
Brain Plasticity Mechanisms Underlying Motor Control Reorganization: Pilot Longitudinal Study on Post-Stroke Subjects

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Set-up
2.3. Patients Functional Assessment and Foot Drop Rehabilitation Procedures
2.4. fMRI Paradigm
2.5. fMRI Image Acquisition and Preprocessing
2.6. Regions of Interest (ROIs) Selection and Extraction
- driving input representing descending voluntary signals—V (combined onsets of the two conditions requiring volitional effort);
- modulatory input encoding the contribution of ascending functional electrical stimulation to proprioceptive input—E (combined onsets of the two conditions requiring stimulation);
- driving input representing underlying proprioceptive input from all movements—P (onsets from all conditions).
- Angular gyrus (AG). AG is assumed to be a multi-sensorial integration hub [28], also able to process discrepancies between intended action and movement consequences, in such a way that they can be consciously detected by the subject [29]. Moreover, in an earlier study [13], AG and SMA have been shown to be differently active in responders and non-responders.
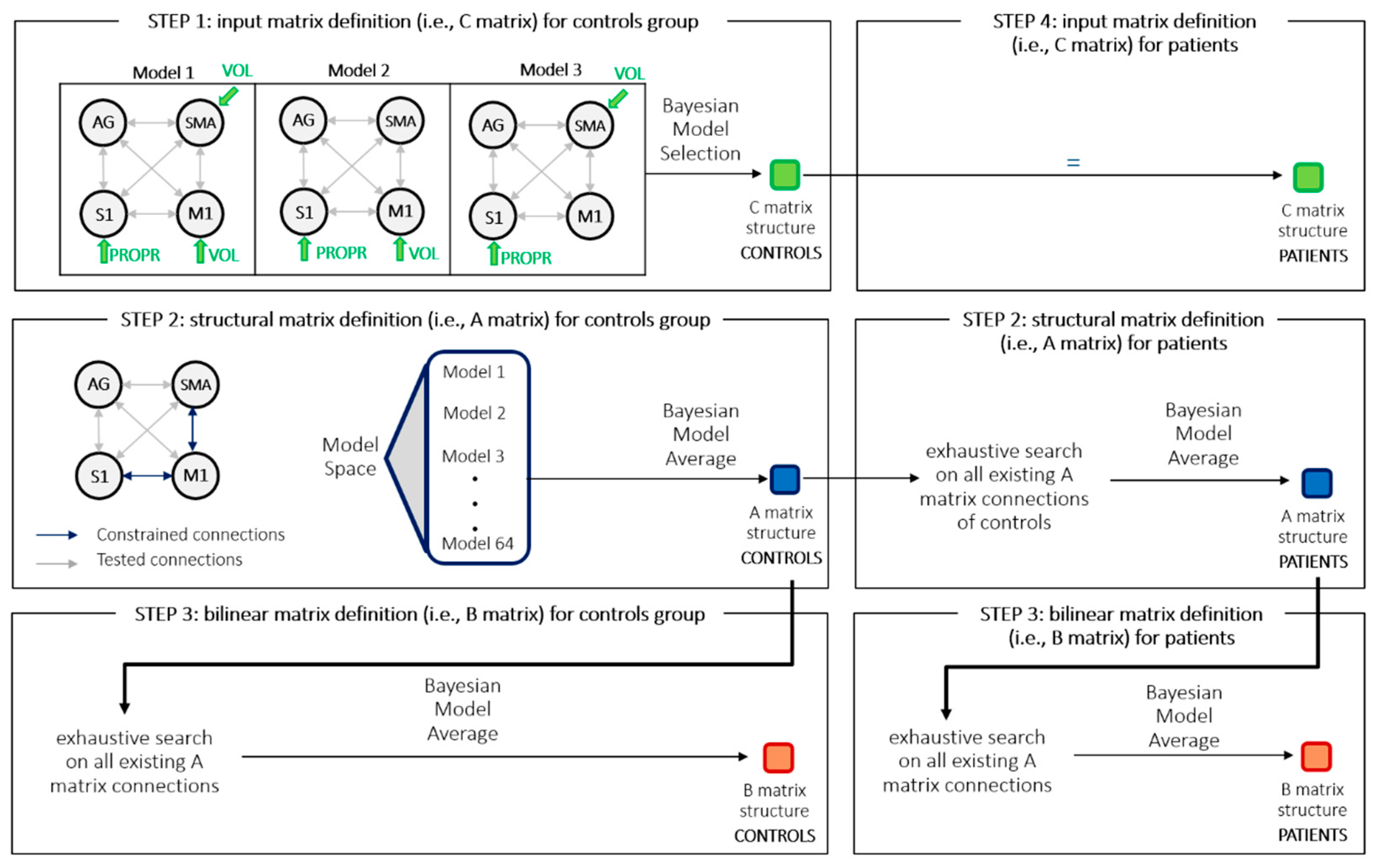
2.7. Dynamic Causal Modeling (DCM) Analysis
- matrix A—average connectivity among brain regions during the experiment, irrespective of task modulation (endogenous connections).
- matrix B—change in endogenous connections that can be elicited by an experimental variable (modulatory inputs).
- matrix C—direct influences of an experimental variable on specific regions (driving inputs).
2.8. Parametric Empirical Bayes (PEB) Model
2.9. Model Structure Identification Procedure
3. Results
3.1. Participants
3.2. Activation Patterns for Healthy and Patients’ Groups (Previous Results)
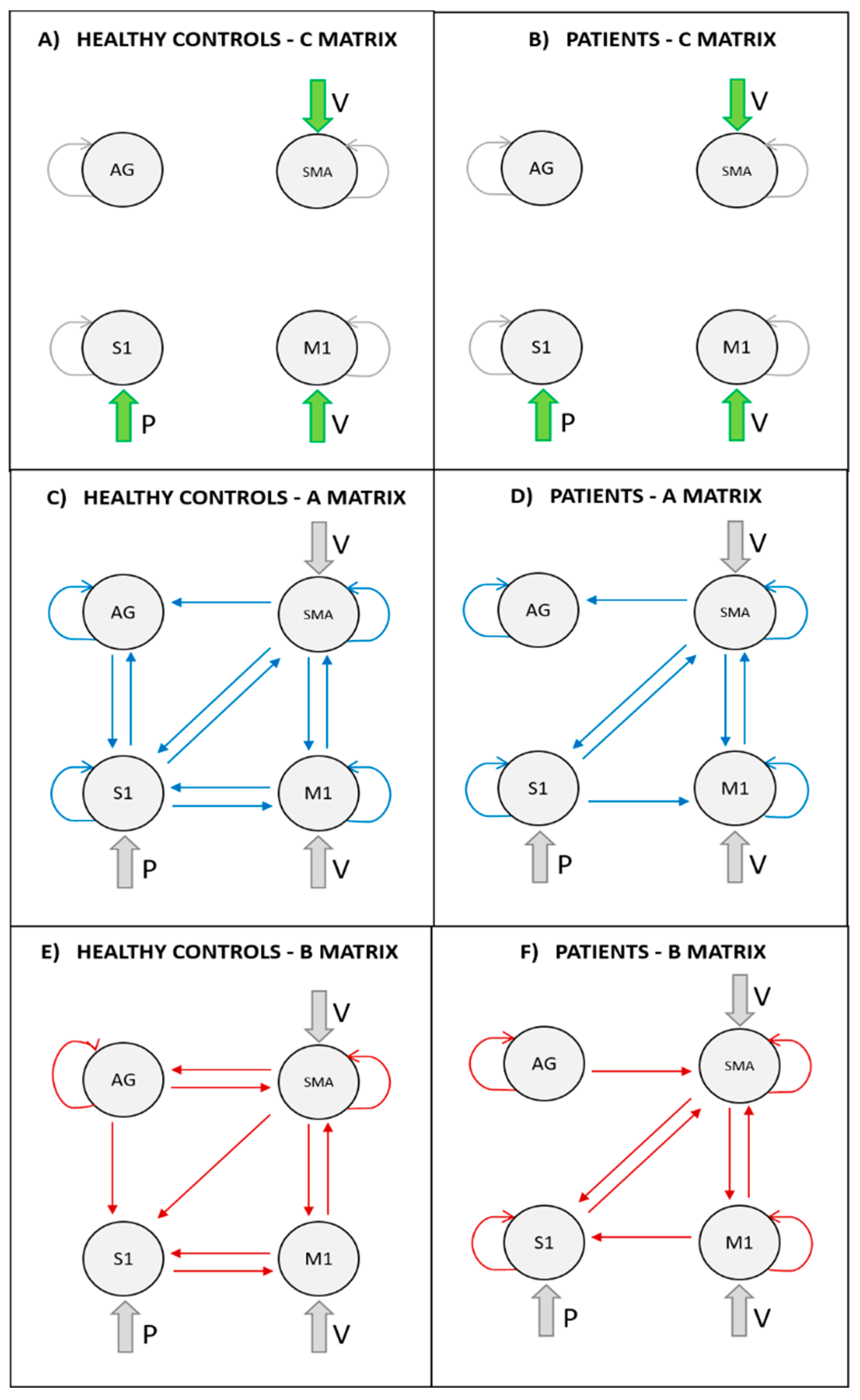
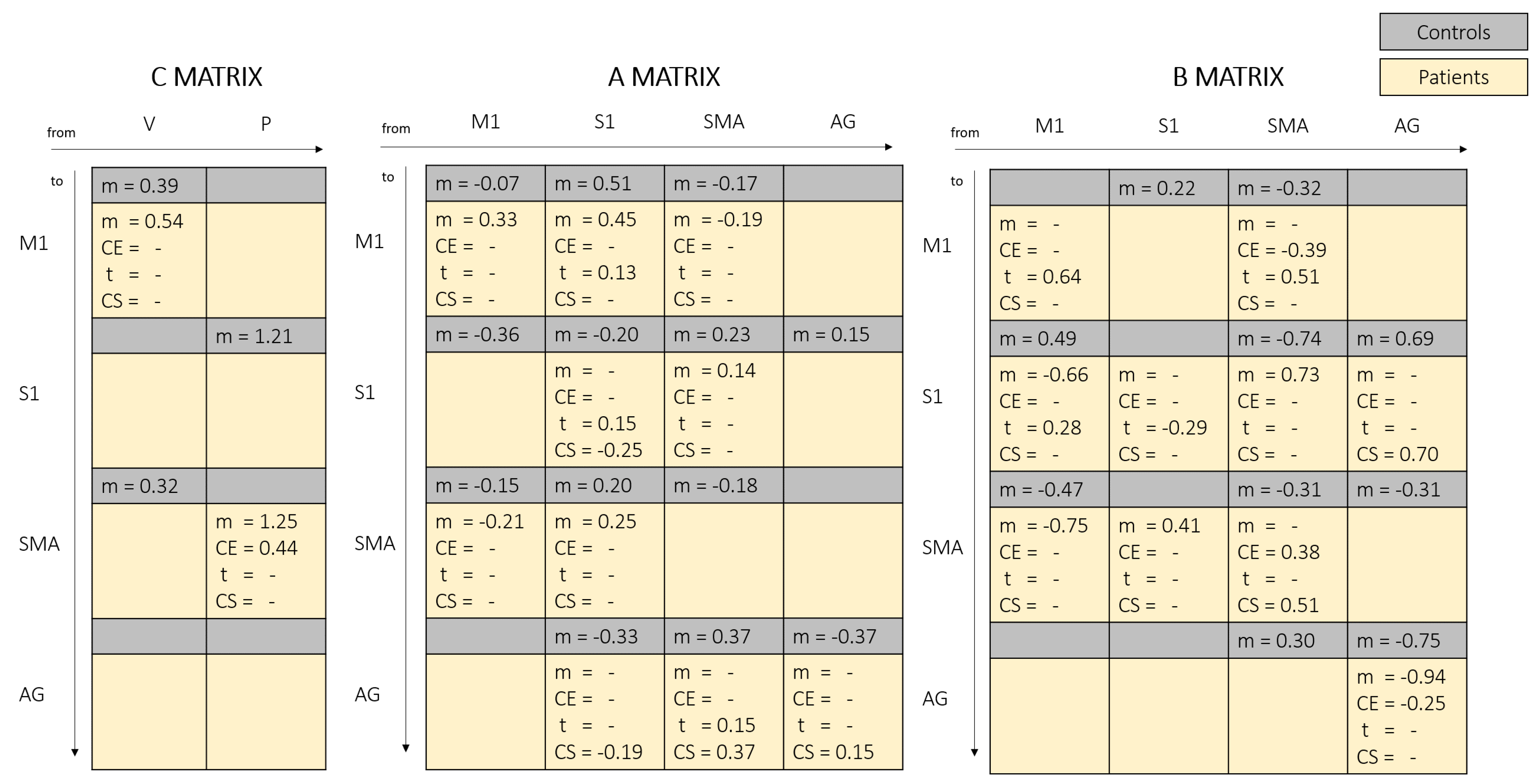
3.3. DCM Results
4. Discussion
4.1. Proprioceptive Input to S1 Area
4.2. Influence of SMA and M1 over S1
4.3. Outcoming Connections from S1
4.4. SMA-M1 Bilateral Connections
4.5. Role of AG Area
4.6. Study Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AG | angular gyrus |
| CE | carry-over group/responders |
| DCM | Dynamic Causal Modelling |
| FES | Functional Electrical Stimulation |
| fMRI | functional Magnetic Resonance Imaging |
| M1 | Primary motor cortex |
| nCE | non-responders |
| PEB | Parametric Empirical Bayes |
| ROI | Region of Interest |
| S1 | primary somatosensory cortex |
| SMA | supplementary motor area |
References
- Takeuchi, N.; Izumi, S.-I. Rehabilitation with Poststroke Motor Recovery: A Review with a Focus on Neural Plasticity Stroke Research and Treatment 2013. Available online: https://www.hindawi.com/journals/srt/2013/128641/ (accessed on 9 January 2019).
- Tacchino, G.; Gandolla, M.; Coelli, S.; Barbieri, R.; Pedrocchi, A.; Bianchi, A.M. EEG Analysis During Active and Assisted Repetitive Movements: Evidence for Differences in Neural Engagement. IEEE Trans. Neural Syst. Rehabil. Eng. 2017, 25, 761–771. [Google Scholar] [CrossRef]
- Burridge, J.H.; Taylor, P.N.; Hagan, S.A.; Wood, D.E.; Swain, I.D. The effects of common peroneal stimulation on the effort and speed of walking: A randomized controlled trial with chronic hemiplegic patients. Clin. Rehabil. 1997, 11, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Liberson, W.T.; Holmquest, H.J.; Scot, D.; Dow, M. Functional electrotherapy: Stimulation of the peroneal nerve synchronized with the swing phase of the gait of hemiplegic patients. Arch Phys Med. 1961, 42, 101–105. [Google Scholar]
- Merletti, R.; Andina, A.; Galante, M.; Furlan, I. Clinical experience of electronic peroneal stimulators in 50 hemiparetic patients. Scand. J. Rehabil. Med. 1979, 11, 111–121. [Google Scholar] [PubMed]
- Gandolla, M.; Molteni, F.; Ward, N.S.; Guanziroli, E.; Ferrigno, G.; Pedrocchi, A. Validation of a quantitative single-subject based evaluation for rehabilitation-induced improvement assessment. Ann. Biomed. Eng. 2015, 43, 2686–2698. [Google Scholar] [CrossRef] [PubMed]
- Ambrosini, E. A myocontrolled neuroprosthesis integrated with a passive exoskeleton to support upper limb activities. J. Electromyogr. Kinesiol. 2014, 24, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Schaechter, J.D. Motor rehabilitation and brain plasticity after hemiparetic stroke. Prog. Neurobiol. 2004, 73, 61–72. [Google Scholar] [CrossRef]
- Li, S. Spasticity, Motor Recovery, and Neural Plasticity after Stroke. Front. Neurol. 2017, 8, 120. [Google Scholar] [CrossRef] [Green Version]
- Iftime-Nielsen, S.D.; Christensen, M.S.; Vingborg, R.J.; Sinkjaer, T.; Roepstorff, A.; Grey, M.J. Interaction of electrical stimulation and voluntary hand movement in SII and the cerebellum during simulated therapeutic functional electrical stimulation in healthy adults. Hum. Brain Mapp. 2012, 33, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.S.; Grey, M.J. Modulation of proprioceptive feedback during functional electrical stimulation: An fMRI study. Eur. J. Neurosci. 2013, 37, 1766–1778. [Google Scholar] [CrossRef]
- Gandolla, M.; Ferrante, S.; Molteni, F.; Guanziroli, E.; Frattini, T.; Martegani, A.; Ferrigno, G.; Friston, K.; Pedrocchi, A.; Ward, N.S. Re-thinking the role of motor cortex: Context-sensitive motor outputs? NeuroImage 2014, 91, 366–374. [Google Scholar] [CrossRef] [Green Version]
- Gandolla, M.; Ward, N.S.; Molteni, F.; Guanziroli, E.; Ferrigno, G.; Pedrocchi, A. The Neural Correlates of Long-Term Carryover following Functional Electrical Stimulation for Stroke. Neural Plast. 2016, 4192718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolpert, D.M.; Flanagan, J.R. Motor prediction. Curr. Biol. 2001, 11, R729–R732. [Google Scholar] [CrossRef] [Green Version]
- Sejnowski, T.; Tesauro, G. The Hebb Rule for Synaptic Plasticity: Algorithms and Implementations. Neural Models Plast. 1989, 94–103. [Google Scholar] [CrossRef]
- Rushton, D.N. Functional electrical stimulation and rehabilitation--an hypothesis. Med Eng. Phys. 2003, 25, 75–78. [Google Scholar] [CrossRef]
- Rosenkranz, K.; Rothwell, J.C. Modulation of proprioceptive integration in the motor cortex shapes human motor learning. J. Neurosci. 2012, 32, 9000–9006. [Google Scholar] [CrossRef] [Green Version]
- Leonard, N.R.; Jha, A.P.; Casarjian, B.; Goolsarran, M.; Garcia, C.; Cleland, C.M.; Gwadz, M.V.; Massey, Z. Mindfulness training improves attentional task performance in incarcerated youth: A group randomized controlled intervention trial. Front. Psychol. 2013, 4, 792. [Google Scholar] [CrossRef] [Green Version]
- Ageranioti-Bélanger, S.A.; Chapman, C.E. Discharge properties of neurones in the hand area of primary somatosensory cortex in monkeys in relation to the performance of an active tactile discrimination task. II. Area 2 as compared to areas 3b and 1. Exp. Brain Res. 1992, 91, 207–228. [Google Scholar] [CrossRef] [PubMed]
- Casellato, C.; Ferrante, S.; Gandolla, M.; Volonterio, N.; Ferrigno, G.; Baselli, G.; Frattini, T.; Martegani, A.; Molteni, F.; Pedrocchi, A.A. Simultaneous measurements of kinematics and fMRI: Compatibility assessment and case report on recovery evaluation of one stroke patient. J. Neuroeng. Rehabil. 2010, 7, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandolla, M.; Ferrante, S.; Casellato, C.; Ferrigno, G.; Molteni, F.; Martegani, A.; Frattini, T.; Pedrocchi, A. fMRI brain mapping during motion capture and FES induced motor tasks: Signal to noise ratio assessment. Med. Eng. Phys. 2011, 33, 1027–1032. [Google Scholar] [CrossRef]
- Burridge, J.H.; Wood, D.E.; Taylor, P.N.; McLellan, D.L. Indices to describe different muscle activation patterns, identified during treadmill walking, in people with spastic drop-foot. Med. Eng. Phys. 2001, 23, 427–434. [Google Scholar] [CrossRef]
- Friston, K.J.; Harrison, L.; Penny, W. Dynamic causal modelling. NeuroImage 2003, 19, 1273–1302. [Google Scholar] [CrossRef]
- Shadmehr, R.; Smith, M.A.; Krakauer, J.W. Error correction, sensory prediction, and adaptation in motor control. Annu. Rev. Neurosci. 2010, 33, 89–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flanders, M. What is the biological basis of sensorimotor integration? Biol. Cybern. 2011, 104, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orgogozo, J.M.; Larsen, B. Activation of the supplementary motor area during voluntary movement in man suggests it works as a supramotor area. Science 1979, 206, 847–850. [Google Scholar] [CrossRef]
- Narayana, S.; Laird, A.R.; Tandon, N.; Franklin, C.; Lancaster, J.L.; Fox, P.T. Electrophysiological and Functional Connectivity of the Human Supplementary Motor Area. Neuroimage 2012, 62, 250–265. [Google Scholar] [CrossRef] [Green Version]
- Seghier, M.L. The angular gyrus: Multiple functions and multiple subdivisions. Neurosci. 2013, 19, 43–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrer, C.; Frey, S.H.; Van Horn, J.D.; Tunik, E.; Turk, D.; Inati, S.; Grafton, S.T. The angular gyrus computes action awareness representations. Cereb. Cortex 2008, 18, 254–261. [Google Scholar] [CrossRef]
- Penny, W.D.; Stephan, K.E.; Mechelli, A.; Friston, K.J. Comparing dynamic causal models. Neuroimage 2004, 22, 1157–1172. [Google Scholar] [CrossRef]
- Friston, K.; Zeidman, P.; Litvak, V. Empirical Bayes for DCM: A Group Inversion Scheme. Front. Syst. Neurosci. 2015, 9, 164. [Google Scholar] [CrossRef] [Green Version]
- Friston, K.J.; Litvak, V.; Oswal, A.; Razi, A.; Stephan, K.E.; Van Wijk, B.C.M.; Ziegler, G.; Zeidman, P. Bayesian model reduction and empirical Bayes for group (DCM) studies. Neuroimage 2016, 128, 413–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsey, J.D.; Hanson, S.J.; Hanson, C.; Halchenko, Y.O.; Poldrack, R.A.; Glymour, C. Six problems for causal inference from fMRI. Neuroimage 2010, 49, 1545–1558. [Google Scholar] [CrossRef] [PubMed]
- Lohmann, G.; Erfurth, K.; Müller, K.; Turner, R. Critical comments on dynamic causal modelling. Neuroimage 2012, 59, 2322–2329. [Google Scholar] [CrossRef] [PubMed]
- Guye, M.; Parker, G.J.; Symms, M.; Boulby, P.; Wheeler-Kingshott, C.A.; Salek-Haddadi, A.; Barker, J.G.; Duncan, J.S. Combined functional MRI and tractography to demonstrate the connectivity of the human primary motor cortex in vivo. Neuroimage 2003, 19, 1349–1360. [Google Scholar] [CrossRef]
- Kinnischtzke, A.K. Cell Type Specific Connections from Primary Motor to Primary Somatosensory Cortex. 2013. Available online: http://d-scholarship.pitt.edu/19434/ (accessed on 15 January 2019).
- Kasess, C.H.; Windischberger, C.; Cunnington, R.; Lanzenberger, R.; Pezawas, L.; Moser, E. The suppressive influence of SMA on M1 in motor imagery revealed by fMRI and dynamic causal modeling. Neuroimage 2008, 40, 828–837. [Google Scholar] [CrossRef]
- Vergani, F.; Lacerda, L.; Martino, J.; Attems, J.; Morris, C.; Mitchell, P.; de Schotten, D.T.; Dell’Acqua, F. White matter connections of the supplementary motor area in humans. J. Neurol. Neurosurg. Psychiatry. 2014, 85, 1377–1385. [Google Scholar] [CrossRef] [PubMed]
- Madigan, D.; Raftery, A.E. Model Selection and Accounting for Model Uncertainty in Graphical Models Using Occam’s Window. J. Am. Stat. Assoc. 1994, 89, 1535–1546. [Google Scholar] [CrossRef]
- Kass, R.E.; Raftery, A.E. Bayes Factors. J. Am. Stat. Assoc. 1995, 90, 773–795. [Google Scholar] [CrossRef]
- Friston, K. The free-energy principle: A unified brain theory? Nat. Rev. Neurosci. 2010, 11, 127–138. [Google Scholar] [CrossRef]
- Adams, R.A.; Shipp, S.; Friston, K.J. Predictions not commands: Active inference in the motor system. Brain Struct. Funct. 2013, 218, 611–643. [Google Scholar] [CrossRef] [Green Version]
- Wardak, C. The Role of the Supplementary Motor Area in Inhibitory Control in Monkeys and Humans. J. Neurosci. 2011, 31, 5181–5183. [Google Scholar] [CrossRef] [Green Version]
- Sanger, T.D. Failure of Motor Learning for Large Initial Errors. Neural Comput. 2004, 16, 1873–1886. [Google Scholar] [CrossRef]
- Ward, N.S.; Brown, M.M.; Thompson, A.J.; Frackowiak, R.S.J. Neural correlates of motor recovery after stroke: A longitudinal fMRI study. Brain 2003, 126, 2476–2496. [Google Scholar] [CrossRef]
- Ward, N.S.; Brown, M.M.; Thompson, A.J.; Frackowiak, R.S.J. Neural correlates of outcome after stroke: A cross-sectional fMRI study. Brain 2003, 126, 1430–1448. [Google Scholar]
- Seki, K.; Fetz, E.E. Gating of Sensory Input at Spinal and Cortical Levels during Preparation and Execution of Voluntary Movement. J. Neurosci. 2012, 32, 890–902. [Google Scholar] [CrossRef] [Green Version]
- Juravle, G.; Binsted, G.; Spence, C. Tactile suppression in goal-directed movement. Psychon. Bull. Rev. 2017, 24, 1060–1076. [Google Scholar] [CrossRef] [Green Version]
- Paillard, T.; Noé, F.; Passelergue, P.; Dupui, P. Electrical stimulation superimposed onto voluntary muscular contraction. Sports Med. 2005, 35, 951–966. [Google Scholar] [CrossRef]
- Jochumsen, M.; Niazi, I.K.; Signal, N.; Nedergaard, R.W.; Holt, K.; Haavik, H.; Taylor, D. Pairing Voluntary Movement and Muscle-Located Electrical Stimulation Increases Cortical Excitability. Front Hum. Neurosci. 2016, 10, 482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potgieser, A.R.E.; de Jong, B.M.; Wagemakers, M.; Hoving, E.W.; Groen, R.J.M. Insights from the supplementary motor area syndrome in balancing movement initiation and inhibition. Front. Hum. Neurosci. 2014, 8, 960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, L.D.; Black, S.E.; Gao, F.; Szilagyi, G.; Danells, C.J.; McIlroy, W.E. Correlating lesion size and location to deficits after ischemic stroke: The influence of accounting for altered peri-necrotic tissue and incidental silent infarcts. Behav. Brain Funct. 2010, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Seghier, M.L.; Fagan, E.; Price, C.J. Functional subdivisions in the left angular gyrus where the semantic system meets and diverges from the default network. J. Neurosci. 2010, 30, 16809–16817. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, E.; Mapelli, L.; Casellato, C.; Garrido, J.A.; Luque, N.; Monaco, J.; Prestori, F.; Pedrocchi, A.; Ros, E. Distributed Circuit Plasticity: New Clues for the Cerebellar Mechanisms of Learning. Cerebellum 2016, 15, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Gandolla, M.; Guanziroli, E.; D’Angelo, A.; Cannaviello, G.; Molteni, F.; Pedrocchi, A. Automatic setting procedure for exoskeleton-assisted overground gait: Proof of concept on stroke population. Front. Neurorobotics 2018, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject | Age [Years Range] | Site of Lesion | Type of Stroke | Time [Months] |
|---|---|---|---|---|
| 01 | 20–25 | R MCA | H | 23 |
| 02 | 35–40 | R GP | I | 23 |
| 03 | 60–65 | L MCA | H + I | 13 |
| 04 | 18–20 | L MCA | H | 44 |
| 05 | 45–50 | L GP | H | 44 |
| 06 | 20–25 | R MCA | I | 30 |
| 07 | 45–50 | R GP | I | 13 |
| 08 | 60–65 | R MCA | H | 58 |
| Subj. | Gait Velocity [m/s] | Endurance Velocity [m/s] | Paretic Step Length [mm] | TAAI | MRC | Capacity SCORE | Carry-Over Effect |
|---|---|---|---|---|---|---|---|
| 01 | Pre: 0.64 Post: 0.71 | Pre: 0.82 Post: 0.92 | Pre: 486 Post: 538 | Pre: 0.60 Post: 0.38 | Pre: 3 Post: 4 | Pre: 20.30 Post: 21.32 | Yes |
| 02 | Pre: 0.25 Post: 0.27 | Pre: 0.28 Post: 0.24 | Pre: 194 Post: 196 | Pre: 0.48 Post: 0.40 | Pre: 2 Post: 3 | Pre: 8.15 Post: 8.17 | No |
| 03 | Pre: 0.32 Post: 0.60 | Pre: 0.50 Post: 0.54 | Pre: 345 Post: 445 | Pre: 0.56 Post: 0.36 | Pre: 3 Post: 3 | Pre: 14.33 Post: 18.15 | Yes |
| 04 | Pre: 0.82 Post: 0.86 | Pre: 1.01 Post: 1.10 | Pre: 561 Post: 567 | Pre: 0.44 Post: 0.53 | Pre: 3 Post: 4 | Pre: 23.42 Post: 23.82 | No |
| 05 | Pre: 0.52 Post: 0.55 | Pre: 0.82 Post: 0.91 | Pre: 513 Post: 554 | Pre: 0.46 Post: 0.44 | Pre: 3 Post: 4 | Pre: 21.26 Post: 23 | Yes |
| 06 | Pre: 0.70 Post: 0.65 | Pre: 0.98 Post: 0.96 | Pre: 591 Post: 544 | Pre: 0.62 Post: 0.71 | Pre: 3 Post: 3 | Pre: 24.50 Post: 22.7 | No |
| 07 | Pre: 0.49 Post: 0.48 | Pre: 0.58 Post: 0.63 | Pre: 420 Post: 410 | Pre: 0.43 Post: 0.22 | Pre: 3 Post: 3 | Pre: 17.31 Post: 16.99 | No |
| 08 | Pre: 0.40 Post: 0.55 | Pre: 1.36 Post: 1.33 | Pre: 430 Post: 460 | Pre: 0.69 Post: 0.75 | Pre: 2 Post: 3 | Pre: 18.50 Post: 19.76 | Yes |
| PATIENTS | CONTROLS | |||||
|---|---|---|---|---|---|---|
| x [mm] | y [mm] | z [mm] | x [mm] | y [mm] | z [mm] | |
| M1 | ||||||
| S1 | ||||||
| SMA | ||||||
| AG | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gandolla, M.; Niero, L.; Molteni, F.; Guanziroli, E.; Ward, N.S.; Pedrocchi, A. Brain Plasticity Mechanisms Underlying Motor Control Reorganization: Pilot Longitudinal Study on Post-Stroke Subjects. Brain Sci. 2021, 11, 329. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11030329
Gandolla M, Niero L, Molteni F, Guanziroli E, Ward NS, Pedrocchi A. Brain Plasticity Mechanisms Underlying Motor Control Reorganization: Pilot Longitudinal Study on Post-Stroke Subjects. Brain Sciences. 2021; 11(3):329. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11030329
Chicago/Turabian StyleGandolla, Marta, Lorenzo Niero, Franco Molteni, Elenora Guanziroli, Nick S. Ward, and Alessandra Pedrocchi. 2021. "Brain Plasticity Mechanisms Underlying Motor Control Reorganization: Pilot Longitudinal Study on Post-Stroke Subjects" Brain Sciences 11, no. 3: 329. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11030329