
Recalled Parental Bonding Interacts with Oxytocin Receptor Gene Polymorphism in Modulating Anxiety and Avoidance in Adult Relationships

 ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
1.1. Familial Bonds in Italy and Singapore
1.2. OXTR Polymorphism rs53576
1.3. Current Study: Hypotheses
- HP1
- Gene*environment interaction on the Western sample: We expected to find a statistically significant effect of the interaction between OXTR polymorphism and parental bonding features (assessed using the PBI scales) over the main features of adult social relationships (measured with the ECR-R), with Italian A-carriers showing different rates of distress in terms of anxiety and avoidance than G/G homozygotes when they reported low levels of parental care and high levels of parental overprotection;
- HP2
- Gene*environment interaction on the Eastern sample: We expected to observe a statistically significant effect of the interaction between OXTR polymorphism and parental bonding features (assessed using the PBI scales) over the main features of adult social relationships (measured with the ECR-R), with Singaporean G-carriers exhibiting different rates of distress in terms of anxiety and avoidance than A/A homozygotes when they recalled low levels of parental care and high levels of parental overprotection.
- HP3
- Culture*gene*environment interactions on the total sample: We predicted a differential susceptibility of OXTR rs53576 to perceived parental bonding (assessed with the PBI subscales) between the Western (i.e., Italian) and the Eastern (i.e., Singaporean) groups in explaining the levels of anxiety and avoidance (measured using the ECR index.
2. Materials and Methods
2.1. Questionnaires
2.2. Genotyping
2.3. Statistical Analyses
2.3.1. Preliminary Analyses
2.3.2. Main Analyses
3. Results
3.1. Preliminary Results
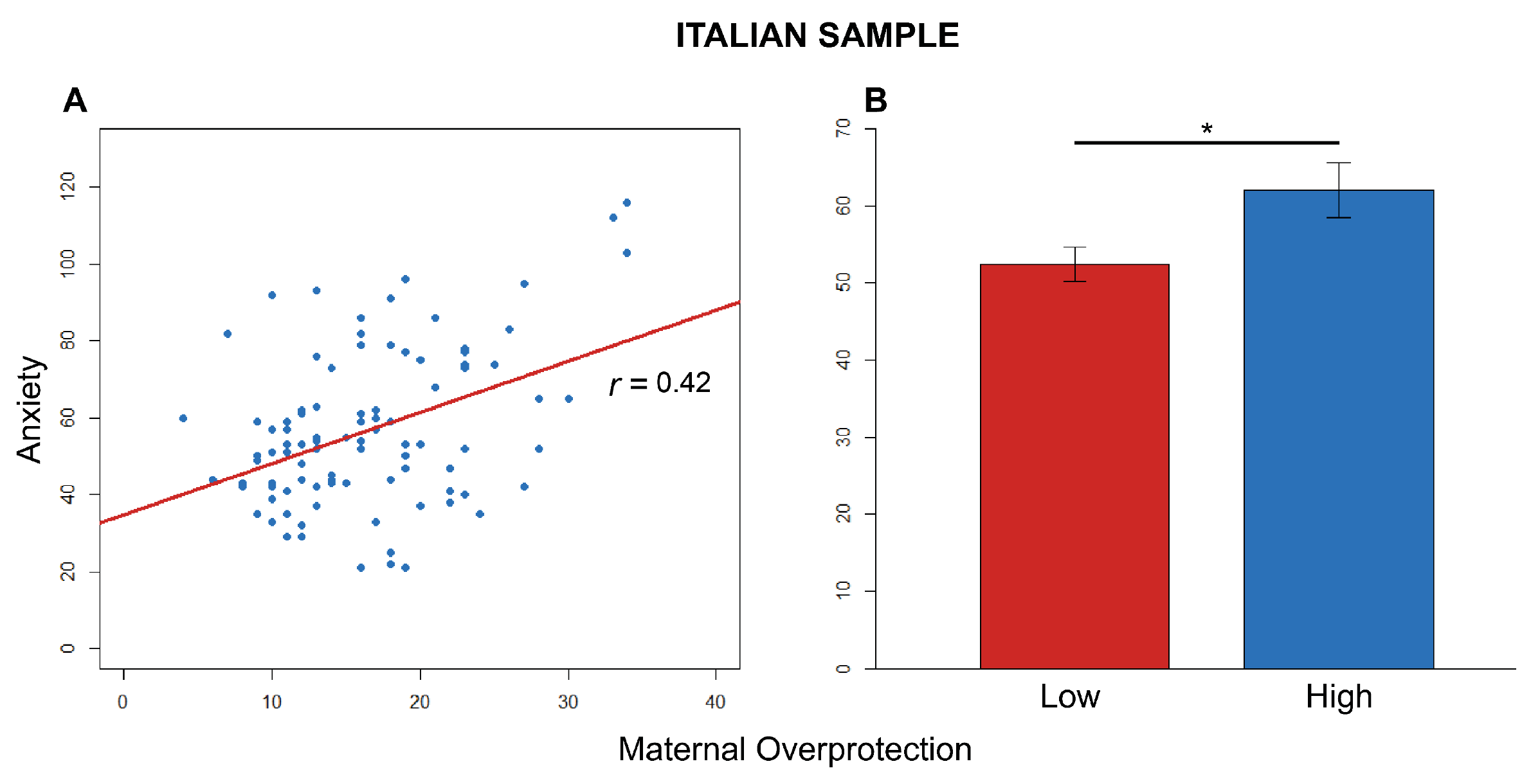
3.2. Italian Sample
3.2.1. Anxiety
3.2.2. Avoidance
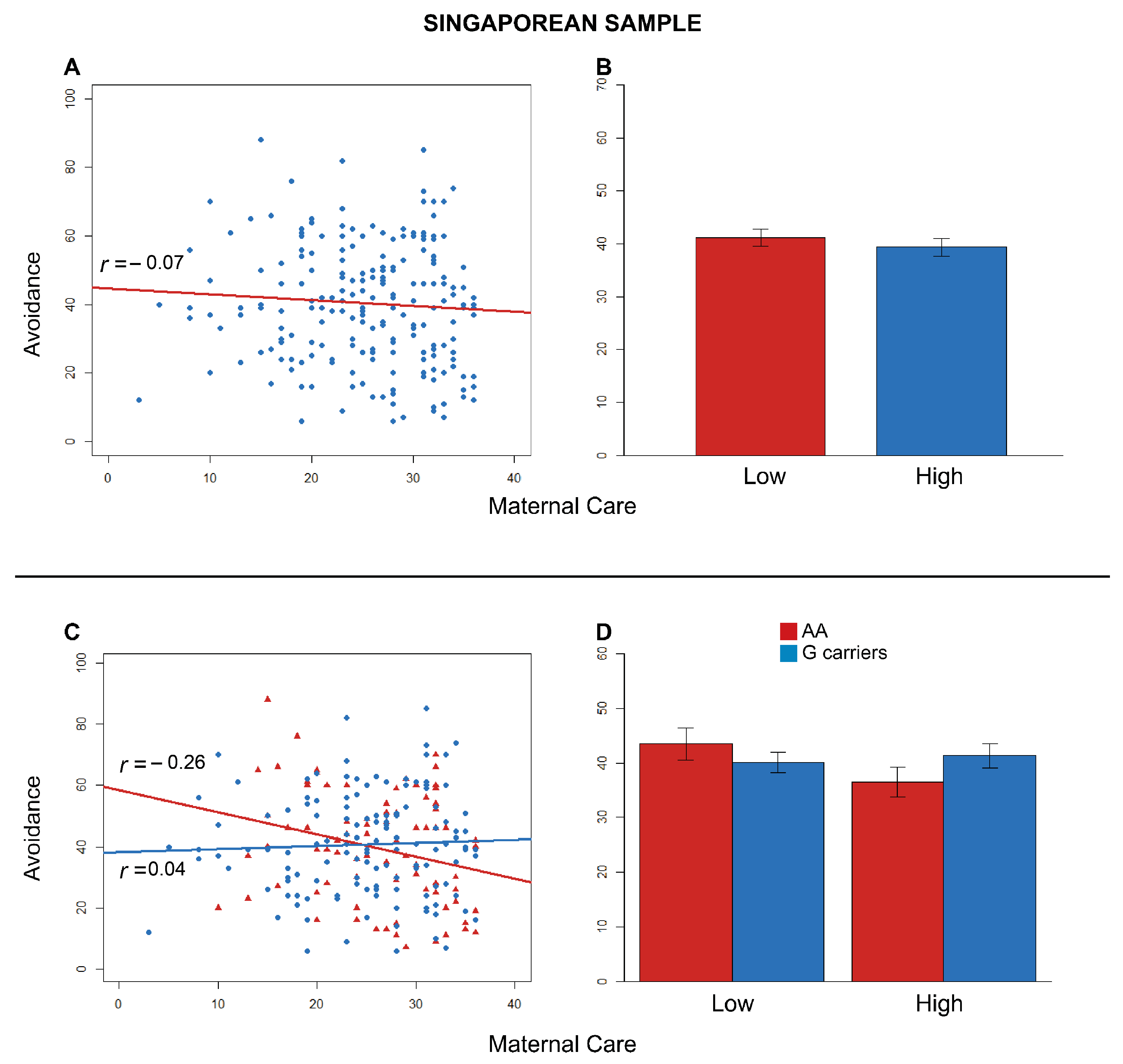
3.3. Singaporean Sample
3.3.1. Anxiety
3.3.2. Avoidance
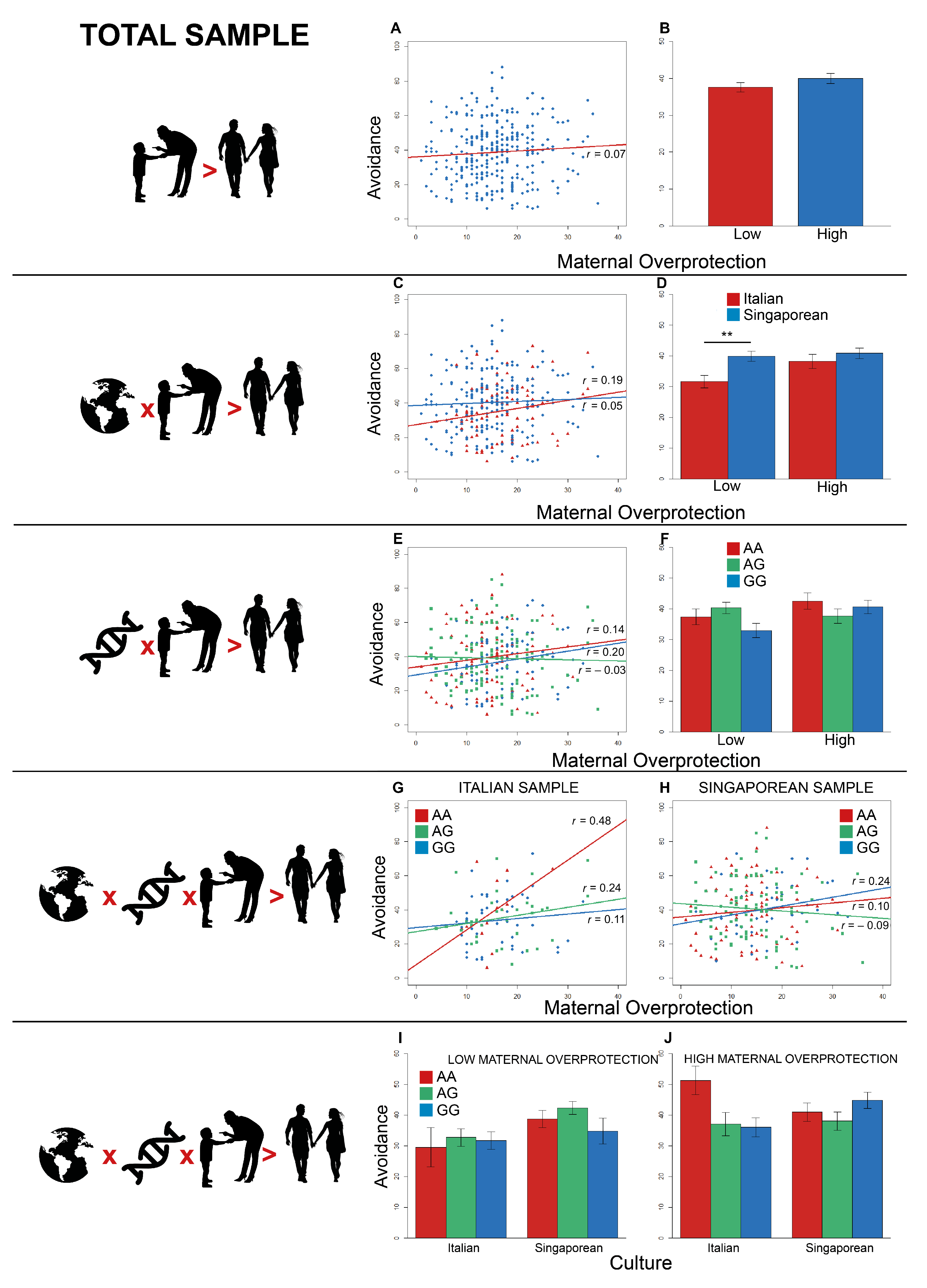
3.4. Total Sample: Italian and Singaporean Participants
3.4.1. Anxiety
3.4.2. Avoidance
4. Discussion
4.1. Gene-by-Environment Interaction within the Western-Oriented Sample and the Eastern-Oriented Sample Evaluated Independently
4.2. Gene-by-Culture Interactions in the Total Sample
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ECR-R | Experience in Close Relationships-Revised |
| PBI | Parental Bonding Instrument |
| OXTR | Oxytocin receptor gene |
| SS | Sum of squares |
| RSS | Residual sum of squares |
| HMR | Hierarchical multiple regression |
References
- Bowlby, J. Attachment and Loss: Volume II: Separation, Anxiety and Anger; The Hogarth Press and the Institute of Psycho-Analysis: London, UK, 1973; pp. 1–429. [Google Scholar]
- Parker, G.; Tupling, H.; Brown, L.B. A parental bonding instrument. Br. J. Med. Psychol. 1979, 52, 1–10. [Google Scholar] [CrossRef]
- Zafiropoulou, M.; Avagianou, P.A.; Vassiliadou, S. Parental bonding and early maladaptive schemas. J. Psychol. Abnorm. Child. 2014, 3. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.; Lee, J.; Kim, B.; Lee, S.M. Students’ perceptions of parental bonding styles and their academic burnout. Asia Pac. Educ. Rev. 2012, 13, 509–517. [Google Scholar] [CrossRef]
- Francis, D.D.; Meaney, M.J. Maternal care and the development of stress responses. Curr. Opin. Neurobiol. 1999, 9, 128–134. [Google Scholar] [CrossRef]
- Hane, A.A.; Cheah, C.; Rubin, K.H.; Fox, N.A. The role of maternal behavior in the relation between shyness and social reticence in early childhood and social withdrawal in middle childhood. Soc. Dev. 2008, 17, 795–811. [Google Scholar] [CrossRef]
- Picardi, A.; Caroppo, E.; Fabi, E.; Proietti, S.; Di Gennaro, G.; Meldolesi, G.N.; Martinotti, G. Attachment and parenting in adult patients with anxiety disorders. Clin. Pract. Epidemiol. Ment. Health CP EMH 2013, 9, 157. [Google Scholar] [CrossRef]
- Rogosch, F.A.; Cicchetti, D.; Aber, J.L. The role of child maltreatment in early deviations in cognitive and affective processing abilities and later peer relationship problems. Dev. Psychopathol. 1995, 7, 591–609. [Google Scholar] [CrossRef]
- Shaver, P.R.; Mikulincer, M. Attachment-related psychodynamics. Attach. Hum. Dev. 2002, 4, 133–161. [Google Scholar] [CrossRef] [PubMed]
- Reti, I.M.; Samuels, J.; Eaton, W.W.; Bienvenu Iii, O.; Costa, P., Jr.; Nestadt, G. Adult antisocial personality traits are associated with experiences of low parental care and maternal overprotection. Acta Psychiatr. Scand. 2002, 106, 126–133. [Google Scholar] [CrossRef]
- Spinrad, T.L.; Gal, D.E. Fostering prosocial behavior and empathy in young children. Curr. Opin. Psychol. 2018, 20, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.S.; Silk, J.S.; Steinberg, L.; Myers, S.S.; Robinson, L.R. The role of the family context in the development of emotion regulation. Soc. Dev. 2007, 16, 361–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porges, S.W. Social engagement and attachment: A phylogenetic perspective. Ann. N. Y. Acad. Sci. 2003, 1008, 31–47. [Google Scholar] [CrossRef] [PubMed]
- Rork, K.E.; Morris, T.L. Influence of parenting factors on childhood social anxiety: Direct observation of parental warmth and control. Child Fam. Behav. Ther. 2009, 31, 220–235. [Google Scholar] [CrossRef]
- Ainsworth, M.D.S. Infancy in Uganda: Infant Care and the Growth of Love; Johns Hopkins Press: Baltimore, MD, USA, 1967. [Google Scholar]
- Behrens, K.Y.; Hesse, E.; Main, M. Mothers’ attachment status as determined by the Adult Attachment Interview predicts their 6-year-olds’ reunion responses: A study conducted in Japan. Dev. Psychol. 2007, 43, 1553. [Google Scholar] [CrossRef]
- Mesman, J.; van IJzendoorn, M.H.; Bakermans-Kranenburg, M.J. Unequal in opportunity, equal in process: Parental sensitivity promotes positive child development in ethnic minority families. Child Dev. Perspect. 2012, 6, 239–250. [Google Scholar] [CrossRef]
- Sun, Q.W.; Ng, K.M.; Guo, L. The link between parental bonding and adult attachment in Chinese graduate students: Gender differences. Fam. J. 2010, 18, 386–394. [Google Scholar] [CrossRef]
- Laudani, C.; Guzzo, G.; Cascio, V.L.; Pace, U.; Cacioppo, M. Does a Mediterranean model of family functioning in the perception of Italian and Spanish adolescents exist? A cross-national study. Mediterr. J. Soc. Sci. 2014, 5, 377. [Google Scholar] [CrossRef]
- Keller, H. Universality claim of attachment theory: Children’s socioemotional development across cultures. Proc. Natl. Acad. Sci. USA 2018, 115, 11414–11419. [Google Scholar] [CrossRef] [Green Version]
- Cataldo, I.; Azhari, A.; Lepri, B.; Esposito, G. Oxytocin receptors (OXTR) and early parental care: An interaction that modulates psychiatric disorders. Res. Dev. Disabil. 2018, 82, 27–38. [Google Scholar] [CrossRef]
- Walum, H.; Lichtenstein, P.; Neiderhiser, J.M.; Reiss, D.; Ganiban, J.M.; Spotts, E.L.; Pedersen, N.L.; Anckarsäter, H.; Larsson, H.; Westberg, L. Variation in the oxytocin receptor gene is associated with pair-bonding and social behavior. Biol. Psychiatry 2012, 71, 419–426. [Google Scholar] [CrossRef] [Green Version]
- Tost, H.; Kolachana, B.; Hakimi, S.; Lemaitre, H.; Verchinski, B.A.; Mattay, V.S.; Weinberger, D.R.; Meyer-Lindenberg, A. A common allele in the oxytocin receptor gene (OXTR) impacts prosocial temperament and human hypothalamic-limbic structure and function. Proc. Natl. Acad. Sci. USA 2010, 107, 13936–13941. [Google Scholar] [CrossRef] [Green Version]
- Feldman, R. Oxytocin and social affiliation in humans. Horm. Behav. 2012, 61, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Zink, C.F.; Meyer-Lindenberg, A. Human neuroimaging of oxytocin and vasopressin in social cognition. Horm. Behav. 2012, 61, 400–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domes, G.; Heinrichs, M.; Gläscher, J.; Büchel, C.; Braus, D.F.; Herpertz, S.C. Oxytocin attenuates amygdala responses to emotional faces regardless of valence. Biol. Psychiatry 2007, 62, 1187–1190. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, S.M.; Saslow, L.R.; Garcia, N.; John, O.P.; Keltner, D. Oxytocin receptor genetic variation relates to empathy and stress reactivity in humans. Proc. Natl. Acad. Sci. USA 2009, 106, 21437–21441. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.E.; Porges, E.C.; Norman, G.J.; Connelly, J.J.; Decety, J. Oxytocin receptor gene variation predicts empathic concern and autonomic arousal while perceiving harm to others. Soc. Neurosci. 2014, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truzzi, A.; Poquérusse, J.; Setoh, P.; Shinohara, K.; Bornstein, M.H.; Esposito, G. Oxytocin receptor gene polymorphisms (rs53576) and early paternal care sensitize males to distressing female vocalizations. Dev. Psychobiol. 2018, 60, 333–339. [Google Scholar] [CrossRef]
- Senese, V.P.; Shinohara, K.; Esposito, G.; Doi, H.; Venuti, P.; Bornstein, M.H. Implicit association to infant faces: Genetics, early care experiences, and cultural factors influence caregiving propensities. Behav. Brain Res. 2017, 325, 163–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norman, G.J.; Hawkley, L.; Luhmann, M.; Ball, A.B.; Cole, S.W.; Berntson, G.G.; Cacioppo, J.T. Variation in the oxytocin receptor gene influences neurocardiac reactivity to social stress and HPA function: A population based study. Horm. Behav. 2012, 61, 134–139. [Google Scholar] [CrossRef]
- Thompson, R.J.; Parker, K.J.; Hallmayer, J.F.; Waugh, C.E.; Gotlib, I.H. Oxytocin receptor gene polymorphism (rs2254298) interacts with familial risk for psychopathology to predict symptoms of depression and anxiety in adolescent girls. Psychoneuroendocrinology 2011, 36, 144–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, N.; Li, Z.; Su, Y. The association between oxytocin receptor gene polymorphism (OXTR) and trait empathy. J. Affect. Disord. 2012, 138, 468–472. [Google Scholar] [CrossRef]
- Lucht, M.J.; Barnow, S.; Sonnenfeld, C.; Rosenberger, A.; Grabe, H.J.; Schroeder, W.; Völzke, H.; Freyberger, H.J.; Herrmann, F.H.; Kroemer, H.; et al. Associations between the oxytocin receptor gene (OXTR) and affect, loneliness and intelligence in normal subjects. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2009, 33, 860–866. [Google Scholar] [CrossRef]
- Apicella, C.L.; Cesarini, D.; Johannesson, M.; Dawes, C.T.; Lichtenstein, P.; Wallace, B.; Beauchamp, J.; Westberg, L. No association between oxytocin receptor (OXTR) gene polymorphisms and experimentally elicited social preferences. PLoS ONE 2010, 5, e11153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhao, Y.; Li, R.; Broster, L.S.; Zhou, C.; Yang, S. Association of oxytocin receptor gene (OXTR) rs53576 polymorphism with sociality: A meta-analysis. PLoS ONE 2015, 10, e0131820. [Google Scholar] [CrossRef] [PubMed]
- Bakermans-Kranenburg, M.J.; van IJzendoorn, M.H. Oxytocin receptor (OXTR) and serotonin transporter (5-HTT) genes associated with observed parenting. Soc. Cogn. Affect. Neurosci. 2008, 3, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Sherman, D.K.; Sasaki, J.Y.; Xu, J.; Chu, T.Q.; Ryu, C.; Suh, E.M.; Graham, K.; Taylor, S.E. Culture, distress, and oxytocin receptor polymorphism (OXTR) interact to influence emotional support seeking. Proc. Natl. Acad. Sci. USA 2010, 107, 15717–15721. [Google Scholar]
- Fraley, R.C.; Waller, N.G.; Brennan, K.A. An item response theory analysis of self-report measures of adult attachment. J. Personal. Soc. Psychol. 2000, 78, 350. [Google Scholar] [CrossRef]
- Scinto, A.; Marinangeli, M.G.; Kalyvoka, A.; Daneluzzo, E.; Rossi, A. The use of the Italian version of the Parental Bonding Instrument (PBI) in a clinical sample and in a student group: An exploratory and confirmatory factor analysis study. Epidemiol. Psychiatr. Sci. 1999, 8, 276–283. [Google Scholar] [CrossRef]
- Busonera, A.; Martini, P.S.; Zavattini, G.C.; Santona, A. Psychometric properties of an Italian version of the experiences in close relationships-revised (ECR-R) scale. Psychol. Rep. 2014, 114, 785–801. [Google Scholar] [CrossRef] [PubMed]
- Parker, G. The parental bonding instrument: Psychometric properties reviewed. Psychiatr. Dev. 1989, 7, 317–335. [Google Scholar]
- Bonassi, A.; Ghilardi, T.; Truzzi, A.; Cataldo, I.; Azhari, A.; Setoh, P.; Shinohara, K.; Esposito, G. Dataset on genetic and physiological adults’ responses to social distress. Data Brief 2017, 13, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Jia, M.; Ruan, Y.; Liu, J.; Guo, Y.; Shuang, M.; Gong, X.; Zhang, Y.; Yang, X.; Zhang, D. Positive association of the oxytocin receptor gene (OXTR) with autism in the Chinese Han population. Biol. Psychiatry 2005, 58, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Cataldo, I.; Neoh, M.J.Y.; Chew, W.F.; Foo, J.N.; Lepri, B.; Esposito, G. Oxytocin receptor gene and parental bonding modulate prefrontal responses to cries: A NIRS Study. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Butovskaya, M.; Rostovtseva, V.; Butovskaya, P.; Burkova, V.; Dronova, D.; Filatova, V.; Sukhodolskaya, E.; Vasiliev, V.; Mesa, T.; Rosa, A.; et al. Oxytocin receptor gene polymorphism (rs53576) and digit ratio associates with aggression: Comparison in seven ethnic groups. J. Physiol. Anthropol. 2020, 39, 1–15. [Google Scholar] [CrossRef]
- Zijlstra, W.P.; van der Ark, L.A.; Sijtsma, K. Outliers in questionnaire data: Can they be detected and should they be removed? J. Educ. Behav. Stat. 2011, 36, 186–212. [Google Scholar] [CrossRef]
- Bradley, R.H.; Corwyn, R.F.; Burchinal, M.; McAdoo, H.P.; García Coll, C. The home environments of children in the United States Part II: Relations with behavioral development through age thirteen. Child Dev. 2001, 72, 1868–1886. [Google Scholar] [CrossRef]
- Elgar, F.J.; Mills, R.S.; McGrath, P.J.; Waschbusch, D.A.; Brownridge, D.A. Maternal and paternal depressive symptoms and child maladjustment: The mediating role of parental behavior. J. Abnorm. Child Psychol. 2007, 35, 943–955. [Google Scholar] [CrossRef]
- Kogan, A.; Saslow, L.R.; Impett, E.A.; Oveis, C.; Keltner, D.; Saturn, S.R. Thin-slicing study of the oxytocin receptor (OXTR) gene and the evaluation and expression of the prosocial disposition. Proc. Natl. Acad. Sci. USA 2011, 108, 19189–19192. [Google Scholar] [CrossRef] [Green Version]
- Senese, V.P.; Azhari, A.; Shinohara, K.; Doi, H.; Venuti, P.; Bornstein, M.H.; Esposito, G. Implicit associations to infant cry: Genetics and early care experiences influence caregiving propensities. Horm. Behav. 2019, 108, 1–9. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.G. Statistical power analyses using G* Power 3.1: Tests for correlation and regression analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef] [Green Version]
- Diedenhofen, B.; Musch, J. cocor: A comprehensive solution for the statistical comparison of correlations. PLoS ONE 2015, 10, e0121945. [Google Scholar] [CrossRef] [Green Version]
- Ravitz, P.; Maunder, R.; Hunter, J.; Sthankiya, B.; Lancee, W. Adult attachment measures: A 25-year review. J. Psychosom. Res. 2010, 69, 419–432. [Google Scholar] [CrossRef]
- Ebstein, R.; Israel, S.; Lerer, E.; Uzefovsky, F.; Shalev, I.; Gritsenko, I.; Riebold, M.; Salomon, S.; Yirmiya, N. Arginine vasopressin and oxytocin modulate human social behavior. Ann. N. Y. Acad. Sci. 2009, 1167, 87–102. [Google Scholar] [CrossRef]
- Ebstein, R.P.; Knafo, A.; Mankuta, D.; Chew, S.H.; San Lai, P. The contributions of oxytocin and vasopressin pathway genes to human behavior. Horm. Behav. 2012, 61, 359–379. [Google Scholar] [CrossRef]
- Feeney, J.A. Early parenting and parental attachment: Links with offspring’s attachment and perceptions of social support. J. Fam. Stud. 2002, 8, 5–23. [Google Scholar] [CrossRef]
- Hao, J.; Chan, R.C.; Wilkinson, R.B. A cross-cultural examination of the Experiences in Close Relationships—Revised—General Short Form (ECR-R-GSF) in an Australian and a Chinese sample. J. Relatsh. Res. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Mastrotheodoros, S.; Chen, B.B.; Motti-Stefanidi, F. Experiences in close relationships-revised (ECR-R): Measurement (non-) invariance across Chinese and Greek samples. Eur. J. Dev. Psychol. 2015, 12, 344–358. [Google Scholar] [CrossRef]
- Bowlby, J. Developmental psychiatry comes of age. Am. J. Psychiatry 1988, 145, 1–10. [Google Scholar]
- Parker, G.; Lipscombe, P. Influences on maternal overprotection. Br. J. Psychiatry 1981, 138, 303–311. [Google Scholar] [CrossRef]
- Raudino, A.; Murray, L.; Turner, C.; Tsampala, E.; Lis, A.; De Pascalis, L.; Cooper, P.J. Child anxiety and parenting in England and Italy: The moderating role of maternal warmth. J. Child Psychol. Psychiatry 2013, 54, 1318–1326. [Google Scholar] [CrossRef]
- Kiel, E.J.; Kalomiris, A.E. Emotional Development and Anxiety. In Handbook of Emotional Development; Springer: Berlin/Heidelberg, Germany, 2019; pp. 665–693. [Google Scholar]
- Smout, A.; Lazarus, R.S.; Hudson, J.L. The Relationship Between Parenting and Anxiety in Emerging Adulthood. Cogn. Ther. Res. 2020, 44, 182–195. [Google Scholar] [CrossRef]
- Land, L.N.; Rochlen, A.B.; Vaughn, B.K. Correlates of adult attachment avoidance: Men’s avoidance of intimacy in romantic relationships. Psychol. Men Masculinity 2011, 12, 64. [Google Scholar] [CrossRef]
- Gallagher, B.; Cartwright-Hatton, S. The relationship between parenting factors and trait anxiety: Mediating role of cognitive errors and metacognition. J. Anxiety Disord. 2008, 22, 722–733. [Google Scholar] [CrossRef] [PubMed]
- Pellerone, M.; Ramaci, T.; Herrera López, M.; Craparo, G. The role of identity development and decision making process on adult attachment: A cross-national study in sicilian and andalusian adolescents. Clin. Neuropsychiatry 2017, 14, 141–150. [Google Scholar]
- Pace, U.; Cacioppo, M.; Cascio, V.L.; Guzzo, G.; Passanisi, A. Are there similar or divergent transitions to adulthood in a Mediterranean context? A cross-national comparison of Italy and Spain. Eur. J. Psychol. 2016, 12, 153. [Google Scholar] [CrossRef] [Green Version]
- Belsky, J.; Pluess, M. Beyond diathesis stress: Differential susceptibility to environmental influences. Psychol. Bull. 2009, 135, 885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, P.; Wang, Q.; Liu, J.; Xi, S.; Yang, X.; Fang, P.; Wang, B.; He, L.; Guo, W.; Zhang, M. The OXTR polymorphisms are not associated with attachment dimensions: A three-approach study. Psychoneuroendocrinology 2020, 120, 104780. [Google Scholar] [CrossRef] [PubMed]
- Gillath, O.; Shaver, P.R.; Baek, J.M.; Chun, D.S. Genetic correlates of adult attachment style. Personal. Soc. Psychol. Bull. 2008, 34, 1396–1405. [Google Scholar] [CrossRef]
- Sümer, N.; Yetkili, O. Cultural aspects of attachment anxiety, avoidance, and life satisfaction: Comparing the US and Turkey. In Close Relationships and Happiness across Cultures; Springer: Berlin/Heidelberg, Germany, 2018; pp. 165–184. [Google Scholar]
- Friedman, M.; Rholes, W.S.; Simpson, J.; Bond, M.; DIAZ-LOVING, R.; Chan, C. Attachment avoidance and the cultural fit hypothesis: A cross-cultural investigation. Pers. Relatsh. 2010, 17, 107–126. [Google Scholar] [CrossRef]
- Luo, S.; Ma, Y.; Liu, Y.; Li, B.; Wang, C.; Shi, Z.; Li, X.; Zhang, W.; Rao, Y.; Han, S. Interaction between oxytocin receptor polymorphism and interdependent culture values on human empathy. Soc. Cogn. Affect. Neurosci. 2015, 10, 1273–1281. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, S.E.; Low, W.Y.; Hashim, A.H. Perceived parenting styles and cultural influences in adolescent’s anxiety: A cross-cultural comparison. J. Child Fam. Stud. 2016, 25, 2102–2110. [Google Scholar] [CrossRef]
- Kim, H.S.; Sherman, D.K.; Mojaverian, T.; Sasaki, J.Y.; Park, J.; Suh, E.M.; Taylor, S.E. Gene–culture interaction: Oxytocin receptor polymorphism (OXTR) and emotion regulation. Soc. Psychol. Personal. Sci. 2011, 2, 665–672. [Google Scholar] [CrossRef]
- Kashdan, T.B.; Barrios, V.; Forsyth, J.P.; Steger, M.F. Experiential avoidance as a generalized psychological vulnerability: Comparisons with coping and emotion regulation strategies. Behav. Res. Ther. 2006, 44, 1301–1320. [Google Scholar] [CrossRef]
- Fulton, J.J.; Kiel, E.J.; Tull, M.T.; Gratz, K.L. Associations between perceived parental overprotection, experiential avoidance, and anxiety. J. Exp. Psychopathol. 2014, 5, 200–211. [Google Scholar] [CrossRef]
- Bonassi, A.; Cataldo, I.; Gabrieli, G.; Foo, J.N.; Lepri, B.; Esposito, G. Oxytocin receptor gene polymorphisms and early parental bonding interact in shaping instagram social behavior. Int. J. Environ. Res. Public Health 2020, 17, 7232. [Google Scholar] [CrossRef]
- Carollo, A.; Bonassi, A.; Cataldo, I.; Gabrieli, G.; Tandiono, M.; Foo, J.N.; Lepri, B.; Esposito, G. The relation between Oxytocin Receptor Gene polymorphisms, adult attachment and Instagram sociability: An exploratory analysis. PsyArXiv 2020. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Genotype | A/A | A/G | G/G | p-Value | |
|---|---|---|---|---|---|
| rs53576ITA | 14 (14.43%) | 35 (36.08%) | 48 (49.49%) | 3.05 | 0.08 |
| rs53576SNG | 80 (37.04%) | 94 (43.52%) | 42 (19.44%) | 2.24 | 0.13 |
| rs53576Total | 94 (30.03%) | 129 (41.21%) | 90 (28–75%) | 9.64 | 0.002 ** |
| Group | Subscale | Cronbach | Skewness | Kurtosis | Mean (SD) | Median |
|---|---|---|---|---|---|---|
| Italy | Age | - | - | - | 23.01 (3.82) | 22 |
| PBI M_Care | 0.93 | −1.15 * | 0.71 | 26.33 (8.50) | 28 | |
| Squared PBI M_Care | - | −0.43 | −0.88 | 764.70 (373.24) | 784 | |
| PBI M_OverP | 0.78 | 0.72 | 0.18 | 16.40 (6.30) | 16 | |
| PBI F_Care | 0.94 | −0.49 | −0.69 | 23.23 (9.30) | 25 | |
| PBI F_OverP | 0.83 | 0.75 | 0.17 | 14.12 (7.01) | 13 | |
| ECR-R Anxiety | 0.91 | 0.70 | 0.14 | 56.59 (20.10) | 53 | |
| ECR-R Avoidance | 0.88 | 0.38 | −0.39 | 35.12 (15.37) | 33 | |
| Singapore | Age | - | - | - | 21.51 (1.83) | 21 |
| PBI M_Care | 0.91 | −0.66 | −0.03 | 25.39 (6.99) | 26 | |
| PBI M_OverP | 0.86 | 0.40 | −0.01 | 15.09 (7.06) | 15 | |
| PBI F_Care | 0.92 | −0.28 | −0.41 | 21.44 (8.14) | 22 | |
| PBI F_OverP | 0.86 | 0.69 | 0.51 | 11.57 (6.63) | 11 | |
| ECR-R Anxiety | 0.93 | −0.04 | −0.39 | 64.79 (20.61) | 66.50 | |
| ECR-R Avoidance | 0.94 | 0.12 | −0.51 | 40.31 (17.13) | 40 | |
| Total | Age | - | - | - | 21.97 (2.70) | 22 |
| PBI M_Care | 0.92 | -0.86 | 0.36 | 25.68 (7.49) | 27 | |
| PBI M_OverP | 0.83 | 0.45 | 0.09 | 15.50 (6.85) | 15 | |
| PBI F_Care | 0.93 | −0.32 | −0.53 | 21.99 (8.54) | 23 | |
| PBI F_OverP | 0.83 | 0.71 | 0.46 | 12.36 (6.84) | 12 | |
| ECR-R Anxiety | 0.92 | 0.18 | −0.44 | 62.28 (20.76) | 62 | |
| ECR-R Avoidance | 0.92 | 0.22 | −0.48 | 38.70 (16.75) | 39 |
| Sample | ECR-R Subscale | Variable | t (df) | p-Value | 95% CI | r |
|---|---|---|---|---|---|---|
| Italy | Anxiety | sex | −1.21 (95) | 0.23 | [−13.31; 3.25] | |
| Age | 0.19 (95) | 0.85 | [−7.38; 8.93] | −0.03 | ||
| Avoidance | sex | 2.19 (95) | 0.03 | [0.67; 13.12] | ||
| Age | −0.49 (95) | 0.62 | [−7.78; 4.69] | 0.19 | ||
| Singapore | Anxiety | sex | 1.69 (214) | 0.09 | [−0.84; 10.80] | |
| Age | −0.03 (214) | 0.97 | [−5.64; 5.45] | 0.03 | ||
| Avoidance | sex | −0.21 (214) | 0.83 | [−5.39; 4.34] | ||
| Age | 1.00 (214) | 0.32 | [−2.27; 6.92] | −0.07 | ||
| Total | Anxiety | sex | 0.54 (311) | 0.59 | [−3.50; 6.16] | |
| Age | −0.14 (311) | 0.89 | [−5.27; 4.57] | −0.05 | ||
| Culture | 3.24 (311) | 0.13 * | [3.18; 13.02] | |||
| Avoidance | sex | 0.81 (311) | 0.42 | [−2.29; 5.50] | ||
| Age | 0.16 (311) | 0.87 | [−3.65; 4.30] | 0.01 | ||
| Culture | 2.56 (311) | 0.01 | [1.19; 9.18] |
| Subscale | M_Care | M_Overp | F_Care | F_Overp | Anxiety | Avoidance |
|---|---|---|---|---|---|---|
| M_Care | −0.29 *** | 0.43 *** | −0.10 | −0.06 | −0.07 | |
| M_Overp | −0.31 * | −0.17 | 0.39 *** | 0.1 | 0.05 | |
| F_Care | 0.49 *** | −0.23 | −0.18 | −0.16 | −0.09 | |
| F_Overp | −0.07 | 0.39 *** | −0.30 * | 0.17 | 0.10 | |
| Anxiety | −0.36 ** | 0.42 *** | −0.37 ** | 0.20 | 0.38 *** | |
| Avoidance | −0.25 | 0.19 | −0.20 | 0.08 | 0.35 ** |
| Step | Variable | SE | t | R | |||
|---|---|---|---|---|---|---|---|
| 1 | 0.07 | 0.01 | 0.01 | ||||
| OXTR rs53576 | 2.84 | 4.09 | 0.69 | ||||
| 2 | 0.37 | 0.14 | 0.13 *** | ||||
| OXTR rs53576 | 3.11 | 3.83 | 0.81 | ||||
| M_Care | −0.02 *** | 0.01 | −3.81 | ||||
| 3 | 0.49 | 0.24 | 0.10 *** | ||||
| OXTR rs53576 | 3.36 | 3.62 | 0.93 | ||||
| M_Care | −0.01 ** | 0.01 | −2.73 | ||||
| M_OverP | 1.08 *** | 0.30 | 3.54 | ||||
| 4 | 0.53 | 0.28 | 0.04 * | ||||
| OXTR rs53576 | 4.06 | 3.55 | 1.14 | ||||
| M_Care | −0.01 | 0.01 | −1.46 | ||||
| M_OverP | 1.01 ** | 0.30 | 3.39 | ||||
| F_Care | −0.50 * | 0.22 | −2.25 | ||||
| 5 | 0.53 | 0.28 | 0 | ||||
| OXTR rs53576 | 4.06 | 3.57 | 1.14 | ||||
| M_Care | −0.01 | 0.01 | −1.42 | ||||
| M_OverP | 1.01 ** | 0.33 | 3.11 | ||||
| F_Care | −0.50 * | 0.23 | −2.13 | ||||
| F_OverP | 0.00 | 0.29 | 0.00 | ||||
| 6 | 0.54 | 0.29 | 0.01 | ||||
| OXTR rs53576 | 18.08 | 17.73 | 1.02 | ||||
| M_Care | −0.01 | 0.01 | −0.72 | ||||
| M_OverP | 1.12 * | 0.47 | 2.40 | ||||
| F_Care | −0.48 | 0.38 | −1.29 | ||||
| F_OverP | 0.20 | 0.44 | 0.45 | ||||
| OXTR rs53576 x M_Care | 0.00 | 0.01 | −0.34 | ||||
| OXTR rs53576 x M_OverP | −0.26 | 0.67 | −0.39 | ||||
| OXTR rs53576 x F_Care | −0.01 | 0.49 | −0.01 | ||||
| OXTR rs53576 x F_OverP | −0.46 | 0.61 | 0.76 |
| Step | Variable | SE | t | R | |||
|---|---|---|---|---|---|---|---|
| 1 | 0.03 | 0.00 | 0.00 | ||||
| OXTR rs53576 | 1.02 | 2.42 | 0.43 | ||||
| 2 | 0.07 | 0.01 | 0.01 | ||||
| OXTR rs53576 | 0.86 | 2.42 | 0.35 | ||||
| M_Care | −0.17 | 0.17 | −0.99 | ||||
| 3 | 0.08 | 0.01 | 0.00 | ||||
| OXTR rs53576 | 0.78 | 2.44 | 0.32 | ||||
| M_Care | −0.15 | 0.18 | −0.84 | ||||
| M_OverP | 0.07 | 0.17 | 0.38 | ||||
| 4 | 0.10 | 0.01 | 0.00 | ||||
| OXTR rs53576 | 1.00 | 2.45 | 0.41 | ||||
| M_Care | −0.08 | 0.19 | −0.39 | ||||
| M_OverP | 0.06 | 0.18 | 0.33 | ||||
| F_Care | −0.15 | 0.16 | −0.92 | ||||
| 5 | 0.13 | 0.02 | 0.01 | ||||
| OXTR rs53576 | 0.95 | 2.45 | 0.39 | ||||
| M_Care | −0.09 | 0.19 | −0.49 | ||||
| M_OverP | −0.03 | 0.19 | −0.16 | ||||
| F_Care | −0.12 | 0.16 | −0.73 | ||||
| F_OverP | 0.24 | 0.19 | 1.25 | ||||
| 6 | 0.22 | 0.05 | 0.03 | ||||
| OXTR rs53576 | −17.16 | 13.28 | −1.29 | ||||
| M_Care | −0.83 * | 0.35 | −2.37 | ||||
| M_OverP | 0.14 | 0.35 | 0.40 | ||||
| F_Care | 0.22 | 0.29 | 0.76 | ||||
| F_OverP | 0.02 | 0.33 | 0.05 | ||||
| OXTR rs53576 x M_Care | 1.03 * | 0.42 | 2.45 | ||||
| OXTR rs53576 x M_OverP | −0.16 | 0.42 | −0.39 | ||||
| OXTR rs53576 x F_Care | −0.45 | 0.35 | −1.27 | ||||
| OXTR rs53576 x F_OverP | 0.30 | 0.41 | 0.73 |
| Step | Variable | SE | t | R | |||
|---|---|---|---|---|---|---|---|
| 1 | 0.14 | 0.02 | 0.02 ** | ||||
| Culture | 5.19 * | 2.03 | 2.55 | ||||
| 2 | 0.14 | 0.02 | 0.00 | ||||
| Culture | 5.05 * | 2.15 | 2.35 | ||||
| OXTR rs53576 A/G-A/A | 0.21 | 2.27 | 0.09 | ||||
| OXTR rs53576 G/G-A/A | −0.39 | 2.59 | −0.15 | ||||
| 3 | 0.18 | 0.03 | 0.01 * | ||||
| Culture | 4.74 * | 2.15 | 2.21 | ||||
| OXTR rs53576 A/G-A/A | −0.01 | 2.27 | −0.01 | ||||
| OXTR rs53576 G/G-A/A | −0.69 | 2.58 | −0.27 | ||||
| M_Care | −0.26 * | 0.13 | −2.05 | ||||
| 4 | 0.19 | 0.04 | 0.01 | ||||
| Culture | 4.88 * | 2.15 | 2.27 | ||||
| OXTR rs53576 A/G-A/A | −0.12 | 2.27 | −0.05 | ||||
| OXTR rs53576 G/G-A/A | −0.99 | 2.60 | −0.38 | ||||
| M_Care | −0.22 | 0.13 | −1.67 | ||||
| M_OverP | 0.14 | 0.15 | 0.99 | ||||
| 5 | 0.21 | 0.04 | 0.00 | ||||
| Culture | 4.75 * | 2.15 | 2.20 | ||||
| OXTR rs53576 A/G-A/A | 0.15 | 2.28 | 0.07 | ||||
| OXTR rs53576 G/G-A/A | −0.58 | 2.62 | −0.22 | ||||
| M_Care | −0.14 | 0.14 | −0.99 | ||||
| M_OverP | 0.13 | 0.15 | 0.88 | ||||
| F_Care | −0.16 | 0.12 | −1.31 | ||||
| 6 | 0.21 | 0.05 | 0.01 | ||||
| Culture | 5.11 * | 2.18 | 2.34 | ||||
| OXTR rs53576 A/G-A/A | 0.26 | 2.28 | 0.12 | ||||
| OXTR rs53576 G/G-A/A | −0.53 | 2.62 | −2.20 | ||||
| M_Care | −0.16 | 0.15 | −1.11 | ||||
| M_OverP | 0.07 | 0.16 | 0.42 | ||||
| F_Care | −0.14 | 0.13 | −1.08 | ||||
| F_OverP | 0.16 | 0.16 | 1.03 | ||||
| 7 | 0.40 | 0.16 | 0.11 * | ||||
| Culture | 11.02 | 27.75 | 0.40 | ||||
| OXTR rs53576 A/G-A/A | 14.40 | 29.92 | 0.48 | ||||
| OXTR rs53576 G/G-A/A | −14.19 | 28.76 | −0.49 | ||||
| M_Care | −0.73 | 0.85 | −0.87 | ||||
| M_OverP | 5.03 ** | 1.71 | 2.95 | ||||
| F_Care | −1.52 * | 0.75 | −2.04 | ||||
| F_OverP | −1.71 | 0.87 | −1.96 | ||||
| Culture xOXTR rs53576 A/G-A/A | −27.14 | 32.77 | −0.83 | ||||
| Culture x OXTR rs53576 G/G-A/A | −12.77 | 33.42 | −0.38 | ||||
| Culture x M_Care | −0.10 | 0.91 | −0.11 | ||||
| Culture x M_OverP | −4.89 ** | 1.74 | −2.81 | ||||
| Culture x F_Care | 1.74 * | 0.80 | 2.19 | ||||
| Culture x F_OverP | 1.73 | 0.93 | 1.87 | ||||
| OXTR rs53576 A/G-A/A x M_Care | 0.04 | 0.97 | 0.05 | ||||
| OXTR rs53576 G/G-A/A x M_Care | 0.78 | 0.89 | 0.87 | ||||
| OXTR rs53576 A/G-A/A x M_OverP | −5.27 ** | 1.79 | −2.96 | ||||
| OXTR rs53576 G/G-A/A x M_OverP | −4.57 ** | 1.76 | −2.59 | ||||
| OXTR rs53576 A/G-A/A x F_Care | 1.25 | 0.86 | 1.46 | ||||
| OXTR rs53576 G/G-A/A x F_Care | 1.60 * | 0.80 | 2.01 | ||||
| OXTR rs53576 A/G-A/A x F_OverP | 2.18 * | 1.06 | 2.06 | ||||
| OXTR rs53576 G/G-A/A x F_OverP | 1.27 | 0.96 | 1.33 | ||||
| Culture x OXTR rs53576 A/G-A/A x M_Care | 1.24 | 1.06 | 1.17 | ||||
| Culture x OXTR rs53576 G/G-A/A x M_Care | −0.11 | 1.03 | −0.11 | ||||
| Culture x OXTR rs53576 A/G-A/A x M_OverP | 4.91 ** | 1.83 | 2.67 | ||||
| Culture x OXTR rs53576 G/G-A/A x M_OverP | 4.79 ** | 1.84 | 2.61 | ||||
| Culture x OXTR rs53576 A/G-A/A x F_Care | −2.06 * | 0.93 | −2.22 | ||||
| Culture x OXTR rs53576 G/G-A/A x F_Care | −1.44 | 0.90 | −1.60 | ||||
| Culture x OXTR rs53576 A/G-A/A x F_OverP | −1.88 | 1.13 | −1.66 | ||||
| Culture x OXTR rs53576 G/G-A/A x F_OverP | −1.09 | 1.10 | −0.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cataldo, I.; Bonassi, A.; Lepri, B.; Foo, J.N.; Setoh, P.; Esposito, G. Recalled Parental Bonding Interacts with Oxytocin Receptor Gene Polymorphism in Modulating Anxiety and Avoidance in Adult Relationships. Brain Sci. 2021, 11, 496. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11040496
Cataldo I, Bonassi A, Lepri B, Foo JN, Setoh P, Esposito G. Recalled Parental Bonding Interacts with Oxytocin Receptor Gene Polymorphism in Modulating Anxiety and Avoidance in Adult Relationships. Brain Sciences. 2021; 11(4):496. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11040496
Chicago/Turabian StyleCataldo, Ilaria, Andrea Bonassi, Bruno Lepri, Jia Nee Foo, Peipei Setoh, and Gianluca Esposito. 2021. "Recalled Parental Bonding Interacts with Oxytocin Receptor Gene Polymorphism in Modulating Anxiety and Avoidance in Adult Relationships" Brain Sciences 11, no. 4: 496. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11040496