
TMS–EEG Co-Registration in Patients with Mild Cognitive Impairment, Alzheimer’s Disease and Other Dementias: A Systematic Review

 ,
,
Abstract
:1. Introduction
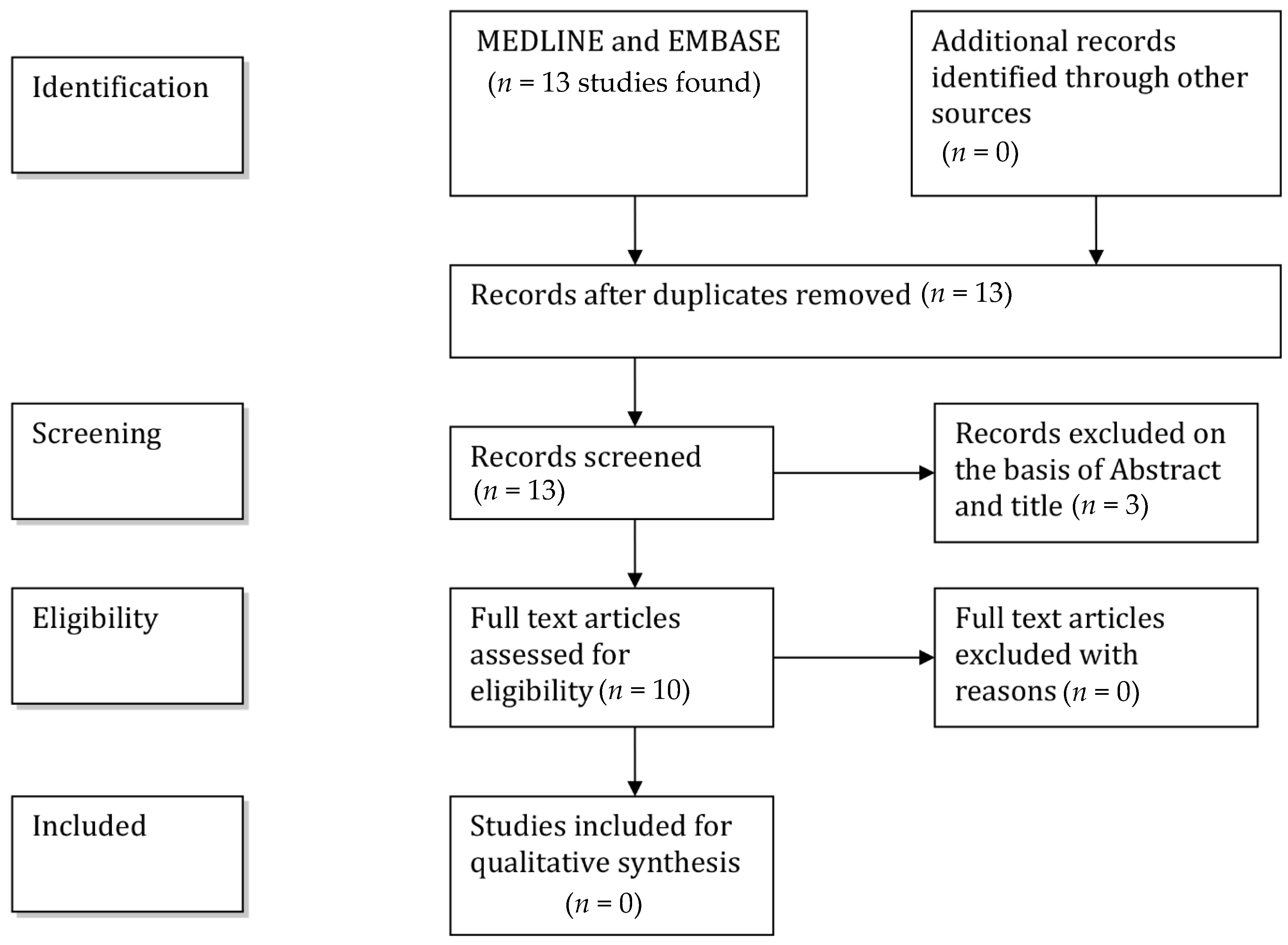
2. Methods
3. TMS–EEG Co-Registration Studies
3.1. TMS–EEG Studies Targeting the Motor Cortex
3.2. TMS–EEG Studies Targeting Other Cortical Areas
3.3. Evaluation of Drugs Effects by Means of TMS–EEG Co-Registration
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Hardy, J.A.; Mann, D.M.A.; Wester, P.; Winblad, B. An integrative hypothesis concerning the pathogenesis and progression of Alzheimer’s disease. Neurobiol. Aging 1986, 7, 489–502. [Google Scholar] [CrossRef]
- Grady, C.L.; Furey, M.L.; Pietrini, P.; Horwitz, B.; Rapoport, S.I. Altered brain functional connectivity and impaired short-term memory in Alzheimer’s disease. Brain 2001, 124, 739–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arendt, T. Synaptic degeneration in Alzheimer’s disease. Acta Neuropathol. 2009, 118, 167–179. [Google Scholar] [CrossRef]
- Grutzendler, J.; Helmin, K.; Tsai, J.; Gan, W.B. Various dendritic abnormalities are associated with fibrillar amyloid deposits in Alzheimer’s disease. Ann. N.Y. Acad. Sci. 2007, 1097, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Thut, G.; Ives, J.R.; Kampmann, F.; Pascual-Leone, A. A new device and protocol for combining TMS and online recordings of EEG and evoked potentials. J. Neurosci. Methods 2005, 141, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Ives, J.R.; Rotenberg, A.; Poma, R.; Thut, G.; Pascual-Leone, A. Electroencephalographic recording during transcranial magnetic stimulation in humans and animals. Clin. Neurophysiol. 2006, 117, 1870–1875. [Google Scholar] [CrossRef]
- Thut, G.; Pascual-Leone, A. A review of combined TMS-EEG studies to characterize lasting effects of repetitive TMS and assess their usefulness in cognitive and clinical neuroscience. Brain Topogr. 2010, 22, 219–232. [Google Scholar] [CrossRef] [Green Version]
- McMackin, R.; Muthuraman, M.; Groppa, S.; Babiloni, C.; Taylor, J.P.; Kiernan, M.C.; Nasseroleslami, B.; Hardiman, O. Measuring network disruption in neurodegenerative diseases: New approaches using signal analysis. J. Neurol. Neurosurg. Psychiatry 2019, 90, 1011–1020. [Google Scholar] [CrossRef]
- Mäki, H.; Ilmoniemi, R.J. The relationship between peripheral and early cortical activation induced by transcranial magnetic stimulation. Neurosci. Lett. 2010, 478, 24–28. [Google Scholar] [CrossRef]
- Raij, T.; Karhu, J.; Kicić, D.; Lioumis, P.; Julkunen, P.; Lin, F.H.; Ahveninen, J.; Ilmoniemi, R.J.; Mäkelä, J.P.; Hämäläinen, M.; et al. Parallel input makes the brain run aster. NeuroImage 2008, 40, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Komssi, S.; Aronen, H.J.; Huttunen, J.; Kesäniemi, M.; Soinne, L.; Nikouline, V.V.; Ollikainen, M.; Roine, R.O.; Karhu, J.; Savolainen, S.; et al. Ipsi- and contralateral EEG reactions to transcranial magnetic stimulation. Clin. Neurophysiol. 2002, 113, 175–184. [Google Scholar] [CrossRef]
- Massimini, M.; Ferrarelli, F.; Huber, R.; Esser, S.K.; Singh, H.; Tononi, G. Neuroscience: Breakdown of cortical effective connectivity during sleep. Science 2005, 309, 2228–2232. [Google Scholar] [CrossRef] [PubMed]
- Rosanova, M.; Casali, A.; Bellina, V.; Resta, F.; Mariotti, M.; Massimini, M. Natural frequencies of human corticothalamic circuits. J. Neurosci. 2009, 29, 7679–7685. [Google Scholar] [CrossRef]
- Ferreri, F.; Pasqualetti, P.; Määttä, S.; Ponzo, D.; Ferrarelli, F.; Tononi, G.; Mervaala, E.; Miniussi, C.; Rossini, P.M. Human brain connectivity during single and paired pulse transcranial magnetic stimulation. NeuroImage 2011, 54, 90–102. [Google Scholar] [CrossRef]
- Ilmoniemi, R.J.; Virtanen, J.; Ruohonen, J.; Karhu, J.; Aronen, H.J.; Näätänen, R.; Katila, T. Neuronal responses to magnetic stimulation reveal cortical reactivity and connectivity. Neuroreport 1997, 8, 3537–3540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paus, T.; Sipilä, P.K.; Strafella, A.P. Synchronization of neuronal activity in the human primary motor cortex by transcranial magnetic stimulation: An EEG study. J. Neurophysiol. 2001, 86, 1983–1990. [Google Scholar] [CrossRef] [Green Version]
- Komssi, S.; Kähkönen, S.; Ilmoniemi, R.J. The effect of stimulus intensity on brain responses evoked by transcranial magnetic stimulation. Hum. Brain Mapp. 2004, 21, 154–164. [Google Scholar] [CrossRef]
- Van Der Werf, Y.D.; Sadikot, A.F.; Strafella, A.P.; Paus, T. The neural response to transcranial magnetic stimulation of the human motor cortex. II. Thalamocortical contributions. Exp. Brain Res. 2006, 175, 246–255. [Google Scholar] [CrossRef]
- Ferreri, F.; Ponzo, D.; Hukkanen, T.; Mervaala, E.; Könönen, M.; Pasqualetti, P.; Vecchio, F.; Rossini, P.M.; Määttä, S. Human brain cortical correlates of short-latency afferent inhibition: A combined EEG-TMS study. J. Neurophysiol. 2012, 108, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, F.; Vecchio, F.; Ponzo, D.; Pasqualetti, P.; Rossini, P.M. Time-varying coupling of EEG oscillations predicts excitability fluctuations in the primary motor cortex as reflected by motor evoked potentials amplitude: An EEG-TMS study. Hum. Brain Mapp. 2014, 35, 1969–1980. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, F.; Rossini, P.M. TMS and TMS-EEG techniques in the study of the excitability, connectivity, and plasticity of the human motor cortex. Rev. Neurosci. 2013, 24, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Rossini, P.M.; Rossi, S.; Babiloni, C.; Polich, J. Clinical neurophysiology of aging brain: From normal aging to neurodegeneration. Prog. Neurobiol. 2007, 83, 375–400. [Google Scholar] [CrossRef] [PubMed]
- Guerra, A.; Assenza, F.; Bressi, F.; Scrascia, F.; Del Duca, M.; Ursini, F.; Vollaro, S.; Trotta, L.; Tombini, M.; Chisari, C.; et al. Transcranial magnetic stimulation studies in Alzheimer’s disease. Int. J. Alzheimers Dis. 2011, 2011, 263817. [Google Scholar] [CrossRef] [Green Version]
- Pascual-Leone, A.; Freitas, C.; Oberman, L.; Horvath, J.C.; Halko, M.; Eldaief, M.; Bashir, S.; Vernet, M.; Shafi, M.; Westover, B.; et al. Characterizing brain cortical plasticity and network dynamics across the age-span in health and disease with TMS-EEG and TMS-fMRI. Brain Topogr. 2011, 24, 302–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yener, G.G.; Başar, E. Biomarkers in Alzheimer’s disease with a special emphasis on event-related oscillatory responses. Suppl. Clin. Neurophysiol. 2013, 62, 237–273. [Google Scholar]
- Higgins, J.P.T.; Green, S. Cochrane Handbook for Systematic Reviews of Interventions4.2.5 [Updated May 2005]; Higgins, J.P.T., Green, S., Eds.; John Wiley and Sons: Hoboken, NJ, USA, 2005; Volume 3. [Google Scholar]
- Julkunen, P.; Jauhiainen, A.M.; Westerén-Punnonen, S.; Pirinen, E.; Soininen, H.; Könönen, M.; Pääkkönen, A.; Määttä, S.; Karhu, J. Navigated TMS combined with EEG in mild cognitive impairment and Alzheimer’s disease: A pilot study. J. Neurosci. Methods 2008, 172, 270–276. [Google Scholar] [CrossRef]
- Julkunen, P.; Jauhiainen, A.M.; Könönen, M.; Pääkkönen, A.; Karhu, J.; Soininen, H. Combining transcranial magnetic stimulation and electroencephalography may contribute to assess the severity of Alzheimer’s disease. Int. J. Alzheimers Dis. 2011, 2011, 654794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casarotto, S.; Määttä, S.; Herukka, S.K.; Pigorini, A.; Napolitani, M.; Gosseries, O.; Niskanen, E.; Könönen, M.; Mervaala, E.; Rosanova, M.; et al. Transcranial magnetic stimulation-evoked EEG/cortical potentials in physiological and pathological aging. Neuroreport 2011, 22, 592–597. [Google Scholar] [CrossRef] [Green Version]
- Ferreri, F.; Vecchio, F.; Vollero, L.; Guerra, A.; Petrichella, S.; Ponzo, D.; Määtta, S.; Mervaala, E.; Könönen, M.; Ursini, F.; et al. Sensorimotor cortex excitability and connectivity in Alzheimer’s disease: A TMS-EEG Co-registration study. Hum. Brain Mapp. 2016, 37, 2083–2096. [Google Scholar] [CrossRef]
- Gandelman-Marton, R.; Aichenbaum, S.; Dobronevsky, E.; Khaigrekht, M.; Rabey, J.M. Quantitative EEG after Brain Stimulation and Cognitive Training in Alzheimer Disease. J. Clin. Neurophysiol. 2017, 34, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Zomorrodi, R.; Ghazala, Z.; Goodman, M.S.; Blumberger, D.M.; Cheam, A.; Fischer, C.; Daskalakis, Z.J.; Mulsant, B.H.; Pollock, B.G.; et al. Extent of Dorsolateral Prefrontal Cortex Plasticity and Its Association with Working Memory in Patients with Alzheimer Disease. JAMA Psychiatry 2017, 74, 1266–1274. [Google Scholar] [CrossRef]
- Koch, G.; Motta, C.; Bonnì, S.; Pellicciari, M.C.; Picazio, S.; Casula, E.P.; Maiella, M.; Di Lorenzo, F.; Ponzo, V.; Ferrari, C.; et al. Effect of Rotigotine vs Placebo on Cognitive Functions among Patients With Mild to Moderate Alzheimer Disease: A Randomized Clinical Trial. JAMA Netw. Open 2020, 3, e2010372. [Google Scholar] [CrossRef] [PubMed]
- Bagattini, C.; Mutanen, T.P.; Fracassi, C.; Manenti, R.; Cotelli, M.; Ilmoniemi, R.J.; Miniussi, C.; Bortoletto, M. Predicting Alzheimer’s disease severity by means of TMS-EEG coregistration. Neurobiol. Aging 2019, 80, 38–45. [Google Scholar] [CrossRef]
- Assogna, M.; Casula, E.P.; Borghi, I.; Bonnì, S.; Samà, D.; Motta, C.; Di Lorenzo, F.; D’Acunto, A.; Porrazzini, F.; Minei, M.; et al. Effects of Palmitoylethanolamide Combined with Luteoline in Frontal Lobe Functions, High Frequency Oscillations, and GABAergic Transmission in Patients with Frontotemporal Dementia. J. Alzheimers Dis. 2020, 76, 1297–1308. [Google Scholar] [CrossRef]
- Bonato, C.; Miniussi, C.; Rossini, P.M. Transcranial magnetic stimulation and cortical evoked potentials: A TMS/EEG co-registration study. Clin. Neurophysiol. 2006, 117, 1699–1707. [Google Scholar] [CrossRef] [PubMed]
- Daskalakis, Z.J.; Moller, B.; Christensen, B.K.; Fitzgerald, P.B.; Gunraj, C.; Chen, R. The effects of repetitive transcranial magnetic stimulation on cortical inhibition in healthy human subjects. Exp. Brain Res. 2006, 174, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Hallett, M. Transcranial magnetic stimulation: A primer. Neuron 2007, 55, 187–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, T.; Valero-Cabre, A.; Pascual-Leone, A. Noninvasive human brain stimulation. Annu. Rev. Biomed. Eng. 2007, 9, 527–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, G.; Bonnì, S.; Pellicciari, M.C.; Casula, E.P.; Mancini, M.; Esposito, R.; Ponzo, V.; Picazio, S.; Di Lorenzo, F.; Serra, L.; et al. Transcranial magnetic stimulation of the precuneus enhances memory and neural activity in prodromal Alzheimer’s disease. Neuroimage 2018, 169, 302–311. [Google Scholar] [CrossRef] [Green Version]
- Stefan, K.; Kunesch, E.; Benecke, R.; Cohen, L.G. Mechanisms of enhancement of human motor cortex excitability induced by interventional paired associative stimulation. J. Physiol. 2002, 43, 699–708. [Google Scholar] [CrossRef]
- Delbeuck, X.; Van der Linden, M. Alzheimer’Disease as a disconnection syndrome? Neuropsychol. Rev. 2003, 13, 79–92. [Google Scholar] [CrossRef]
- Tiitinen, H.; Virtanen, J.; Ilmoniemi, R.J.; Kamppuri, J.; Ollikainen, M.; Ruohonen, J.; Näätänen, R. Separation of contamination caused by coil clicks from responses elicited by transcranial magnetic stimulation. Clin. Neurophysiol. 1999, 110, 982–985. [Google Scholar] [CrossRef]
- Koenig, T.; Prichep, L.; Dierks, T.; Hubl, D.; Wahlund, L.O.; John, E.R.; Jelic, V. Decreased EEG synchronization in Alzheimer’s disease and mild cognitive impairment. Neurobiol. Aging 2005, 6, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Nikouline, V.; Ruohonen, J.; Ilmoniemi, R.J. The role of the coil click in TMS assessed with simultaneous EEG. Clin. Neurophysiol. 1999, 110, 1325–1328. [Google Scholar] [CrossRef]
- Sanes, J.N.; Donoghue, J.P. Plasticity and primary motor cortex. Annu. Rev. Neurosci. 2000, 23, 393–415. [Google Scholar] [CrossRef] [Green Version]
- Basso, D.; Lotze, M.; Vitale, L.; Ferreri, F.; Bisiacchi, P.; Olivetti Belardinelli, M.; Rossini, P.M.; Birbaumer, N. The role of prefrontal cortex in visuo-spatial planning: A repetitive TMS study. Exp. Brain Res. 2006, 171, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Di Pino, G.; Pellegrino, G.; Assenza, G.; Capone, F.; Ferreri, F.; Formica, D.; Ranieri, F.; Tombini, M.; Ziemann, U.; Rothwell, J.C.; et al. Modulation of brain plasticity in stroke: Anovel model for neurorehabilitation. Nat. Rev. Neurol. 2014, 10, 597–608. [Google Scholar] [CrossRef]
- Ferreri, F.; Curcio, G.; Pasqualetti, P.; De Gennaro, L.; Fini, R.; Rossini, P.M. Mobile phone emissions and human brain excitability. Ann. Neurol. 2006, 60, 188–196. [Google Scholar] [CrossRef]
- Sharbrough, F.W. Electrical fields and recording techniques. In Current Practice of Clinical Electroencephalography; Daly, D.D., Pedley, T.A., Eds.; Lippincott-Raven: Philadelphia, PA, USA, 1997; pp. 29–49. [Google Scholar]
- Babiloni, C.; Infarinato, F.; Aujard, F.; Bastlund, J.F.; Bentivoglio, M.; Bertin, G.; Del Percio, C.; Fabene, P.F.; Forloni, G.; Herrero Ezquerro, M.T.; et al. Effects of pharmacological agents, sleep deprivation hypoxia and transcranial magnetic stimulation on electroencephalographic rhythms in rodents: Towards translational challenge models for drug discovery in Alzheimer’s disease. Clin. Neurophysiol. 2013, 124, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Steriade, M. Cellular substrates of brain rhythms. In Electroencephalogaphy; Basic Principles, Clinical Applications, Related Fields; Niedermeyer, E., Lopes Da Silva, F., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1999; pp. 28–69. [Google Scholar]
- Mesulam, M.; Shaw, P.; Mash, D.; Weintraub, S. Cholinergic nucleus basalis tauopathy emerges early in the aging-MCI-AD continuum. Ann. Neurol. 2004, 55, 815–828. [Google Scholar] [CrossRef] [PubMed]
- Schreiter-Gasser, U.; Gasser, T.; Ziegler, P. Quantitative EEG analysis in early onset Alzheimer’s disease: Correlations with severity, clinical characteristics, visual EEG and CCT. Electroencephalogr. Clin. Neurophysiol 1994, 90, 267–272. [Google Scholar] [CrossRef]
- Agosta, F.; Pievani, M.; Geroldi, C.; Copetti, M.; Frisoni, G.B.; Filippi, M. Resting state fMRI in Alzheimer’s disease: Beyond the default mode network. Neurobiol. Aging 2012, 33, 1564–1578. [Google Scholar] [CrossRef]
- Zhang, Y.; Simon-Vermot, L.; Araque Caballero, M.T.; Gesierich, B.; Taylor, A.N.W.; Duering, M.; Dichgans, M.; Ewers, M. Enhanced resting-state functional connectivity between core memory-task activation peaks is associated with memory impairment in MCI. Neurobiol. Aging 2016, 45, 43–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opitz, A.; Fox, M.D.; Craddock, R.C.; Colcombe, S.; Milham, M.P. An integratedframework for targeting functional networks via transcranial magnetic stimulation. Neuroimage 2016, 127, 86–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckner, R.L.; Andrews-Hanna, J.R.; Schacter, D.L. The brain’s default network: Anatomy, function, and relevance to disease. Ann. N. Y. Acad. Sci. 2008, 1124, 1–38. [Google Scholar] [CrossRef] [Green Version]
- Raichle, M.E.; MacLeod, A.M.; Snyder, A.Z.; Powers, W.J.; Gusnard, D.A.; Shulman, G.L. A default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Feurra, M.; Galli, G.; Pavone, E.F.; Rossi, A.; Rossi, S. Frequency-specific insight into short-term memory capacity. J. Neurophysiol. 2016, 116, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Dubois, B.; Hampel, H.; Feldman, H.H.; Scheltens, P.; Aisen, P.; Andrieu, S.; Bakardjian, H.; Benali, H.; Bertram, L.; Blennow, K.; et al. Preclinical Alzheimer’s disease: Definition, natural history, and diagnostic criteria. Alzheimers Dement. 2016, 12, 292–323. [Google Scholar] [CrossRef]
- Bai, W.; Liu, T.; Dou, M.; Xia, M.; Lu, J.; Tian, X. Repetitive Transcranial Magnetic Stimulation Reverses Aβ1-42-induced Dysfunction in Gamma Oscillation during Working Memory. Curr. Alzheimer Res. 2018, 6, 570–577. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Studies | Nr Disease | Age (y) | Gender F/M | Education (y) | DD | MSE | Principal Neurophysiological Findings |
|---|---|---|---|---|---|---|---|
| Jalkunen et al., 2008 [27] | 5 MCI | 74.0 ± 8.1 | 2/3 | 7.6 ± 3.0 | ? | 25.4 ± 3.2 | Significantly reduced P30 in the ipsilateral temporo-parietal area |
| 5 AD | 73.26 ± 8.1 | 2/3 | 9.6 ± 2.1 | 22.0 ± 5.1 | and in the contralateral centro-frontal cortex | ||
| Jalkunen et al., 2011 [28] | 5 MCI | 74 ± 8 | 2/3 | 7.6 ± 3.0 | ? | ? | P30 amplitude correlated with cognitive decline and showed good specificity |
| 5 AD | 73 ± 8 | 9.6 ± 2.1 | ? | ? | and sensitivity in identifying subjects with MCI or AD | ||
| Casarotto et al., 2011 [29] | 9 AD | 72 ± 7.1 | 5/4 | 10 ± 3.5 | ? | 18 ± 4.5 | The EEG response to TMS of the left superior frontal cortex is not |
| affected by physiological aging but is altered by cognitive impairment | |||||||
| Ferreri et al., 2016 [30] | 12 AD | 72.4 ± 5.9 | 7/5 | 8 ± 3.5 | ? | 20.8 ± 2.7 | The sensorimotor system is hyperexcitable in patients with AD |
| Gandelman-Marton et al., 2017 [31] | 7 AD | 75.5 ± 4.3 | 1/7 | 11.6 ± 2.7 | 2.8 ± 1 | ? | After rTMS significant increase in delta activity over temporal region |
| Positive correlation between alfa power and MMSE | |||||||
| Kumar et al., 2017 [32] | 32 AD | 76.3 ± 6.3 | 17/15 | 13.5 ± 3.8 | ? | 22.6 ± 3.2 | DLPFC plasticity in patients with AD |
| Koch et al. 2018 [33] | 14 AD | 70.0 ± 5.1 | 7/7 | 7.2 ± 3.0 | ? | 26.1 ± 1.8 | rTMS increased neural activity in PC, enhanced brain oscillations in the beta |
| band and modified functional connections between PC and frontal areas | |||||||
| Bagattini et al., 2019 [34] | 26 AD | 76.5 ± 4.7 | 23/3 | 7.7 ± 4.1 | ? | 20.8 ± 2.5 | Higher P30 amplitudes predicted poorer cognitive performances |
| Koch et al., 2020 [33] | 94 AD | 73.9 ± 5.6 | 58/36 | 1 8.5 ± 4.2 | ? | 1 22.9 ± 2,3 | Rotigotine increases frontal cortical activity, but did not significantly affect |
| 2 9.4 ± 4.3 | 2 23.6 ± 2.4 | global cognition | |||||
| Assogna et al., 2020 [35] | 17 AD | 62.35 ± 9.4 | 11/6 | 12.47 ± 3.41 | 2.6 ± 1.3 | ? | Treatment with PEA-LUT restores LICI at ISI 100 ms, increases TMS-evoked |
| frontal lobe activity and high-frequency oscillations in beta/gamma range |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nardone, R.; Sebastianelli, L.; Versace, V.; Ferrazzoli, D.; Saltuari, L.; Trinka, E. TMS–EEG Co-Registration in Patients with Mild Cognitive Impairment, Alzheimer’s Disease and Other Dementias: A Systematic Review. Brain Sci. 2021, 11, 303. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11030303
Nardone R, Sebastianelli L, Versace V, Ferrazzoli D, Saltuari L, Trinka E. TMS–EEG Co-Registration in Patients with Mild Cognitive Impairment, Alzheimer’s Disease and Other Dementias: A Systematic Review. Brain Sciences. 2021; 11(3):303. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11030303
Chicago/Turabian StyleNardone, Raffaele, Luca Sebastianelli, Viviana Versace, Davide Ferrazzoli, Leopold Saltuari, and Eugen Trinka. 2021. "TMS–EEG Co-Registration in Patients with Mild Cognitive Impairment, Alzheimer’s Disease and Other Dementias: A Systematic Review" Brain Sciences 11, no. 3: 303. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11030303