
Nucleus Accumbens Chemogenetic Inhibition Suppresses Amphetamine-Induced Ultrasonic Vocalizations in Male and Female Rats

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
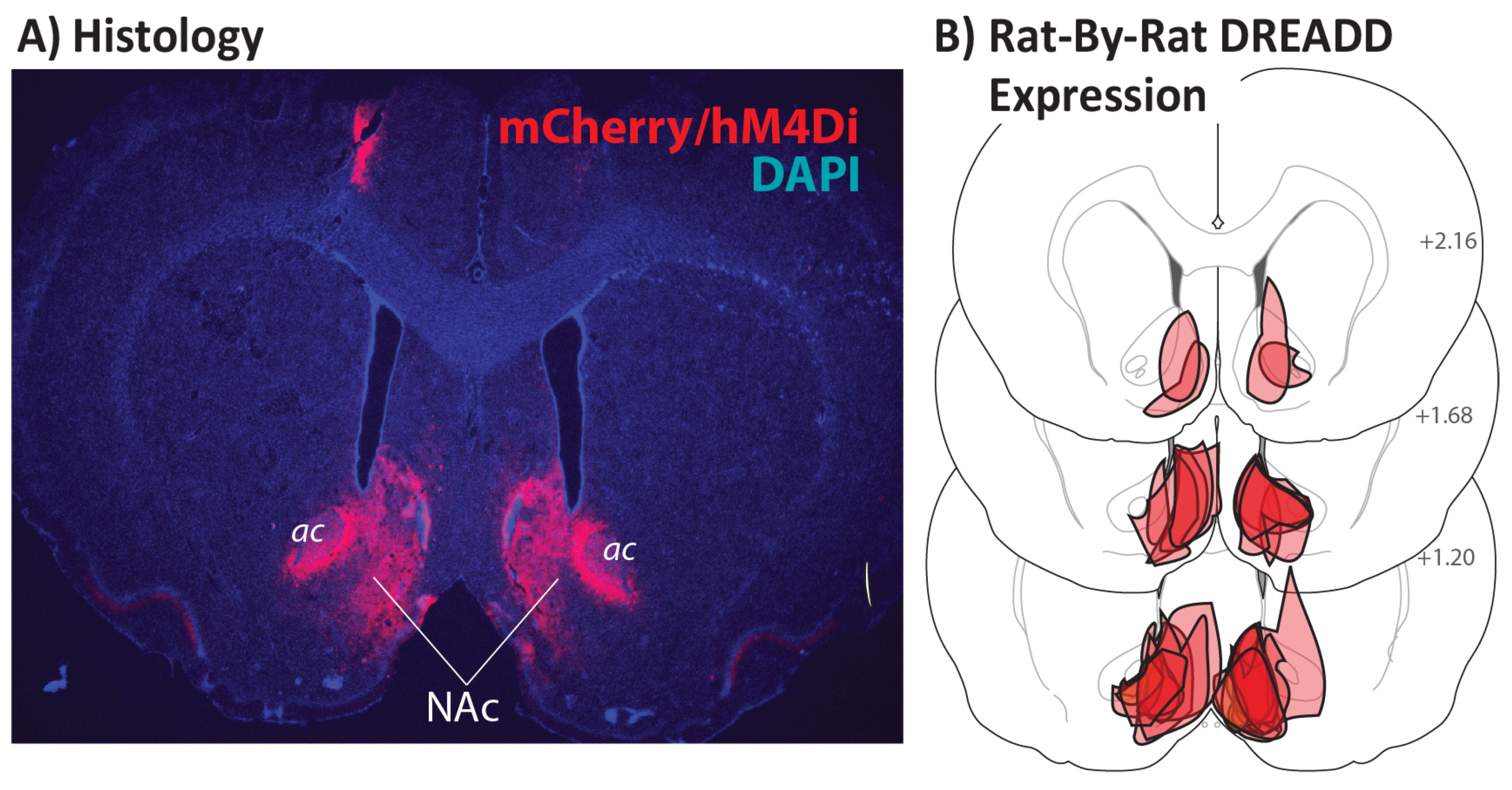
3.1. Localization of DREADD Expression
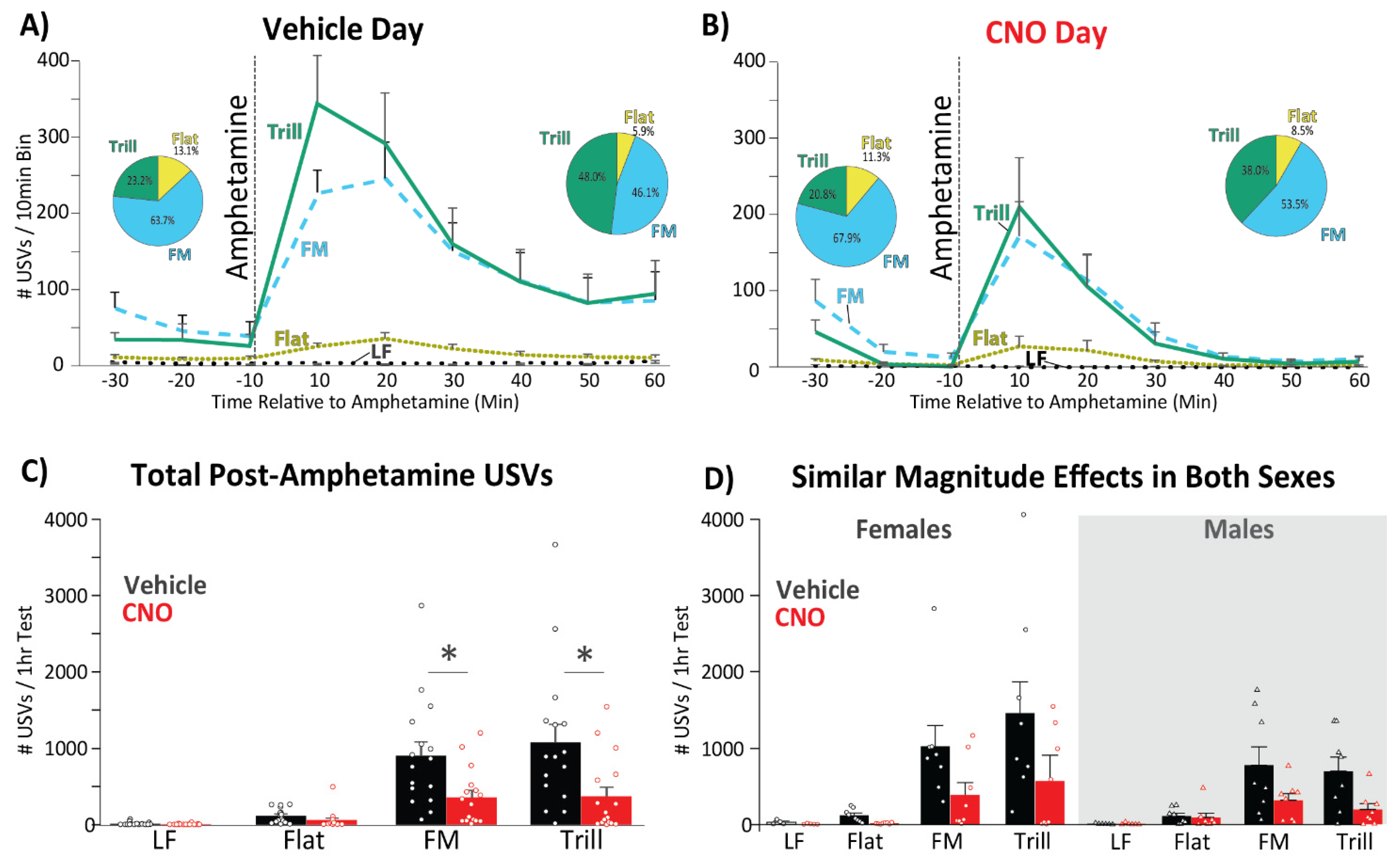
3.2. Amphetamine Increases the Number of High Frequency USVs Emitted in Both Sexes
3.3. CNO Effects on the Number of USVs Emitted Prior to Amphetamine
3.4. CNO Effects on the Number of Amphetamine-Induced USVs
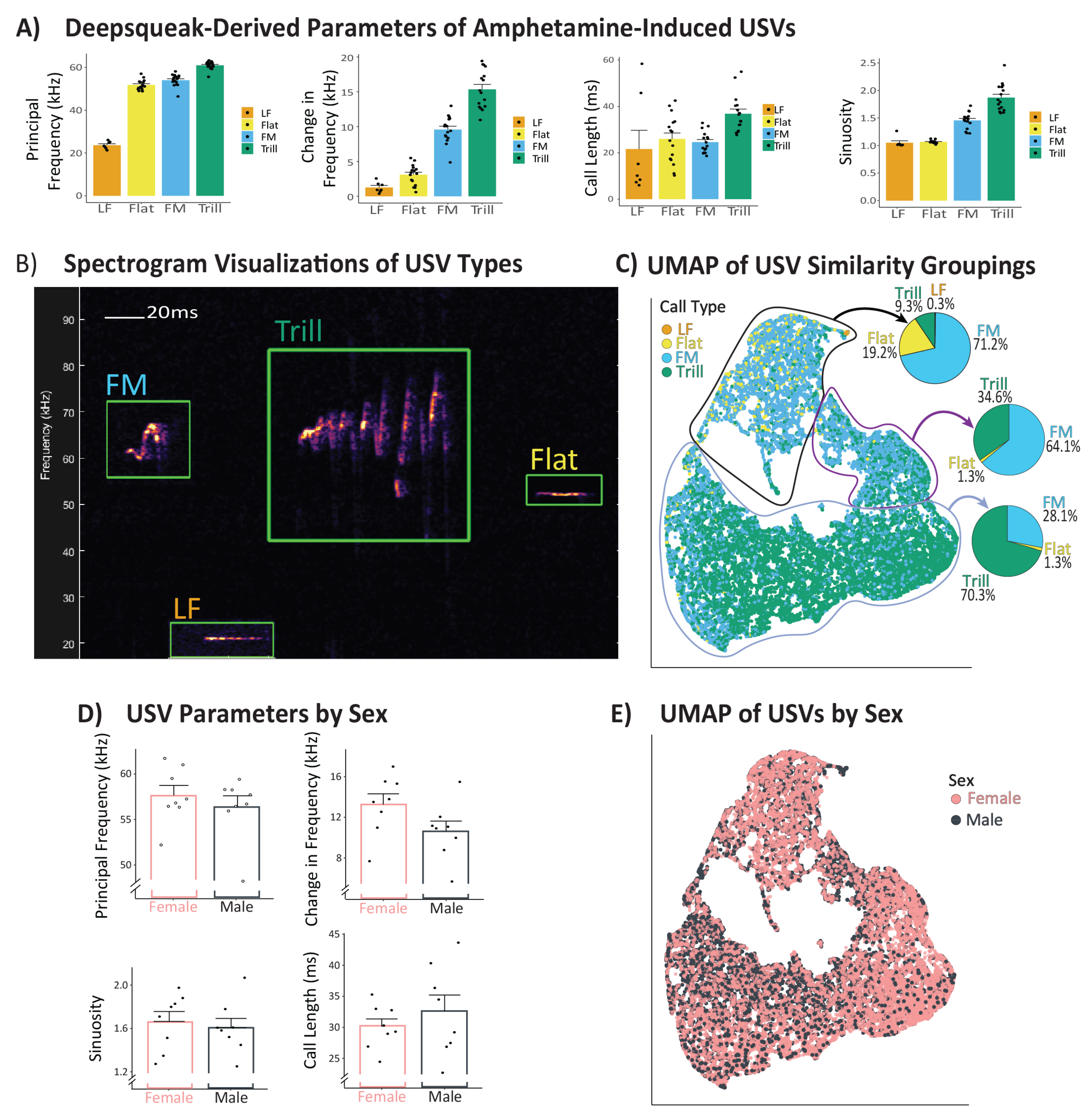
3.5. Amphetamine-Induced USV Characteristics, and Sex Differences
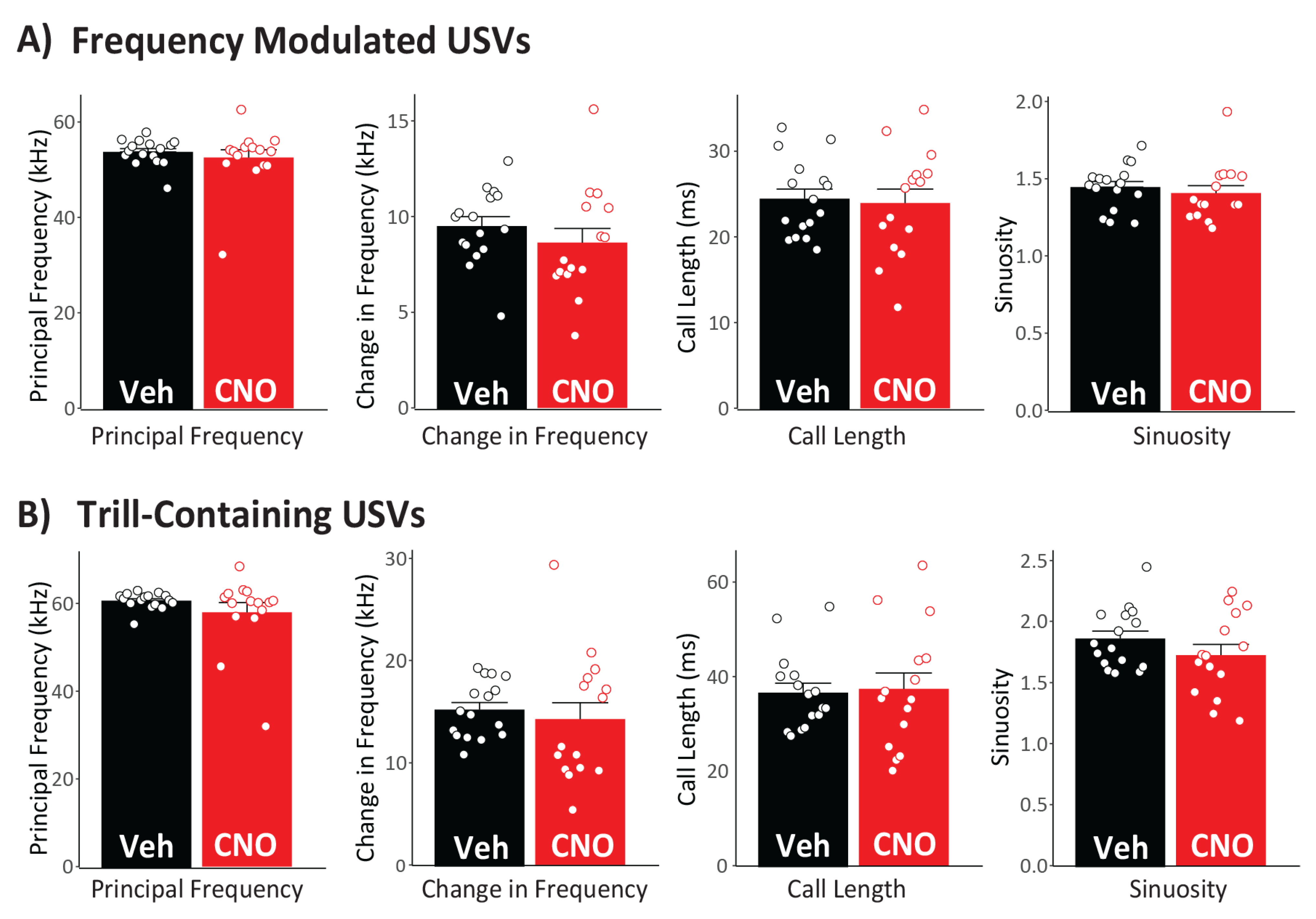
3.6. Effects of CNO on USV Characteristics
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Best, L.M.; Zhao, L.L.; Scardochio, T.; Clarke, P.B.S. Effects of repeated morphine on ultrasonic vocalizations in adult rats: Increased 50-kHz call rate and altered subtype profile. Psychopharmacology 2017, 234, 155–165. [Google Scholar] [CrossRef]
- Brudzynski, S.M. Ethotransmission: Communication of emotional states through ultrasonic vocalization in rats. Curr. Opin. Neurobiol. 2013, 23, 310–317. [Google Scholar] [CrossRef]
- Burgdorf, J.; Brudzynski, S.; Moskal, J. Using rat ultrasonic vocalization to study the neurobiology of emotion: From basic science to the development of novel therapeutics for affective disorders. Curr. Opin. Neurobiol. 2020, 60, 192–200. [Google Scholar] [CrossRef]
- Burgdorf, J.; Kroes, R.A.; Moskal, J.R.; Pfaus, J.G.; Brudzynski, S.M.; Panksepp, J. Ultrasonic vocalizations of rats (Rattus norvegicus) during mating, play, and aggression: Behavioral concomitants, relationship to reward, and self-administration of playback. J. Comp. Psychol. 2008, 122, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Engelhardt, K.-A.; Fuchs, E.; Schwarting, R.K.W.; Wöhr, M. Effects of amphetamine on pro-social ultrasonic communication in juvenile rats: Implications for mania models. Eur. Neuropsychopharmacol. 2017, 27, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Himmler, B.T.; Kisko, T.M.; Euston, D.R.; Kolb, B.; Pellis, S.M. Are 50-kHz calls used as play signals in the playful interactions of rats? I. Evidence from the timing and context of their use. Behav. Process. 2014, 106, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Kisko, T.M.; Himmler, B.T.; Himmler, S.M.; Euston, D.R.; Pellis, S.M. Are 50-kHz calls used as play signals in the playful interactions of rats? II. Evidence from the effects of devocalization. Behav. Process. 2015, 111, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Kisko, T.M.; Euston, D.R.; Pellis, S.M. Are 50-khz calls used as play signals in the playful interactions of rats? III. The effects of devocalization on play with unfamiliar partners as juveniles and as adults. Behav. Process. 2015, 113, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Wöhr, M.; Schwarting, R.K.W. Ultrasonic Communication in Rats: Can Playback of 50-kHz Calls Induce Approach Behavior? PLoS ONE 2007, 2, e1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seffer, D.; Schwarting, R.K.W.; Wöhr, M. Pro-social ultrasonic communication in rats: Insights from playback studies. J. Neurosci. Methods 2014, 234, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Tasaka, G.-I.; Feigin, L.; Maor, I.; Groysman, M.; DeNardo, L.A.; Schiavo, J.K.; Froemke, R.C.; Luo, L.; Mizrahi, A. The Temporal Association Cortex Plays a Key Role in Auditory-Driven Maternal Plasticity. Neuron 2020, 107, 566–579.e7. [Google Scholar] [CrossRef] [PubMed]
- Brudzynski, S.M. Communication of Adult Rats by Ultrasonic Vocalization: Biological, Sociobiological, and Neuroscience Approaches. ILAR J. 2009, 50, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Wöhr, M.; Schwarting, R.K.W. Affective communication in rodents: Ultrasonic vocalizations as a tool for research on emotion and motivation. Cell Tissue Res. 2013, 354, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, R.J.; Blanchard, D.C.; Agullana, R.; Weiss, S.M. Twenty-two kHz alarm cries to presentation of a predator, by laboratory rats living in visible burrow systems. Physiol. Behav. 1991, 50, 967–972. [Google Scholar] [CrossRef]
- Brudzynski, S.M.; Ociepa, D. Ultrasonic vocalization of laboratory rats in response to handling and touch. Physiol. Behav. 1992, 52, 655–660. [Google Scholar] [CrossRef]
- Borta, A.; Wöhr, M.; Schwarting, R.K.W. Rat ultrasonic vocalization in aversively motivated situations and the role of individual differences in anxiety-related behavior. Behav. Brain Res. 2006, 166, 271–280. [Google Scholar] [CrossRef]
- Chang, S.; Fan, Y.; Shin, J.H.; Ryu, Y.; Kim, M.S.; Steffensen, S.C.; Kim, H.K.; Kim, J.M.; Lee, B.H.; Jang, E.Y.; et al. Unpleasant Sound Elicits Negative Emotion and Reinstates Drug Seeking. Mol. Neurobiol. 2019, 56, 7594–7607. [Google Scholar] [CrossRef]
- Inagaki, H. Chapter 32—Sex Differences in Ultrasonic Vocal Expression of Negative Emotional States in Rats. In Handbook of Behavioral Neuroscience; Brudzynski, S.M., Ed.; Handbook of Ultrasonic Vocalization; Elsevier: Amsterdam, The Netherlands, 2018; Volume 25, pp. 337–344. [Google Scholar]
- Kroes, R.A.; Burgdorf, J.; Otto, N.J.; Panksepp, J.; Moskal, J.R. Social defeat, a paradigm of depression in rats that elicits 22-kHz vocalizations, preferentially activates the cholinergic signaling pathway in the periaqueductal gray. Behav. Brain Res. 2007, 182, 290–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgdorf, J.; Panksepp, J.; Moskal, J.R. Frequency-modulated 50 kHz ultrasonic vocalizations: A tool for uncovering the molecular substrates of positive affect. Neurosci. Biobehav. Rev. 2011, 35, 1831–1836. [Google Scholar] [CrossRef]
- Burgdorf, J.; Wood, P.L.; Kroes, R.A.; Moskal, J.R.; Panksepp, J. Neurobiology of 50-kHz ultrasonic vocalizations in rats: Electrode mapping, lesion, and pharmacology studies. Behav. Brain Res. 2007, 182, 274–283. [Google Scholar] [CrossRef]
- Hori, M.; Shimoju, R.; Tokunaga, R.; Ohkubo, M.; Miyabe, S.; Ohnishi, J.; Murakami, K.; Kurosawa, M. Tickling increases dopamine release in the nucleus accumbens and 50 kHz ultrasonic vocalizations in adolescent rats. NeuroReport 2013, 24, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Redecker, T.M.; Kisko, T.M.; Schwarting, R.K.W.; Wöhr, M. Effects of Cacna1c haploinsufficiency on social interaction behavior and 50-kHz ultrasonic vocalizations in adult female rats. Behav. Brain Res. 2019, 367, 35–52. [Google Scholar] [CrossRef] [PubMed]
- Mahler, S.V.; Moorman, D.E.; Feltenstein, M.W.; Cox, B.M.; Ogburn, K.B.; Bachar, M.; McGonigal, J.T.; Ghee, S.M.; See, R.E. A rodent “self-report” measure of methamphetamine craving? Rat ultrasonic vocalizations during methamphetamine self-administration, extinction, and reinstatement. Behav. Brain Res. 2013, 236, 78–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, M.; Andreatini, R.; Schwarting, R.K.W.; Brenes, J.C. Amphetamine-induced appetitive 50-kHz calls in rats: A marker of affect in mania? Psychopharmacology 2014, 231, 2567–2577. [Google Scholar] [CrossRef] [PubMed]
- Schwarting, R.K.W.; Jegan, N.; Wöhr, M. Situational factors, conditions and individual variables which can determine ultrasonic vocalizations in male adult Wistar rats. Behav. Brain Res. 2007, 182, 208–222. [Google Scholar] [CrossRef]
- Wright, J.M.; Gourdon, J.C.; Clarke, P.B.S. Identification of multiple call categories within the rich repertoire of adult rat 50-kHz ultrasonic vocalizations: Effects of amphetamine and social context. Psychopharmacology 2010, 211, 1–13. [Google Scholar] [CrossRef]
- Ahrens, A.M.; Ma, S.T.; Maier, E.Y.; Duvauchelle, C.L.; Schallert, T. Repeated intravenous amphetamine exposure: Rapid and persistent sensitization of 50-kHz ultrasonic trill calls in rats. Behav. Brain Res. 2009, 197, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Wright, J.M.; Dobosiewicz, M.R.S.; Clarke, P.B.S. α- and β-Adrenergic Receptors Differentially Modulate the Emission of Spontaneous and Amphetamine-Induced 50-kHz Ultrasonic Vocalizations in Adult Rats. Neuropsychopharmacology 2012, 37, 808–821. [Google Scholar] [CrossRef] [Green Version]
- Buck, C.L.; Vendruscolo, L.F.; Koob, G.F.; George, O. Dopamine D1 and μ-opioid receptor antagonism blocks anticipatory 50 kHz ultrasonic vocalizations induced by palatable food cues in Wistar rats. Psychopharmacology 2014, 231, 929–937. [Google Scholar] [CrossRef] [Green Version]
- Ciucci, M.R.; Ahrens, A.M.; Ma, S.T.; Kane, J.R.; Windham, E.B.; Woodlee, M.T.; Schallert, T. Reduction of dopamine synaptic activity: Degradation of 50-kHz ultrasonic vocalization in rats. Behav. Neurosci. 2009, 123, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Barker, D.J.; Herrera, C.; West, M.O. Automated Detection of 50-kHz Ultrasonic Vocalizations Using Template Matching in XBAT. J. Neurosci. Methods 2014, 236, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Coffey, K.R.; Marx, R.G.; Neumaier, J.F. DeepSqueak: A deep learning-based system for detection and analysis of ultrasonic vocalizations. Neuropsychopharmacology 2019, 44, 859–868. [Google Scholar] [CrossRef] [Green Version]
- Premoli, M.; Baggi, D.; Bianchetti, M.; Gnutti, A.; Bondaschi, M.; Mastinu, A.; Migliorati, P.; Signoroni, A.; Leonardi, R.; Memo, M.; et al. Automatic classification of mice vocalizations using Machine Learning techniques and Convolutional Neural Networks. PLoS ONE 2021, 16, e0244636. [Google Scholar] [CrossRef]
- Reno, J.M.; Marker, B.; Cormack, L.K.; Schallert, T.; Duvauchelle, C.L. Automating Ultrasonic Vocalization Analyses: The WAAVES Program. J. Neurosci. Methods 2013, 219, 155–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Segbroeck, M.; Knoll, A.T.; Levitt, P.; Narayanan, S. MUPET—Mouse Ultrasonic Profile ExTraction: A Signal Processing Tool for Rapid and Unsupervised Analysis of Ultrasonic Vocalizations. Neuron 2017, 94, 465–485.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zala, S.M.; Reitschmidt, D.; Noll, A.; Balazs, P.; Penn, D.J. Automatic mouse ultrasound detector (A-MUD): A new tool for processing rodent vocalizations. PLoS ONE 2017, 12, e0181200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamed, A.; Daszczuk, P.; Kursa, M.B.; Turzyńska, D.; Sobolewska, A.; Lehner, M.; Boguszewski, P.M.; Szyndler, J. Non-parametric analysis of neurochemical effects and Arc expression in amphetamine-induced 50-kHz ultrasonic vocalization. Behav. Brain Res. 2016, 312, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, K.G.; Brudzynski, S.M. Effect of microinjections of dopamine into the nucleus accumbens shell on emission of 50 kHz USV: Comparison with effects of d-amphetamine. Pharmacol. Biochem. Behav. 2019, 176, 23–32. [Google Scholar] [CrossRef]
- Simola, N.; Brudzynski, S.M. Rat 50-kHz ultrasonic vocalizations as a tool in studying neurochemical mechanisms that regulate positive emotional states. J. Neurosci. Methods 2018, 310, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Willuhn, I.; Tose, A.; Wanat, M.J.; Hart, A.S.; Hollon, N.G.; Phillips, P.E.M.; Schwarting, R.K.W.; Wöhr, M. Phasic dopamine release in the nucleus accumbens in response to pro-social 50 kHz ultrasonic vocalizations in rats. J. Neurosci. Off. J. Soc. Neurosci. 2014, 34, 10616–10623. [Google Scholar] [CrossRef]
- Armbruster, B.N.; Li, X.; Pausch, M.H.; Herlitze, S.; Roth, B.L. Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc. Natl. Acad. Sci. USA 2007, 104, 5163–5168. [Google Scholar] [CrossRef] [Green Version]
- Rogan, S.C.; Roth, B.L. Remote Control of Neuronal Signaling. Pharmacol. Rev. 2011, 63, 291–315. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.S.; Bucci, D.J.; Luikart, B.W.; Mahler, S.V. DREADDs: Use and Application in Behavioral Neuroscience. Behav. Neurosci. 2016, 130, 137–155. [Google Scholar] [CrossRef] [PubMed]
- National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press (US): Washington, DC, USA, 2011. [Google Scholar]
- Farrell, M.R.; Ruiz, C.M.; Castillo, E.; Faget, L.; Khanbijian, C.; Liu, S.; Schoch, H.; Rojas, G.; Huerta, M.Y.; Hnasko, T.S.; et al. Ventral pallidum is essential for cocaine relapse after voluntary abstinence in rats. Neuropsychopharmacology 2019, 44, 2174–2185. [Google Scholar] [CrossRef] [PubMed]
- McInnes, L.; Healy, J.; Melville, J. UMAP: Uniform Manifold Approximation and Projection for Dimension Reduction. arXiv 2020, arXiv:1802.03426. [Google Scholar]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates: Hard Cover Edition; Elsevier: Amsterdam, The Netherlands, 2006; p. 451. [Google Scholar]
- Becht, E.; McInnes, L.; Healy, J.; Dutertre, C.-A.; Kwok, I.W.H.; Ng, L.G.; Ginhoux, F.; Newell, E.W. Dimensionality reduction for visualizing single-cell data using UMAP. Nat. Biotechnol. 2019, 37, 38–44. [Google Scholar] [CrossRef]
- Bender, N.K.; Heilig, C.E.; Droll, B.; Wohlgemuth, J.; Armbruster, F.P.; Heilig, B. Immunogenicity, efficacy and adverse events of adalimumab in RA patients. Rheumatol. Int. 2007, 27, 269–274. [Google Scholar] [CrossRef]
- Dorrity, M.W.; Saunders, L.M.; Queitsch, C.; Fields, S.; Trapnell, C. Dimensionality reduction by UMAP to visualize physical and genetic interactions. Nat. Commun. 2020, 11, 1537. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Rico, M.; Alvarado, J.M. A Machine Learning Approach for Studying the Comorbidities of Complex Diagnoses. Behav. Sci. 2019, 9, 122. [Google Scholar] [CrossRef] [Green Version]
- Kisko, T.M.; Schwarting, R.K.W.; Wöhr, M. Sex differences in the acoustic features of social play-induced 50-kHz ultrasonic vocalizations: A detailed spectrographic analysis in wild-type Sprague–Dawley and Cacna1c haploinsufficient rats. Dev. Psychobiol. 2021, 63, 262–276. [Google Scholar] [CrossRef]
- Barker, D.J.; Root, D.H.; Ma, S.; Jha, S.; Megehee, L.; Pawlak, A.P.; West, M.O. Dose-dependent differences in short ultrasonic vocalizations emitted by rats during cocaine self-administration. Psychopharmacology 2010, 211, 435–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brudzynski, S.M.; Bihari, F.; Ociepa, D.; Fu, X.-W. Analysis of 22 kHz ultrasonic vocalization in laboratory rats: Long and short calls. Physiol. Behav. 1993, 54, 215–221. [Google Scholar] [CrossRef]
- Mahler, S.V.; Vazey, E.M.; Beckley, J.T.; Keistler, C.R.; McGlinchey, E.M.; Kaufling, J.; Wilson, S.P.; Deisseroth, K.; Woodward, J.J.; Aston-Jones, G. Designer receptors show role for ventral pallidum input to ventral tegmental area in cocaine seeking. Nat. Neurosci. 2014, 17, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Farrell, M.R.; Esteban, J.S.D.; Faget, L.; Floresco, S.B.; Hnasko, T.S.; Mahler, S.V. Ventral Pallidum GABA Neurons Mediate Motivation Underlying Risky Choice. J. Neurosci. 2021, 41, 4500–4513. [Google Scholar] [CrossRef] [PubMed]
- McGlinchey, E.M.; James, M.H.; Mahler, S.V.; Pantazis, C.; Aston-Jones, G. Prelimbic to Accumbens Core Pathway Is Recruited in a Dopamine-Dependent Manner to Drive Cued Reinstatement of Cocaine Seeking. J. Neurosci. 2016, 36, 8700–8711. [Google Scholar] [CrossRef] [Green Version]
- Thompson, B.; Leonard, K.C.; Brudzynski, S.M. Amphetamine-induced 50kHz calls from rat nucleus accumbens: A quantitative mapping study and acoustic analysis. Behav. Brain Res. 2006, 168, 64–73. [Google Scholar] [CrossRef]
- Wöhr, M. Measuring mania-like elevated mood through amphetamine-induced 50-kHz ultrasonic vocalizations in rats. Br. J. Pharmacol. 2021, 1–19. [Google Scholar] [CrossRef]
- Wintink, A.J.; Brudzynski, S.M. The related roles of dopamine and glutamate in the initiation of 50-kHz ultrasonic calls in adult rats. Pharmacol. Biochem. Behav. 2001, 70, 317–323. [Google Scholar] [CrossRef]
- Simola, N.; Fenu, S.; Costa, G.; Pinna, A.; Plumitallo, A.; Morelli, M. Pharmacological characterization of 50-kHz ultrasonic vocalizations in rats: Comparison of the effects of different psychoactive drugs and relevance in drug-induced reward. Neuropharmacology 2012, 63, 224–234. [Google Scholar] [CrossRef]
- Burgdorf, J.; Knutson, B.; Panksepp, J.; Ikemoto, S. Nucleus accumbens amphetamine microinjections unconditionally elicit 50-kHz ultrasonic vocalizations in rats. Behav. Neurosci. 2001, 115, 940–944. [Google Scholar] [CrossRef]
- Wright, J.M.; Dobosiewicz, M.R.S.; Clarke, P.B.S. The role of dopaminergic transmission through D1-like and D2-like receptors in amphetamine-induced rat ultrasonic vocalizations. Psychopharmacology 2013, 225, 853–868. [Google Scholar] [CrossRef]
- Brudzynski, S.M. Pharmacology of Ultrasonic Vocalizations in adult Rats: Significance, Call Classification and Neural Substrate. Curr. Neuropharmacol. 2015, 13, 180–192. [Google Scholar] [CrossRef] [Green Version]
- Ciucci, M.R.; Ma, S.T.; Kane, J.R.; Ahrens, A.M.; Schallert, T. Limb use and complex ultrasonic vocalization in a rat model of Parkinson’s disease: Deficit-targeted training. Parkinsonism Relat. Disord. 2008, 14, S172–S175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahler, S.V.; Brodnik, Z.D.; Cox, B.M.; Buchta, W.C.; Bentzley, B.S.; Quintanilla, J.; Cope, Z.A.; Lin, E.C.; Riedy, M.D.; Scofield, M.D.; et al. Chemogenetic Manipulations of Ventral Tegmental Area Dopamine Neurons Reveal Multifaceted Roles in Cocaine Abuse. J. Neurosci. 2019, 39, 503–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, D.J.; Simmons, S.J.; Servilio, L.C.; Bercovicz, D.; Ma, S.; Root, D.H.; Pawlak, A.P.; West, M.O. Ultrasonic Vocalizations: Evidence for an affective opponent process during cocaine self-administration. Psychopharmacology 2014, 231, 909–918. [Google Scholar] [CrossRef] [Green Version]
- Gomez, J.L.; Bonaventura, J.; Lesniak, W.; Mathews, W.B.; Sysa-Shah, P.; Rodriguez, L.A.; Ellis, R.J.; Richie, C.T.; Harvey, B.K.; Dannals, R.F.; et al. Chemogenetics revealed: DREADD occupancy and activation via converted clozapine. Science 2017, 357, 503–507. [Google Scholar] [CrossRef] [Green Version]
- MacLaren, D.A.A.; Browne, R.W.; Shaw, J.K.; Radhakrishnan, S.K.; Khare, P.; España, R.A.; Clark, S.D. Clozapine N-Oxide Administration Produces Behavioral Effects in Long–Evans Rats: Implications for Designing DREADD Experiments. eNeuro 2016, 3. [Google Scholar] [CrossRef] [PubMed]
- Mahler, S.V.; Aston-Jones, G. CNO Evil? Considerations for the Use of DREADDs in Behavioral Neuroscience. Neuropsychopharmacology 2018, 43, 934–936. [Google Scholar] [CrossRef]
- Manvich, D.F.; Webster, K.A.; Foster, S.L.; Farrell, M.S.; Ritchie, J.C.; Porter, J.H.; Weinshenker, D. The DREADD agonist clozapine N-oxide (CNO) is reverse-metabolized to clozapine and produces clozapine-like interoceptive stimulus effects in rats and mice. Sci. Rep. 2018, 8, 3840. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lawson, K.A.; Flores, A.Y.; Hokenson, R.E.; Ruiz, C.M.; Mahler, S.V. Nucleus Accumbens Chemogenetic Inhibition Suppresses Amphetamine-Induced Ultrasonic Vocalizations in Male and Female Rats. Brain Sci. 2021, 11, 1255. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11101255
Lawson KA, Flores AY, Hokenson RE, Ruiz CM, Mahler SV. Nucleus Accumbens Chemogenetic Inhibition Suppresses Amphetamine-Induced Ultrasonic Vocalizations in Male and Female Rats. Brain Sciences. 2021; 11(10):1255. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11101255
Chicago/Turabian StyleLawson, Kate A., Abigail Y. Flores, Rachael E. Hokenson, Christina M. Ruiz, and Stephen V. Mahler. 2021. "Nucleus Accumbens Chemogenetic Inhibition Suppresses Amphetamine-Induced Ultrasonic Vocalizations in Male and Female Rats" Brain Sciences 11, no. 10: 1255. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11101255