
Recognition Memory in Noonan Syndrome

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Tasks
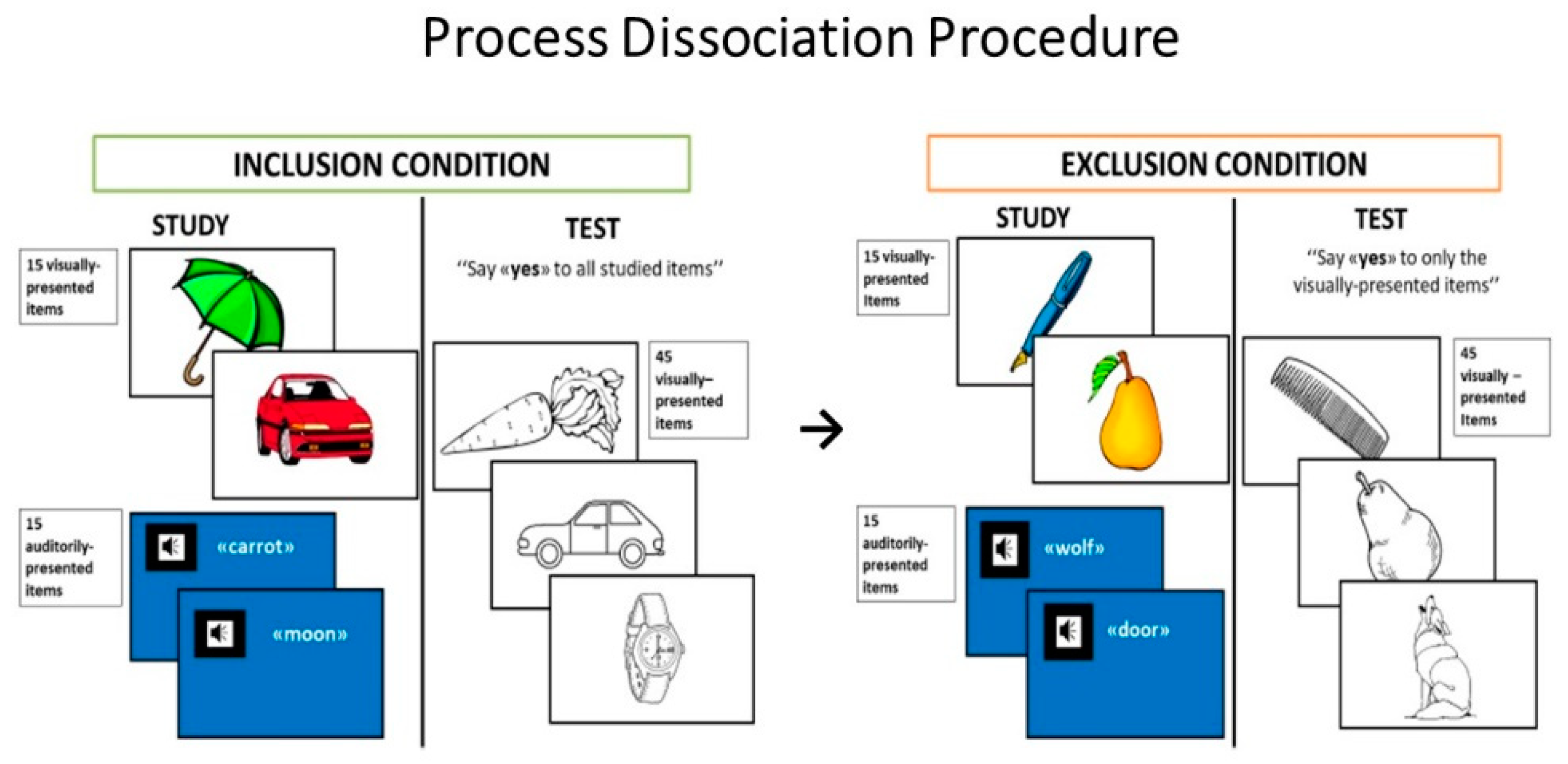
2.2.1. Process Dissociation Procedure
Stimuli
Procedure
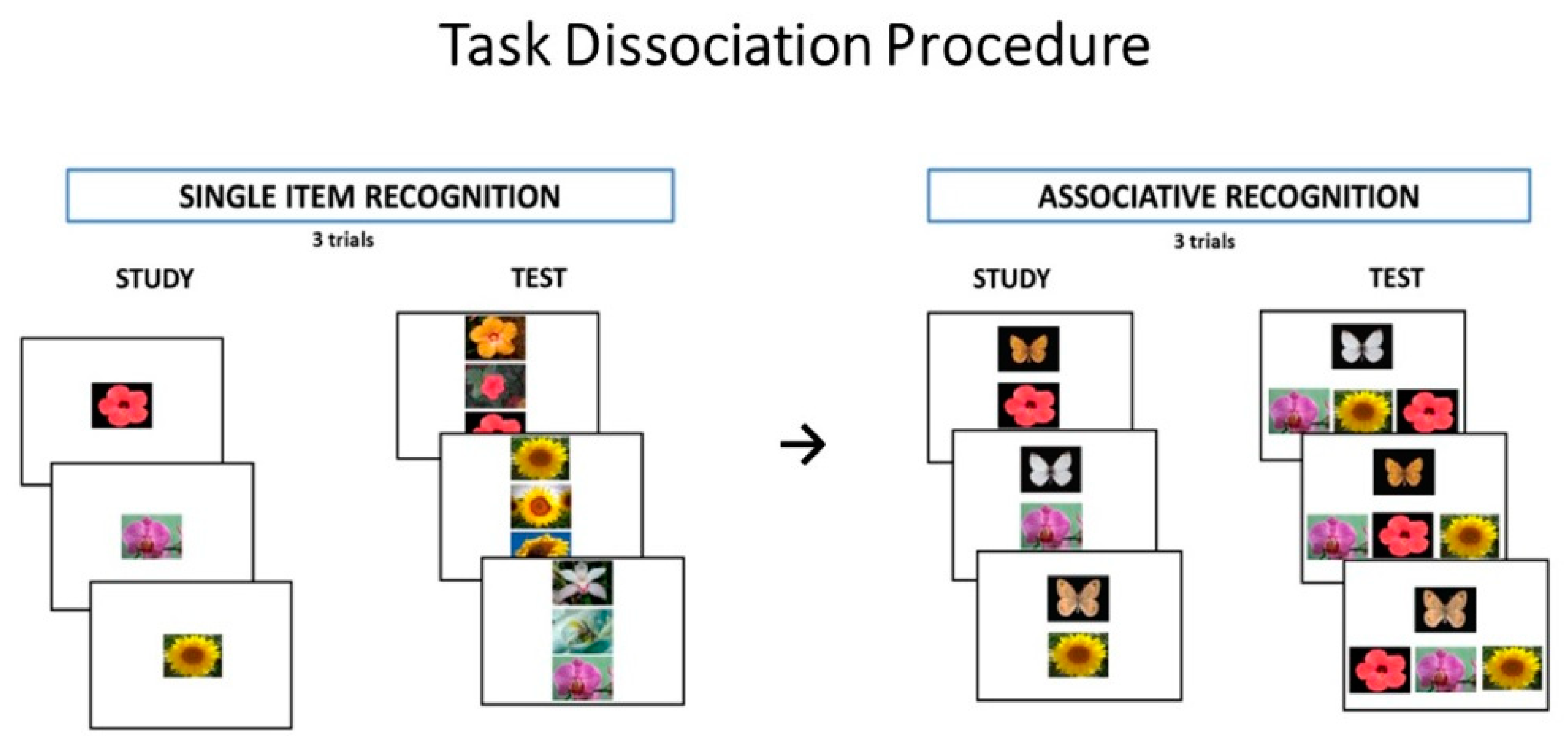
2.2.2. Task Dissociation Procedure
Stimuli
Procedure
2.3. Statistical Analysis
3. Results
3.1. Process Dissociation Procedure
3.2. Task Dissociation Procedure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allanson, J.E. Noonan syndrome. Am. J. Med. Genet. Part C Semin. Med. Genet. 2007, 145, 274–279. [Google Scholar] [CrossRef]
- Roberts, A.E.; Allanson, J.E.; Tartaglia, M.; Gelb, B.D. Noonan syndrome. Lancet 2013, 381, 333–342. [Google Scholar] [CrossRef] [Green Version]
- Romano, A.A.; Allanson, J.E.; Dahlgren, J.; Gelb, B.D.; Hall, B.; Pierpont, M.E.; Roberts, A.E.; Robinson, W.; Takemoto, C.M.; Noonan, J.A. Noonan syndrome: Clinical features, diagnosis, and management guidelines. Pediatrics 2010, 126, 746–759. [Google Scholar] [CrossRef] [Green Version]
- Van der Burgt, I. Noonan syndrome. Orphanet J. Rare Dis. 2007, 2, 4. [Google Scholar] [CrossRef] [PubMed]
- Aoki, Y.; Niihori, T.; Inoue, S.; Matsubara, Y. Recent advances in RASopathies. J. Hum. Genet. 2016, 61, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Tartaglia, M.; Gelb, B.D. Disorders of dysregulated signal traffic through the RAS-MAPK pathway: Phenotypic spectrum and molecular mechanisms. Ann. N. Y. Acad. Sci. 2010, 1214, 99–121. [Google Scholar] [CrossRef] [Green Version]
- Fey, D.; Matallanas, D.; Rauch, J.; Rukhlenko, O.S.; Kholodenko, B.N. The complexities and versatility of the RAS-to-ERK signalling system in normal and cancer cells. Semin. Cell Dev. Biol. 2016, 58, 96–107. [Google Scholar] [CrossRef]
- Ryu, H.H.; Lee, Y.S. Cell type-specific roles of RAS-MAPK signaling in learning and memory: Implications in neurodevelopmental disorders. Neurobiol. Learn. Mem. 2016, 135, 13–21. [Google Scholar] [CrossRef]
- Krab, L.C.; Goorden, S.M.; Elgersma, Y. Oncogenes on my mind: ERK and MTOR signaling in cognitive diseases. Trends Genet. TIG 2008, 24, 498–510. [Google Scholar] [CrossRef]
- Lee, D.A.; Portnoy, S.; Hill, P.; Gillberg, C.; Patton, M.A. Psychological profile of children with Noonan syndrome. Dev. Med. Child Neurol. 2005, 47, 35–38. [Google Scholar] [CrossRef]
- Wingbermuehle, E.; Egger, J.; van der Burgt, I.; Verhoeven, W. Neuropsychological and behavioral aspects of Noonan syndrome. Horm. Res. 2009, 72, 15–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesarini, L.; Alfieri, P.; Pantaleoni, F.; Vasta, I.; Cerutti, M.; Petrangeli, V.; Mariotti, P.; Leoni, C.; Ricci, D.; Vicari, S.; et al. Cognitive profile of disorders associated with dysregulation of the RAS/MAPK signaling cascade. Am. J. Med. Genet. Part A 2009, 149, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Pierpont, E.I.; Pierpont, M.E.; Mendelsohn, N.J.; Roberts, A.E.; Tworog-Dube, E.; Seidenberg, M.S. Genotype differences in cognitive functioning in Noonan syndrome. Genes Brain Behav. 2009, 8, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Sweatt, J.D. The neuronal MAP kinase cascade: A biochemical signal integration system subserving synaptic plasticity and memory. J. Neurochem. 2001, 76, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sweatt, J.D.; Weeber, E.J.; Lombroso, P.J. Genetics of childhood disorders: LI. Learning and memory, Part 4: Human cognitive disorders and the ras/ERK/CREB pathway. J. Am. Acad. Child Adolesc. Psychiatry 2003, 42, 741–744. [Google Scholar] [CrossRef] [Green Version]
- Sweatt, J.D.; Weeber, E.J. Genetics of childhood disorders: LII. Learning and memory, part 5: Human cognitive disorders and the ras/ERK/CREB pathway. J. Am. Acad. Child Adolesc. Psychiatry 2003, 42, 873–876. [Google Scholar] [CrossRef] [Green Version]
- Sweatt, J.D. Mitogen-activated protein kinases in synaptic plasticity and memory. Curr. Opin. Neurobiol. 2004, 14, 311–317. [Google Scholar] [CrossRef]
- Pagani, M.R.; Oishi, K.; Gelb, B.D.; Zhong, Y. The phosphatase SHP2 regulates the spacing effect for long-term memory induction. Cell 2009, 139, 186–198. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S.; Ehninger, D.; Zhou, M.; Oh, J.Y.; Kang, M.; Kwak, C.; Ryu, H.H.; Butz, D.; Araki, T.; Cai, Y.; et al. Mechanism and treatment for learning and memory deficits in mouse models of Noonan syndrome. Nat. Neurosci. 2014, 17, 1736–1743. [Google Scholar] [CrossRef]
- Alfieri, P.; Cesarini, L.; Mallardi, M.; Piccini, G.; Caciolo, C.; Leoni, C.; Mirante, N.; Pantaleoni, F.; Digilio, M.C.; Gambardella, M.L.; et al. Long term memory profile of disorders associated with dysregulation of the RAS-MAPK signaling cascade. Behav. Genet. 2011, 41, 423–429. [Google Scholar] [CrossRef]
- Schwartz, D.D.; Katzenstein, J.M.; Hopkins, E.; Stabley, D.L.; Sol-Church, K.; Gripp, K.W.; Axelrad, M.E. Verbal memory functioning in adolescents and young adults with Costello syndrome: Evidence for relative preservation in recognition memory. Am. J. Med. Genet. Part A 2013, 161, 2258–2265. [Google Scholar] [CrossRef] [Green Version]
- Pierpont, E.I.; Tworog-Dube, E.; Roberts, A.E. Learning and memory in children with Noonan syndrome. Am. J. Med. Genet. Part A 2013, 161, 2250–2257. [Google Scholar] [CrossRef] [PubMed]
- Mandler, G. Recognizing: The judgement of previous occurrence. Psychol. Rev. 1980, 87, 252–271. [Google Scholar] [CrossRef]
- Tulving, E. Memory and consciousness. Can. Psychol. 1985, 26, 1–12. [Google Scholar] [CrossRef]
- Yonelinas, A.P. The nature of recollection and familiarity: A review of 30 years of research. J. Mem. Lang. 2002, 46, 441–517. [Google Scholar] [CrossRef] [Green Version]
- Montaldi, D.; Mayes, A.R. The role of recollection and familiarity in the functional differentiation of the medial temporal lobes. Hippocampus 2010, 20, 1291–1314. [Google Scholar] [CrossRef]
- Yonelinas, A.P.; Kroll, N.E.; Dobbins, I.; Lazzara, M.; Knight, R.T. Recollection and familiarity deficits in amnesia: Convergence of remember-know, process dissociation, and receiver operating characteristic data. Neuropsychology 1998, 12, 323–339. [Google Scholar] [CrossRef]
- Wixted, J.T. Dual-process theory and signal-detection theory of recognition memory. Psychol. Rev. 2007, 114, 152–176. [Google Scholar] [CrossRef] [Green Version]
- Wais, P.E. FMRI signals associated with memory strength in the medial temporal lobes: A meta-analysis. Neuropsychologia 2008, 46, 3185–3196. [Google Scholar] [CrossRef]
- Wais, P.E. Hippocampal signals for strong memory when associative memory is available and when it is not. Hippocampus 2011, 21, 9–21. [Google Scholar] [CrossRef]
- Wais, P.E. The limited usefulness of models based on recollection and familiarity. J. Neurophysiol. 2013, 109, 1687–1689. [Google Scholar] [CrossRef] [Green Version]
- Wais, P.E.; Mickes, L.; Wixted, J.T. Remember/know judgments probe degrees of recollection. J. Cogn. Neurosci. 2008, 20, 400–405. [Google Scholar] [CrossRef]
- Wais, P.E.; Squire, L.R.; Wixted, J.T. In search of recollection and familiarity signals in the hippocampus. J. Cogn. Neurosci. 2010, 22, 109–123. [Google Scholar] [CrossRef] [Green Version]
- Norman, K.A.; O’Reilly, R.C. Modeling hippocampal and neocortical contributions to recognition memory: A complementary-learning-systems approach. Psychol. Rev. 2003, 110, 611–646. [Google Scholar] [CrossRef] [Green Version]
- Evans, L.H.; Wilding, E.L. 20 Recollection and familiarity make independent contributions to memory judgments. J. Neurosci. 2012, 32, 7253–7257. [Google Scholar] [CrossRef]
- Perner, J.; Ruffman, T. Episodic memory and autonoetic consciousness: Developmental evidence and a theory of childhood amnesia. J. Exp. Child Psychol. 1995, 59, 516–548. [Google Scholar] [CrossRef]
- Brainerd, C.J.; Holliday, R.E.; Reyna, V.F. Behavioral measurement of remembering phenomenologies: So simple a child can do it. Child Dev. 2004, 75, 505–522. [Google Scholar] [CrossRef]
- Holliday, R.E. Reducing misinformation effects in children with cognitive interviews: Dissociating recollection and familiarity. Child Dev. 2003, 74, 728–751. [Google Scholar] [CrossRef]
- Holliday, R.E.; Hayes, B.K. Dissociating automatic and intentional processes in children’s eyewitness memory. J. Exp. Child Psychol. 2000, 75, 1–42. [Google Scholar] [CrossRef]
- Koenig, L.; Wimmer, M.C.; Hollins, T.J. Process dissociation of familiarity and recollection in children: Response deadline affects recollection but not familiarity. J. Exp. Child Psychol. 2015, 131, 120–134. [Google Scholar] [CrossRef]
- Ofen, N.; Kao, Y.C.; Sokol-Hessner, P.; Kim, H.; Whitfield-Gabrieli, S.; Gabrieli, J.D. Development of the declarative memory system in the human brain. Nat. Neurosci. 2007, 10, 1198–1205. [Google Scholar] [CrossRef]
- Ofen, N.; Tang, L.; Yu, Q.; Johnson, E.L. Memory and the developing brain: From description to explanation with innovation in methods. Dev. Cogn. Neurosci. 2019, 36, 100613. [Google Scholar] [CrossRef]
- Riggins, T.; Geng, F.; Botdorf, M.; Canada, K.; Cox, L.; Hancock, G.R. Protracted hippocampal development is associated with age-related improvements in memory during early childhood. NeuroImage 2018, 174, 127–137. [Google Scholar] [CrossRef]
- Schneider, W.; Ornstein, P.A. The development of children’s memory. Child Dev. Perspect. 2015, 9, 190–195. [Google Scholar]
- Daugherty, A.M.; Flinn, R.; Ofen, N. Hippocampal CA3-dentate gyrus volume uniquely linked to improvement in associative memory from childhood to adulthood. NeuroImage 2017, 153, 75–85. [Google Scholar] [CrossRef]
- DeMaster, D.; Pathman, T.; Lee, J.K.; Ghetti, S. Structural development of the hippocampus and episodic memory: Developmental differences along the anterior/posterior axis. Cereb. Cortex 2014, 24, 3036–3045. [Google Scholar] [CrossRef] [Green Version]
- Schlichting, M.L.; Guarino, K.F.; Schapiro, A.C.; Turk-Browne, N.B.; Preston, A.R. Hippocampal structure predicts statistical learning and associative inference abilities during development. J. Cogn. Neurosci. 2017, 29, 37–51. [Google Scholar] [CrossRef] [Green Version]
- Keresztes, A.; Bender, A.R.; Bodammer, N.C.; Lindenberger, U.; Shing, Y.L.; Werkle-Bergner, M. Hippocampal maturity promotes memory distinctiveness in childhood and adolescence. Proc. Natl. Acad. Sci. USA 2017, 114, 9212–9217. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Shafer, A.T.; Ofen, N. Prefrontal cortex contributions to the development of memory formation. Cereb. Cortex 2018, 28, 3295–3308. [Google Scholar] [CrossRef] [Green Version]
- Bigham, S.; Boucher, J.; Mayes, A.; Anns, S. Assessing recollection and familiarity in autistic spectrum disorders: Methods and findings. J. Autism Dev. Disord. 2010, 40, 878–889. [Google Scholar] [CrossRef]
- Costanzo, F.; Vicari, S.; Carlesimo, G.A. Familiarity and recollection in Williams syndrome. Cortex 2013, 49, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, L.L. A process dissociation framework: Separating automatic from intentional uses of memory. J. Mem. Lang. 1991, 30, 513–541. [Google Scholar] [CrossRef]
- Koenig, L.; Wimmer, M.C.; Trippas, D. Item repetition and response deadline affect familiarity and recollection differently across childhood. Memory 2020, 28, 900–907. [Google Scholar] [CrossRef]
- Belacchi, C.; Scalisi, T.G.; Cannoni, E.; Cornoldi, C.; Manuale, C.P.M. Coloured Progressive Matrices. Standardizzazione Italiana; Giunti, O.S., Ed.; Organizzazioni Speciali: Firenze, Italy, 2008. [Google Scholar]
- Stella, G.; Pizzoli, C.; Tressoldi, P.E. PPVT-R, Peabody Picture Vocabulary Test-Revised; Omega: Torino, Italy, 2000. [Google Scholar]
- Snodgrass, J.G.; Vanderwart, M. A standardized set of 260 pictures: Norms for name agreement, image agreement, familiarity, and visual complexity. J. Exp. Psychol. Hum. Learn. 1980, 6, 174–215. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, P.; Barca, L.; Burani, C. A database for semantic, grammatical, and frequency properties of the first words acquired by Italian children. Behav. Res. Methods Instrum. Comput. 2004, 36, 525–530. [Google Scholar] [CrossRef] [Green Version]
- Tamnes, C.K.; Walhovd, K.B.; Engvig, A.; Grydeland, H.; Krogsrud, S.K.; Østby, Y.; Holland, D.; Dale, A.M.; Fjell, A.M. Regional hippocampal volumes and development predict learning and memory. Dev. Neurosci. 2014, 36, 161–174. [Google Scholar] [CrossRef] [Green Version]
- Bastin, C.; Linden, M.; Charnallet, A.; Denby, C.; Montaldi, D.; Roberts, N.; Andrew, M. Dissociation between recall and recognition memory performance in an amnesic patient with hippocampal damage following carbon monoxide poisoning. Neurocase 2004, 10, 330–344. [Google Scholar] [CrossRef]
- Zhu, J.J.; Qin, Y.; Zhao, M.; Van Aelst, L.; Malinow, R. Ras and Rap control AMPA receptor trafficking during synaptic plasticity. Cell 2002, 110, 443–455. [Google Scholar] [CrossRef] [Green Version]
- Josselyn, S.A.; Frankland, P.W. Infantile amnesia: A neurogenic hypothesis. Learn. Mem. 2012, 19, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Lavenex, P.; Banta Lavenex, P. Building hippocampal circuits to learn and remember: Insights into the development of human memory. Behav. Brain Res. 2013, 254, 8–21. [Google Scholar] [CrossRef]
- Johnson, E.M.; Ishak, A.D.; Naylor, P.E.; Stevenson, D.A.; Reiss, A.L.; Green, T. PTPN11 Gain-of-Function Mutations Affect the Developing Human Brain, Memory, and Attention. Cereb. Cortex 2019, 29, 2915–2923. [Google Scholar] [CrossRef] [PubMed]
- Altmüller, F.; Pothula, S.; Annamneedi, A.; Nakhaei-Rad, S.; Montenegro-Venegas, C.; Pina-Fernández, E.; Marini, C.; Santos, M.; Schanze, D.; Montag, D.; et al. Correction: Aberrant neuronal activity-induced signaling and gene expression in a mouse model of RASopathy. PLoS Genet. 2017, 13, e1006843. [Google Scholar] [CrossRef] [PubMed]
- Squire, L.R.; Wixted, J.T.; Clark, R.E. Recognition memory and the medial temporal lobe: A new perspective. Nat. Rev. Neurosci. 2007, 8, 872–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fattah, M.; Raman, M.M.; Reiss, A.L.; Green, T. PTPN11 Mutations in the Ras-MAPK Signaling Pathway Affect Human White Matter Microstructure. Cereb. Cortex 2020. [Google Scholar] [CrossRef]
- Canada, K.L.; Ngo, C.T.; Newcombe, N.S.; Geng, F.; Riggins, T. It’s All in the Details: Relations Between Young Children’s Developing Pattern Separation Abilities and Hippocampal Subfield Volumes. Cereb. Cortex 2019, 29, 3427–3433. [Google Scholar] [CrossRef]
- Davachi, L. Item, context and relational episodic encoding in humans. Curr. Opin. Neurobiol. 2006, 16, 693–700. [Google Scholar] [CrossRef]
- Rugg, M.D.; Vilberg, K.L.; Mattson, J.T.; Yu, S.S.; Johnson, J.D.; Suzuki, M. Item memory, context memory and the hippocampus: fMRI evidence. Neuropsychologia 2012, 50, 3070–3079. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.G.; Biederman, I. Where do objects become scenes? Cereb. Cortex 2011, 21, 1738–1746. [Google Scholar] [CrossRef]
- Rémy, F.; Vayssière, N.; Saint-Aubert, L.; Bacon-Macé, N.; Pariente, J.; Barbeau, E.; Fabre-Thorpe, M. Age effects on the neural processing of object-context associations in briefly flashed natural scenes. Neuropsychologia 2020, 136, 107264. [Google Scholar] [CrossRef]
- Kafkas, A.; Migo, E.M.; Morris, R.G.; Kopelman, M.D.; Montaldi, D.; Mayes, A.R. Material Specificity Drives Medial Temporal Lobe Familiarity But Not Hippocampal Recollection. Hippocampus 2017, 27, 194–209. [Google Scholar] [CrossRef] [Green Version]
- Ofen, N.; Chai, X.J.; Schuil, K.D.; Whitfield-Gabrieli, S.; Gabrieli, J.D. The development of brain systems associated with successful memory retrieval of scenes. J. Neurosci. 2012, 32, 10012–10020. [Google Scholar] [CrossRef] [Green Version]
- Rajaram, S. Perceptual effects on remembering: Recollective processes in picture recognition memory. J. Exp. Psychol. 1996, 22, 365–377. [Google Scholar] [CrossRef]
- Craik, F.I.; Govoni, R.; Naveh-Benjamin, M.; Anderson, N.D. The effects of divided attention on encoding and retrieval processes in human memory. J. Exp. Psychol. Gen. 1996, 125, 159–180. [Google Scholar] [CrossRef]
- Troyer, A.K.; Winocur, G.; Craik, F.I.; Moscovitch, M. Source memory and divided attention: Reciprocal costs to primary and secondary tasks. Neuropsychology 1999, 13, 467–474. [Google Scholar] [CrossRef]
- Alfieri, P.; Cesarini, L.; De Rose, P.; Ricci, D.; Selicorni, A.; Menghini, D.; Guzzetta, A.; Baranello, G.; Tinelli, F.; Mallardi, M.; et al. Visual processing in Noonan syndrome: Dorsal and ventral stream sensitivity. Am. J. Med. Genet. Part A 2011, 155, 2459–2464. [Google Scholar] [CrossRef]
- Piccini, G.; Menghini, D.; D’Andrea, A.; Caciolo, C.; Pontillo, M.; Armando, M.; Perrino, F.; Mandolesi, L.; Salerni, A.; Buzzonetti, L.; et al. Visual perception skills: A comparison between patients with Noonan syndrome and 22q11.2 deletion syndrome. Genes Brain Behav. 2017, 16, 627–634. [Google Scholar] [CrossRef] [Green Version]
- Alfieri, P.; Piccini, G.; Caciolo, C.; Perrino, F.; Gambardella, M.L.; Mallardi, M.; Cesarini, L.; Leoni, C.; Leone, D.; Fossati, C.; et al. Behavioral profile in RASopathies. Am. J. Med. Genet. Part A 2014, 164, 934–942. [Google Scholar] [CrossRef]
- Perrino, F.; Licchelli, S.; Serra, G.; Piccini, G.; Caciolo, C.; Pasqualetti, P.; Cirillo, F.; Leoni, C.; Digilio, M.C.; Zampino, G.; et al. Psychopathological features in Noonan syndrome. Eur. J. Paediatr. Neurol. 2018, 22, 170–177. [Google Scholar] [CrossRef]
- Cattaneo, V.; San Martin, A.; Lew, S.E.; Gelb, B.D.; Pagani, M.R. Repeating or spacing learning sessions are strategies for memory improvement with shared molecular and neuronal components. Neurobiol. Learn. Mem. 2020. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Subject | Sex | Age | IQ | LQ | Disorder | Gene | Amino Acid Change |
|---|---|---|---|---|---|---|---|
| 1 | M | 6.08 | 90 | 84 | NS | SOS1 | Gly434Arg |
| 2 | F | 13 | 90 | 90 | NS | PTPN11 | Asn308Ser |
| 3 | F | 6.75 | 120 | 98 | NS | PTPN11 | Thr42Ala |
| 4 | F | 8.58 | 100 | 106 | NS | SOS1 | Ile733Phe |
| 5 | M | 6.42 | 120 | 84 | NS | PTPN11 | Ala72Ser |
| 6 | M | 10.08 | 102 | 94 | NS | RIT1 | Ser35Thr |
| 7 | F | 12.5 | 80 | 95 | NS | SOS1 | Arg552Lys |
| 8 | M | 13.75 | 100 | 111 | NS | PTPN11 | Met504Thr |
| 9 | F | 5.75 | 90 | 84 | NS | SOS1 | Arg552Ser |
| 10 | M | 6.25 | 110 | 89 | NS | PTPN11 | E139D |
| 11 | F | 5.75 | 130 | 102 | NS | SOS1 | Glu433Lys |
| 12 | M | 7.33 | 100 | 71 | NS | PTPN11 | Asn308Ser |
| 13 | F | 5.41 | 110 | 74 | NS | RAF1 | S257L |
| 14 | F | 7.66 | 90 | 73 | NS | PTPN11 | Phe285Ser |
| 15 | M | 13.08 | 120 | 80 | NS | PTPN11 | Gly503Arg |
| 16 | F | 7.42 | 101 | 120 | NSML | PTPN11 | Tyr279Ser |
| 17 | F | 12.33 | 125 | 122 | NSML | PTPN11 | Thr468Met |
| 18 | M | 9.08 | 120 | 108 | NSML | PTPN11 | Thr468Met |
| Type of Item | Inclusion Condition | Exclusion Condition |
|---|---|---|
| NS/NMLS | ||
| Colored drawings | 0.87 (0.19) | 0.87 (0.19) |
| Spoken words | 0.80 (0.17) | 0.08 (0.08) |
| New | 0.03 (0.05) | 0.02 (0.04) |
| TD | ||
| Colored drawings | 0.92 (0.12) | 0.89 (0.13) |
| Spoken words | 0.93 (0.08) | 0.05 (0.09) |
| New | 0.04 (0.07) | 0.03 (0.05) |
| Task | NS/NMLS | TD |
|---|---|---|
| SINGLE ITEM | ||
| Trial 1 | 0.93 (0.12) | 0.95 (0.14) |
| Trial 2 | 0.96 (0.08) | 0.98 (0.03) |
| Trial 3 | 0.96 (0.07) | 1.00 (0.00) |
| ASSOCIATIVE RECOGNITION | ||
| Trial 1 | 0.40 (0.15) | 0.49 (0.22) |
| Trial 2 | 0.50 (0.22) | 0.64 (0.20) |
| Trial 3 | 0.54 (0.25) | 0.75 (0.14) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costanzo, F.; Alfieri, P.; Caciolo, C.; Bergonzini, P.; Perrino, F.; Zampino, G.; Leoni, C.; Menghini, D.; Digilio, M.C.; Tartaglia, M.; et al. Recognition Memory in Noonan Syndrome. Brain Sci. 2021, 11, 169. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11020169
Costanzo F, Alfieri P, Caciolo C, Bergonzini P, Perrino F, Zampino G, Leoni C, Menghini D, Digilio MC, Tartaglia M, et al. Recognition Memory in Noonan Syndrome. Brain Sciences. 2021; 11(2):169. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11020169
Chicago/Turabian StyleCostanzo, Floriana, Paolo Alfieri, Cristina Caciolo, Paola Bergonzini, Francesca Perrino, Giuseppe Zampino, Chiara Leoni, Deny Menghini, Maria Cristina Digilio, Marco Tartaglia, and et al. 2021. "Recognition Memory in Noonan Syndrome" Brain Sciences 11, no. 2: 169. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11020169