
Gene–Toxicant Interactions in Gulf War Illness: Differential Effects of the PON1 Genotype

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
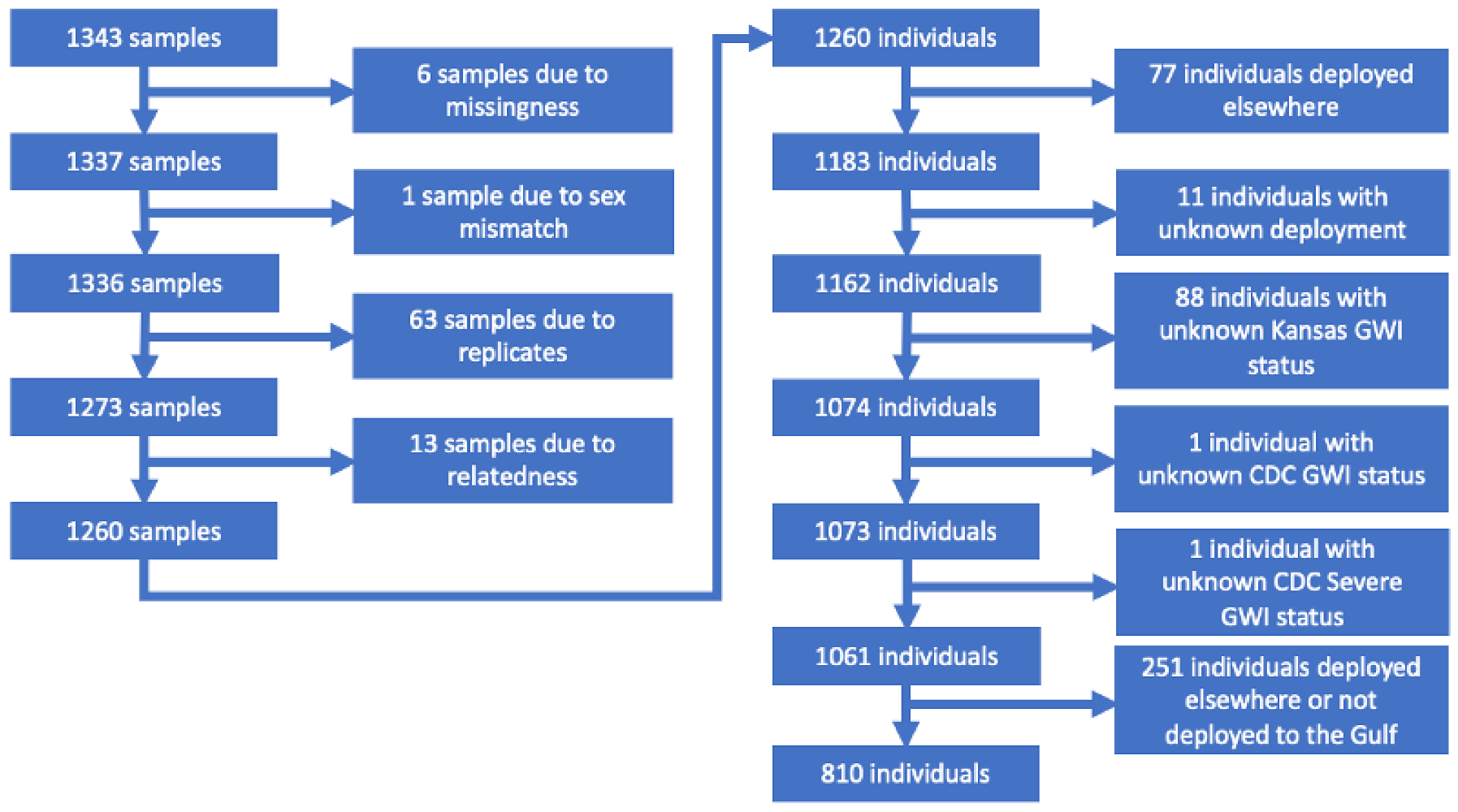
2.1. Introduction to GWECB
2.2. GWI Phenotypes
2.3. Gulf War Exposures
2.4. Genotypes and Genetic Data Cleaning
2.5. Statistical Analysis
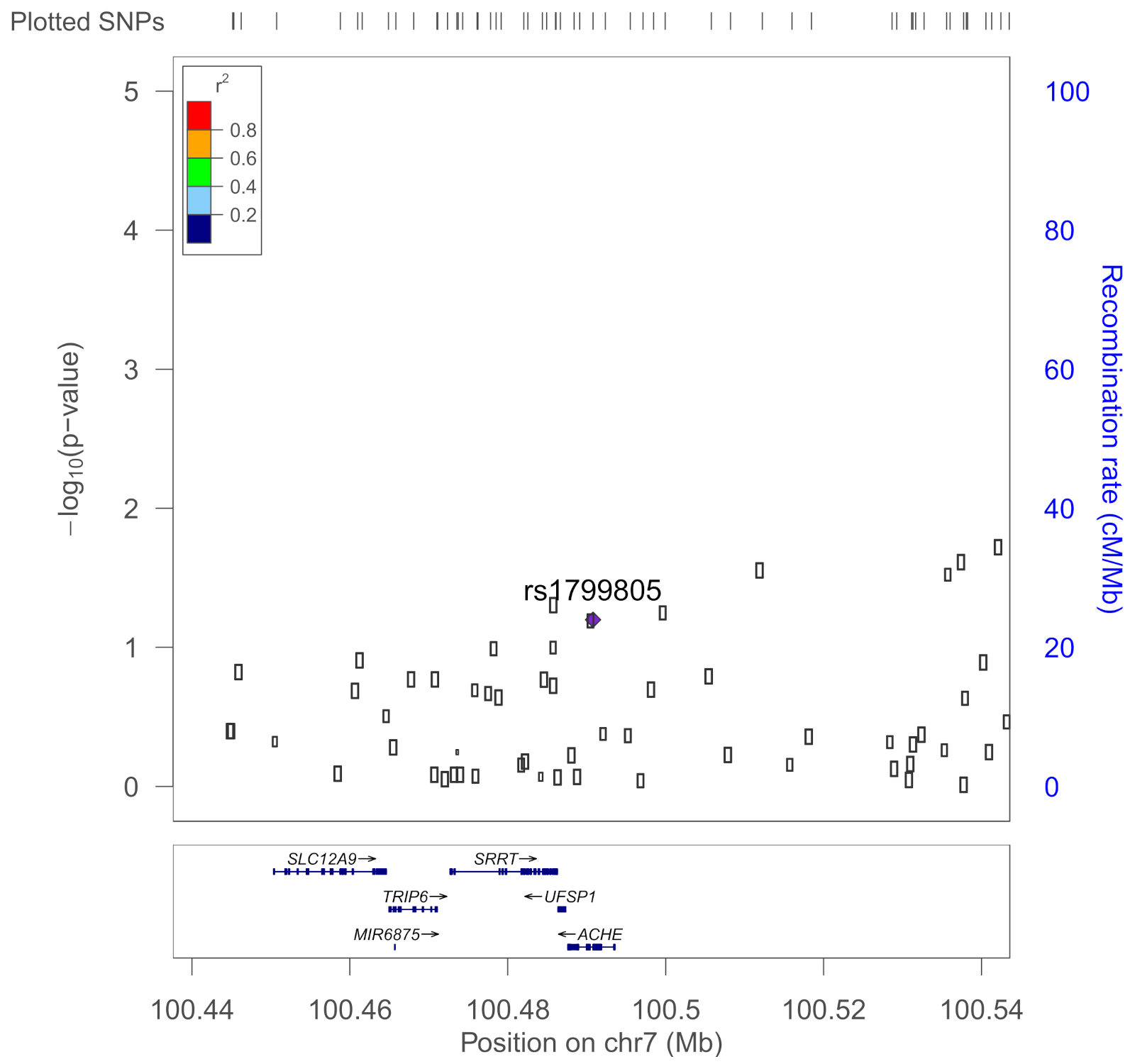
2.6. SNP Selection
3. Results
3.1. Demographics and Outcomes
3.2. Association of Tier 1 SNP/Exposure Interactions with Primary and Secondary Outcomes
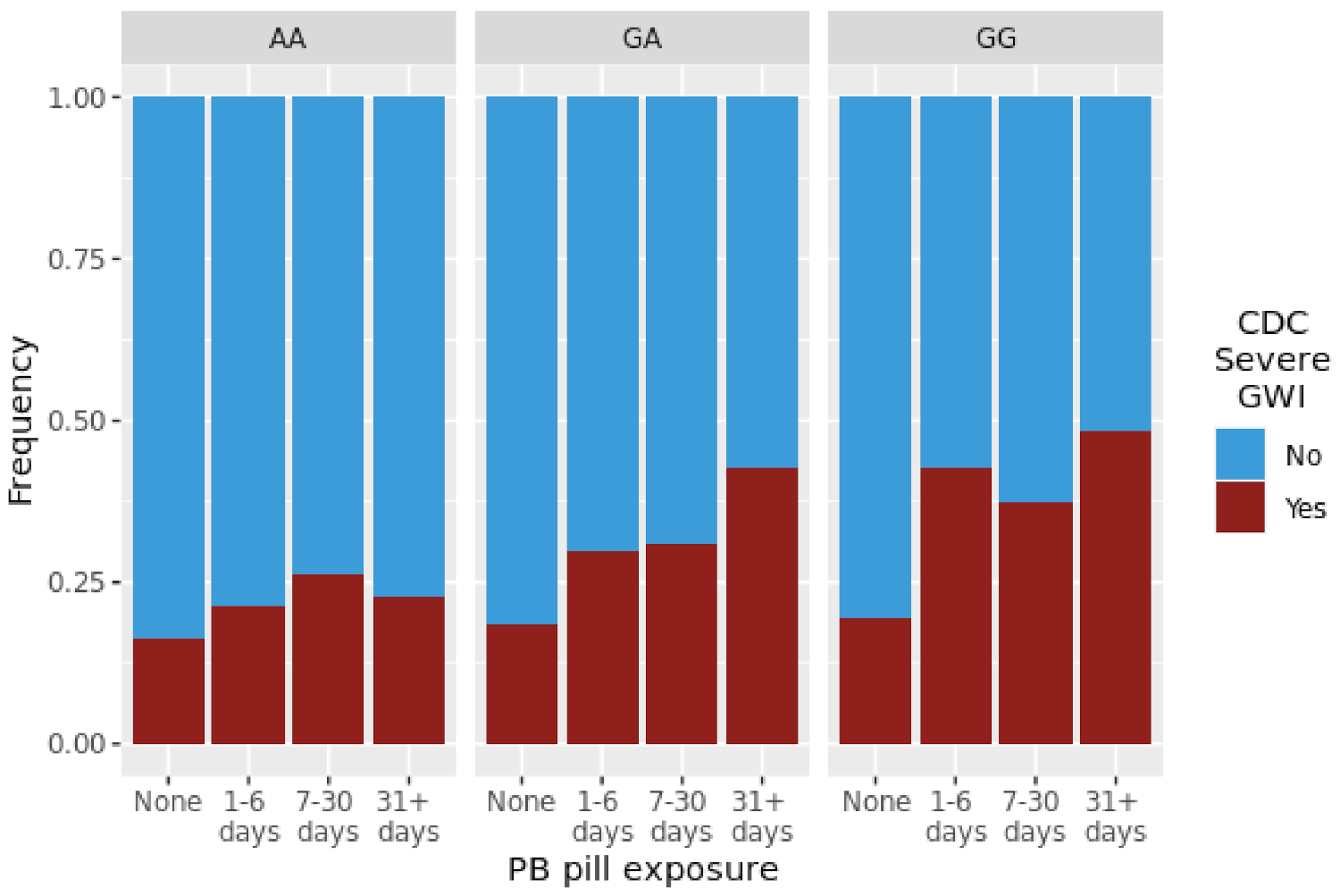
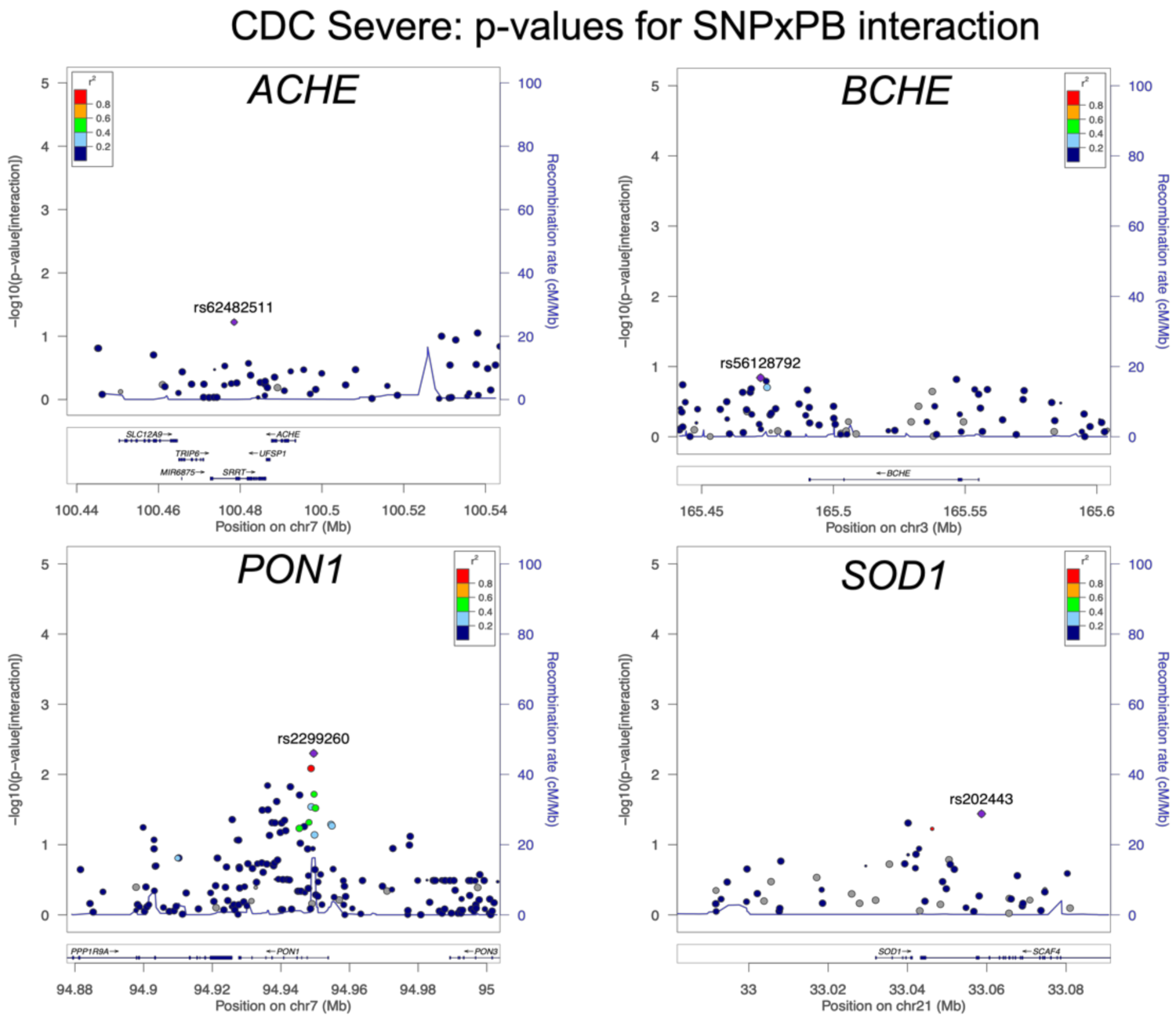
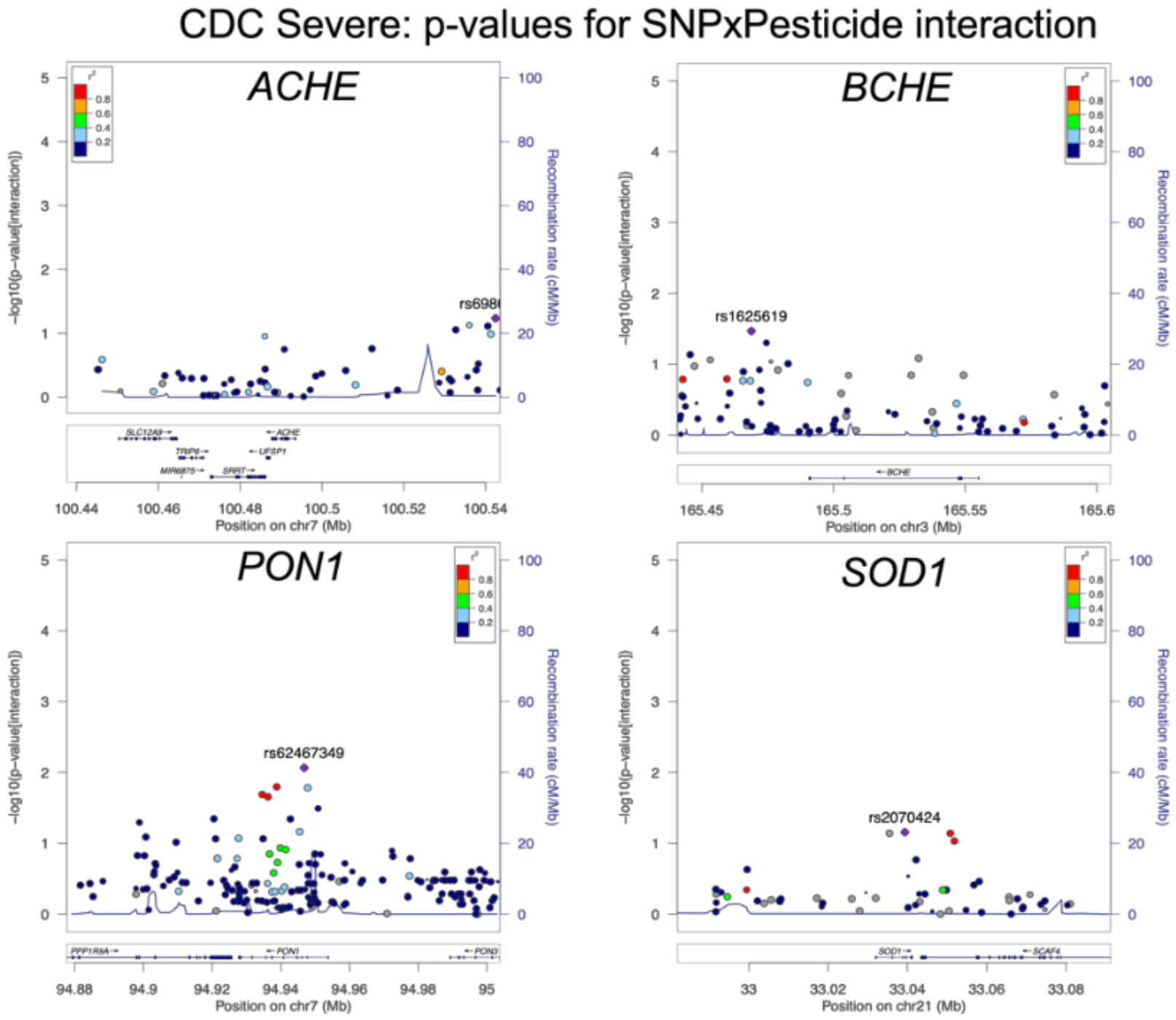
3.3. Association of Tier 2 SNP/Exposure interactions with CDC Severe GWI
3.4. Association of Tier 2 SNP/Exposure Interactions with Secondary Outcomes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Overall | Pesticide Exposure | |||||
|---|---|---|---|---|---|---|
| None | Missing | Short | Medium | Long | ||
| Count | 810 | 346 | 106 | 78 | 124 | 156 |
| CDC Severe: Fatigue Domain | 16% | 12% | 18% | 19% | 19% | 21% |
| CDC Severe: Mood Domain | 33% | 25% | 40% | 32% | 37% | 45% |
| CDC Severe: Musculoskeletal Domain | 33% | 24% | 46% | 37% | 36% | 41% |
| CDC Severe GWI | 26% | 19% | 35% | 24% | 30% | 33% |
| Kansas: Fatigue Domain | 80% | 70% | 86% | 81% | 86% | 92% |
| Kansas: Respiratory Domain | 36% | 25% | 43% | 47% | 40% | 45% |
| Kansas: Gastrointestinal Domain | 39% | 30% | 41% | 47% | 36% | 55% |
| Kansas: Pain Domain | 73% | 62% | 84% | 80% | 76% | 83% |
| Kansas: Skin Domain | 33% | 22% | 35% | 42% | 40% | 42% |
| Kansas: Mood Domain | 87% | 80% | 90% | 87% | 95% | 93% |
| Kansas Symptom Criteria | 72% | 59% | 81% | 78% | 77% | 89% |
| Kansas: Exclusionary Criterion | 40% | 40% | 44% | 46% | 39% | 38% |
| Overall | PB Pill Exposure | |||||
|---|---|---|---|---|---|---|
| None | Missing | Short | Medium | Long | ||
| Count | 810 | 311 | 138 | 98 | 114 | 149 |
| CDC Severe: Fatigue Domain | 16% | 9% | 20% | 22% | 17% | 24% |
| CDC Severe: Mood Domain | 33% | 24% | 37% | 34% | 41% | 44% |
| CDC Severe: Musculoskeletal Domain | 33% | 24% | 36% | 38% | 34% | 48% |
| CDC Severe GWI | 26% | 18% | 30% | 28% | 31% | 36% |
| Kansas: Fatigue Domain | 80% | 71% | 80% | 84% | 89% | 89% |
| Kansas: Respiratory Domain | 36% | 27% | 40% | 43% | 40% | 43% |
| Kansas: Gastrointestinal Domain | 39% | 28% | 38% | 40% | 52% | 51% |
| Kansas: Pain Domain | 73% | 61% | 80% | 77% | 77% | 87% |
| Kansas: Skin Domain | 33% | 24% | 33% | 40% | 38% | 41% |
| Kansas: Mood Domain | 87% | 79% | 87% | 90% | 93% | 94% |
| Kansas Symptom Criteria | 72% | 59% | 75% | 74% | 83% | 87% |
| Kansas: Exclusionary Criterion | 40% | 38% | 46% | 42% | 39% | 41% |











References
- Institute of Medicine. Chronic Multisymptom Illness in Gulf War Veterans: Case Definitions Reexamined; The National Academies Press: Washington, DC, USA, 2014. [Google Scholar]
- Kipen, H.M.; Hallman, W.; Kang, H.; Fiedler, N.; Natelson, B.H. Prevalence of Chronic Fatigue and Chemical Sensitivities in Gulf Registry Veterans. Arch. Environ. Health Int. J. 1999, 54, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, K.; Strauss, S.; Hickie, I.; Sharpe, M.; Dobbins, J.; Komaroff, A. The Chronic Fatigue Syndrome: A Comprehensive Approach to Its Definition and Study. J. Chronic Fatigue Syndr. 1995, 1, 67–84. [Google Scholar] [CrossRef]
- Fukuda, K.; Nisenbaum, R.; Stewart, G.; Thompson, W.W.; Robin, L.; Washko, R.M.; Noah, D.L.; Barrett, D.H.; Randall, B.; Herwaldt, B.L.; et al. Chronic Multisymptom Illness Affecting Air Force Veterans of the Gulf War. JAMA 1998, 280, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Steele, L. Prevalence and Patterns of Gulf War Illness in Kansas Veterans: Association of Symptoms with Characteristics of Person, Place, and Time of Military Service. Am. J. Epidemiol. 2000, 152, 992–1002. [Google Scholar] [CrossRef]
- Gifford, E.J.; Vahey, J.; Hauser, E.R.; Sims, K.J.; Efird, J.T.; Dursa, E.K.; Steele, L.; Helmer, D.A.; Provenzale, D. Gulf War illness in the Gulf War Era Cohort and Biorepository: The Kansas and Centers for Disease Control definitions. Life Sci. 2021, 278, 119454. [Google Scholar] [CrossRef]
- White, R.F.; Steele, L.; O’Callaghan, J.P.; Sullivan, K.; Binns, J.H.; Golomb, B.A.; Bloom, F.E.; Bunker, J.A.; Crawford, F.; Graves, J.C.; et al. Recent research on Gulf War illness and other health problems in veterans of the 1991 Gulf War: Effects of toxicant exposures during deployment. Cortex 2016, 74, 449–475. [Google Scholar] [CrossRef] [Green Version]
- Eisen, S.A.; Kang, H.K.; Murphy, F.M.; Blanchard, M.S.; Reda, D.J.; Henderson, W.G.; Toomey, R.; Jackson, L.W.; Alpern, R.; Parks, B.J.; et al. Gulf War Veterans’ Health: Medical Evaluation of a U.S. Cohort. Ann. Intern. Med. 2005, 142, 881–890. [Google Scholar] [CrossRef] [Green Version]
- Blanchard, M.S.; Eisen, S.A.; Alpern, R.; Karlinsky, J.; Toomey, R.; Reda, D.J.; Murphy, F.M.; Jackson, L.W.; Kang, H.K. Chronic Multisymptom Illness Complex in Gulf War I Veterans 10 Years Later. Am. J. Epidemiol. 2005, 163, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Smith, B.N.; Wang, J.M.; Vogt, D.; Vickers, K.; King, D.W.; King, L.A. Gulf War Illness. J. Occup. Environ. Med. 2013, 55, 104–110. [Google Scholar] [CrossRef]
- Cory-Slechta, D.A.; Wedge, R. Gulf War and Health. Gulf War Health 2016, 10, 1–292. [Google Scholar] [CrossRef]
- Updated Scientific Findings and Recommendations Research Advisory Committee on Gulf War Veterans’ Illnesses; Research Advisory Committee: Washington, DC, USA, 2014.
- Steele, L.; Lockridge, O.; Gerkovich, M.M.; Cook, M.R.; Sastre, A. Butyrylcholinesterase genotype and enzyme activity in relation to Gulf War illness: Preliminary evidence of gene-exposure interaction from a case–control study of 1991 Gulf War veterans. Environ. Health 2015, 14, 4. [Google Scholar] [CrossRef] [Green Version]
- Haley, R.W.; Billecke, S.; La Du, B.N. Association of Low PON1 Type Q (Type A) Arylesterase Activity with Neurologic Symptom Complexes in Gulf War Veterans. Toxicol. Appl. Pharmacol. 1999, 157, 227–233. [Google Scholar] [CrossRef]
- Abdullah, L.; Evans, J.E.; Montague, H.; Reed, J.M.; Moser, A.; Crynen, G.; Gonzalez, A.; Zakirova, Z.; Ross, I.; Mullan, C.; et al. Chronic elevation of phosphocholine containing lipids in mice exposed to Gulf War agents pyridostigmine bromide and permethrin. Neurotoxicol. Teratol. 2013, 40, 74–84. [Google Scholar] [CrossRef]
- Abdel-Rahman, A.; Abou-Donia, S.M.; El-Masry, E.M.; Shetty, A.; Abou-Donia, M.B. Stress and Combined Exposure to Low Doses of Pyridostigmine Bromide, DEET, and Permethrin Produce Neurochemical and Neuropathological Alterations in Cerebral Cortex, Hippocampus, and Cerebellum. J. Toxicol. Environ. Health Part A 2004, 67, 163–192. [Google Scholar] [CrossRef]
- Ashbrook, D.G.; Hing, B.; Michalovicz, L.T.; Kelly, K.A.; Miller, J.V.; De Vega, W.C.; Miller, D.B.; Broderick, G.; O’Callaghan, J.P.; McGowan, P.O. Epigenetic impacts of stress priming of the neuroinflammatory response to sarin surrogate in mice: A model of Gulf War illness. J. Neuroinflamm. 2018, 15, 86. [Google Scholar] [CrossRef] [Green Version]
- Nettleman, M. Gulf War Illness: Challenges Persist. Trans. Am. Clin. Climatol. Assoc. 2015, 126, 237–247. [Google Scholar]
- Emmerich, T.; Zakirova, Z.; Klimas, N.; Sullivan, K.; Shetty, A.K.; Evans, J.E.; Ait-Ghezala, G.; Laco, G.S.; Hattiangady, B.; Shetty, G.A.; et al. Phospholipid profiling of plasma from GW veterans and rodent models to identify potential biomarkers of Gulf War Illness. PLoS ONE 2017, 12, e0176634. [Google Scholar] [CrossRef]
- Chao, L.; Abadjian, L.R.; Esparza, I.L.; Reeb, R. Insomnia Severity, Subjective Sleep Quality, and Risk for Obstructive Sleep Apnea in Veterans With Gulf War Illness. Mil. Med. 2016, 181, 1127–1134. [Google Scholar] [CrossRef] [Green Version]
- Belgrad, J.; Dutta, D.J.; Bromley-Coolidge, S.; Kelly, K.A.; Michalovicz, L.T.; Sullivan, K.A.; O’Callaghan, J.P.; Fields, R.D. Oligodendrocyte involvement in Gulf War Illness. Glia 2019, 67, 2107–2124. [Google Scholar] [CrossRef] [Green Version]
- Gulf War Illness and the Health of Gulf War Veterans; Research Advisory Committee: Washington, DC, USA, 2008.
- Ojo, J.O.; Abdullah, L.; Evans, J.; Reed, J.M.; Montague, H.; Mullan, M.J.; Crawford, F.C. Exposure to an organophosphate pesticide, individually or in combination with other Gulf War agents, impairs synaptic integrity and neuronal differentiation, and is accompanied by subtle microvascular injury in a mouse model of Gulf War agent exposure. Neuropathology 2014, 34, 109–127. [Google Scholar] [CrossRef]
- Miller, J.V.; LeBouf, R.; A Kelly, K.; Michalovicz, L.T.; Ranpara, A.; Locker, A.R.; Miller, D.B.; O’Callaghan, J.P. The Neuroinflammatory Phenotype in a Mouse Model of Gulf War Illness is Unrelated to Brain Regional Levels of Acetylcholine as Measured by Quantitative HILIC-UPLC-MS/MS. Toxicol. Sci. 2018, 165, 302–313. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, J.; Gordon, H.; Ludwig, H.; Wagner, J.; Harn, D.; Norberg, T.; Filipov, N. Neurochemical and neuroinflammatory perturbations in two Gulf War Illness models: Modulation by the immunotherapeutic LNFPIII. NeuroToxicology 2020, 77, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Zakirova, Z.; Tweed, M.; Crynen, G.; Reed, J.; Abdullah, L.; Nissanka, N.; Mullan, M.; Mullan, M.J.; Mathura, V.; Crawford, F.; et al. Gulf War Agent Exposure Causes Impairment of Long-Term Memory Formation and Neuropathological Changes in a Mouse Model of Gulf War Illness. PLoS ONE 2015, 10, e0119579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, S.; Fried, D.E.; Grubišić, V.; McClain, J.L.; Gulbransen, B.D. Gastrointestinal neuroimmune disruption in a mouse model of Gulf War illness. FASEB J. 2019, 33, 6168–6184. [Google Scholar] [CrossRef] [PubMed]
- Koo, B.-B.; Michalovicz, L.T.; Calderazzo, S.; Kelly, K.; Sullivan, K.; Killiany, R.J.; O’Callaghan, J.P. Corticosterone potentiates DFP-induced neuroinflammation and affects high-order diffusion imaging in a rat model of Gulf War Illness. Brain Behav. Immun. 2018, 67, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Michalovicz, L.T.; Kelly, K.A.; Sullivan, K.; O’Callaghan, J.P. Acetylcholinesterase inhibitor exposures as an initiating factor in the development of Gulf War Illness, a chronic neuroimmune disorder in deployed veterans. Neuropharmacology 2020, 171, 108073. [Google Scholar] [CrossRef]
- Mackness, B.; Durrington, P.N.; Mackness, M.I. Low Paraoxonase in Persian Gulf War Veterans Self-Reporting Gulf War Syndrome. Biochem. Biophys. Res. Commun. 2000, 276, 729–733. [Google Scholar] [CrossRef]
- Saccon, R.A.; Bunton-Stasyshyn, R.K.A.; Fisher, E.M.; Fratta, P. Is SOD1 loss of function involved in amyotrophic lateral sclerosis? Brain 2013, 136, 2342–2358. [Google Scholar] [CrossRef]
- Haley, R.W. Excess incidence of ALS in young Gulf War veterans. Neurology 2003, 61, 750–756. [Google Scholar] [CrossRef]
- Horner, R.; Kamins, K.; Feussner, J.; Grambow, S.; Hoff-Lindquist, J.; Harati, Y.; Mitsumoto, H.; Pascuzzi, R.; Spencer, P.; Tim, R.; et al. Occurrence of amyotrophic lateral sclerosis among Gulf War veterans. Neurology 2003, 61, 742–749. [Google Scholar] [CrossRef]
- Merwin, S.J.; Obis, T.; Nunez, Y.; Re, D.B. Organophosphate neurotoxicity to the voluntary motor system on the trail of environment-caused amyotrophic lateral sclerosis: The known, the misknown, and the unknown. Arch. Toxicol. 2017, 91, 2939–2952. [Google Scholar] [CrossRef]
- A van Es, M.; Dahlberg, C.; Birve, A.; Veldink, J.H.; Berg, L.H.V.D.; Andersen, P.M. Large-scale SOD1 mutation screening provides evidence for genetic heterogeneity in amyotrophic lateral sclerosis. J. Neurol. Neurosurg. Psychiatry 2010, 81, 562–566. [Google Scholar] [CrossRef]
- Zhu, G.-D.; Dawson, E.; Huskey, A.; Gordon, R.J.; Del Tredici, A.L. Genetic Testing for BCHE Variants Identifies Patients at Risk of Prolonged Neuromuscular Blockade in Response to Succinylcholine. Pharmacogenomics Pers. Med. 2020, 13, 405–414. [Google Scholar] [CrossRef]
- Costa, L.G.; Cole, T.B.; Jarvik, G.P.; Furlong, C.E. Functional Genomics of the Paraoxonase (PON1) Polymorphisms: Effects on Pesticide Sensitivity, Cardiovascular Disease, and Drug Metabolism. Annu. Rev. Med. 2003, 54, 371–392. [Google Scholar] [CrossRef]
- Slowik, A.; Tomik, B.; Wolkow, P.P.; Partyka, D.; Turaj, W.; Malecki, M.T.; Pera, J.; Dziedzic, T.; Szczudlik, A.; Figlewicz, D.A. Paraoxonase gene polymorphisms and sporadic ALS. Neurology 2006, 67, 766–770. [Google Scholar] [CrossRef]
- Verde, F.; Tiloca, C.; Morelli, C.; Doretti, A.; Poletti, B.; Maderna, L.; Messina, S.; Gentilini, D.; Fogh, I.; Ratti, A.; et al. PON1 is a disease modifier gene in amyotrophic lateral sclerosis: Association of the Q192R polymorphism with bulbar onset and reduced survival. Neurol. Sci. 2019, 40, 1469–1473. [Google Scholar] [CrossRef]
- Huo, X.; Guo, Y.; Zhang, Y.; Li, J.; Wen, X.; Liu, J. Paraoxonase 1 gene (Q192R) polymorphism confers susceptibility to coronary artery disease in type 2 diabetes patients: Evidence from case-control studies. Drug Discov. Ther. 2019, 13, 80–88. [Google Scholar] [CrossRef] [Green Version]
- Jarvik, G.P.; Hatsukami, T.S.; Carlson, C.; Richter, R.J.; Jampsa, R.; Brophy, V.H.; Margolin, S.; Rieder, M.; Nickerson, D.; Schellenberg, G.D.; et al. Paraoxonase Activity, But Not Haplotype Utilizing the Linkage Disequilibrium Structure, Predicts Vascular Disease. Arter. Thromb. Vasc. Biol. 2003, 23, 1465–1471. [Google Scholar] [CrossRef] [Green Version]
- Jarvik, G.P.; Jampsa, R.; Richter, R.J.; Carlson, C.S.; Rieder, M.J.; Nickerson, D.A.; Furlong, C.E. Novel paraoxonase (PON1) nonsense and missense mutations predicted by functional genomic assay of PON1 status. Pharmacogenetics Genom. 2003, 13, 291–295. [Google Scholar] [CrossRef]
- Shapira, M.; Tur-Kaspa, I.; Bosgraaf, L.; Livni, N.; Grant, A.D.; Grisaru, D.; Korner, M.; Ebstein, R.P.; Soreq, H. A transcription-activating polymorphism in the ACHE promoter associated with acute sensitivity to anti-acetylcholinesterases. Hum. Mol. Genet. 2000, 9, 1273–1281. [Google Scholar] [CrossRef] [Green Version]
- Khalil, L.; McNeil, R.B.; Sims, K.J.; A Felder, K.; Hauser, E.R.; Goldstein, K.M.; I Voils, C.; Klimas, N.G.; Brophy, M.T.; Thomas, C.M.; et al. The Gulf War Era Cohort and Biorepository: A Longitudinal Research Resource of Veterans of the 1990–1991 Gulf War Era. Am. J. Epidemiol. 2018, 187, 2279–2291. [Google Scholar] [CrossRef]
- Vahey, J.; Hauser, E.R.; Sims, K.J.; Helmer, D.A.; Provenzale, D.; Gifford, E.J. Research tool for classifying Gulf War illness using survey responses: Lessons for writing replicable algorithms for symptom-based conditions. Life Sci. 2021, 282, 119808. [Google Scholar] [CrossRef]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.A.M.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience 2015, 4, 7. [Google Scholar] [CrossRef]
- Chen, Y.; Meyer, J.; Hill, H.Z.; Lange, G.; Condon, M.R.; Klein, J.C.; Ndirangu, D.; Falvo, M.J. Role of mitochondrial DNA damage and dysfunction in veterans with Gulf War Illness. PLoS ONE 2017, 12, e0184832. [Google Scholar] [CrossRef] [Green Version]
- Pruim, R.J.; Welch, R.P.; Sanna, S.; Teslovich, T.M.; Chines, P.S.; Gliedt, T.P.; Boehnke, M.; Abecasis, G.; Willer, C.J. LocusZoom: Regional visualization of genome-wide association scan results. Bioinformatics 2010, 26, 2336–2337. [Google Scholar] [CrossRef] [Green Version]
- Goodall, R. Cholinesterase: Phenotyping and genotyping. Ann. Clin. Biochem. Int. J. Lab. Med. 2004, 41, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Dardiotis, E.; Aloizou, A.-M.; Siokas, V.; Tsouris, Z.; Rikos, D.; Marogianni, C.; Aschner, M.; Kovatsi, L.; Bogdanos, D.P.; Tsatsakis, A. Paraoxonase-1 genetic polymorphisms in organophosphate metabolism. Toxicology 2019, 411, 24–31. [Google Scholar] [CrossRef]
- Davies, H.G.; Richter, R.J.; Keifer, M.; Broomfield, C.A.; Sowal, J.; Furlong, C.E. The Effect of the Human Serum Paraoxonase Polymorphism is Reversed with Diazoxon, Soman and Sarin. Nat. Genet. 1996, 14, 334–336. [Google Scholar] [CrossRef] [PubMed]
- Machiela, M.J.; Chanock, S.J. LDlink: A web-based application for exploring population-specific haplotype structure and linking correlated alleles of possible functional variants: Fig. Bioinformatics 2015, 31, 3555–3557. [Google Scholar] [CrossRef] [PubMed]
- Furlong, C.E.; Marsillach, J.; Jarvik, G.P.; Costa, L.G. Paraoxonases-1, -2 and -3: What are their functions? Chem. Interact. 2016, 259, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Buniello, A.; MacArthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E.; et al. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics. Nucleic Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef] [Green Version]
- Sun, B.B.; Maranville, J.C.; Peters, J.E.; Stacey, D.; Staley, J.R.; Blackshaw, J.; Burgess, S.; Jiang, T.; Paige, E.; Surendran, P.; et al. Genomic atlas of the human plasma proteome. Nature 2018, 558, 73–79. [Google Scholar] [CrossRef]
- Marees, A.T.; De Kluiver, H.; Stringer, S.; Vorspan, F.; Curis, E.; Marie-Claire, C.; Derks, E. A tutorial on conducting genome-wide association studies: Quality control and statistical analysis. Int. J. Methods Psychiatr. Res. 2018, 27, e1608. [Google Scholar] [CrossRef]
- Radhakrishnan, K.; Hauser, E.; Polimanti, R.; Helmer, D.; Provenzale, D.; McNeil, R.; Maffucci, A.; Quaden, R.; Zhao, H.; Whitbourne, S.; et al. Genomics of Gulf War Illness in U.S. Veterans Who Served during the 1990–1991 Persian Gulf War: Methods and Rationale for Veterans Affairs Cooperative Study #2006. Brain Sci. 2021, 11, 845. [Google Scholar] [CrossRef]
- Parihar, V.K.; Hattiangady, B.; Shuai, B.; Shetty, A.K. Mood and Memory Deficits in a Model of Gulf War Illness Are Linked with Reduced Neurogenesis, Partial Neuron Loss, and Mild Inflammation in the Hippocampus. Neuropsychopharmacology 2013, 38, 2348–2362. [Google Scholar] [CrossRef]
- Golomb, B.A. Acetylcholinesterase inhibitors and Gulf War illnesses. Proc. Natl. Acad. Sci. USA 2008, 105, 4295–4300. [Google Scholar] [CrossRef] [Green Version]
- Janulewicz, P.; Krengel, M.; Quinn, E.; Heeren, T.; Toomey, R.; Killiany, R.; Zundel, C.; Ajama, J.; O’Callaghan, J.; Steele, L.; et al. The Multiple Hit Hypothesis for Gulf War Illness: Self-Reported Chemical/Biological Weapons Exposure and Mild Traumatic Brain Injury. Brain Sci. 2018, 8, 198. [Google Scholar] [CrossRef] [Green Version]
- Phillips, K.F.; Deshpande, L.S. Repeated low-dose organophosphate DFP exposure leads to the development of depression and cognitive impairment in a rat model of Gulf War Illness. NeuroToxicology 2016, 52, 127–133. [Google Scholar] [CrossRef]
- Haley, R.W. Self-reported Exposure to Neurotoxic Chemical Combinations in the Gulf WarA Cross-sectional Epidemiologic Study. JAMA 1997, 277, 231–237. [Google Scholar] [CrossRef]




| Tier 1 SNPs: BCHE, PON1, and ACHE from Prior Studies | ||||
|---|---|---|---|---|
| SNP | Gene | Variant | MAF | Citations |
| rs1799807 | BCHE | Atypical (A); lower catalytic rate; succinylcholine susceptibility | 0.020 | Zhu et al., 2020 [36], Goodall 2004 [49], Steele et al., 2015 [13] |
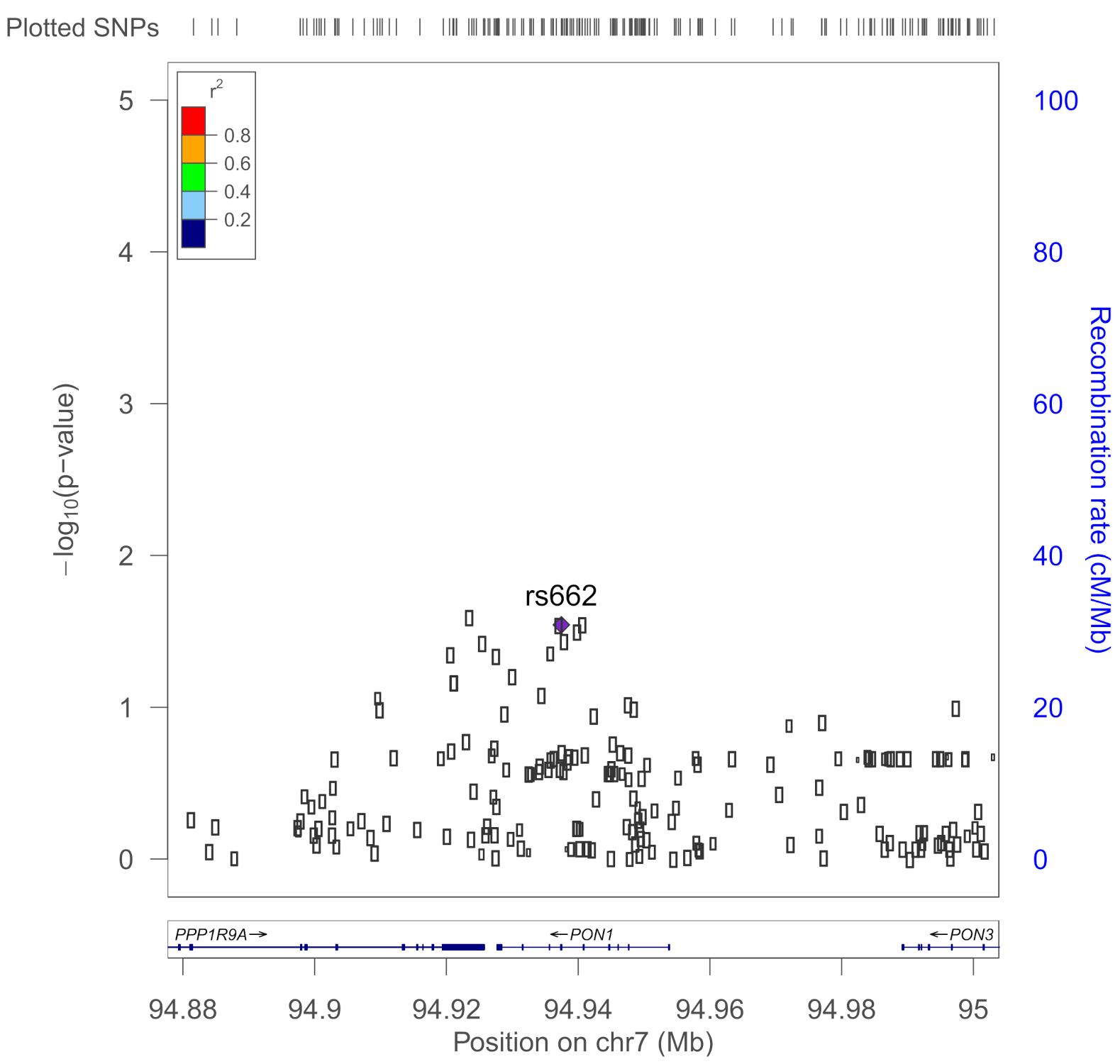
| rs662 | PON1 | 192Q/R; functional variant of Pon1 Modifier for risk of sporadic ALS | 0.396 | Dardiotis et al., 2018 [50], Davies et al., 1996 [51], Haley et al., 1999 [14]; Verde et al., 2019 [39] |
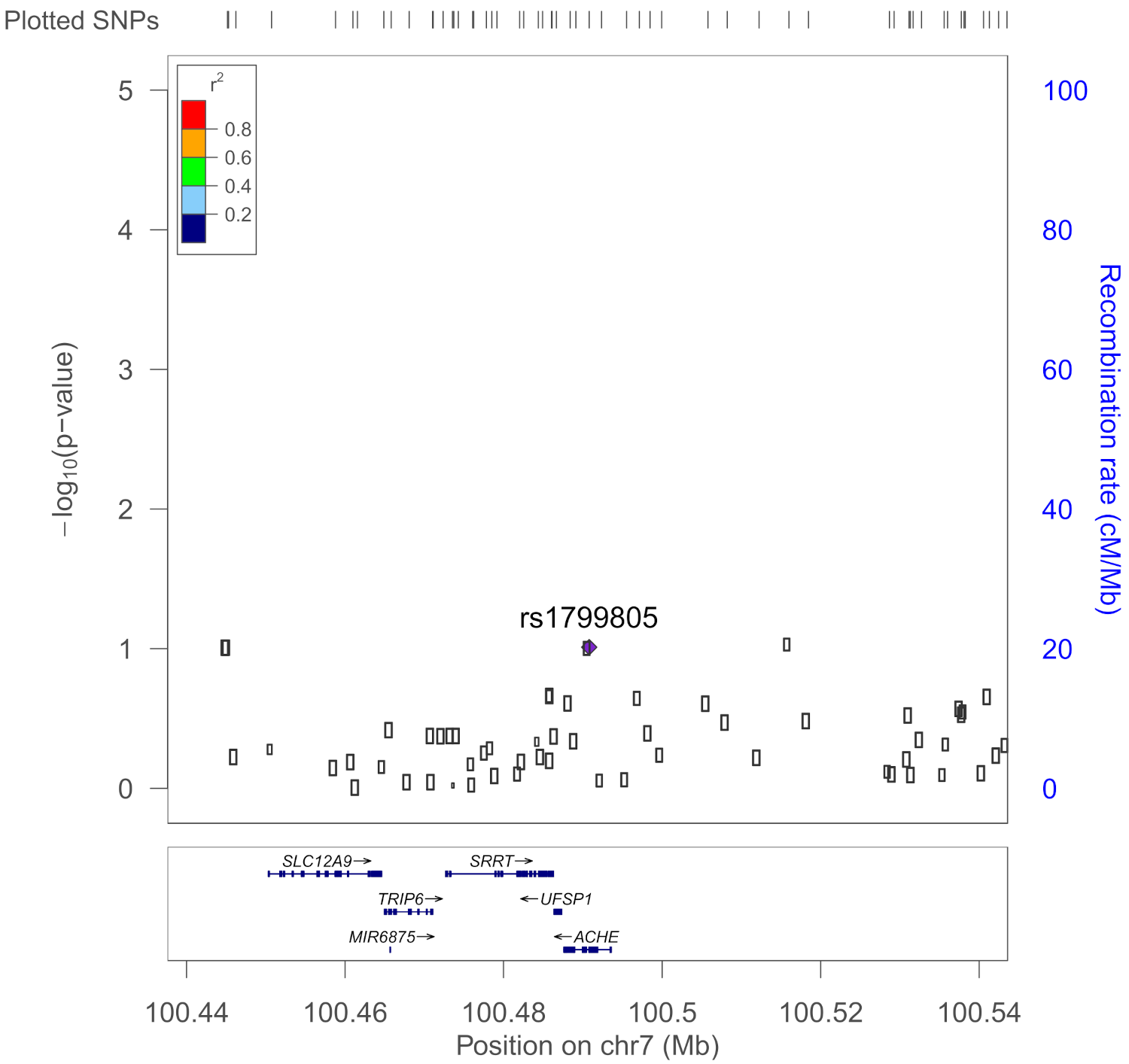
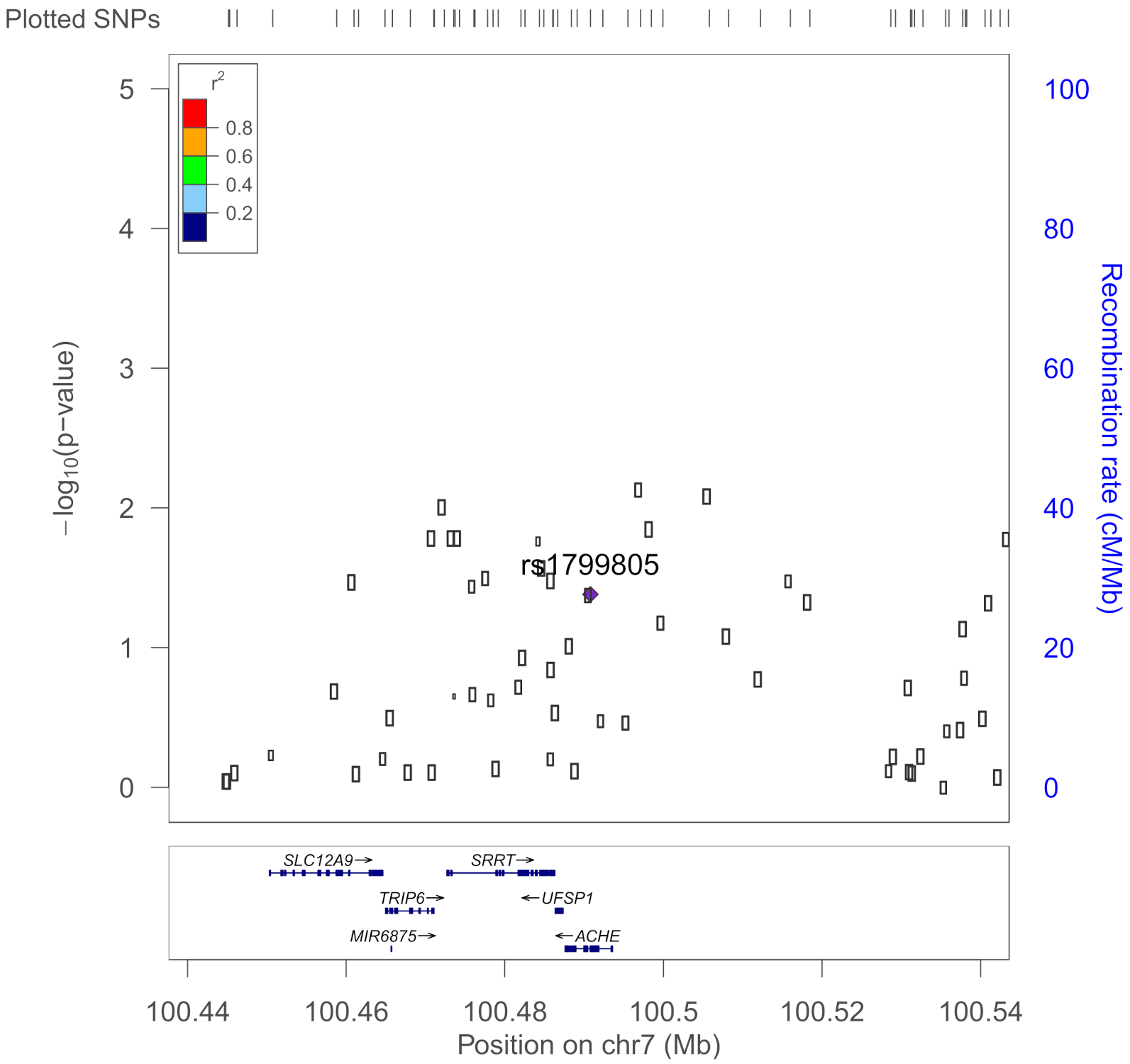
| rs1799805 | ACHE | H322N; Yt blood group; | 0.037 | Shapira et al., 2000 [43] |
| Tier 2 SNPs: Remaining SNPs in BCHE, PON1, ACHE, and SOD1 | ||||
| All SNPs in GWECB that are within 50 kb of the gene | Gene | Location (hg19) | Number of SNPs | |
| BCHE | Chr 3. 165490692–165555211 (+/− 50 kb) | 84 | ||
| PON1 | Chr 7. 94926988–94953844 (+/− 50 kb) | 178 | ||
| ACHE | Chr 7. 100487615–100494614 (+/− 50 kb) | 59 | ||
| SOD1 | Chr 21. 33032006–33041244 (+/− 50 kb) | 53 | ||
| Demographics | All (n = 810) | CDC Severe GWI | ||
|---|---|---|---|---|
| Yes (n = 212) | No (n = 598) | |||
| Sex | Female | 180 (22%) | 55 (26%) | 125 (21%) |
| Male | 630 (78%) | 157 (74%) | 473 (79%) | |
| Age Group | 40–49 | 321 (40%) | 98 (46%) | 223 (37%) |
| 50–59 | 296 (37%) | 74 (35%) | 222 (37%) | |
| 60+ | 193 (24%) | 40 (19%) | 153 (26%) | |
| Race/Ethnicity | White, non-Hispanic | 529 (65%) | 101 (48%) | 428 (72%) |
| Black, non-Hispanic | 137 (17%) | 43 (20%) | 94 (16%) | |
| Hispanic | 76 (9%) | 37 (17%) | 39 (7%) | |
| Other or missing | 68 (8%) | 31 (15%) | 37 (6%) | |
| Income | Under $30,000 | 84 (10%) | 33 (16%) | 51 (9%) |
| $30,000–59,999 | 179 (22%) | 60 (28%) | 119 (20%) | |
| $60,000–99,999 | 239 (30%) | 65 (31%) | 174 (29%) | |
| $100,000+ | 246 (30%) | 35 (17%) | 211 (35%) | |
| Education ^ | High school, GED, or less | 75 (9%) | 17 (8%) | 58 (10%) |
| Some college, or associate’s or bachelor’s degree | 544 (67%) | 161 (76%) | 383 (64%) | |
| Advanced degree | 166 (20%) | 24 (11%) | 142 (24%) | |
| Service Branch^ | Army only | 366 (45%) | 114 (54%) | 252 (42%) |
| Navy only | 139 (17%) | 28 (13%) | 111 (19%) | |
| Air Force only | 80 (10%) | 12 (6%) | 68 (11%) | |
| Marine Corps only | 105 (13%) | 26 (12%) | 79 (13%) | |
| National Guard, any | 81 (10%) | 21 (10%) | 60 (10%) | |
| Military Component | Active Duty | 470 (58%) | 136 (64%) | 334 (56%) |
| Both Active Duty and Reserves | 208 (26%) | 59 (28%) | 149 (25%) | |
| Reserves Only | 127 (16%) | 17 (8%) | 110 (18%) | |
| OEF/OIF Deployment a | Yes | 187 (23%) | 38 (18%) | 149 (25%) |
| No | 614 (76%) | 168 (79%) | 446 (75%) | |
| Kansas Exclusionary Criterion | Yes | 327 (40%) | 112 (53%) | 215 (36%) |
| No | 483 (60%) | 100 (47%) | 384 (64%) | |
| rs1799807 (BCHE) | Homozygous reference (AA) | 773 (95%) | 206 (97%) | 567 (95%) |
| Heterozygous (AG) | 36 (4%) | 6 (3%) | 30 (5%) | |
| Homozygous Atypical Variant (GG) | 0 (0%) | 0 (0%) | 0 (0%) | |
| rs1799805 (ACHE) | Homozygous reference (CC) | 746 (92%) | 199 (94%) | 547 (91%) |
| Heterozygous (CA) | 63 (8%) | 13 (6%) | 50 (8%) | |
| Homozygous YT variant (AA) | 0 (0%) | 0 (0%) | 0 (0%) | |
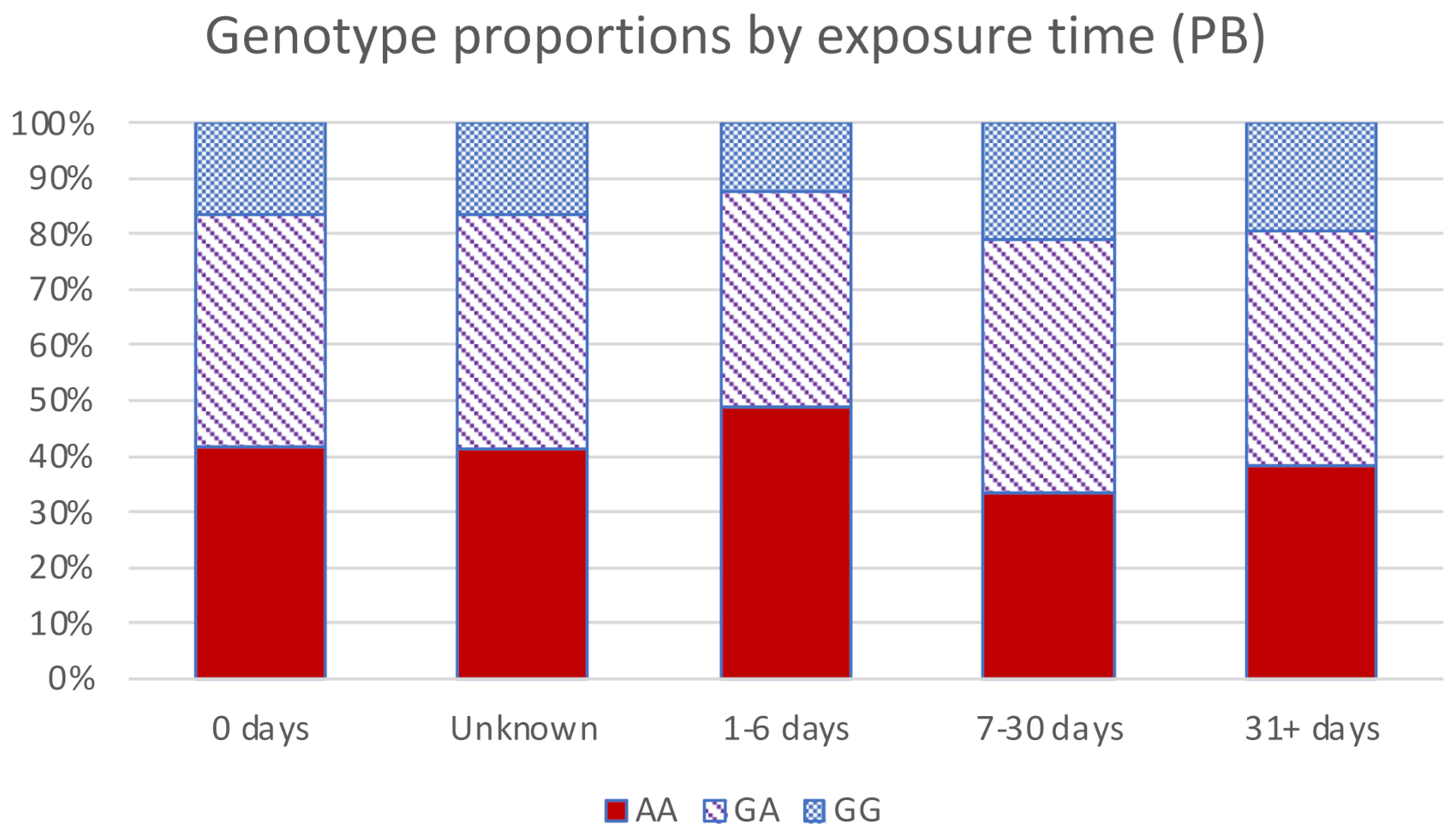
| rs662 (PON1) | Homozygous 192Q variant (AA) | 329 (41%) | 67 (32%) | 262 (44%) |
| Heterozygous (AG) | 340 (42%) | 100 (47%) | 240 (40%) | |
| Homozygous 192R variant (GG) | 141 (17%) | 45 (21%) | 96 (16%) | |
| Exposure | Length of Time | All | CDC Severe GWI | |
|---|---|---|---|---|
| Yes | No | |||
| All | 810 | 212 (26%) | 598 (74%) | |
| PB pill exposure | None | 311 | 55 (18%) | 256 (82%) |
| Unclear or missing | 138 | 41 (30%) | 97 (70%) | |
| 1–6 days | 98 | 27 (28%) | 71 (72%) | |
| 7–30 days | 114 | 35 (31%) | 79 (69%) | |
| 31+ days | 149 | 54 (36%) | 95 (64%) | |
| Pesticide exposure | None | 346 | 67 (19%) | 279 (81%) |
| Unclear or missing | 106 | 37 (35%) | 69 (65%) | |
| 1–30 days | 78 | 19 (24%) | 59 (76%) | |
| 31–62 days | 124 | 37 (30%) | 87 (70%) | |
| 63+ days | 156 | 52 (33%) | 104 (67%) | |
| Outcome | Exposure | ACHE | BCHE | PON1 | |||
|---|---|---|---|---|---|---|---|
| rs1799805 | rs1799807 | rs662 | |||||
| OR | p | OR | p | OR | p | ||
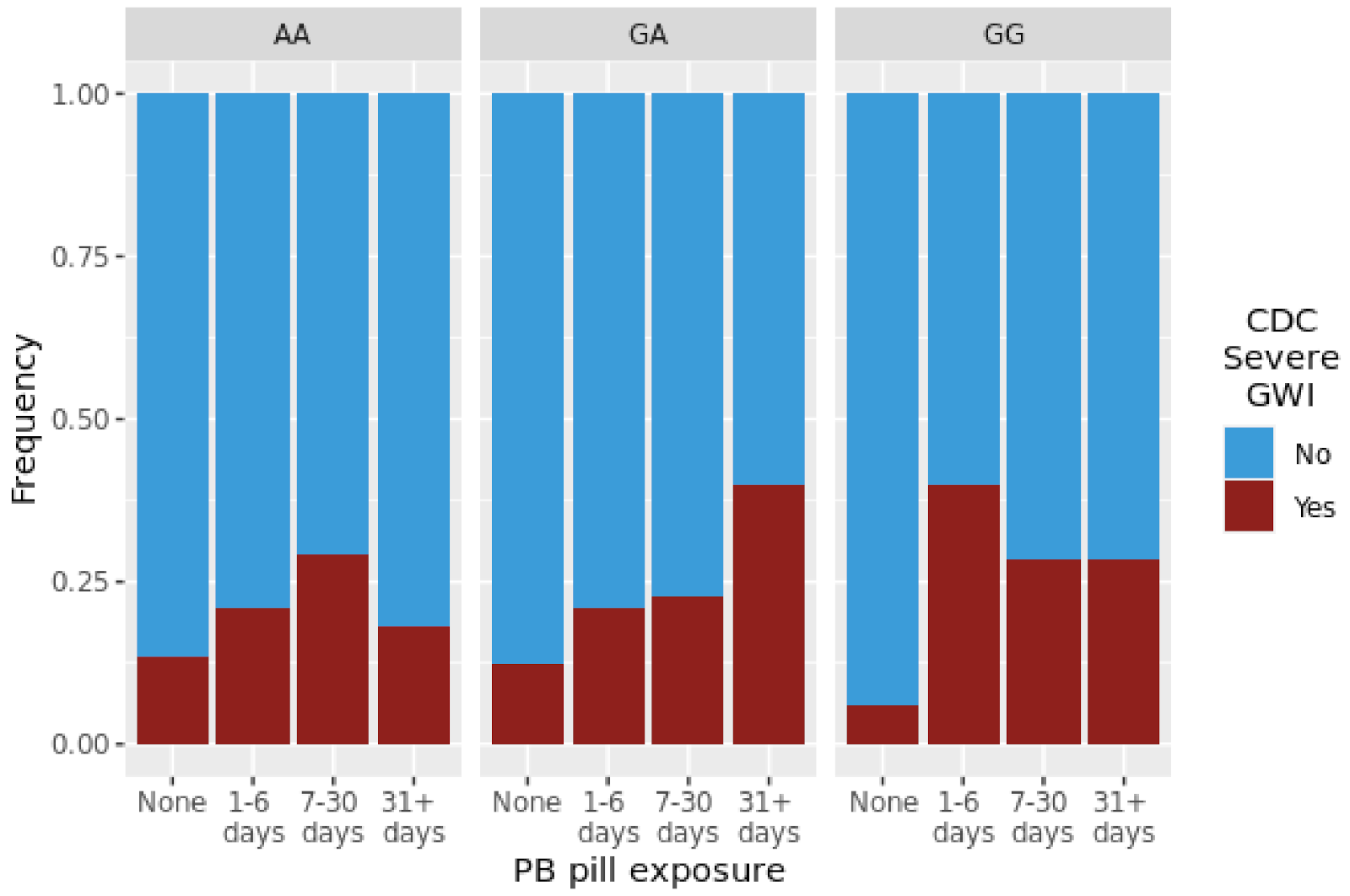
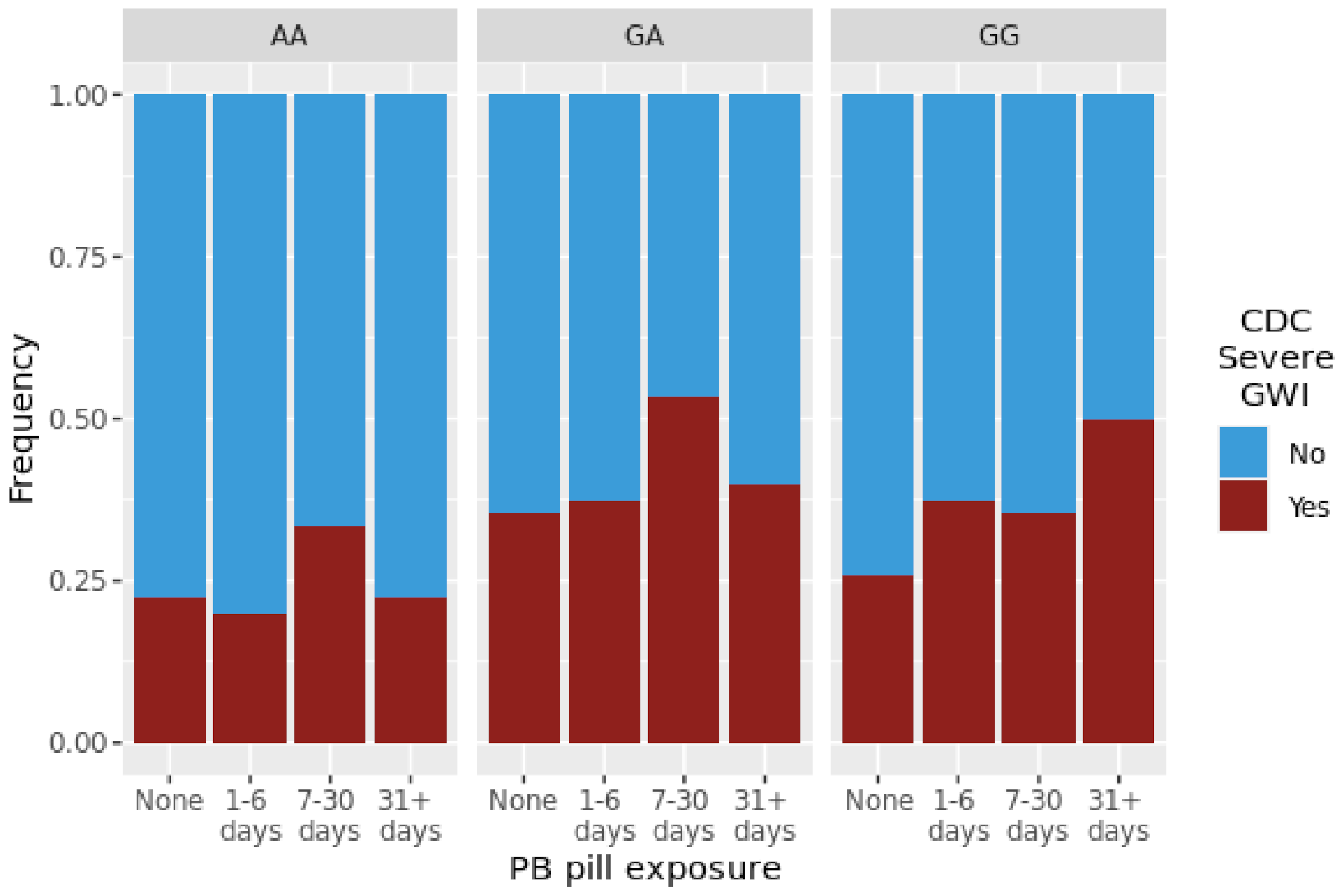
| CDC Severe GWI | PB | 1.11 | 0.725 | 0.93 | 0.848 | 1.22 | 0.049 * |
| CDC Severe: Fatigue | PB | 1.50 | 0.268 | 1.63 | 0.254 | 1.21 | 0.105 |
| CDC Severe: Mood/Cognitive | PB | 1.05 | 0.856 | 1.13 | 0.721 | 1.13 | 0.201 |
| CDC Severe: Musculoskeletal | PB | 0.99 | 0.953 | 0.86 | 0.639 | 1.13 | 0.202 |
| Kansas GWI symptom criteria | PB | 0.93 | 0.821 | 1.28 | 0.599 | 0.99 | 0.923 |
| Kansas: Fatigue | PB | 0.93 | 0.838 | 1.01 | 0.979 | 1.03 | 0.814 |
| Kansas: Pain | PB | 1.26 | 0.443 | 1.61 | 0.356 | 1.16 | 0.225 |
| Kansas: Gastrointestinal | PB | 0.96 | 0.883 | 1.18 | 0.614 | 0.96 | 0.663 |
| Kansas: Respiratory | PB | 1.15 | 0.581 | 0.95 | 0.868 | 1.18 | 0.080 |
| Kansas: Mood/Neurological/ Cognitive | PB | 0.58 | 0.251 | 1.09 | 0.855 | 1.21 | 0.278 |
| Kansas: Skin | PB | 0.59 | 0.042 * | 1.3 | 0.408 | 1.01 | 0.897 |
| CDC Severe GWI | Pesticides | 0.67 | 0.179 | 0.78 | 0.548 | 1.07 | 0.485 |
| CDC Severe: Fatigue | Pesticides | 0.89 | 0.739 | 1.27 | 0.587 | 1.14 | 0.235 |
| CDC Severe: Mood/Cognitive | Pesticides | 0.65 | 0.123 | 1.17 | 0.676 | 1.00 | 0.958 |
| CDC Severe: Musculoskeletal | Pesticides | 0.61 | 0.063 | 0.95 | 0.887 | 1.04 | 0.684 |
| Kansas GWI symptom criteria | Pesticides | 0.60 | 0.128 | 2.11 | 0.189 | 1.05 | 0.660 |
| Kansas: Fatigue | Pesticides | 0.76 | 0.484 | 0.63 | 0.281 | 0.97 | 0.815 |
| Kansas: Pain | Pesticides | 0.67 | 0.151 | 1.47 | 0.362 | 1.17 | 0.185 |
| Kansas: Gastrointestinal | Pesticides | 0.84 | 0.490 | 1.11 | 0.747 | 0.82 | 0.029 * |
| Kansas: Respiratory | Pesticides | 0.66 | 0.103 | 1.39 | 0.320 | 1.00 | 0.970 |
| Kansas: Mood/Neurological/ Cognitive | Pesticides | 1.09 | 0.893 | 1.72 | 0.353 | 1.11 | 0.562 |
| Kansas: Skin | Pesticides | 0.65 | 0.098 | 1.28 | 0.451 | 1.00 | 0.96 |
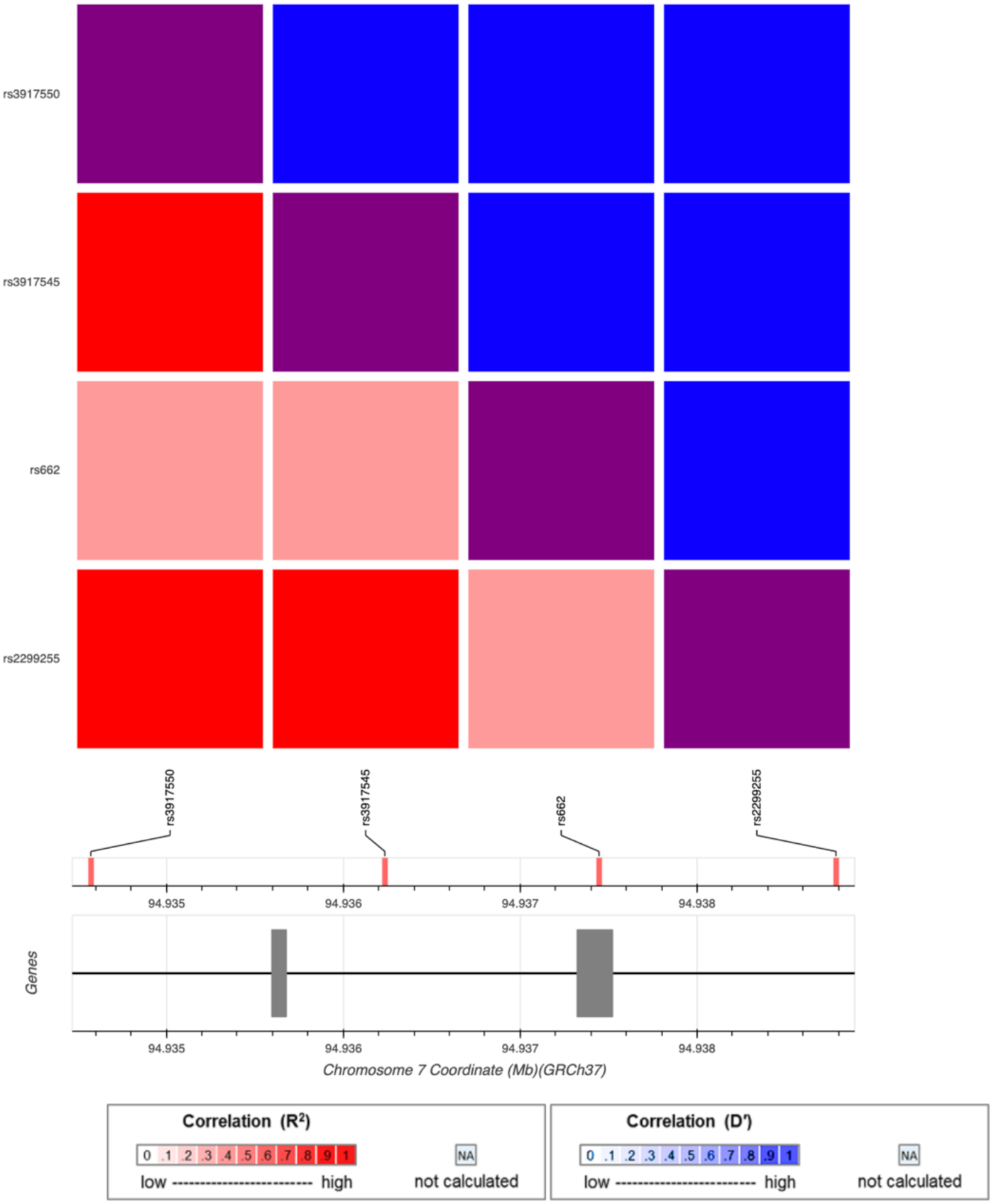
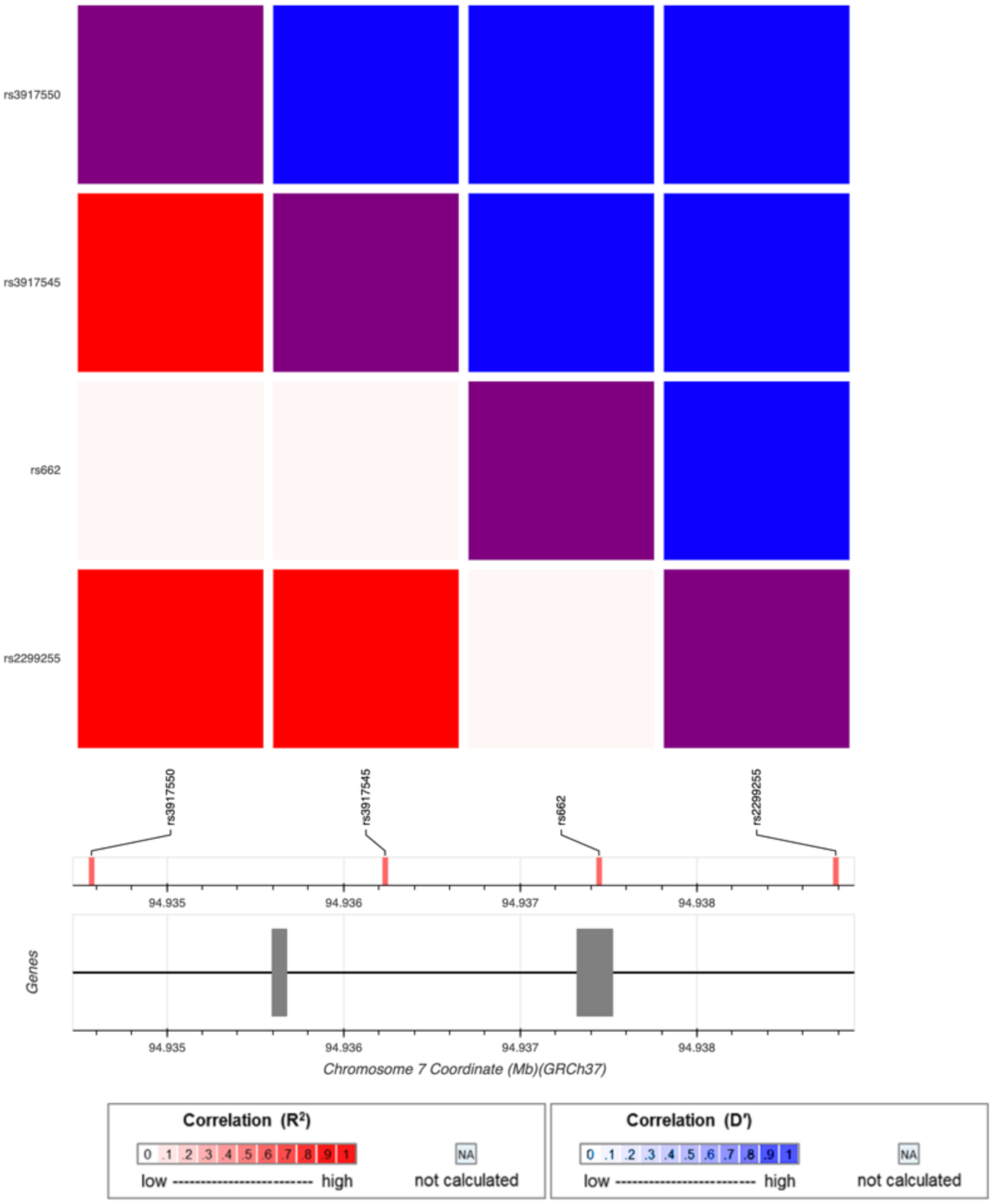
| Outcome | Exposure | PON1 | PON1 | PON1 | |||
|---|---|---|---|---|---|---|---|
| rs3917545 | rs2299260 | rs62467349 | |||||
| OR | p | OR | p | OR | p | ||
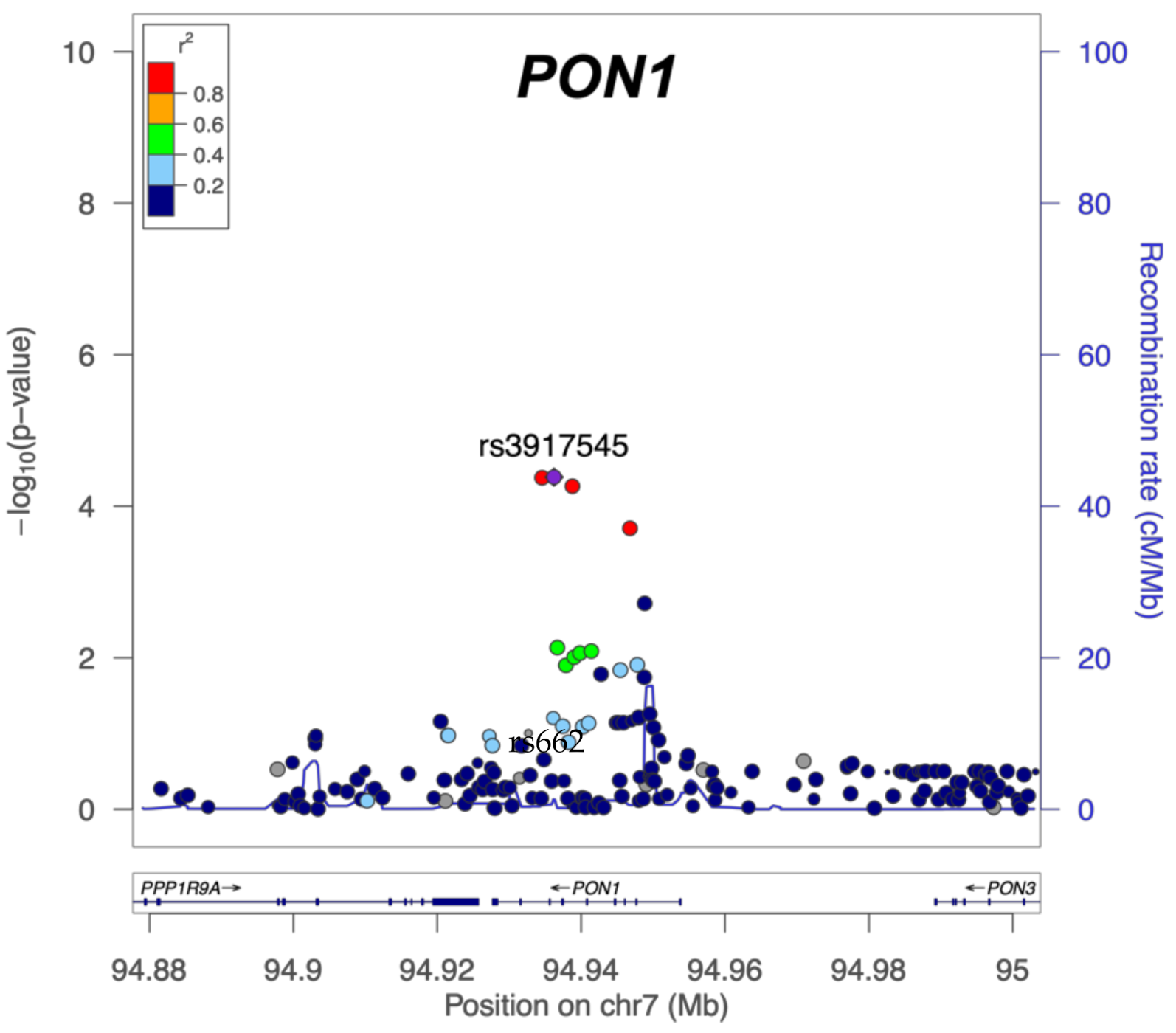
| CDC Severe GWI | PB | 1.45 | 3.20 × 10−2 | 0.68 | 5.00 × 10−3 | 1.39 | 5.50 × 10−2 |
| CDC Severe: Fatigue | PB | 1.54 | 2.50 × 10−2 | 0.64 | 7.60 × 10−3 | 1.52 | 3.10 × 10−2 |
| CDC Severe: Mood/Cognitive | PB | 1.25 | 1.70 × 10−1 | 0.82 | 1.20 × 10−1 | 1.23 | 2.10 × 10−1 |
| CDC Severe: Musculoskeletal | PB | 1.44 | 2.80 × 10−2 | 0.76 | 3.00 × 10−2 | 1.38 | 5.00 × 10−2 |
| Kansas GWI symptom criteria | PB | 1.23 | 3.20 × 10−1 | 0.99 | 9.70 × 10−1 | 1.23 | 3.20 × 10−1 |
| Kansas: Pain | PB | 1.28 | 2.30 × 10−1 | 0.73 | 3.50 × 10−2 | 1.31 | 1.90 × 10−1 |
| Kansas: Gastrointestinal | PB | 1.09 | 5.90 × 10−1 | 0.79 | 6.10 × 10−2 | 1.03 | 8.60 × 10−1 |
| Kansas: Mood/Neurological/Cognitive | PB | 2.30 | 3.30 × 10−2 | 0.89 | 5.60 × 10−1 | 2.18 | 4.40 × 10−2 |
| Kansas: Fatigue | PB | 1.14 | 5.70 × 10−1 | 0.95 | 7.90 × 10−1 | 1.13 | 5.80 × 10−1 |
| Kansas: Respiratory | PB | 2.00 | 4.1 × 10−5 * | 0.78 | 5.50 × 10−2 | 1.86 | 2.00 × 10−4 |
| Kansas: Skin | PB | 1.11 | 5.20 × 10−1 | 0.92 | 4.80 × 10−1 | 1.11 | 5.10 × 10−1 |
| CDC Severe GWI | Pesticides | 1.45 | 2.20 × 10−2 | 0.89 | 3.70 × 10−1 | 1.54 | 8.60 × 10−3 |
| CDC Severe: Fatigue | Pesticides | 1.41 | 5.70 × 10−2 | 0.97 | 8.60 × 10−1 | 1.38 | 7.50 × 10−2 |
| CDC Severe: Mood/Cognitive | Pesticides | 1.05 | 7.70 × 10−1 | 0.94 | 5.90 × 10−1 | 1.07 | 6.50 × 10−1 |
| CDC Severe: Musculoskeletal | Pesticides | 1.19 | 2.40 × 10−1 | 0.86 | 2.20 × 10−1 | 1.26 | 1.40 × 10−1 |
| Kansas GWI symptom criteria | Pesticides | 1.30 | 1.80 × 10−1 | 0.85 | 2.70 × 10−1 | 1.37 | 1.20 × 10−1 |
| Kansas: Pain | Pesticides | 0.99 | 9.50 × 10−1 | 1.05 | 7.30 × 10−1 | 1.05 | 7.80 × 10−1 |
| Kansas: Gastrointestinal | Pesticides | 0.84 | 2.20 × 10−1 | 1.07 | 5.50 × 10−1 | 0.83 | 2.00 × 10−1 |
| Kansas: Mood/Neurological/Cognitive | Pesticides | 1.41 | 1.90 × 10−1 | 0.86 | 4.30 × 10−1 | 1.38 | 2.20 × 10−1 |
| Kansas: Fatigue | Pesticides | 0.99 | 9.60 × 10−1 | 0.79 | 1.40 × 10−1 | 1.03 | 8.80 × 10−1 |
| Kansas: Respiratory | Pesticides | 1.28 | 1.00 × 10−1 | 0.98 | 8.80 × 10−1 | 1.27 | 1.10 × 10−1 |
| Kansas: Skin | Pesticides | 1.04 | 7.90 × 10−1 | 1.09 | 4.90 × 10−1 | 0.99 | 9.20 × 10−1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vahey, J.; Gifford, E.J.; Sims, K.J.; Chesnut, B.; Boyle, S.H.; Stafford, C.; Upchurch, J.; Stone, A.; Pyarajan, S.; Efird, J.T.; et al. Gene–Toxicant Interactions in Gulf War Illness: Differential Effects of the PON1 Genotype. Brain Sci. 2021, 11, 1558. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11121558
Vahey J, Gifford EJ, Sims KJ, Chesnut B, Boyle SH, Stafford C, Upchurch J, Stone A, Pyarajan S, Efird JT, et al. Gene–Toxicant Interactions in Gulf War Illness: Differential Effects of the PON1 Genotype. Brain Sciences. 2021; 11(12):1558. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11121558
Chicago/Turabian StyleVahey, Jacqueline, Elizabeth J. Gifford, Kellie J. Sims, Blair Chesnut, Stephen H. Boyle, Crystal Stafford, Julie Upchurch, Annjanette Stone, Saiju Pyarajan, Jimmy T. Efird, and et al. 2021. "Gene–Toxicant Interactions in Gulf War Illness: Differential Effects of the PON1 Genotype" Brain Sciences 11, no. 12: 1558. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11121558