subscript 1 models, the visual part of the sensorimotor system, and subscript 2 models the motor part. The RT emerges because of a delay between the two neurons. Observations over 81 athletes (see below Figures 4–6 of the experimental results) showed that the RT is reduced by 10.8%. This means it is shifted because of the training in the mean from 0.69 → 0.61 s of RTs for the athletes’ sensorimotor system.
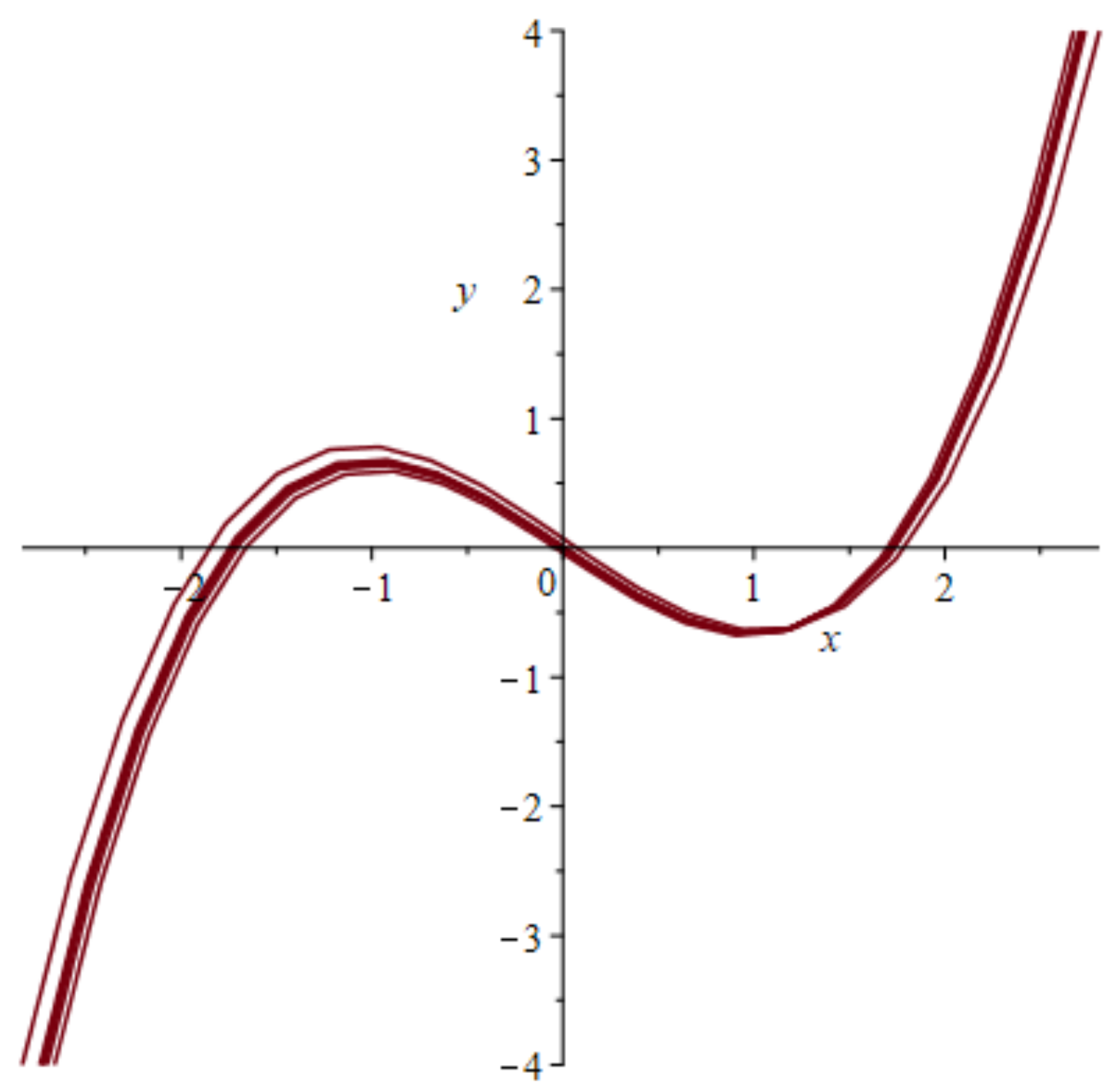
Equation (1) has three stable points, as seen from left to right in the phase plot of Figure 3, at the zero crossings of the curved line. The system is started from a stable state at the origin, moves to the first unstable state at the first crossing to the right, and is pushed further by excitation to the second stable state at the point x = 1.8, which is again a stable state.
The two-effector model can evoke a speed-up in RT if it bifurcates to a novel stable state. Evidence for such a scenario is independently gained from various sources [37,84,85] by studying neural fields. We take two neurons 1 and 2 along with the generic two-variable reaction–diffusion model for excitable and inhibitory media—the FritzHugh-Nagumo model (1)—such that the excitory field 1 is modeled with a membrane potential at time . The electric membrane property is captured by and , which is the potential of the membrane at rest at the starting time zero of excitation (see Kaminski et al. [36]).
Memory effects of such novel obtained bi-stable states last for days and weeks (variable among subjects) and are characterized by time-scales of magnitude larger than those of perceptual reversals [72,74]. We study sensorimotor dynamics through the lens of a mathematical model. The new stable state is shown in Figure 2. An analog electrical model simulating the FHN equations gave the same stability result [86], based upon FitzHugh’s prior model [87].
The delay time is in the brain, which is not necessarily symmetrical (forward versus feedback). We make it symmetrical [78] because we only need proof of two different feedforward delays including one measurement prior to and one after the MIEK Visual Performance Training. Moreover, Panchuk et al. [88] give proof to convert an FHN system with asymmetric delays into a system with one delay.
This plot of Figure 3 in the phase plane shows a nullcline of a single FHN system passing three stability points at the horizontal axis including the zero points of the curve, which are the stable states with the middle one in an intermediate meta-stable state. The phase lines of the system are omitted. In a full phase plot of hundreds of behaviors, the stable points would be difficult to detect. In spite of the variability of neurons’ spiking behavior, Panchuk et al. [45] discover an invariant to detect coherent spiking of the two FHN systems if the number of involved neurons is small. Otherwise, coherent solutions cannot be realized due to the refractory phases of the subsystems.
4.4. Results of the Experiments
5. Conclusions
Bi-stability is shown to be feasible in the applied FHN model, which also shows the retention of the obtained changes.
The exact shape of fields of cooperating neurons is variable and not fully classified. Researchers [89,90,91] defend ring-like architectures of cooperating FHN neurons. Neural field propagation happens in various ways, such as a non-ring traveling wave (see Figure 1). We do not follow [90,91] because they do not include a propagation delay.
The explanation of reduced RTs by strobotraining might need a complimentary explanation. As is well known, dexterous behavior depends upon the context of the task at hand. It might well be that the stroboscopic training forces athletes out of their habitual or comfort zone by perceived similarity or reminiscence of the training with known and familiar tasks. This phenomenon is discussed by Güllich [26]. He discovered that training of only one stable state (comfort zone) is less productive to achieve the world top than to train many sports, or in our model terms, to train many stable states.
What still has to be done is understanding the context of multiple tasks (sports) and its inputs. The quenching of neural variability was discovered by Churchland [92] and explained by Chang [93] in terms of reverse engineering, and the optimal dose of input to maximize the effect/output. Rathelot et al. [3] paved the way for RTs reduction to gain in the future. This scenario is supported by Reference [94] and by reversed application of Plotnikov’s [95] adaptive control without prior knowledge of parameters, or search of independent FHN model coefficients [96] and by estimating these very accurately for the models [97].
A second issue for further research is detailing the successive subprocesses with their neural fields. For instance, Chang and Jazayeri [98] introduced a multi-stimuli concept to explain time to contact a ball, that is the RT between seeing and catching a ball. Because the observed variability [99] of a 200 msec increase in RTs (depending on eye-blinks) has detailed processes accumulating sensorimotor RTs and practical application for research in driver behavior and sports. To gauge FHN models, Safaari et al. [100] modelled cortical state dynamics and single trial sensory processing.
Author Contributions
Conceptualization, H.K. Data curation, M.v.W. Formal analysis, H.K. and K.K. Investigation, H.K., P.K.M., G.S., and M.v.W. Methodology, H.K. and G.S. Project administration, H.K. and G.S. Supervision, H.K. and G.S. Validation, S.K. Writing—original draft, H.K., P.K.M., and K.K. Writing—review & editing, H.K., P.K.M., K.K., and S.K.
Funding
The material cost for this research was funded by Gymi Sports from 2016 – 2018. The APC was also funded by Gymi Sports.
Acknowledgments
We thank B. Bakker (Amphia Hospital, Breda, the Netherlands), R. van den Brule (MSc, Wageningen University, the Netherlands), C. Jenniskens (Stolmed Eye Clinics, Bergen op Zoom, the Netherlands), and R.H.E.M. Koppelaar (Imperial College, London) for their support.
Ethics Approval
The Ethics Board of the Lorestan University of Medical Sciences approves this research. The Ethics code granted to this research is: IR.LUMS.REC.1397.202.
Conflicts of Interest
The first four authors declare that no competing interests exist. The last two authors declare a possible interest in bringing the results to commercial profit in their respective companies.
Appendix A
The FHN models spiking neurons, with precise similarity of the pulses. Its analytical expressions obtained via the Lienard transform are by families of functions in hyperbolic geometry. The classes appear because of the freedom of parameters in solutions, e.g., and are independent constants -arbitrarily to be chosen- in . This mimics rather than spikes train curves. Existing families in the FHN models are not yet explored. The number of results might be wieldy, as discovered in other types of equations [103,104,105]. We restrict ourselves to a single plot of a traveling wave in Figure 1, obtained from the system of FHN Equations (1).
To accommodate travelling waves, we extend the FHN systems (1) with a 1D spatial component [90,106], without loss of generality, along the spatial axis s.
This generates the spatial behavior of neural signals, as depicted in Figure 1.
References
- Abernethy, B. Training the Visual-Perceptual Skills of Athletes: Insights From the Study of Motor Expertise. Am. J. Sports Med. 1996, 24, 89–92. [Google Scholar] [CrossRef]
- Burris, K.; Vittetoe, K.; Ramger, B.; Suresh, S.; Tokdar, S.T.; Reiter, J.P.; Appelbaum, L.G. Sensorimotor abilities predict on-field performance in professional baseball. Sci. Rep. 2018, 8, 1–9. [Google Scholar]
- Rathelot, J.; Dum, R.P.; Strick, P.L. Posterior parietal cortex contains a command apparatus for hand movements. Proc. Natl. Acad. Sci. USA 2017, 114, 4255–4260. [Google Scholar] [CrossRef]
- Scott, S.H. The computational and neural basis of voluntary motor control and planning. Trends Cogn. Sci. 2012, 16, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Jana, S.; Gopal, A.; Murthy, A. A Computational Framework for Understanding Eye-Hand Coordination. J. Indian Inst. Sci. 2017, 97, 543–554. [Google Scholar] [CrossRef]
- Jana, S.; Gopal, A.; Murthy, A. Evidence of common and separate eye and hand accumulators underlying flexible eye-hand coordination. J. Neurophysiol. 2017, 117, 348–364. [Google Scholar] [CrossRef] [PubMed]
- Gopal, A.; Jana, S.; Murthy, A. Contrasting speed accuracy trade-offs for eye and hand movements reveal the optimal nature of saccade kinematics. J. Neurophysiol. 2017, 118, 1664–1667. [Google Scholar] [CrossRef] [PubMed]
- Weiler, J.; Gribble, P.L.; Pruszynski, J.A. Spinal stretch reflexes support efficient hand control. Nat. Neurosci. 2019, 1–11. [Google Scholar] [CrossRef]
- Appelbaum, L.G.; Schroeder, J.E.; Cain, M.S.; Mitroff, S.R. Improved visual cognition through stroboscopic training. Front. Psychol. 2011, 2, 1–13. [Google Scholar] [CrossRef]
- Savazzi, S.; Marzi, C.A. Speeding Up Reaction Time with Invisible Stimuli. Curr. Biol. 2002, 12, 403–407. [Google Scholar] [PubMed]
- Curtis, C.E.; Connolly, J.D. Saccade Preparation Signals in the Human Frontal and Parietal Cortices. J. Neurophysiol. 2007, 99, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Britten, K.H.; Shadlen, M.N.; Newsome, W.T.; Movshon, J.A. The Analysis of Visual Motion: A Comparison of Neuronal and Psychophysical Performance. J. Neurosci. 1992, 12, 4745–4765. [Google Scholar] [CrossRef]
- Smith, T.Q.; Mitroff, S.R. Stroboscopic Training Enhances Anticipatory Timing. Int. J. Exerc. Sci. 2012, 5, 344–353. [Google Scholar] [PubMed]
- Appelbaum, L.G.; Cain, M.S.; Schroeder, J.E.; Darling, E.F.; Mitroff, S.R. Stroboscopic visual training improves information encoding in short-term memory. Atten. Percept. Psychophys. 2012, 74, 1681–1691. [Google Scholar] [CrossRef]
- Mitroff, S.R.; Friesen, P.; Bennett, D.; Yoo, H.; Reichow, A.W. Enhancing Ice Hockey Skills Through Stroboscopic Visual Training: A Pilot Study. Athl. Train. Sport. Health Care 2013, 5, 261–264. [Google Scholar] [CrossRef]
- Smith, P.L.; Lilburn, S.D.; Corbett, E.A.; Sewell, D.K. The attention-weighted sample-size model of visual short-term memory: Attention capture predicts resource allocation and memory load. Cogn. Psychol. 2016, 89, 71–105. [Google Scholar] [CrossRef] [PubMed]
- Brazier, M.A.B. Studies of the EEG activity of limbic structures in man. Electroencephalogr. Clin. Neurophysiol. 1968, 25, 309–318. [Google Scholar] [CrossRef]
- Melnik, A.; Hairston, W.D.; Ferris, D.P.; König, P. EEG correlates of sensorimotor processing: Independent components involved in sensory and motor processing. Sci. Rep. 2017, 7, 4461. [Google Scholar] [CrossRef] [PubMed]
- Samoilenko, A.; Petryshyn, R. Multifrequency Oscillations of Nonlinear Systems; KluwerAcademic Publishers: New York, NY, USA, 2004; ISBN 1402020317. [Google Scholar]
- Schmidt, H.; Avitabile, D.; Montbrio, E.; Roxin, A. Network mechanisms underlying the role of oscillations in cognitive tasks. PLoS Comput. Biol. 2018, 14, e1006430. [Google Scholar]
- Ahissar, M. Perceptual training: A tool for both modifying the brain and exploring it. Proc. Natl. Acad. Sci. USA 2001, 98, 11842–11843. [Google Scholar] [CrossRef]
- Wright, B.A.; Fitzgerald, M.B. Different patterns of human discrimination learning for two interaural cues to sound-source location. Proc. Natl. Acad. Sci. USA 2001, 98, 12307–12312. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.T.; Leung, J.; Burr, D.C.; Alais, D.; Ho, H.T.; Leung, J.; Burr, D.C.; Alais, D.; Morrone, M.C. Auditory Sensitivity and Decision Criteria Oscillate at Different Frequencies Separately for the Two Ears. Curr. Biol. 2017, 27, 3643–3649. [Google Scholar] [CrossRef] [PubMed]
- Marek, S.; Tervo-Clemmens, B.; Klein, N.; Foran, W.; Ghuman, A.S.; Luna, B. Adolescent development of cortical oscillations: Power, phase, and support of cognitive maturation. PLoS Biol. 2018, 16, e2004188. [Google Scholar] [CrossRef] [PubMed]
- Kriegeskorte, N.; Douglas, P.K. Cognitive computational neuroscience. Nat. Neurosci. 2018, 21, 1148–1160. [Google Scholar] [CrossRef]
- Güllich, A. International medallists’ and non-medallists’ developmental sport activities—A matched-pairs analysis. J. Sports Sci. 2017, 35, 2281–2288. [Google Scholar] [CrossRef]
- Cecchi, G.A.; Sigman, M.; Alonso, J.-M.; Martinez, L.; Chialvo, D.R.; Magnasco, M.O. Noise in neurons is message-dependent. Proc. Natl. Acad. Sci. USA 2000, 97, 5557–5561. [Google Scholar] [CrossRef]
- Vidoni, E.D.; McCarley, J.S.; Edwards, J.D.; Boyd, L.A. Manual and oculomotor performance develop contemporaneously but independently during continuous tracking. Exp. Brain Res. 2009, 195, 611–620. [Google Scholar] [CrossRef]
- Dean, H.L.; Martí, D.; Tsui, E.; Rinzel, J.; Pesaran, B. Reaction Time Correlations during Eye–Hand Coordination: Behavior and Modeling. J. Neurosci. 2011, 31, 2399–2412. [Google Scholar] [CrossRef] [PubMed]
- Haak, K.V.; Beckmann, C.F. Objective analysis of the topological organization of the human cortical visual connectome suggests three visual pathways. Cortex 2018, 98, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Ambrosio, B.; Aziz-Alaoui, M.A. Synchronization and control of coupled reaction-diffusion systems of the FitzHugh-Nagumo type. Comput. Math. Appl. 2012, 64, 934–943. [Google Scholar] [CrossRef]
- Ebsch, C.; Rosenbaum, R. Imbalanced amplification: A mechanism of amplification and suppression from local imbalance of excitation and inhibition in cortical circuits. PLoS Comput. Biol. 2018, 14, e1006048. [Google Scholar] [CrossRef]
- Graham, N.V. Beyond multiple pattern analyzers modeled as linear filters (as classical V1 simple cells): Useful additions of the last 25 years. Vis. Res. 2011, 51, 1397–1430. [Google Scholar] [CrossRef]
- Alexik, M. Modelling and identification of eye-hand dynamics. Simul. Pract. Theory 2000, 8, 25–38. [Google Scholar] [CrossRef]
- Schmidt-Hieber, C.; Nolan, M.F. Synaptic integrative mechanisms for spatial cognition. Nat. Neurosci. 2017, 20, 1483–1492. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, M.; Ding, M.; Truccolo, W.A.; Bressler, S.L. Evaluating causal relations in neural systems: Granger causality, directed transfer function and statistical assessment of significance. Biol. Cybern. 2001, 85, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Stefanescu, R.A.; Jirsa, V.K. A low dimensional description of globally coupled heterogeneous neural networks of excitatory and inhibitory neurons. PLoS Comput. Biol. 2008, 4, e1000219. [Google Scholar] [CrossRef] [PubMed]
- Senk, J.; Schuecker, J.; Hagen, E.; Diesmann, M.; Helias, M. Conditions for traveling waves in spiking neural networks. arXiv, 2018; arXiv:1801.06046. [Google Scholar]
- Bastin, J.; Lebranchu, P.; Jerbi, K.; Kahane, P.; Orban, G.; Lachaux, J.; Berthoz, A. NeuroImage Direct recordings in human cortex reveal the dynamics of gamma-band [50–150 Hz] activity during pursuit eye movement control. Neuroimage 2012, 63, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Hafed, Z.M. Alteration of Visual Perception prior to Microsaccades. Neuron 2013, 77, 775–786. [Google Scholar] [CrossRef]
- Irwin, D.E. Where does attention go when you blink? Attent. Percept. Psychophys. 2011, 73, 1374–1384. [Google Scholar] [CrossRef] [PubMed]
- Terhune, D.B.; Sullivan, J.G.; Simola, J.M. Time dilates after spontaneous blinking. Curr. Biol. 2016, 26, R459–R460. [Google Scholar] [CrossRef]
- Hoppe, D.; Helfmann, S.; Rothkopf, C.A. Humans quickly learn to blink strategically in response to environmental task demands. Proc. Natl. Acad. Sci. USA 2018, 115, 2246–2251. [Google Scholar] [CrossRef] [PubMed]
- Donner, T.H.; Siegel, M.; Oostenveld, R.; Fries, P.; Bauer, M.; Engel, A.K. Population Activity in the Human Dorsal Pathway Predicts the Accuracy of Visual Motion Detection. J. Neurosci. 2007, 98, 345–359. [Google Scholar] [CrossRef]
- Panchuk, A.; Rosin, D.P.; Hoevel, P.; Schoell, E. Synchronization of coupled neural oscillators with heterogeneous delays. Int. J. Bifurc. Chaos 2012, 23, 1330039. [Google Scholar] [CrossRef]
- Toit, P.J.; Krüger, P.E.; Mahomed, A.F.; Kleynhans, M.; Jay-du Preez, T.; Govender, C.; Mercier, J. The effect of ports vision exercises on the visual skills of university students. Afr. J. Phys. Health Educ. Recreat. Dance 2011, 17, 429–440. [Google Scholar]
- Clark, J.F.; Ellis, J.K.; Bench, J.; Khoury, J.; Graman, P. High-performance vision training improves batting statistics for University of Cincinnati baseball players. PLoS ONE 2012, 7, e29109. [Google Scholar] [CrossRef] [PubMed]
- Melstrom, A.J. Effectiveness of a Low-Budget Sports Vision Training Program for Improving Statistics of an NCAA Division I Baseball Team; South Dakota State University: Brookings, SD, USA, 2018. [Google Scholar]
- Kioumourtzoglou, E.; Kourtessis, T.; Michalopoulou, M.; Derri, V. Differences in Several Perceptual Abilities between Experts and Novices in Basketball, Volleyball and Water-Polo. Percept. Mot. Skills 1998, 86, 899–912. [Google Scholar] [CrossRef] [PubMed]
- Ghuntla, T.P.; Mehta, H.B.; Gokhale, P.A.; Shah, C.J. A comparative study of visual reaction time in basketball players and healthy controls. Indian J. Physiol. Pharmacol. 2012, 3, 49–51. [Google Scholar]
- Ghasemi, A.; Momeni, M.; Rezaee, M.; Gholami, A. The Difference in Visual Skills Between Expert Versus Novice Soccer Referees. J. Hum. Kinet. 2009, 22, 15–20. [Google Scholar] [CrossRef]
- Wilkins, L.; Nelson, C.; Tweddle, S. Stroboscopic Visual Training: A Pilot Study with Three Elite Youth Football Goalkeepers. J. Cogn. Enhanc. 2017, 2, 3–11. [Google Scholar] [CrossRef]
- Ellison, P.H. Eye-Hand Coordination: An Exploration of Measurement and Different Training methods using the SVT; Edge Hill University: Ormskirk, UK, 2015. [Google Scholar]
- Du Toit, P.J.; Van Vuuren, P.J.; Le Roux, S.; Henning, E.; Kleynhans, M.; Terblanche, H.C.; Crafford, D.; Grobbelaar, C.; Wood, P.S.; Grant, C.C.; et al. The effect of sport specific exercises on the visual skills of rugby players. Conf. Proc. IEEE Int. Conf. Syst. Man Cybern. 2012, 6, 1158–1161. [Google Scholar]
- Tseng, C.H.; Gobell, J.L.; Lu, Z.-L.L.; Sperling, G. When motion appears stopped: Stereo motion standstill. Proc. Natl. Acad. Sci. USA 2006, 103, 14953–14958. [Google Scholar] [CrossRef]
- Appelbaum, L.G.; Lu, Y.; Khanna, R.; Detwiler, K.R. The Effects of Sports Vision Training on Sensorimotor Abilities in Collegiate Softball Athletes. Athl. Train. Sport. Health Care 2016, 8, 154–163. [Google Scholar] [CrossRef]
- Hughes, P.K.; Bhundell, N.L.; Waken, J.M. Visual and psychomotor performance of elite, intermediate and novice table tennis competitors. Clin. Exp. Optom. 1993, 76, 51–60. [Google Scholar] [CrossRef]
- Reschke, M.F.; Somers, J.T.; Ford, G. Stroboscopic Vision as a Treatment forMotion Sickness: Strobe Lightning vs. Shutter Glasses. Aviat. Space Environ. Med. 2006, 77, 2–7. [Google Scholar] [PubMed]
- Berry, A.S.; Zanto, T.P.; Clapp, W.C.; Hardy, J.L.; Delahunt, P.B.; Mahncke, H.W.; Gazzaley, A. The Influence of Perceptual Training on Working Memory in Older Adults. PLoS ONE 2010, 5, e11537. [Google Scholar] [CrossRef] [PubMed]
- Nyquist, J.B.; Lappin, J.S.; Zhang, R.; Tadin, D. Perceptual training yields rapid improvements in visually impaired youth. Sci. Rep. 2016, 6, 37431. [Google Scholar] [CrossRef]
- Schmidt, J.T.; Buzzard, M. Activity-driven sharpening of the retinotectal projection in goldfish: Development under stroboscopic illumination prevents sharpening. J. Neurobiol. 1993, 24, 384–399. [Google Scholar] [CrossRef] [PubMed]
- Erickson, G. Sports Vision; Butterworth-Heinemann Elsevier Ltd.: Oxford, UK, 2007; ISBN 978-7506-7577-2. [Google Scholar]
- Singhal, A.; Culham, J.C.; Chinellato, E.; Goodale, M.A. Dual-task interference is greater in delayed grasping than in visually guided grasping. J. Vis. 2007, 7, 1–12. [Google Scholar] [PubMed]
- Vidnyanszky, Z.; Sohn, W. Learning to suppress task-irrelevant visual stimuli with attention. Vis. Res. 2005, 45, 677–685. [Google Scholar] [CrossRef]
- Paradiso, M.A.; Meshi, D.; Pisarcik, J.; Levine, S. Eye movements reset visual perception. J. Vis. 2012, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Appelbaum, L.G.; Erickson, G. Sports vision training: A review of the state-of-the-art in digital training techniques. Int. Rev. Sport Exerc. Psychol. 2016, 11, 160–189. [Google Scholar] [CrossRef]
- FitzHugh, R. Impulses and physiological states in theoretical models of nerve membrane. Biophys. J. 1961, 1, 445–446. [Google Scholar] [CrossRef]
- Nagumo, J.; Arimoto, S.; Yoshizawa, S. An active pulse transmission line simulating nerve axon. Proc. Inst. Radio Eng. IRE 1962, 50, 2061–2070. [Google Scholar] [CrossRef]
- Dmochowski, J.P.; Norcia, A.M. Cortical Components of Reaction-Time during Perceptual Decisions in Humans. PLoS ONE 2015, 10, e0143339. [Google Scholar]
- Marino, B.; Borghi, A.M.; Gemmi, L.; Cacciari, C.; Riggio, L. Neural Adaptation Effects in Conceptual Processing. Behav. Sci. 2015, 5, 353–371. [Google Scholar] [PubMed]
- Mamassian, P.; Goutcher, R. Temporal dynamics in bistable perception. J. Vis. 2005, 5, 361–375. [Google Scholar] [CrossRef]
- Noest, A.J.; van Ee, R.; Nijs, M.M.; van Wezel, R.J.A. Percept-choice sequences driven by interrupted ambiguous stimuli: A low-level neural model. J. Vis. 2007, 7, 10. [Google Scholar] [CrossRef]
- Fürstenau, N. Computational nonlinear dynamics model of percept switching with ambiguous stimuli. In Proceedings of the Second International Conference on Digital Human Modeling, San Diego, CA, USA, 19–24 July 2009. [Google Scholar]
- Kloosterman, N.A. Brain state and changes of mind: Probing the neural bases of multi-stable perceptual dynamics. Ph.D. Thesis, University of Amsterdam, Amsterdam, The Netherlands, 2015. [Google Scholar]
- Pisarchik, A.N.; Bashkirtseva, I.A.; Ryashko, L.B. Controlling bistability in a stochastic perception model. Eur. Phys. J. Spec. Top. 2015, 224, 1477–1484. [Google Scholar]
- Safaai, H.; Neves, R.; Eschenko, O.; Logothetis, N.K.; Panzeri, S. Modeling the effect of locus coeruleus firing on cortical state dynamics and single-trial sensory processing. Proc. Natl. Acad. Sci. USA 2015, 112, 12834–12839. [Google Scholar] [CrossRef]
- Montbrió, E.; Pazó, D.; Roxin, A. Macroscopic description for networks of spiking neurons. Phys. Rev. X 2015, 5, 1–15. [Google Scholar] [CrossRef]
- Jia, J.; Liu, H.; Xu, C.; Yan, F. Dynamic effects of time delay on a coupled FitzHugh-Nagumo neural system. Alexandria Eng. J. 2015, 54, 241–250. [Google Scholar] [CrossRef]
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerves. J. Physiol. 1952, 117, 500–544. [Google Scholar] [CrossRef]
- Loffing, F. Left-handedness and time pressure in elite interactive ball games. Biol. Lett. 2017, 13, 10–13. [Google Scholar] [CrossRef]
- Chen, N.; Majda, A.J. Beating the curse of dimension with accurate statistics for the Fokker–Planck equation in complex turbulent systems. Proc. Natl. Acad. Sci. USA 2017, 114, 12864–12869. [Google Scholar] [CrossRef] [PubMed]
- Dahlem, M.A.; Hiller, G.; Panchuk, A.; Schoell, E. Dynamics of delay-coupled excitable neural systems. Int. J. Bifurc. Chaos 2008, 19, 745–753. [Google Scholar] [CrossRef]
- Caksan, C.; Lehnert, J.; Schoell, E. Heterogeneous delays in neural networks. Eur. Phys. J. B 2014, 87, 54. [Google Scholar] [CrossRef]
- Zhou, J.; Yu, W.; Li, X.; Small, M.; Lu, J. Identifying the Topology of a Coupled FitzHugh–Nagumo Neurobiological Network via a Pinning Mechanism. IEEE Trans. Neural Netw. 2009, 20, 1679–1684. [Google Scholar] [CrossRef]
- Zeng, C.; Zeng, C.; Gong, A.; Nie, L. Effect of time delay in FitzHugh–Nagumo neural model with correlations between multiplicative and additive noises. Physica A 2010, 389, 5117–5127. [Google Scholar] [CrossRef]
- Marquie, P.; Comte, J.C.; Morfu, S. Analog simulation of neural information propagation using an electrical FitzHugh – Nagumo lattice. Chaos Solitons Fractals 2004, 19, 27–30. [Google Scholar] [CrossRef]
- FitzHugh, R. An electronic model of the nerve membrane for demonstration purposes. J. Appl. Physiol. 1966, 21, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Panchuk, A.; Dahlem, M.; Schoell, E. Regular spiking in asymmetrically delay-coupled FitzHugh-Nagumo systems. arXiv, 2009; arXiv:0911.2071. [Google Scholar]
- Yanchuk, S.; Perlikowski, P.; Popovych, O.V.; Tass, P.A. Variability of spatio-temporal patterns in non-homogeneous rings of spiking neurons. Chaos 2011, 21, 047511. [Google Scholar] [CrossRef]
- Shepelev, I.A.; Shamshin, D.V.; Strelkova, G.I.; Vadivasova, T.E. Bifurcations of spatiotemporal structures in a medium of FitzHugh–Nagumo neurons with diffusive coupling. Chaos Solit. Fract. 2017, 104, 153–160. [Google Scholar] [CrossRef]
- Shepelev, I.A.; Vadivasova, T.E.; Bukh, A.V.; Strelkova, G.I.; Anishchenko, V.S. New type of chimera structures in a ring of bistable FitzHugh–Nagumo oscillators with nonlocal interaction. Phys. Lett. Sect. A Gen. At. Solid State Phys. 2017, 381, 1398–1404. [Google Scholar] [CrossRef]
- Churchland, M.M.; Yu, B.M.; Cunningham, J.P.; Sugrue, L.P.; Cohen, M.R.; Corrado, G.S.; Newsome, W.T.; Clark, A.M.; Hosseini, P.; Scott, B.B.; et al. Stimulus onset quenches neural variability: A widespread cortical phenomenon. Nat. Neurosci. 2010, 13, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Chang, J. Flipping Biological Switches: Solving for Optimal Control. Ph.D. Thesis, University of Massachesetts, Worcester, MA, USA, 2015. [Google Scholar]
- Kwon, O.-S.; Tadin, D.; Knill, D.C. Unifying account of visual motion and position perception. Proc. Natl. Acad. Sci. USA 2015, 112, 8142–8147. [Google Scholar] [CrossRef]
- Plotnikov, S.A.; Lehnert, J.; Fradkov, A.L.; Schöll, E. Synchronization in heterogeneous FitzHugh-Nagumo networks with hierarchical architecture. Int. J. Bifurc. Chaos 2016, 94, 012203. [Google Scholar] [CrossRef]
- Manafian, J.; Lakestani, M. New Improvement of the Expansion Methods for Solving the Generalized Fitzhugh-Nagumo Equation with Time-Dependent Coefficients. Int. J. Eng. Math. 2015, 2015, 107978. [Google Scholar] [CrossRef]
- Singh, B.K.; Arora, G.; Singh, M.K. A numerical scheme for the generalized Burgers-Huxley equation. J. Egypt. Math. Soc. 2016, 24, 629–637. [Google Scholar] [CrossRef]
- Chang, C.-J.; Jazayeri, M. Integration of speed and time for estimating time to contact. Proc. Natl. Acad. Sci. USA 2018, 115, E2879–E2887. [Google Scholar] [CrossRef] [PubMed]
- Johns, M.; Crowley, K.; Chapman, R.; Tucker, A.; Hocking, C. The effect of blinks and saccadic eye movements on visual reaction times. Atten. Percept. Psychophys. 2009, 71, 783–788. [Google Scholar] [CrossRef]
- Van Vugt, B.; Dagnino, B.; Vartak, D.; Safaai, H.; Panzeri, S.; Dehaene, S. The threshold for conscious report: Signal loss and response bias in visual and frontal cortex. Science 2018, 360, 537–542. [Google Scholar] [CrossRef]
- Ghadirzadeh, A.; Maki, A.; Bjorkman, M. A sensorimotor approach for self-learning of hand-eye coordination. IEEE Int. Conf. Intell. Robot. Syst. 2015, 4969–4975. [Google Scholar]
- Rao, H.M.; Khanna, R.; Zielinski, D.J.; Lu, Y.; Clements, J.M.; Potter, N.D.; Sommer, M.A.; Kopper, R. Sensorimotor Learning during a Marksmanship Task in Immersive Virtual Reality. Front. Psychol. 2018, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.T.; Khan, K.; Akbar, M.A. Study of Nonlinear Evolution Equations to Construct Traveling Wave Solutions via Modified Simple Equation Method. Phys. Rev. Res. Int. 2013, 3, 490–503. [Google Scholar]
- Khan, K.; Akbar, M.A.; Koppelaar, H. Study of coupled nonlinear partial differential equations for finding exact analytical solutions. R. Soc. Open Sci. 2015, 2, 140406. [Google Scholar] [CrossRef]
- Khan, K.; Koppelaar, H.; Akbar, M.A. Exact and numerical soliton solutions to nonlinear wave equations. Casp. J. Comput. Math. Eng. 2016, 2, 5–22. [Google Scholar]
- Vasiev, B.N. Classification of patterns in excitable systems with lateral inhibition. Phys. Lett. A 2004, 323, 194–203. [Google Scholar] [CrossRef]
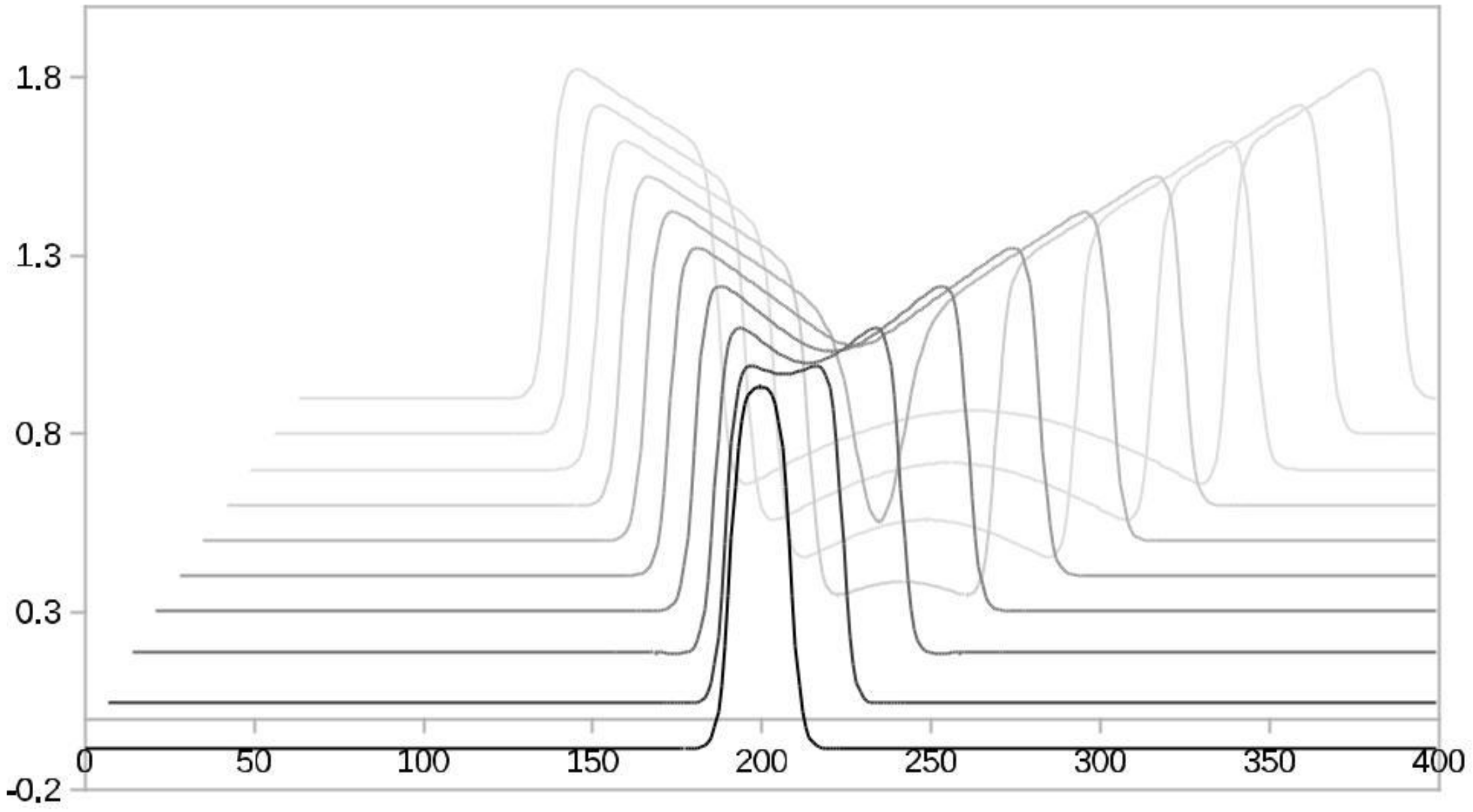
Figure 1.
An FHN activator neuron’s initial spike splits and spreads in time. The time delays are visible by decreasingly paler black shades.
Figure 1.
An FHN activator neuron’s initial spike splits and spreads in time. The time delays are visible by decreasingly paler black shades.

Figure 2.
From Reference [45] and adapted from Figure 4 in Reference [20]. Two FitzHugh-Nagumo neuron groups with coupling strength C and internal feedback strength K exhibit different delay times forward and backward.

Figure 3.
Scaled FHN Nullclines in the phase plane showing two stable points (zero-crossings of the third order Nullcline curve) and one unstable point in the origin.
Figure 3.
Scaled FHN Nullclines in the phase plane showing two stable points (zero-crossings of the third order Nullcline curve) and one unstable point in the origin.

Figure 4.
Summary of the above result: Reaction Times: 60 s without delay FITLIGHT Trainer™ equipment. Sample size of 85 men and women. Baseline sample mean 0.721 s, 0.1075, and Final mean 0.646 s with 0.0989. P-value 0.0000914155. Result: Hypothesis of NO-Effect of training is: Rejected.
Figure 4.
Summary of the above result: Reaction Times: 60 s without delay FITLIGHT Trainer™ equipment. Sample size of 85 men and women. Baseline sample mean 0.721 s, 0.1075, and Final mean 0.646 s with 0.0989. P-value 0.0000914155. Result: Hypothesis of NO-Effect of training is: Rejected.

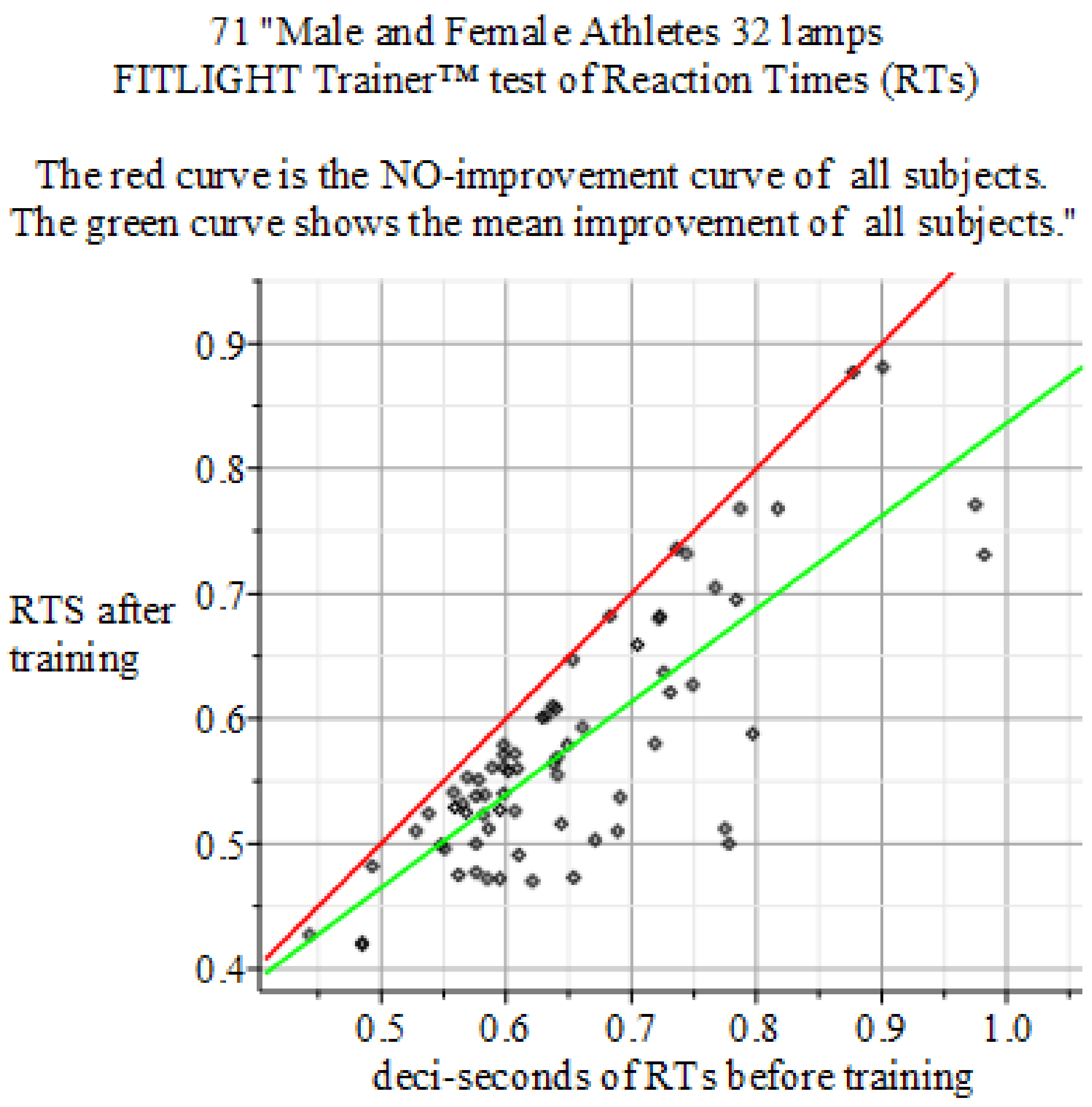
Figure 5.
Summary of the above result. Reaction Times: 32 lamps FITLIGHT Trainer™ equipment. Sample size of 71 men and women. Baseline sample mean 0.650 s, 0.1075 and Final mean 0.577 s with 0.0988966. P-value 0.0000383454. Result: Hypothesis of NO-Effect of training is: Rejected.
Figure 5.
Summary of the above result. Reaction Times: 32 lamps FITLIGHT Trainer™ equipment. Sample size of 71 men and women. Baseline sample mean 0.650 s, 0.1075 and Final mean 0.577 s with 0.0988966. P-value 0.0000383454. Result: Hypothesis of NO-Effect of training is: Rejected.

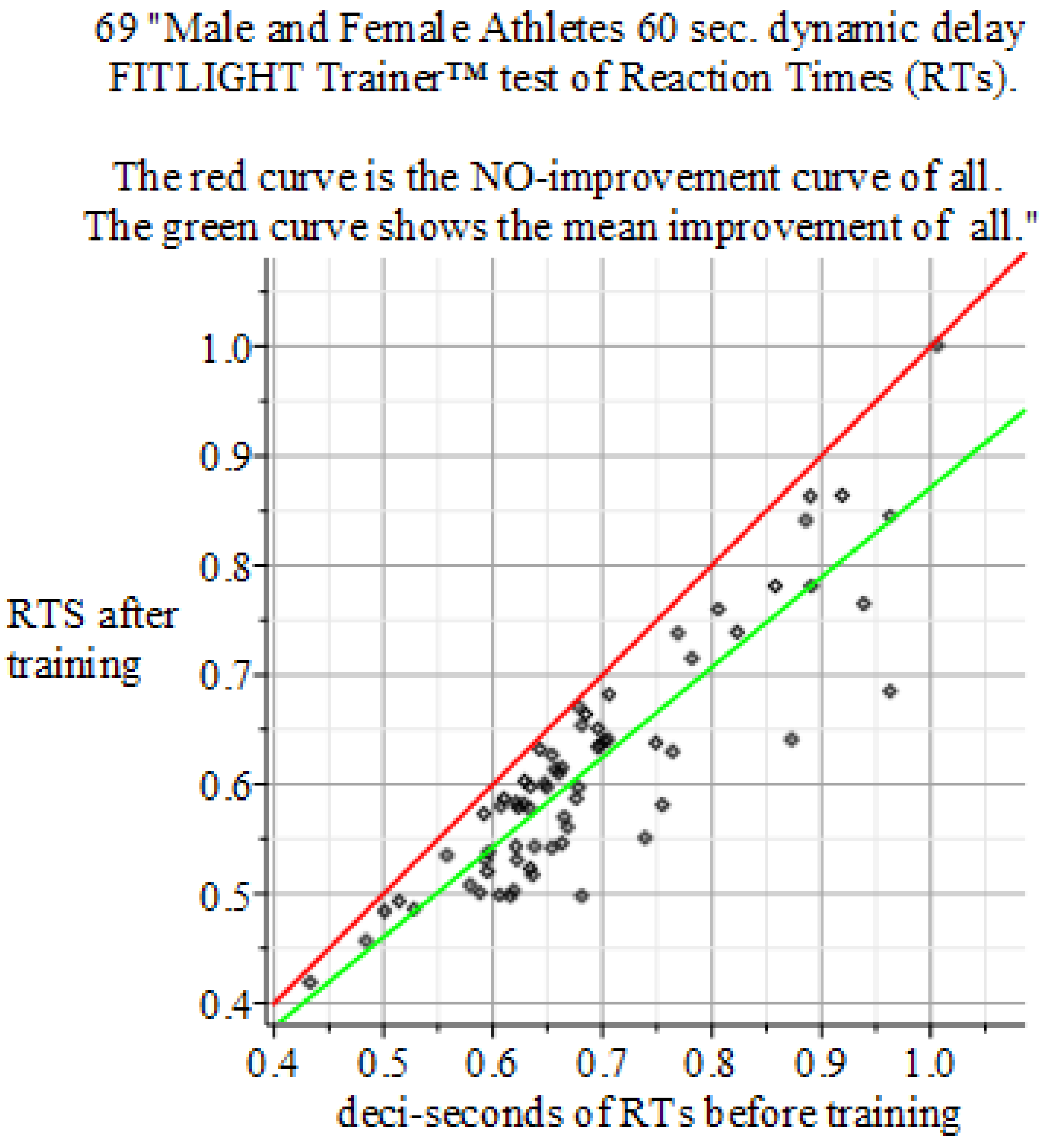
Figure 6.
Summary of the above results. Reaction Times: 60 s dynamic delay FITLIGHT Trainer™ equipment. Sample size of 69 men and women. Baseline sample mean 0.687 s, 0.1194, and Final mean 0.614 s with 0.1099. P-value 0.0000914155. Result: Hypothesis of NO-Effect of training is: Rejected. The mean outcome of the 225 Reaction Time experiments is: Reaction Time performance improvement of +10.8% against the initial intake measurement.
Figure 6.
Summary of the above results. Reaction Times: 60 s dynamic delay FITLIGHT Trainer™ equipment. Sample size of 69 men and women. Baseline sample mean 0.687 s, 0.1194, and Final mean 0.614 s with 0.1099. P-value 0.0000914155. Result: Hypothesis of NO-Effect of training is: Rejected. The mean outcome of the 225 Reaction Time experiments is: Reaction Time performance improvement of +10.8% against the initial intake measurement.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Koppelaar, H.; Kordestani Moghadam, P.; Khan, K.; Kouhkani, S.; Segers, G.; van Warmerdam, M. Reaction Time Improvements by Neural Bistability. Behav. Sci. 2019, 9, 28. https://0-doi-org.brum.beds.ac.uk/10.3390/bs9030028
AMA Style
Koppelaar H, Kordestani Moghadam P, Khan K, Kouhkani S, Segers G, van Warmerdam M. Reaction Time Improvements by Neural Bistability. Behavioral Sciences. 2019; 9(3):28. https://0-doi-org.brum.beds.ac.uk/10.3390/bs9030028
Chicago/Turabian StyleKoppelaar, Henk, Parastou Kordestani Moghadam, Kamruzzaman Khan, Sareh Kouhkani, Gijs Segers, and Martin van Warmerdam. 2019. "Reaction Time Improvements by Neural Bistability" Behavioral Sciences 9, no. 3: 28. https://0-doi-org.brum.beds.ac.uk/10.3390/bs9030028
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.