Neurochemical and Behavioral Characterization after Acute and Repeated Exposure to Novel Synthetic Cannabinoid Agonist 5-MDMB-PICA

 ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
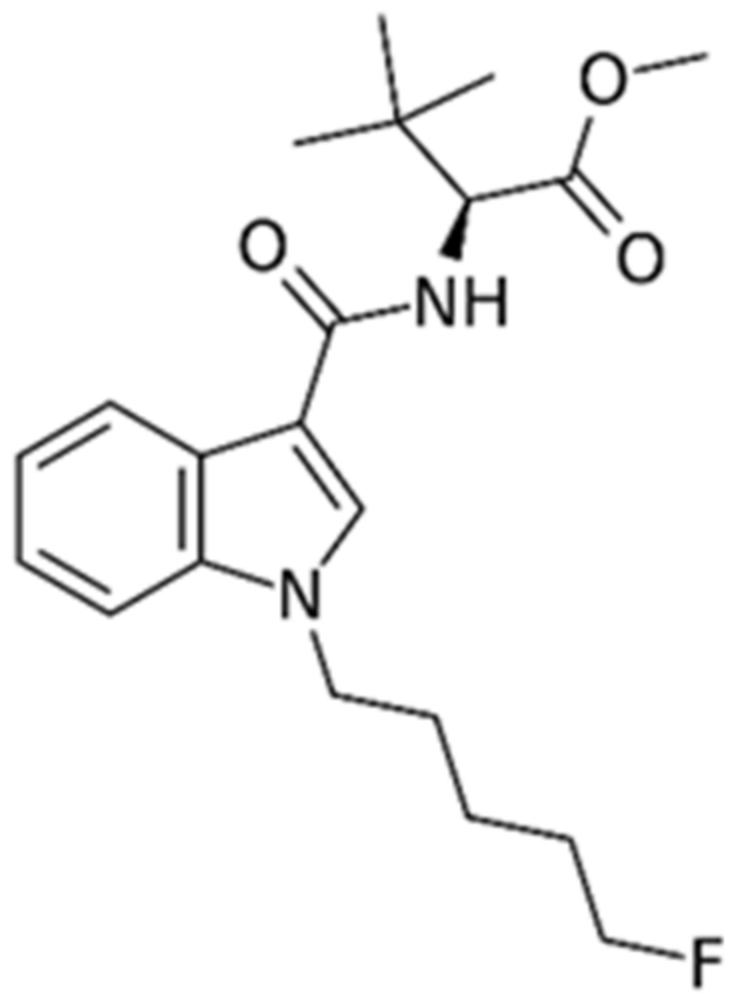
2.2. Drugs
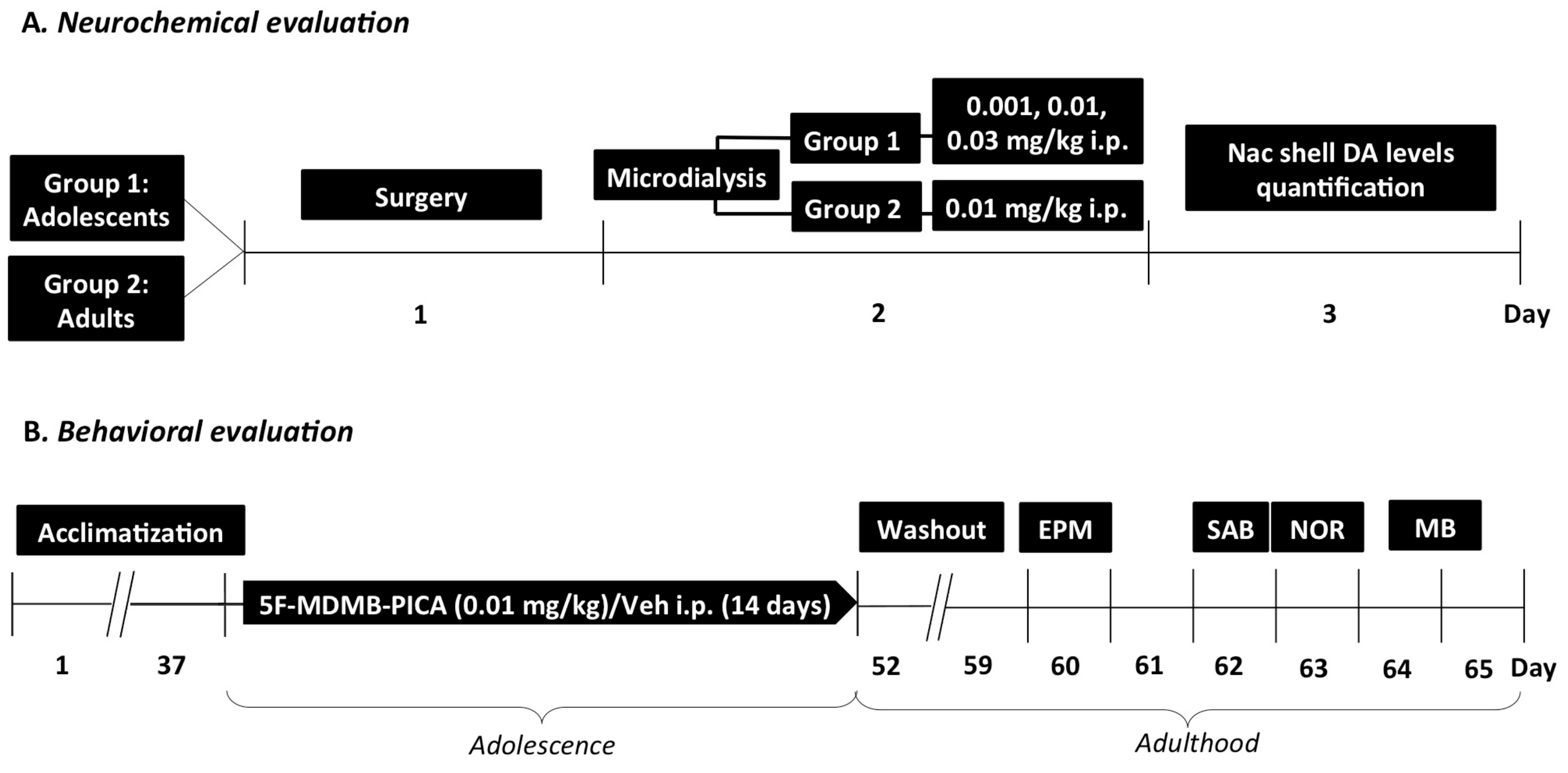
2.3. Experimental Design Timeline
2.4. In Vivo Brain Microdialysis
2.4.1. Preparation of Microdialysis Probe
2.4.2. Surgery
2.4.3. Dopamine Assessment
2.4.4. Histology
2.5. Behavioral Tests
2.5.1. Elevated Plus Maze
2.5.2. Spontaneous Alternation Behavior in a Y-Maze
2.5.3. Novel Object Recognition Test
2.5.4. Marble Burying
2.6. Statistical Analysis
3. Results
3.1. In Vivo Neurochemical Effects of 5F-MDMB PICA
3.1.1. Microdialysis Studies
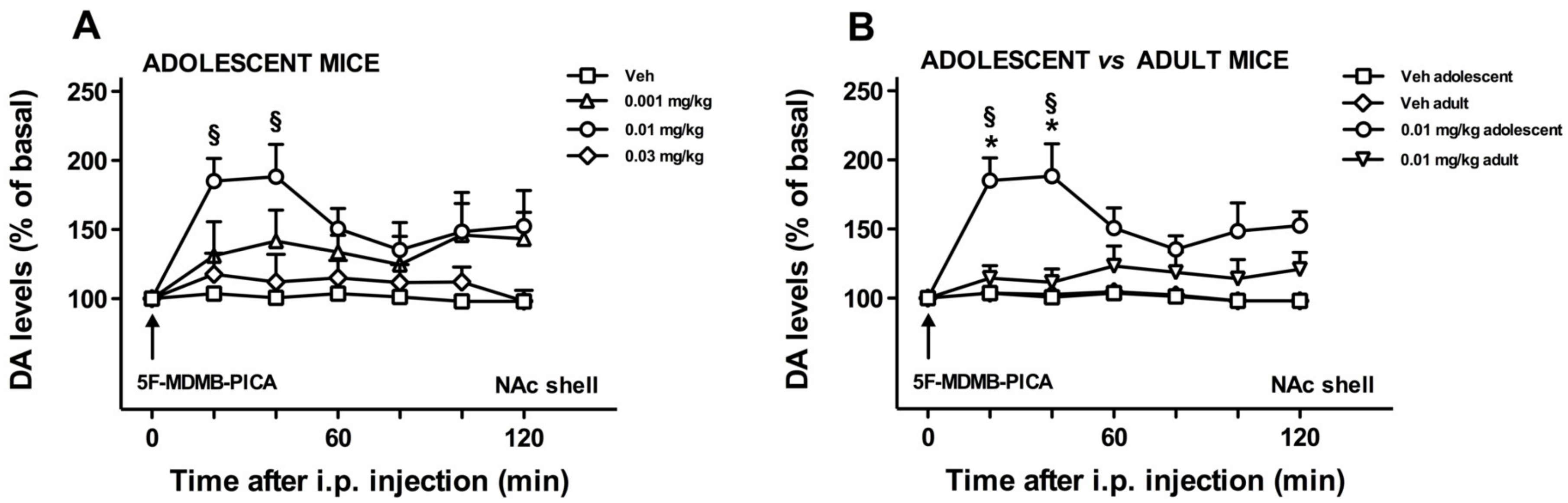
3.1.2. Effects of 5F-MDMB-PICA Administration on DA Transmission in the NAc Shell of Adolescent and Adult Mice
3.1.3. Role of Age on the NAc Shell DA Stimulation Induced by 5F-MDMB-PICA
3.2. Behavioral Effects at Adulthood after 5F-MDMB-PICA Repeated Treatment during Adolescence
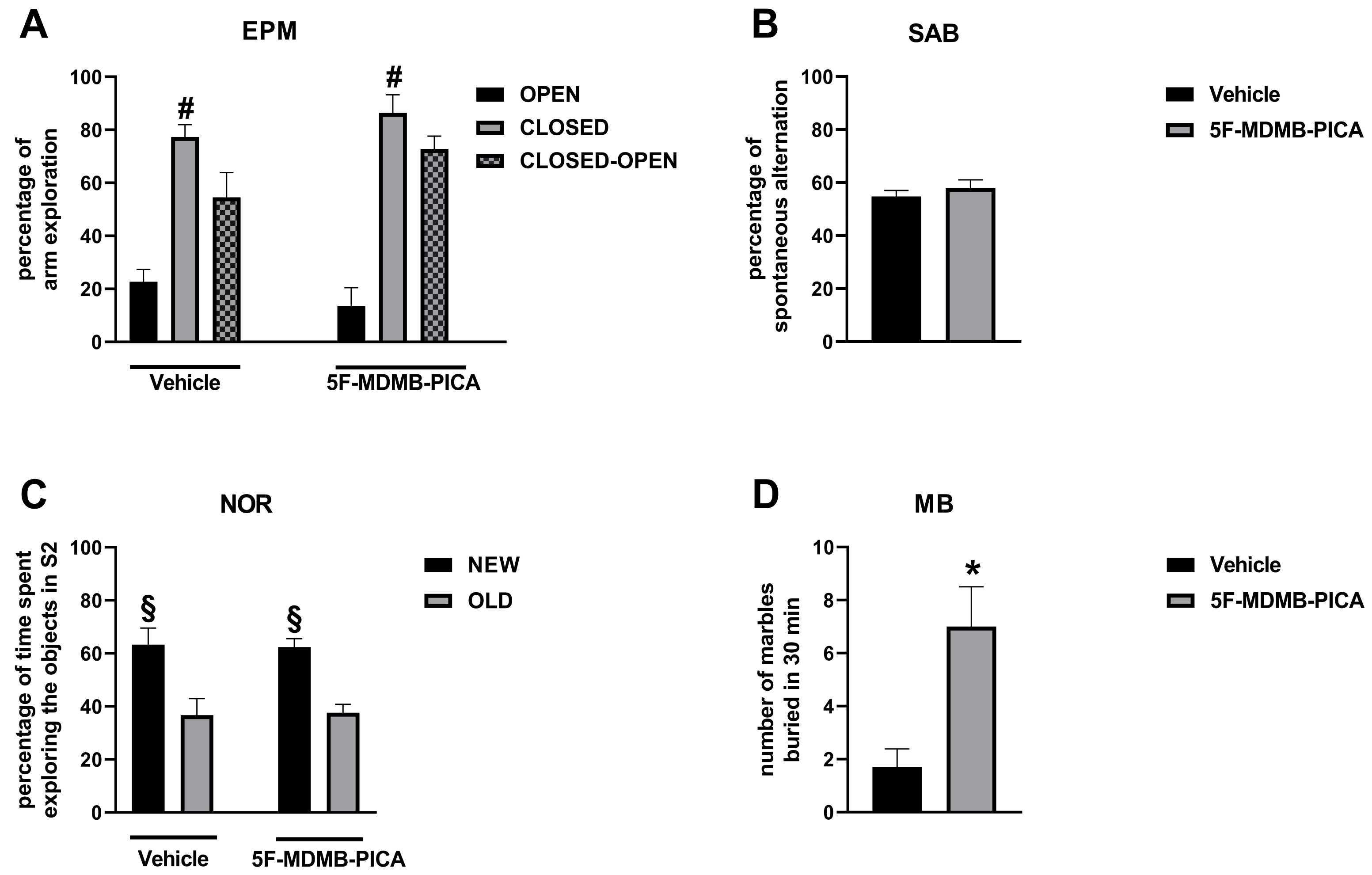
3.2.1. EPM
3.2.2. SAB
3.2.3. NOR
3.2.4. MB
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- United Nations Office on Drugs and Crime. World Drug Report 2013; Sales No. E.13.XI.6; United Nations Office on Drugs and Crime (United Nations publication): Vienna, Austria, 2013. [Google Scholar]
- European Monitoring Centre for Drugs and Drug Addiction. European Drug Report; European Union Publications Office: Luxembourg, 2019; Available online: http://www.emcdda.europa.eu/system/files/publications/4541/TDAT17001ENN.pdf_en (accessed on 15 October 2020).
- Huffman, J.W.; Padgett, L.W. Recent developments in the medicinal chemistry of cannabimimetic indoles, pyrroles and indenes. Curr. Med. Chem. 2005, 12, 1395–1411. [Google Scholar] [CrossRef]
- Wintermeyer, A.; Möller, I.; Thevis, M.; Jübner, M.; Beike, J.; Rothschild, M.A.; Bender, K. In vitro phase I metabolism of the synthetic cannabimimetic JWH-018. Anal. Bioanal. Chem. 2010, 398, 2141–2153. [Google Scholar] [CrossRef] [PubMed]
- Every-Palmer, S. Synthetic cannabinoid JWH-018 and psychosis: An explorative study. Drug Alcohol Depend. 2011, 117, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Seely, K.A.; Lapoint, J.; Moran, J.H.; Fattore, L. Spice drugs are more than harmless herbal blends: A review of the pharmacology and toxicology of synthetic cannabinoids. Prog. Neuropsychopharmacol. Biol. Psychiatry 2012, 39, 234–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papanti, D.; Schifano, F.; Botteon, G.; Bertossi, F.; Mannix, J.; Vidoni, D.; Impagnatiello, M.; Pascolo-Fabrici, E.; Bonavigo, T. “Spiceophrenia”: A systematic overview of “spice”—Related psychopathological issues and a case report. Hum. Psychopharmacol. 2013, 28, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Fantegrossi, W.E.; Moran, J.H.; Radominska-Pandya, A.; Prather, P.L. Distinct pharmacology and metabolism of K2 synthetic cannabinoids compared to Δ(9)-THC: Mechanism underlying greater toxicity? Life Sci. 2014, 97, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Wagner, K.D.; Armenta, R.F.; Roth, A.M.; Maxwell, J.C.; Cuevas-Mota, J.; Garfein, R.S. Use of synthetic cathinones and cannabimimetics among injection drug users in San Diego, California. Drug Alcohol Depend. 2014, 141, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.R.; Boomhower, S.R.; Newland, M.C. Behavioral effects of chronic WIN 55,212-2 administration during adolescence and adulthood in mice. Exp. Clin. Psychopharmacol. 2019, 27, 348–358. [Google Scholar] [CrossRef]
- Mogler, L.; Franz, F.; Rentsch, D.; Angerer, V.; Weinfurtner, G.; Longworth, M.; Banister, S.D.; Kassiou, M.; Moosmann, B.; Auwärter, V. Detection of the recently emerged synthetic cannabinoid 5F-MDMB-PICA in ‘legal high’ products and human urine samples. Drug Test. Anal. 2018, 10, 196–205. [Google Scholar] [CrossRef]
- WHO. 5F-MDMB-PICA. In Proceedings of the Expert Committee on Drug Dependence Forty-second Meeting, Geneva, Switzerland, 21–25 October 2019. [Google Scholar]
- UNODC Early Warning Advisory (EWA) Tox-Portal. Available online: https://www.unodc.org/tox/-/login (accessed on 15 October 2020).
- Truver, M.T.; Watanabe, S.; Åstrand, A.; Vikingsson, S.; Green, H.; Swortwood, M.J.; Kronstrand, R. 5F-MDMB-PICA metabolite identification and cannabinoid receptor activity. Drug Test. Anal. 2020, 12, 127–135. [Google Scholar] [CrossRef]
- Noble, C.; Cannaert, A.; Linnet, K.; Stove, C.P. Application of an activity-based receptor bioassay to investigate the in vitro activity of selected indole- and indazole-3-carboxamide-based synthetic cannabinoids at CB1 and CB2 receptors. Drug Test. Anal. 2019, 11, 501–511. [Google Scholar] [CrossRef]
- Sachdev, S.; Vemuri, K.; Banister, S.D.; Longworth, M.; Kassiou, M.; Santiago, M.; Makriyannis, A.; Connor, M. In vitro determination of the efficacy of illicit synthetic cannabinoids at CB1 receptors. Br. J. Pharmacol. 2019, 176, 4653–4665. [Google Scholar] [CrossRef]
- De Luca, M.A.; Castelli, M.P.; Loi, B.; Porcu, A.; Martorelli, M.; Miliano, C.; Kellett, K.; Davidson, C.; Stair, J.L.; Schifano, F.; et al. Native CB1 receptor affinity, intrinsic activity and accumbens shell dopamine stimulant properties of third generation SPICE/K2 cannabinoids: BB-22, 5F-PB-22, 5F-AKB-48 and STS-135. Neuropharmacology 2016, 105, 630–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Luca, M.A.; Bimpisidis, Z.; Melis, M.; Marti, M.; Caboni, P.; Valentini, V.; Margiani, G.; Pintori, N.; Polis, I.; Marsicano, G.; et al. Stimulation of in vivo dopamine transmission and intravenous self-administration in rats and mice by JWH-018, a Spice cannabinoid. Neuropharmacology 2015, 99, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Banister, S.D.; Longworth, M.; Kevin, R.; Sachdev, S.; Santiago, M.; Stuart, J.; Mack, J.B.; Glass, M.; McGregor, I.S.; Connor, M.; et al. Pharmacology of Valinate and tert-Leucinate Synthetic Cannabinoids 5F-AMBICA, 5F-AMB, 5F-ADB, AMB-FUBINACA, MDMB-FUBINACA, MDMB-CHMICA, and Their Analogues. ACS Chem. Neurosci. 2016, 7, 1241–1254. [Google Scholar] [CrossRef] [PubMed]
- Miliano, C.; Margiani, G.; Fattore, L.; De Luca, M.A. Sales and Advertising Channels of New Psychoactive Substances (NPS): Internet, Social Networks, and Smartphone Apps. Brain Sci. 2018, 8, 123. [Google Scholar] [CrossRef] [Green Version]
- Koob, G.F.; Volkow, N.D. Neurocircuitry of addiction. Neuropsychopharmacology 2010, 35, 217–238. [Google Scholar] [CrossRef] [Green Version]
- Di Chiara, G.; Bassareo, V.; Fenu, S.; De Luca, M.A.; Spina, L.; Cadoni, C.; Acquas, E.; Carboni, E.; Valentini, V.; Lecca, D. Dopamine and drug addiction: The nucleus accumbens shell connection. Neuropharmacology 2004, 47 (Suppl 1), 227–241. [Google Scholar] [CrossRef]
- Canazza, I.; Ossato, A.; Trapella, C.; Fantinati, A.; De Luca, M.A.; Margiani, G.; Vincenzi, F.; Rimondo, C.; Di Rosa, F.; Gregori, A.; et al. Effect of the novel synthetic cannabinoids AKB48 and 5F-AKB48 on “tetrad”, sensorimotor, neurological and neurochemical responses in mice. In vitro and in vivo pharmacological studies. Psychopharmacology 2016, 233, 3685–3709. [Google Scholar] [CrossRef]
- Ossato, A.; Uccelli, L.; Bilel, S.; Canazza, I.; Di Domenico, G.; Pasquali, M.; Pupillo, G.; De Luca, M.A.; Boschi, A.; Vincenzi, F.; et al. Psychostimulant Effect of the Synthetic Cannabinoid JWH-018 and AKB48: Behavioral, Neurochemical, and Dopamine Transporter Scan Imaging Studies in Mice. Front. Psychiatry 2017, 8, 130. [Google Scholar] [CrossRef] [Green Version]
- Bilel, S.; Tirri, M.; Arfè, R.; Stopponi, S.; Soverchia, L.; Ciccocioppo, R.; Frisoni, P.; Strano-Rossi, S.; Miliano, C.; De-Giorgio, F.; et al. Pharmacological and Behavioral Effects of the Synthetic Cannabinoid AKB48 in Rats. Front. Neurosci. 2019, 13, 1163. [Google Scholar] [CrossRef] [PubMed]
- Zeitlin, H. Psychiatric comorbidity with substance misuse in children and teenagers. Drug Alcohol Depend. 1999, 55, 225–234. [Google Scholar] [CrossRef]
- Leeman, R.F.; DeMartini, K.S.; Gueorguieva, R.; Nogueira, C.; Corbin, W.R.; Neighbors, C.; O’Malley, S.S. Randomized controlled trial of a very brief, multicomponent web-based alcohol intervention for undergraduates with a focus on protective behavioral strategies. J. Consult. Clin. Psychol. 2016, 84, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Richmond-Rakerd, L.S.; Slutske, W.S.; Lynskey, M.T.; Agrawal, A.; Madden, P.A.; Bucholz, K.K.; Heath, A.C.; Statham, D.J.; Martin, N.G. Age at first use and later substance use disorder: Shared genetic and environmental pathways for nicotine, alcohol, and cannabis. J. Abnorm. Psychol. 2016, 125, 946–959. [Google Scholar] [CrossRef]
- Di Forti, M.; Quattrone, D.; Freeman, T.P.; Tripoli, G.; Gayer-Anderson, C.; Quigley, H.; Rodriguez, V.; Jongsma, H.E.; Ferraro, L.; La Cascia, C.; et al. EU-GEI WP2 Group: The contribution of cannabis use to variation in the incidence of psychotic disorder across Europe (EU-GEI): A multicentre case-control study. Lancet Psychiatry. Lancet Psychiatry 2019, 6, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Khokhar, J.Y.; Dwiel, L.L.; Henricks, A.M.; Doucette, W.T.; Green, A.I. The link between schizophrenia and substance use disorder: A unifying hypothesis. Schizophr. Res. 2018, 194, 78–85. [Google Scholar] [CrossRef]
- Gleason, K.A.; Birnbaum, S.G.; Shukla, A.; Ghose, S. Susceptibility of the adolescent brain to cannabinoids: Long-term hippocampal effects and relevance to schizophrenia. Transl. Psychiatry 2012, 2, e199. [Google Scholar] [CrossRef] [Green Version]
- Rice, D.; Barone, S., Jr. Critical periods of vulnerability for the developing nervous system: Evidence from humans and animal models. Environ. Health Perspect. 2000, 108 (Suppl. 3), 511–533. [Google Scholar] [CrossRef]
- De Luca, M.A.; Buczynski, M.W.; Di Chiara, G. Loren Parsons’ contribution to addiction neurobiology. Addict. Biol. 2018, 23, 1207–1222. [Google Scholar] [CrossRef]
- Pellow, S.; Chopin, P.; File, S.E.; Briley, M. Validation of open:closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J. Neurosci. Methods 1985, 14, 149–167. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Green, M.R.; Martin, B.R. Pharmacological characterization of cannabinoids in the elevated plus maze. J. Pharmacol. Exp. Ther. 1990, 253, 1002–1009. [Google Scholar] [PubMed]
- Hughes, R.N. The value of spontaneous alternation behavior (SAB) as a test of retention in pharmacological investigations of memory. Neurosci. Biobehav. Rev. 2004, 28, 497–505. [Google Scholar] [CrossRef]
- Costa, G.; Simola, N.; Morelli, M. MDMA administration during adolescence exacerbates MPTP-induced cognitive impairment and neuroinflammation in the hippocampus and prefrontal cortex. Psychopharmacology 2014, 231, 4007–4018. [Google Scholar] [CrossRef] [PubMed]
- Ennaceur, A.; Delacour, J. A new one-trial test for neurobiological studies of memory in rats. 1: Behavioral data. Behav. Brain Res. 1988, 31, 47–59. [Google Scholar] [CrossRef]
- Albelda, N.; Joel, D. Current animal models of obsessive compulsive disorder: An update. Neuroscience 2012, 211, 83–106. [Google Scholar] [CrossRef]
- Margiani, G.; Caria, F.; Musa, A.; De Luca, M.A. Intravenous Self-administration of the synthetic cannabinoid receptor agonist 5F-MDMB-PICA in adolescent mice: Neurobiological sequelae in adulthood. (in preparation)
- Tanda, G.; Pontieri, F.E.; Di Chiara, G. Cannabinoid and heroin activation of mesolimbic dopamine transmission by a common mu1 opioid receptor mechanism. Science 1997, 276, 2048–2050. [Google Scholar] [CrossRef] [Green Version]
- Lecca, D.; Cacciapaglia, F.; Valentini, V.; Di Chiara, G. Monitoring extracellular dopamine in the rat nucleus accumbens shell and core during acquisition and maintenance of intravenous WIN 55,212-2 self-administration. Psychopharmacology 2006, 188, 63–74. [Google Scholar] [CrossRef]
- De Luca, M.A.; Di Chiara, G.; Cadoni, C.; Lecca, D.; Orsolini, L.; Papanti, D.; Corkery, J.; Schifano, F. Cannabis; Epidemiological, Neurobiological and Psychopathological Issues: An Update. CNS Neurol. Disord. Drug Targets 2017, 16, 598–609. [Google Scholar] [CrossRef] [Green Version]
- Corongiu, S.; Dessì, C.; Cadoni, C. Adolescence versus adulthood: Differences in basal mesolimbic and nigrostriatal dopamine transmission and response to drugs of abuse. Addict. Biol. 2020, 25, e12721. [Google Scholar] [CrossRef]
- Frantz, K.J.; O’Dell, L.E.; Parsons, L.H. Behavioral and neurochemical responses to cocaine in periadolescent and adult rats. Neuropsychopharmacology 2007, 32, 625–637. [Google Scholar] [CrossRef] [Green Version]
- Shahbazi, M.; Moffett, A.M.; Williams, B.F.; Frantz, K.J. Age- and sex-dependent amphetamine self-administration in rats. Psychopharmacology 2008, 196, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Doremus-Fitzwater, T.L.; Varlinskaya, E.I.; Spear, L.P. Motivational systems in adolescence: Possible implications for age differences in substance abuse and other risk-taking behaviors. Brain Cogn. 2010, 72, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Natividad, L.A.; Tejeda, H.A.; Torres, O.V.; O’Dell, L.E. Nicotine withdrawal produces a decrease in extracellular levels of dopamine in the nucleus accumbens that is lower in adolescent versus adult male rats. Synapse 2010, 64, 136–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natividad, L.A.; Buczynski, M.W.; Parsons, L.H.; Torres, O.V.; O’Dell, L.E. Adolescent rats are resistant to adaptations in excitatory and inhibitory mechanisms that modulate mesolimbic dopamine during nicotine withdrawal. J. Neurochem. 2012, 123, 578–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellgren, M.; Artmann, A.; Tkalych, O.; Gupta, A.; Hansen, H.S.; Hansen, S.H.; Devi, L.A.; Hurd, Y.L. Dynamic changes of the endogenous cannabinoid and opioid mesocorticolimbic systems during adolescence: THC effects. Eur. Neuropsychopharmacol. 2008, 18, 826–834. [Google Scholar] [CrossRef] [Green Version]
- Meyer, M.R. Toxicokinetics of NPS: Update 2017. Handb. Exp. Pharmacol. 2018, 252, 441–459. [Google Scholar] [CrossRef]
- Chadwick, B.; Miller, M.L.; Hurd, Y.L. Cannabis Use during Adolescent Development: Susceptibility to Psychiatric Illness. Front. Psychiatry 2013, 4, 129. [Google Scholar] [CrossRef] [Green Version]
- Rubino, T.; Zamberletti, E.; Parolaro, D. Endocannabinoids and Mental Disorders. Handb. Exp. Pharmacol. 2015, 231, 261–283. [Google Scholar] [CrossRef]
- Rubino, T.; Parolaro, D. Sex-dependent vulnerability to cannabis abuse in adolescence. Front. Psychiatry 2015, 6, 56. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Gomez, C.; Osentoski, A.; Woods, J.H. Pharmacological evaluation of the adequacy of marble burying as an animal model of compulsion and/or anxiety. Behav. Pharmacol. 2011, 22, 711–713. [Google Scholar] [CrossRef] [Green Version]
- Frontera, J.L.; Gonzalez Pini, V.M.; Messore, F.L.; Brusco, A. Exposure to cannabinoid agonist WIN 55,212-2 during early adolescence increases alcohol preference and anxiety in CD1 mice. Neuropharmacology 2018, 137, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Zanda, M.T.; Fadda, P.; Antinori, S.; Di Chio, M.; Fratta, W.; Chiamulera, C.; Fattore, L. Methoxetamine affects brain processing involved in emotional response in rats. Br. J. Pharmacol. 2017, 174, 3333–3345. [Google Scholar] [CrossRef] [PubMed]
- Tomas-Roig, J.; Benito, E.; Agis-Balboa, R.C.; Piscitelli, F.; Hoyer-Fender, S.; Di Marzo, V.; Havemann-Reinecke, U. Chronic exposure to cannabinoids during adolescence causes long-lasting behavioral deficits in adult mice. Addict. Biol. 2017, 22, 1778–1789. [Google Scholar] [CrossRef]
- Pushkin, A.N.; Eugene, A.J.; Lallai, V.; Torres-Mendoza, A.; Fowler, J.P.; Chen, E.; Fowler, C.D. Cannabinoid and nicotine exposure during adolescence induces sex-specific effects on anxiety- and reward-related behaviors during adulthood. PLoS ONE 2019, 14, e0211346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, G.; De Luca, M.A.; Piras, G.; Marongiu, J.; Fattore, L.; Simola, N. Neuronal and peripheral damages induced by synthetic psychoactive substances: An update of recent findings from human and animal studies. Neural Regen. Res. 2020, 15, 802–816. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Musa, A.; Simola, N.; Piras, G.; Caria, F.; Onaivi, E.S.; De Luca, M.A. Neurochemical and Behavioral Characterization after Acute and Repeated Exposure to Novel Synthetic Cannabinoid Agonist 5-MDMB-PICA. Brain Sci. 2020, 10, 1011. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10121011
Musa A, Simola N, Piras G, Caria F, Onaivi ES, De Luca MA. Neurochemical and Behavioral Characterization after Acute and Repeated Exposure to Novel Synthetic Cannabinoid Agonist 5-MDMB-PICA. Brain Sciences. 2020; 10(12):1011. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10121011
Chicago/Turabian StyleMusa, Aurora, Nicola Simola, Gessica Piras, Francesca Caria, Emmanuel Shan Onaivi, and Maria Antonietta De Luca. 2020. "Neurochemical and Behavioral Characterization after Acute and Repeated Exposure to Novel Synthetic Cannabinoid Agonist 5-MDMB-PICA" Brain Sciences 10, no. 12: 1011. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10121011