
NLM-HS: Navigation Learning Model Based on a Hippocampal–Striatal Circuit for Explaining Navigation Mechanisms in Animal Brains


Abstract
:1. Introduction
2. Materials and Methods
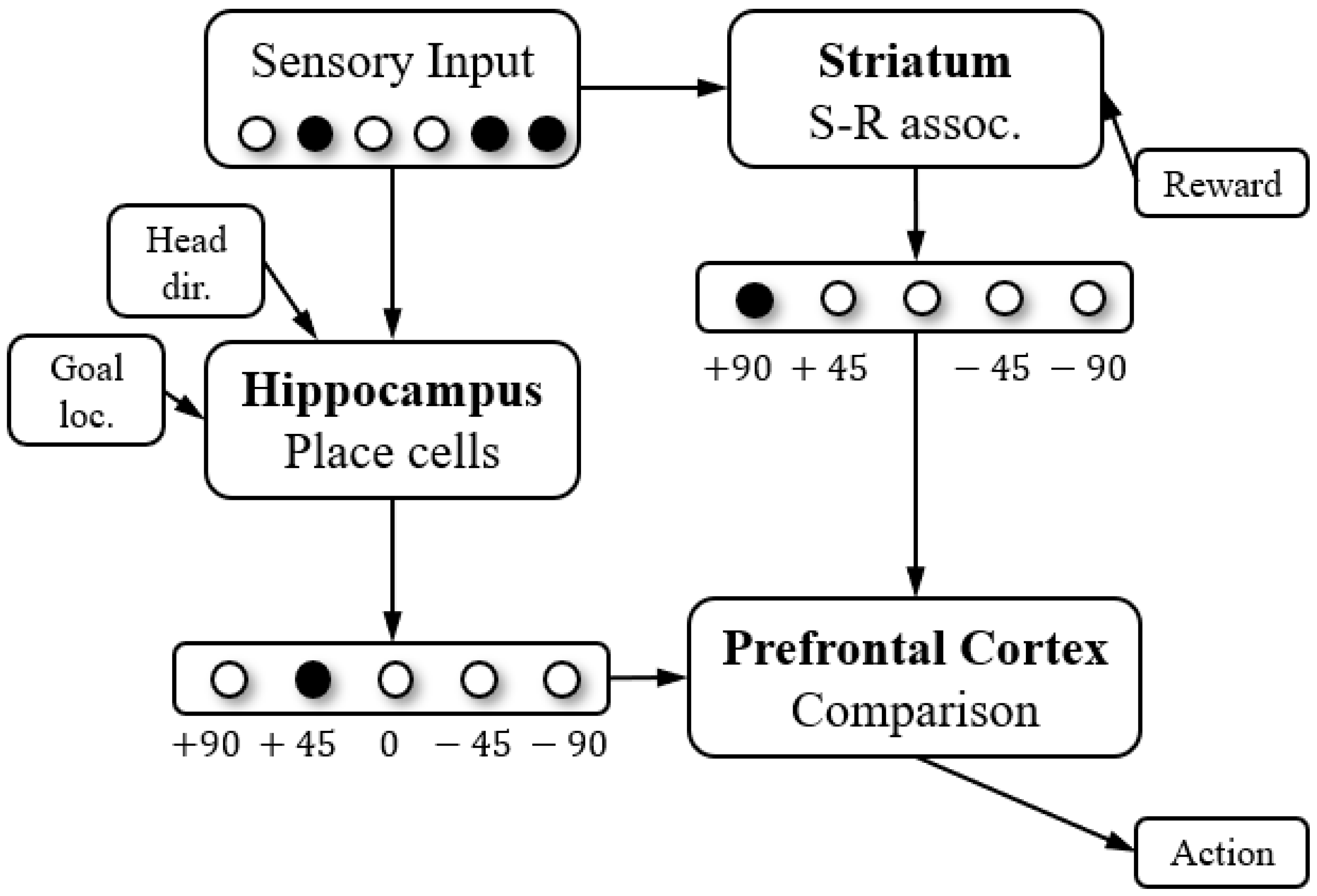
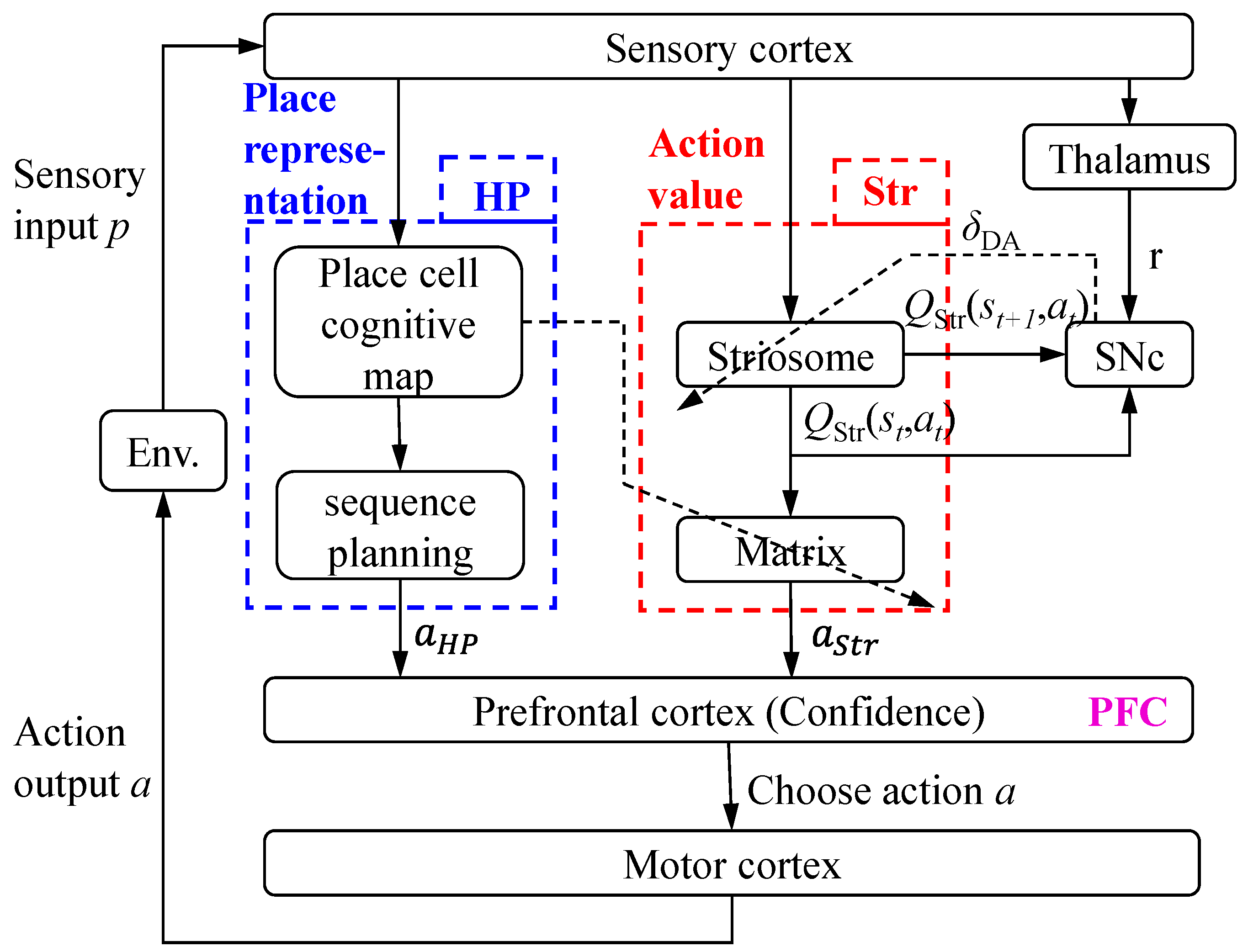
2.1. Model Construction
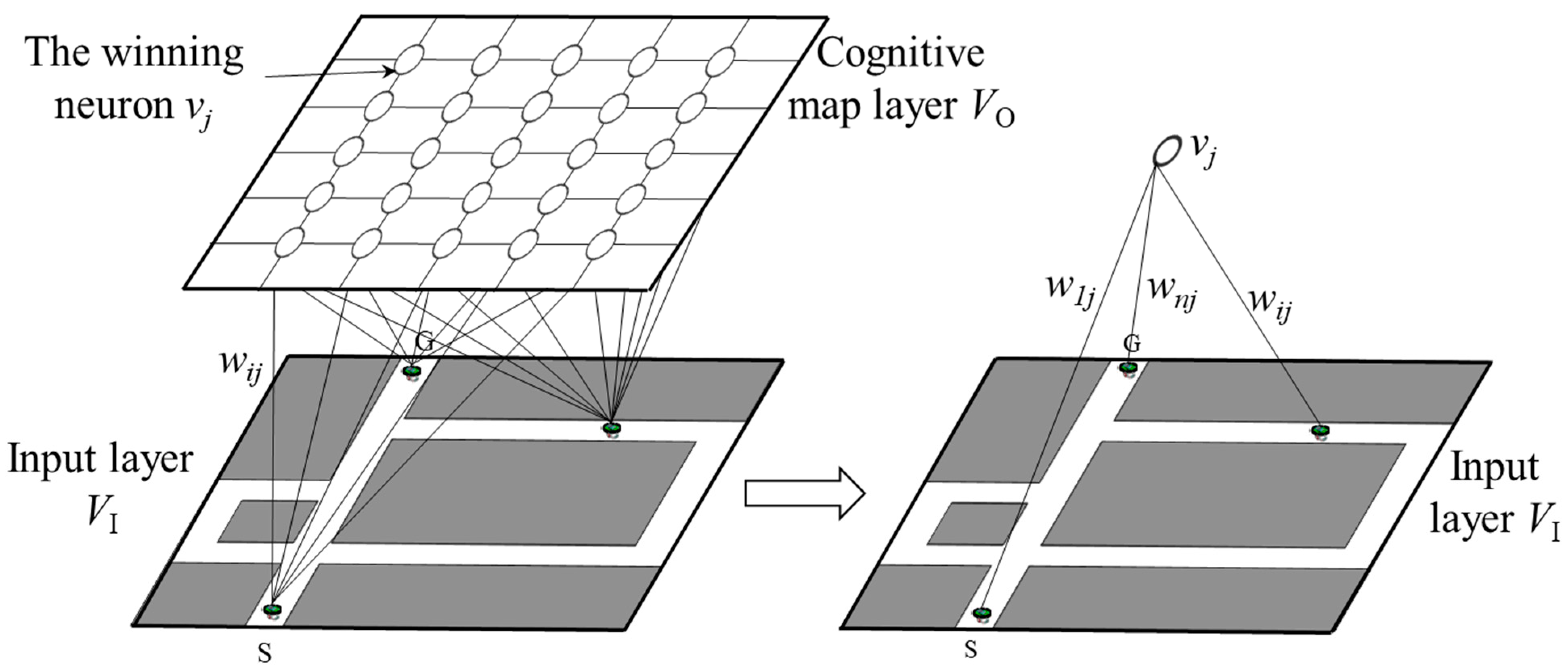
2.2. Model of the Hippocampus
2.3. Model of the Striatum
2.4. Model of the Prefrontal Cortex
3. Results
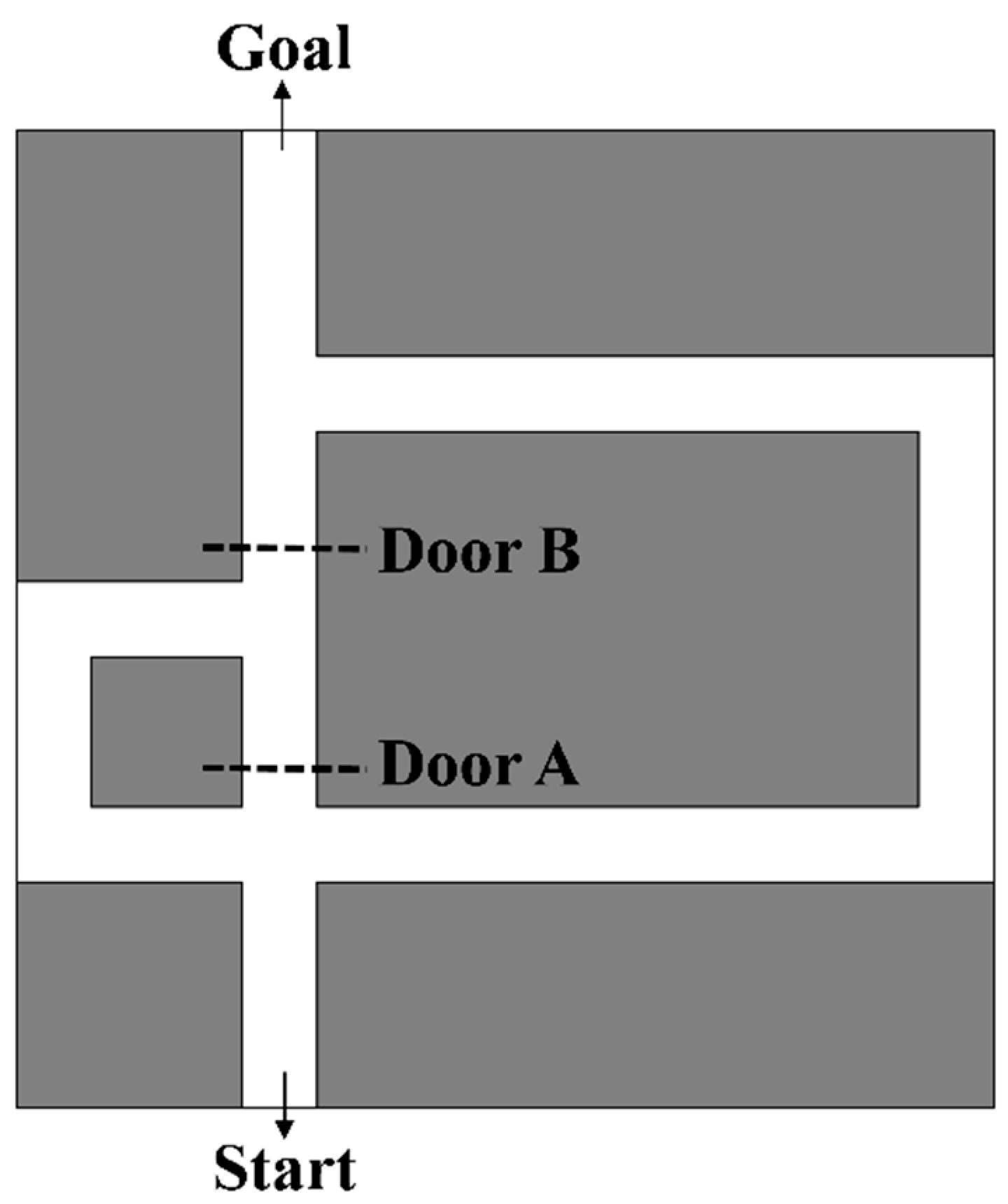
3.1. Experiment Design
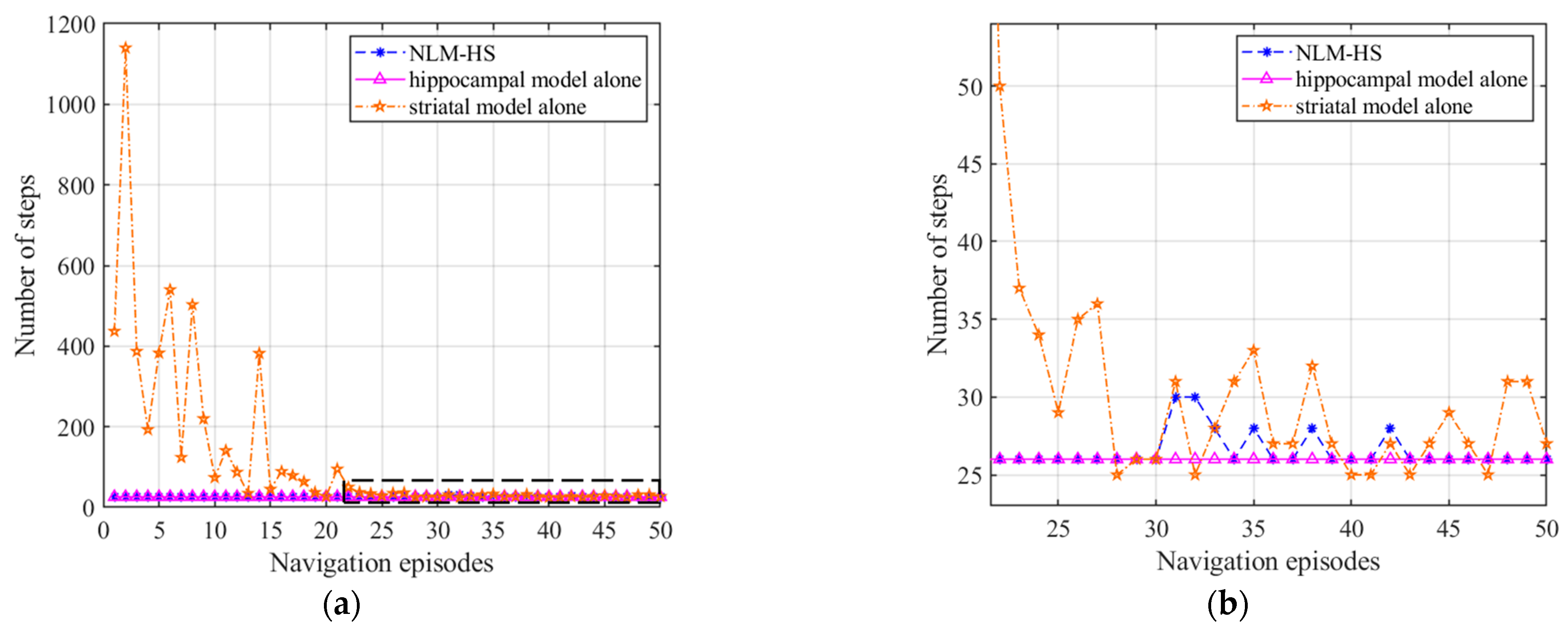
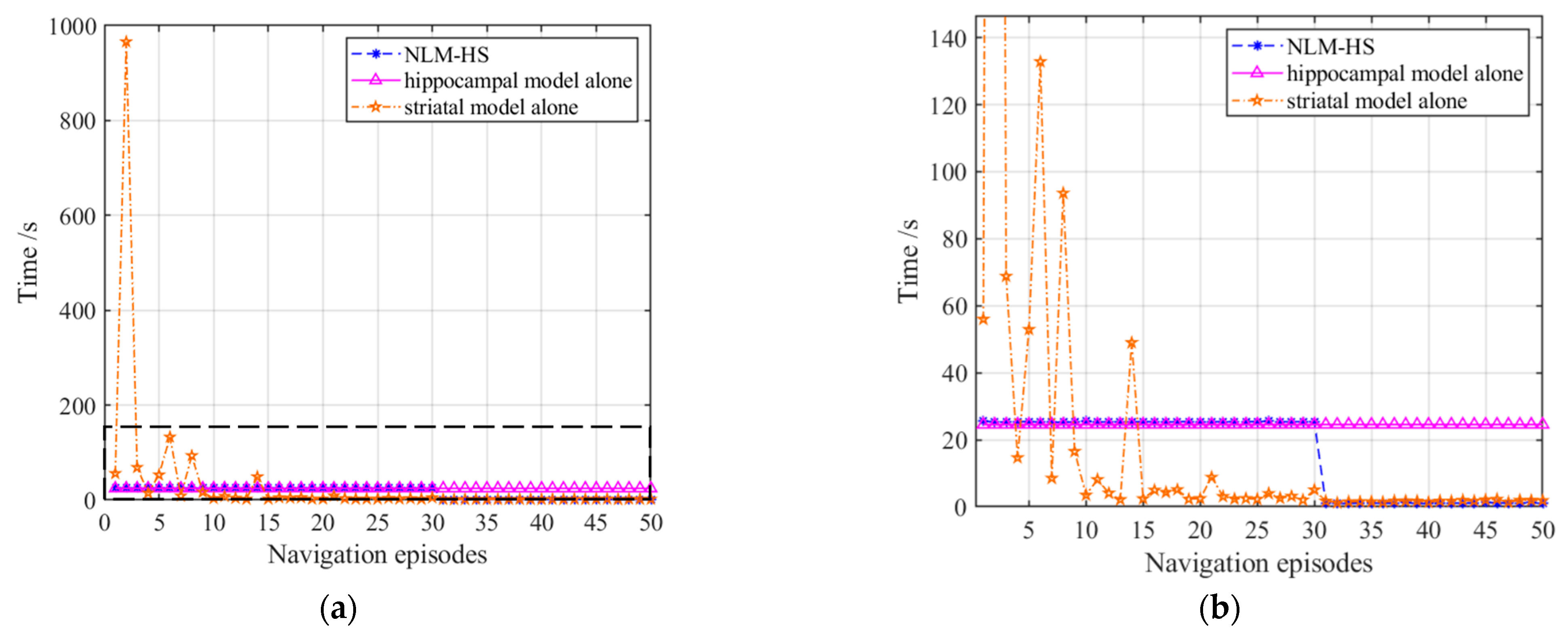
3.2. Basic Navigation Experiments
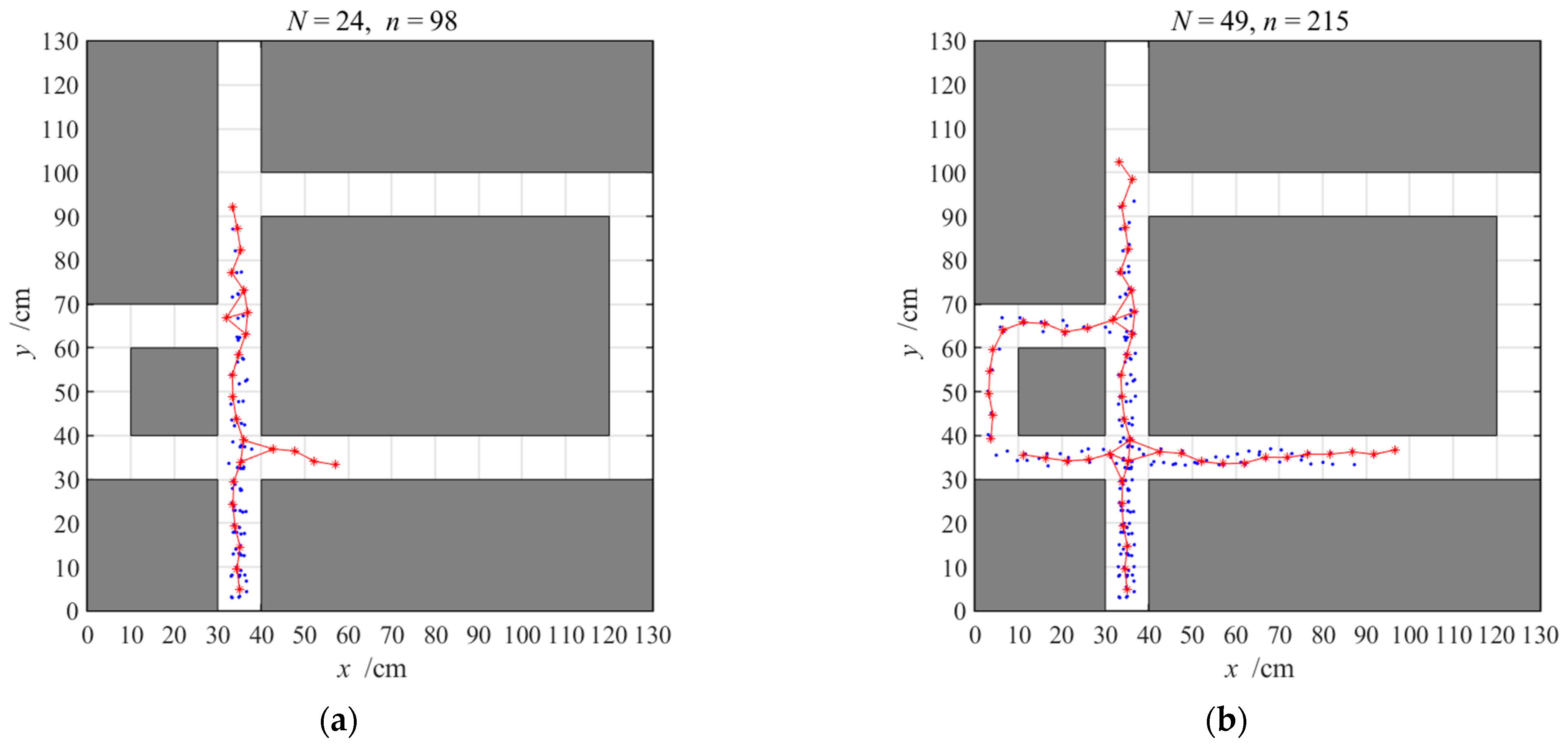
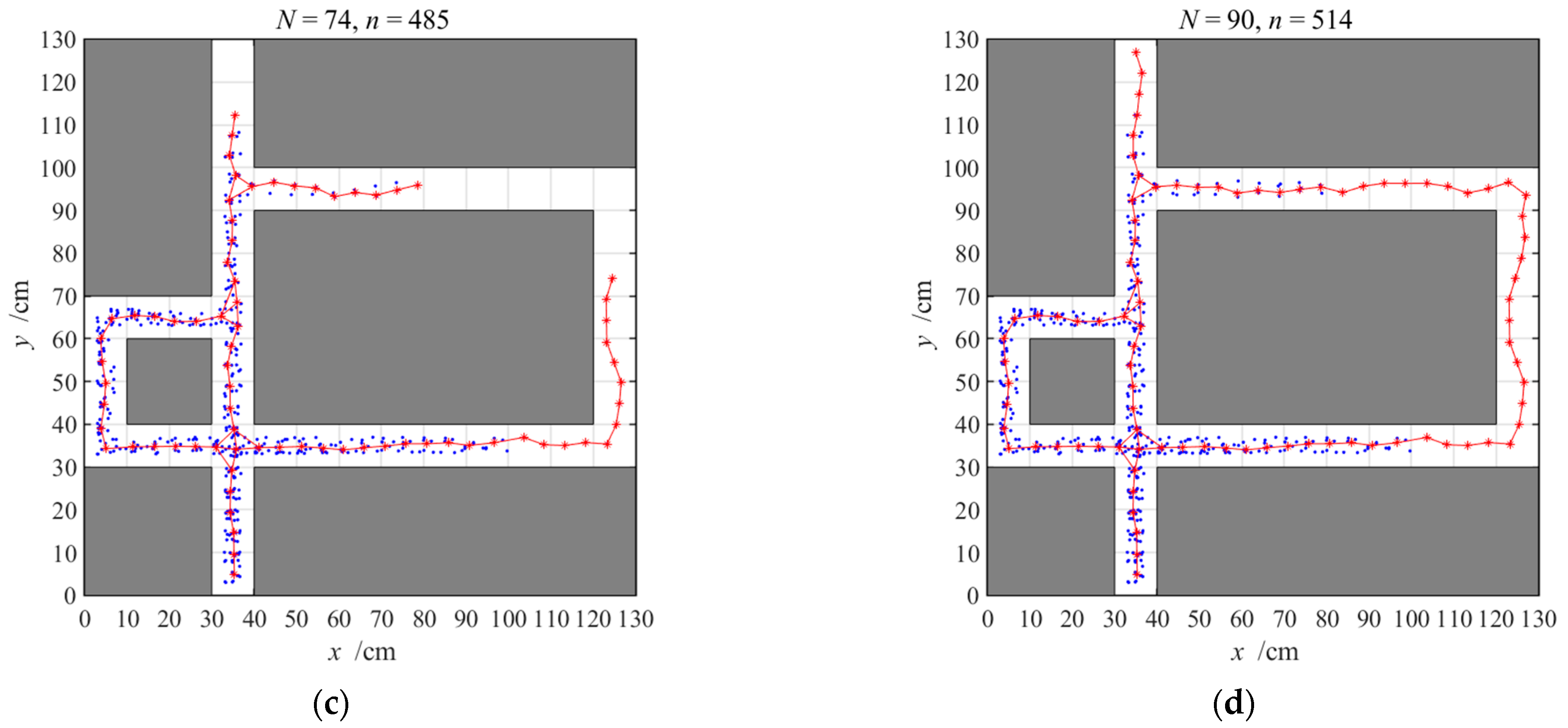
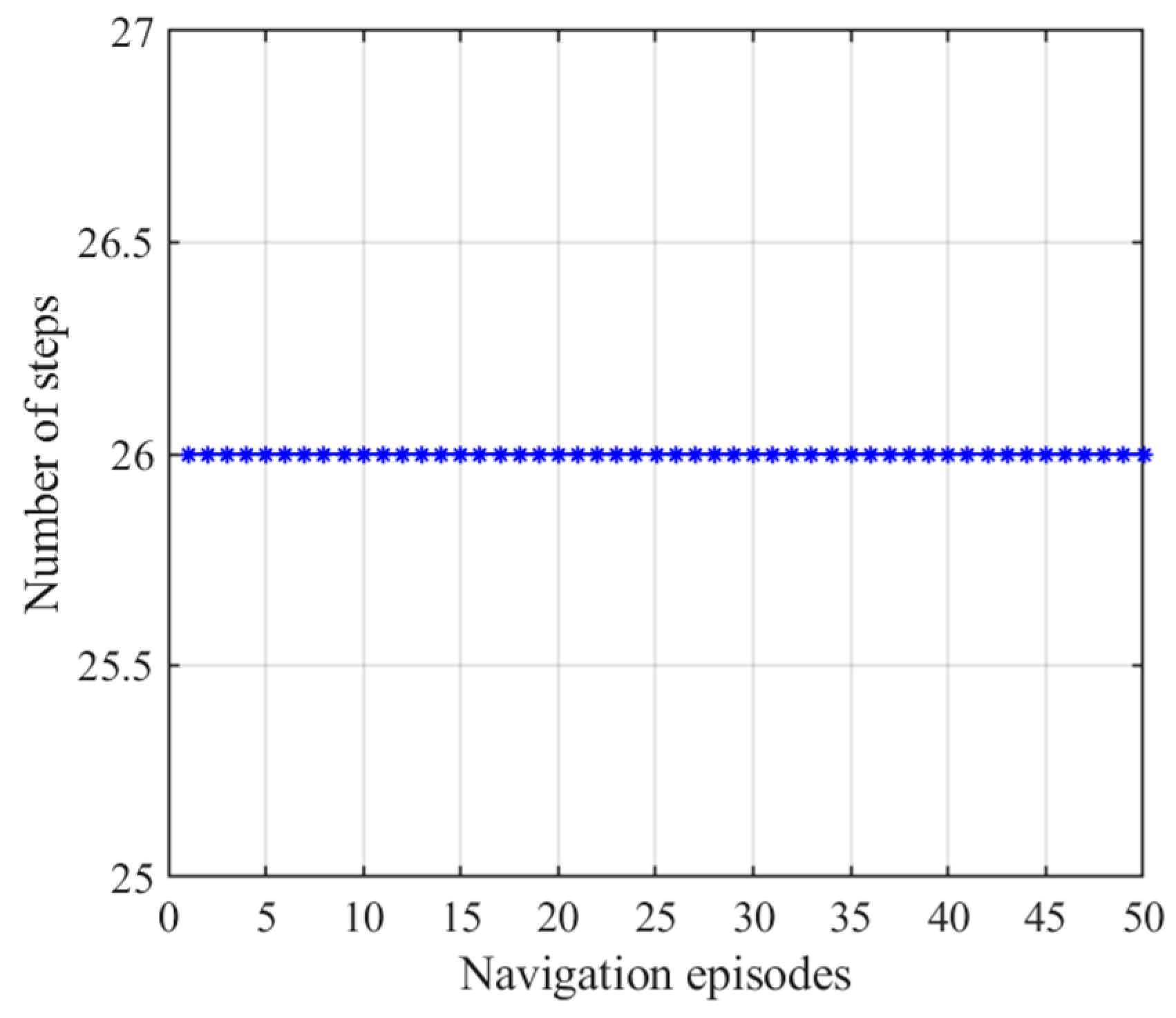
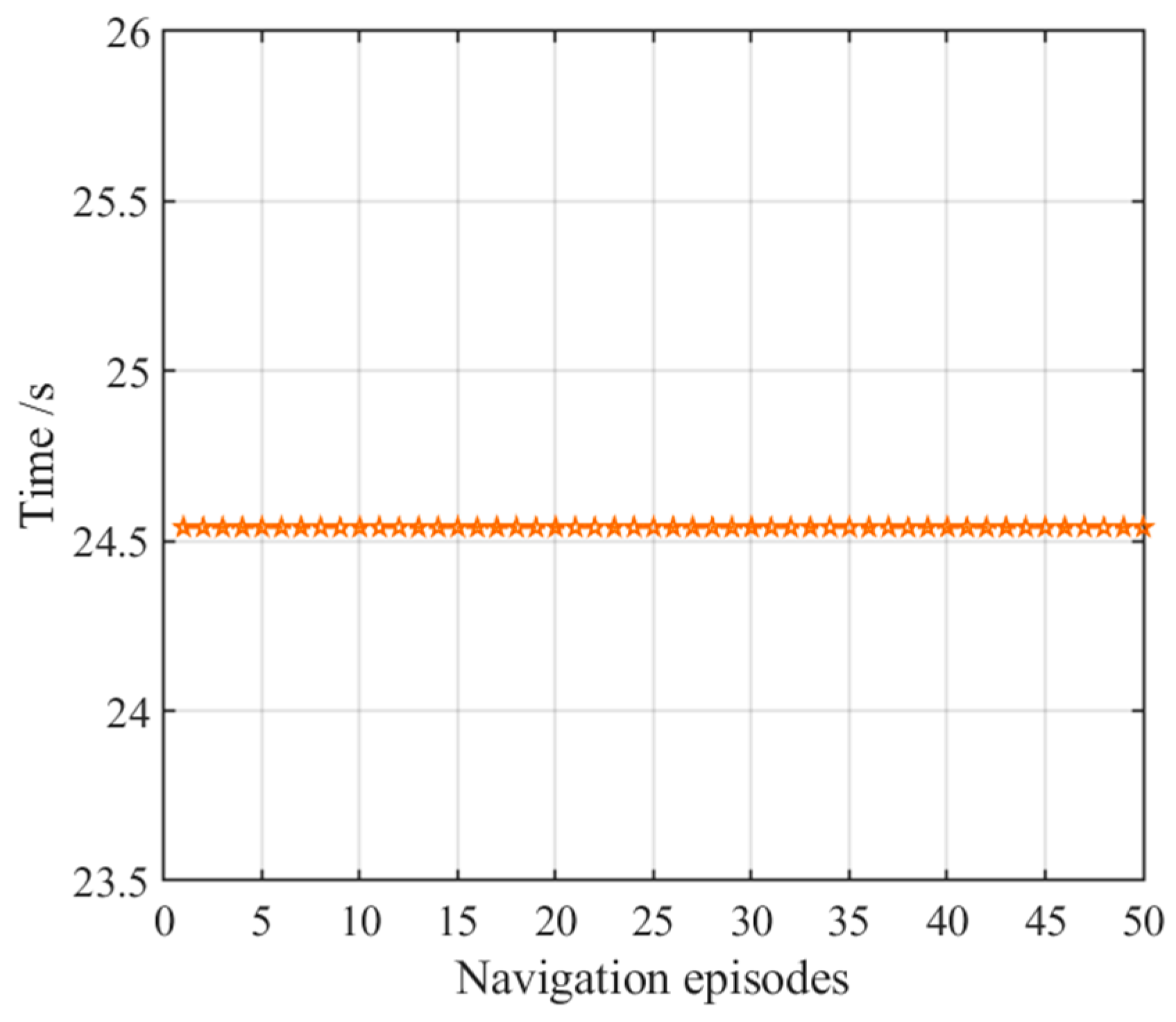
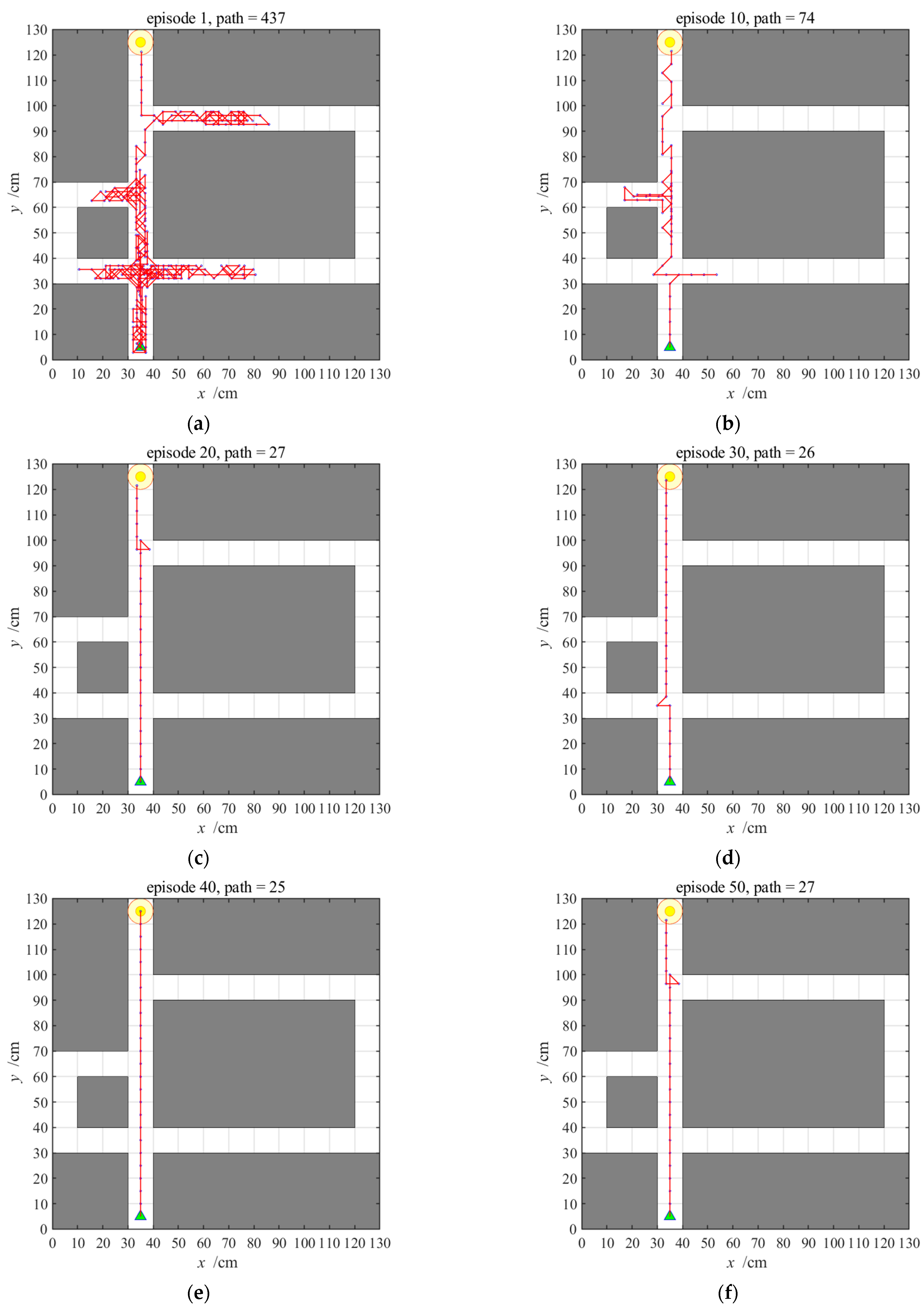
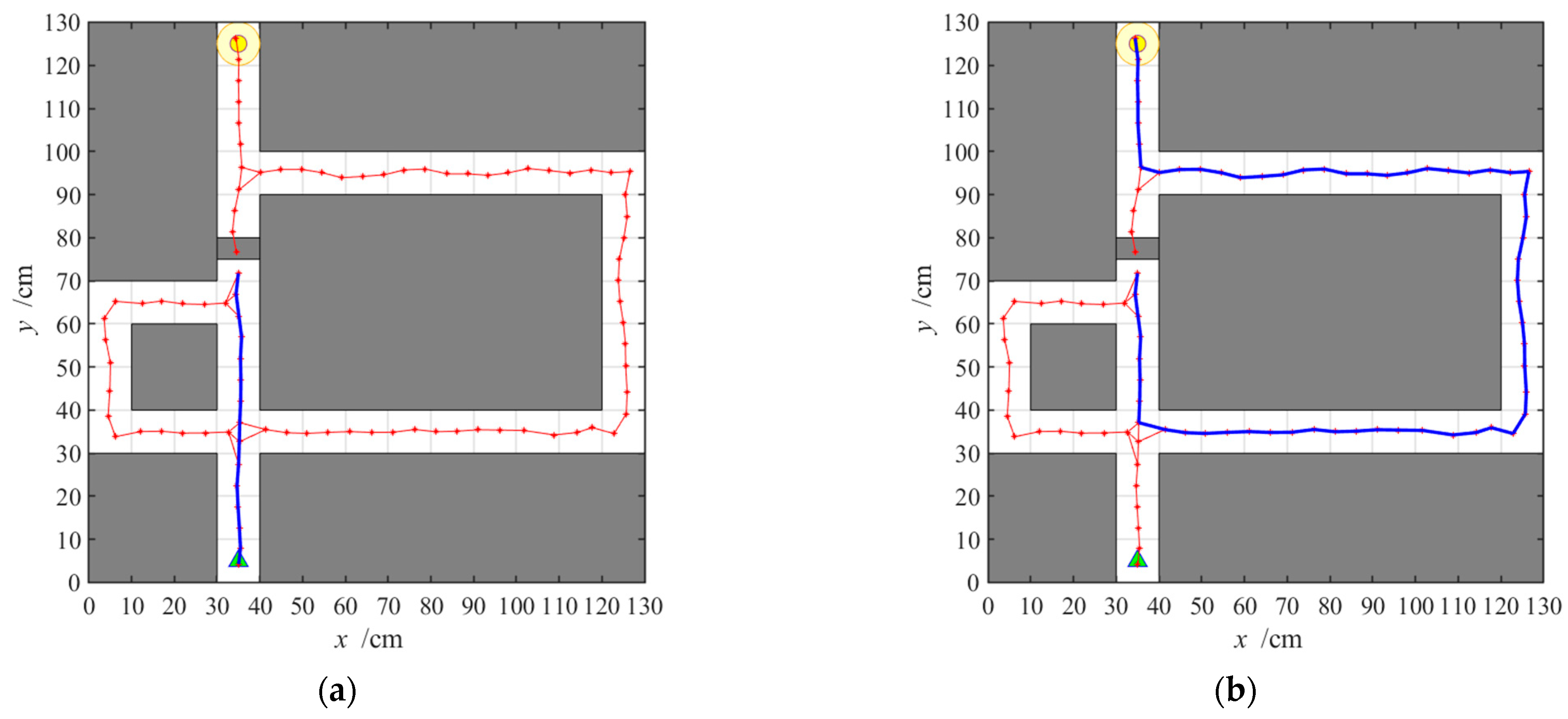
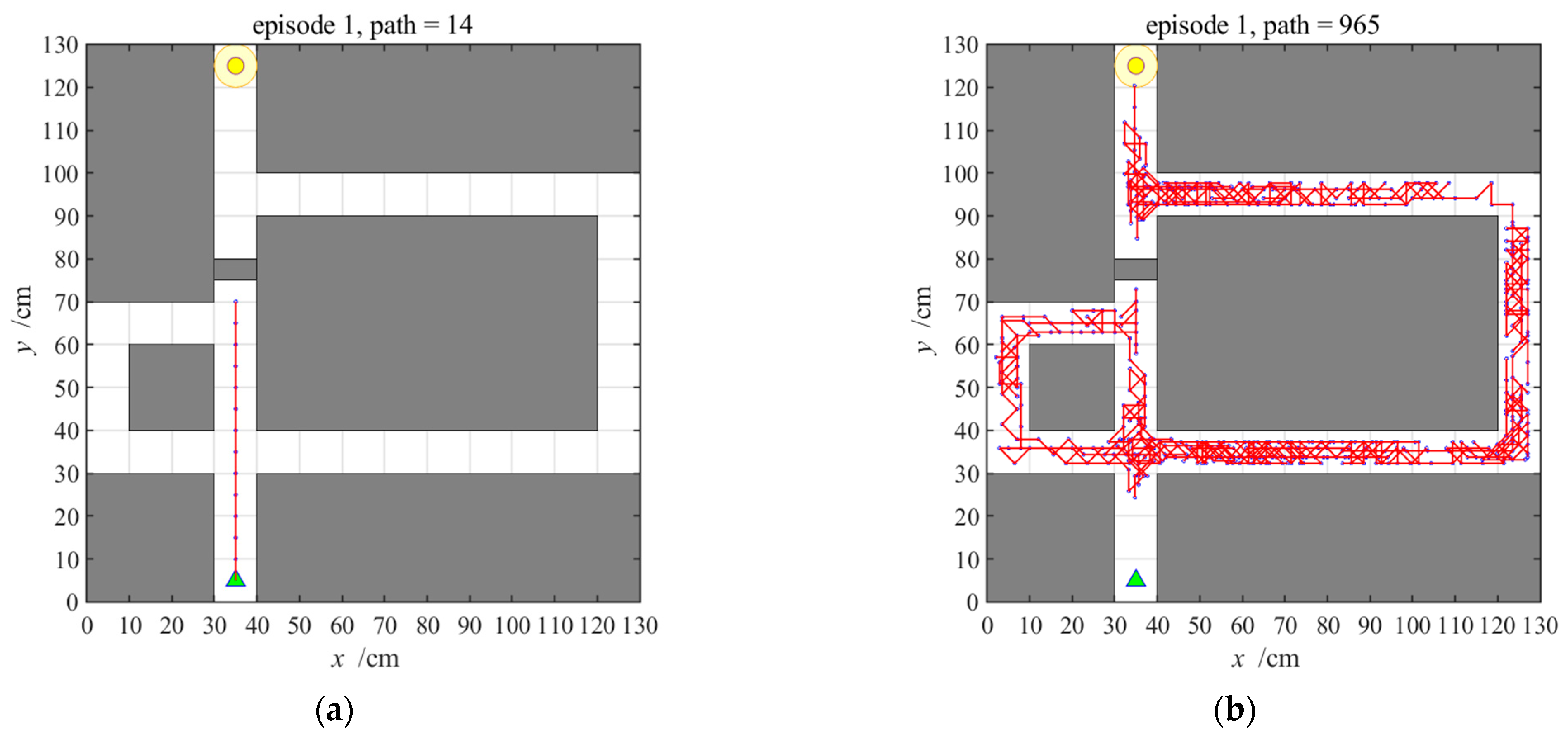
3.2.1. Navigation with the Hippocampal Model Alone
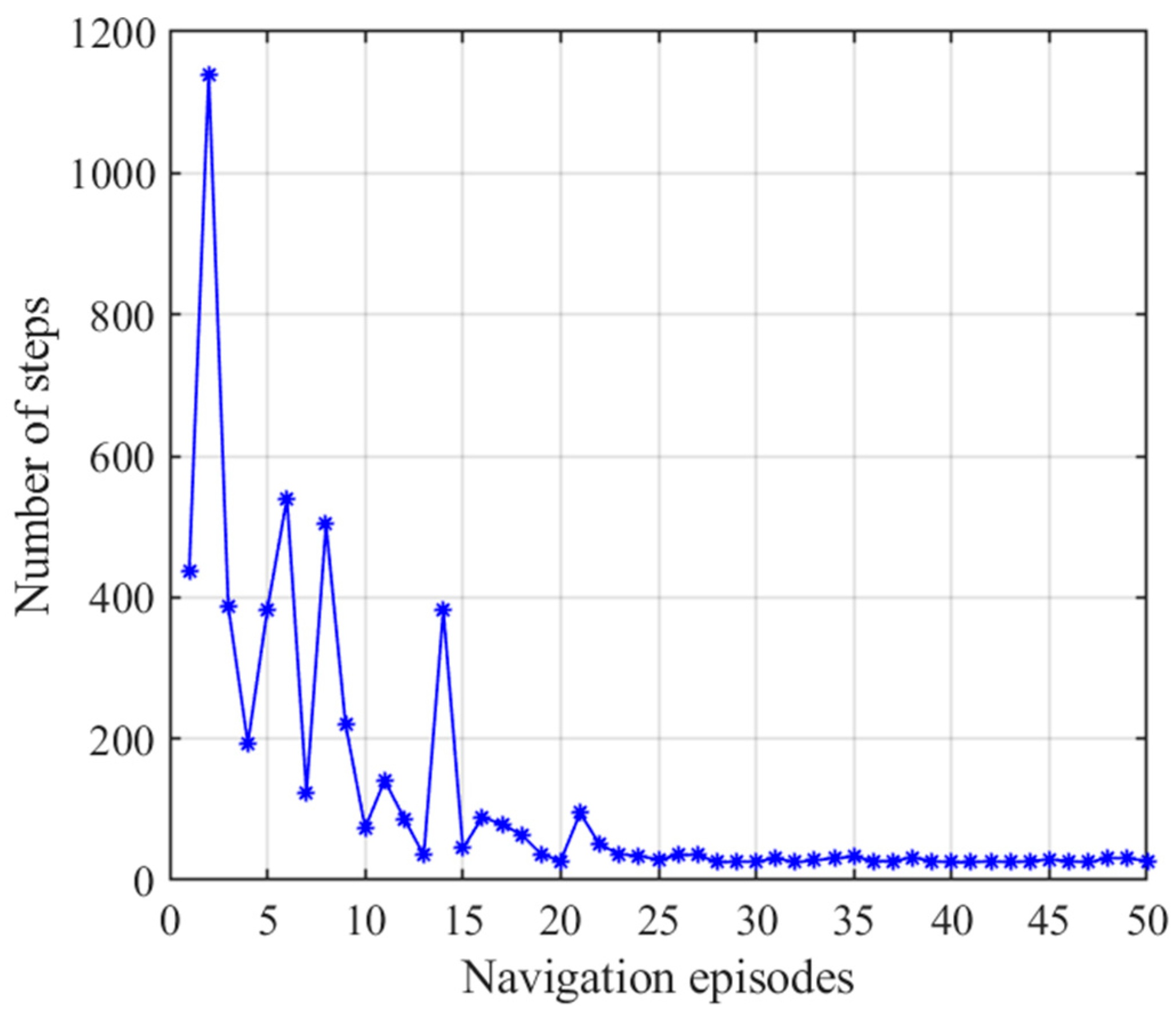
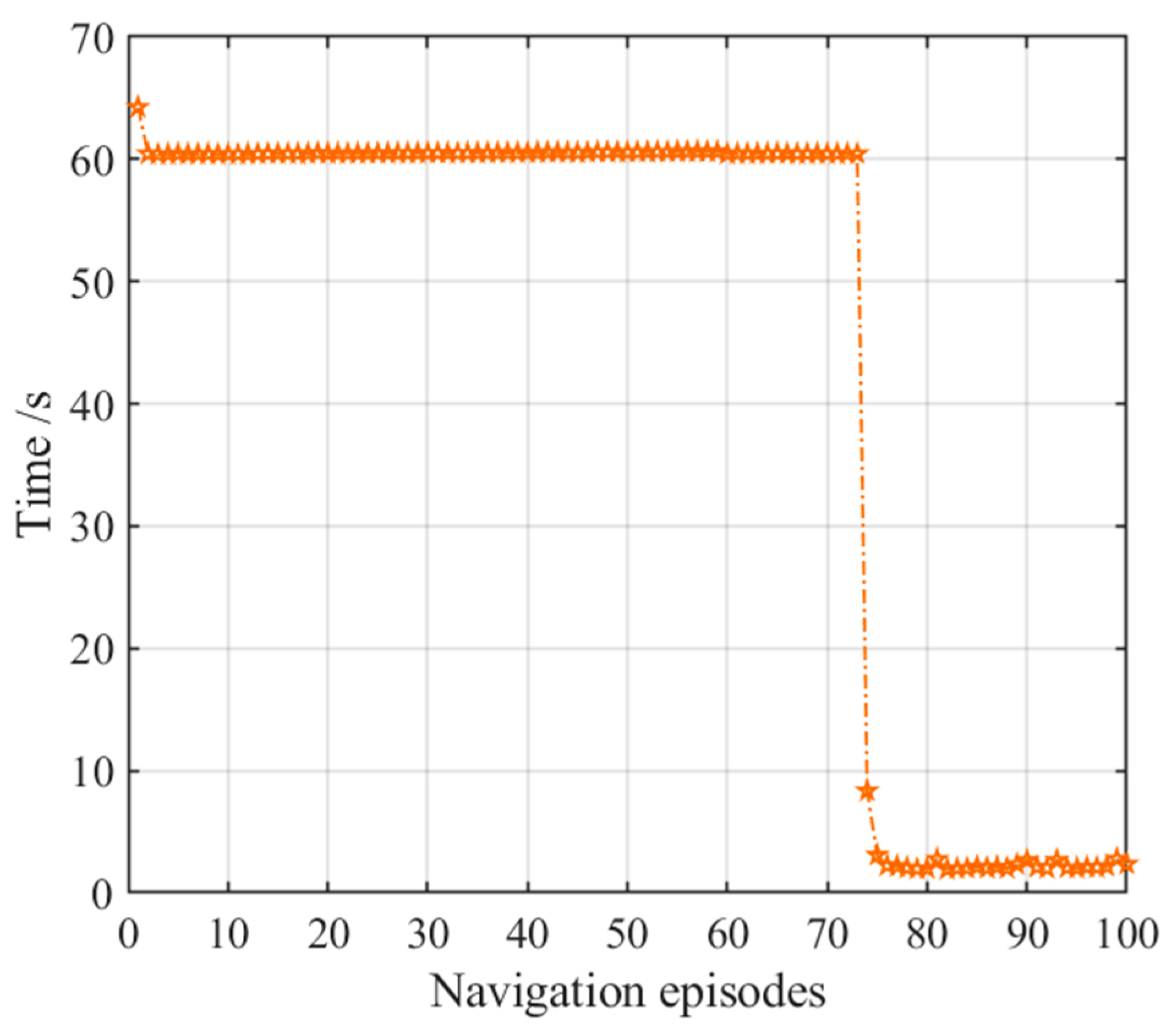
3.2.2. Navigation with the Striatal Model Alone
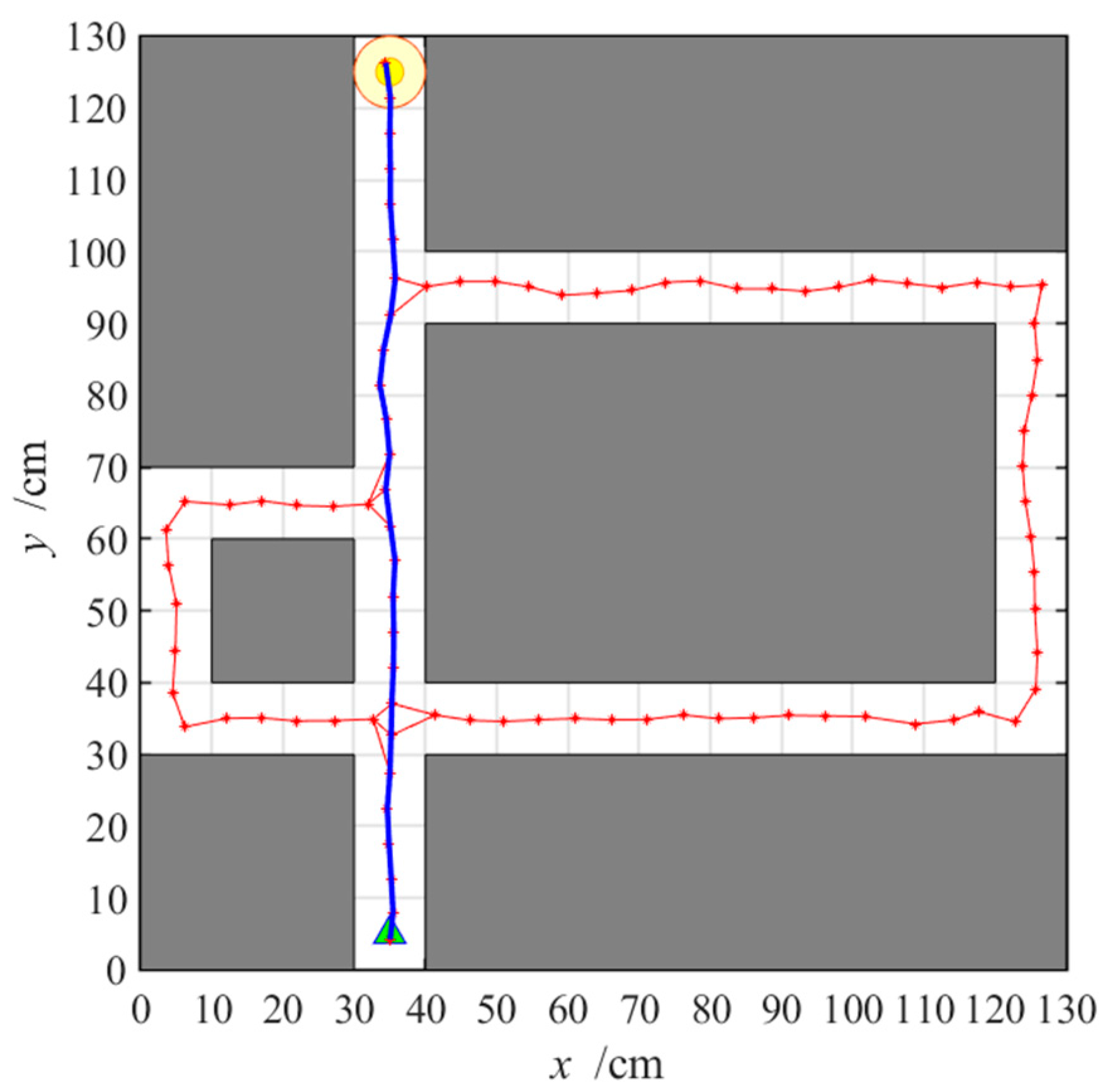
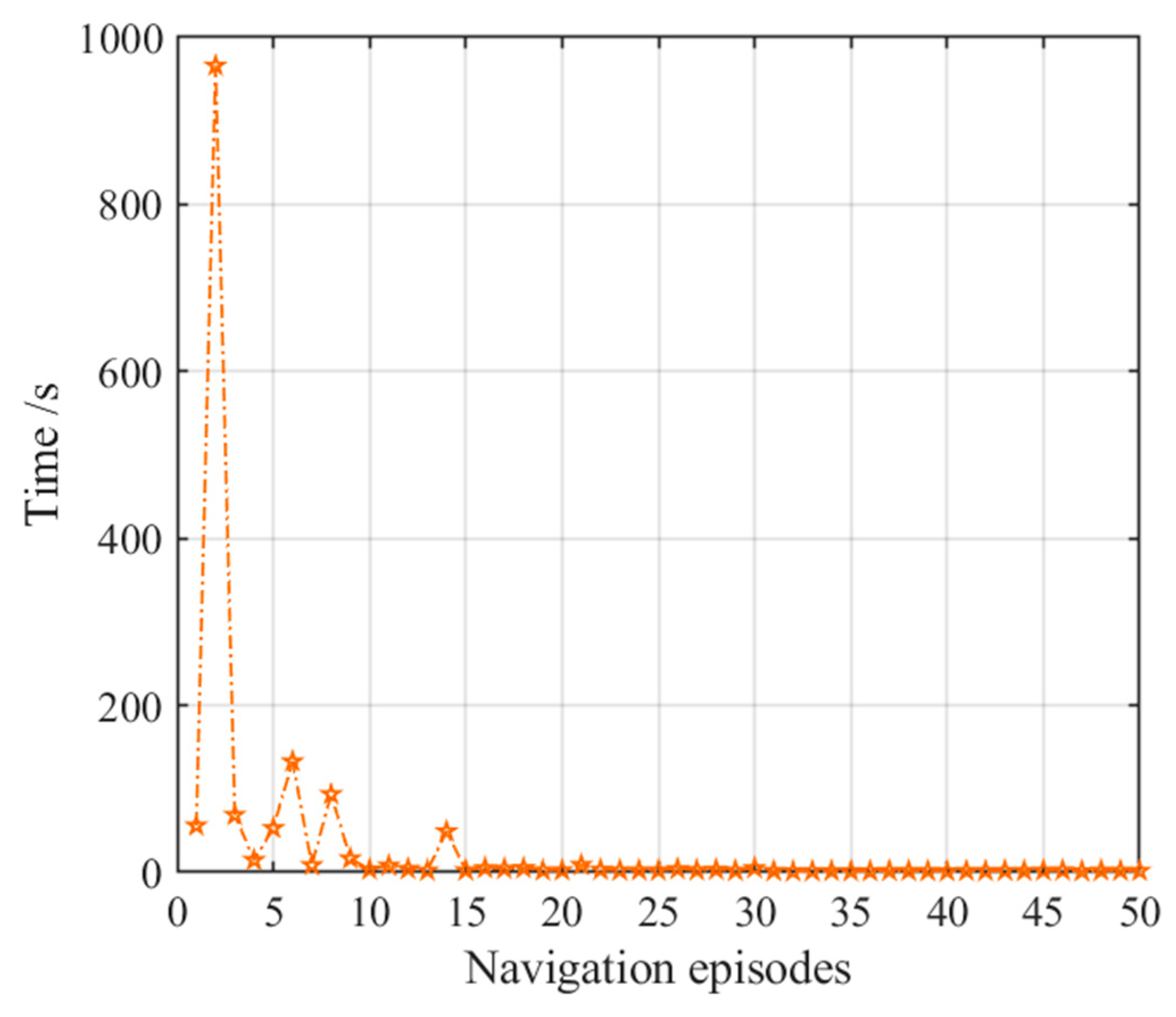
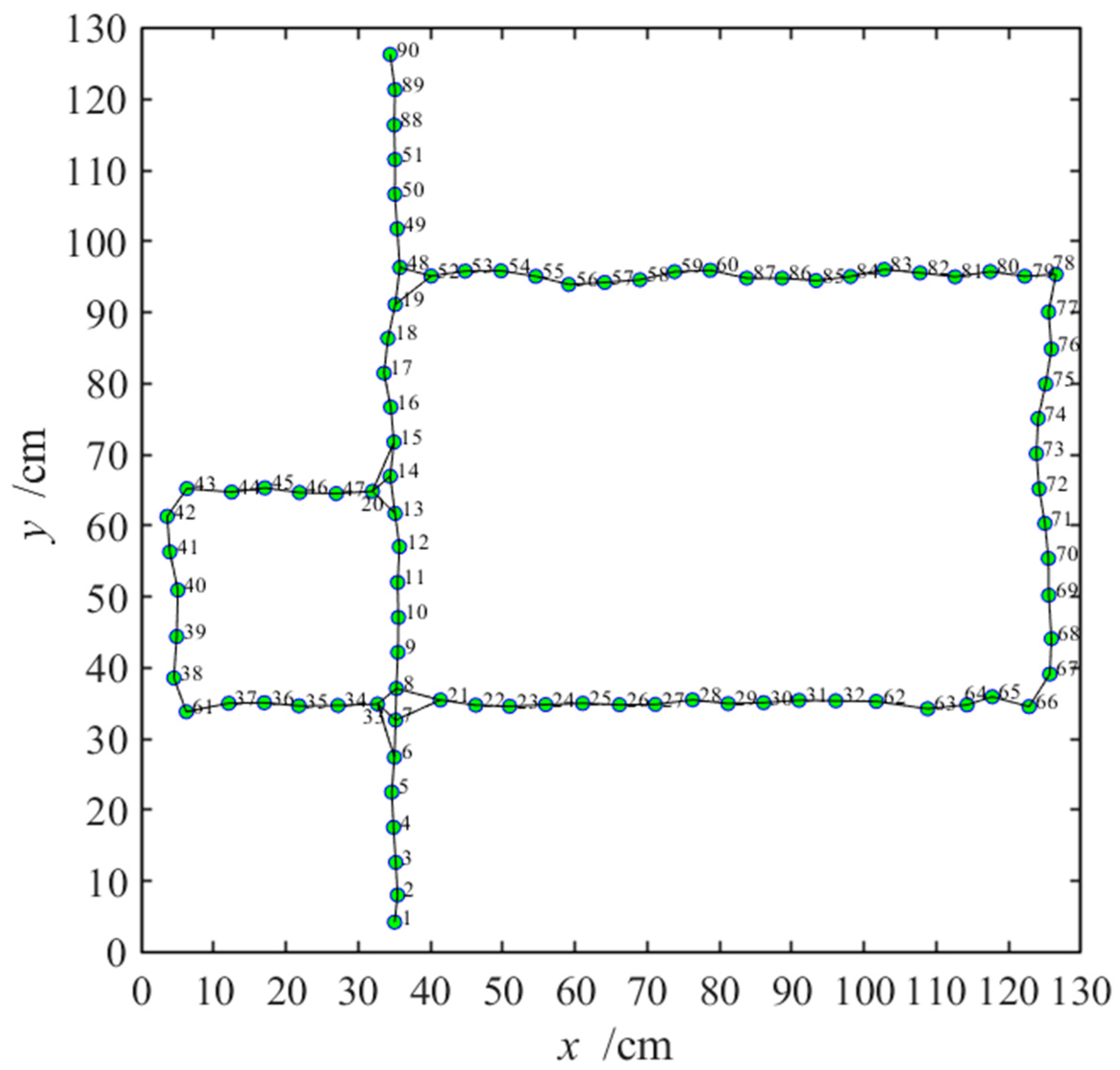
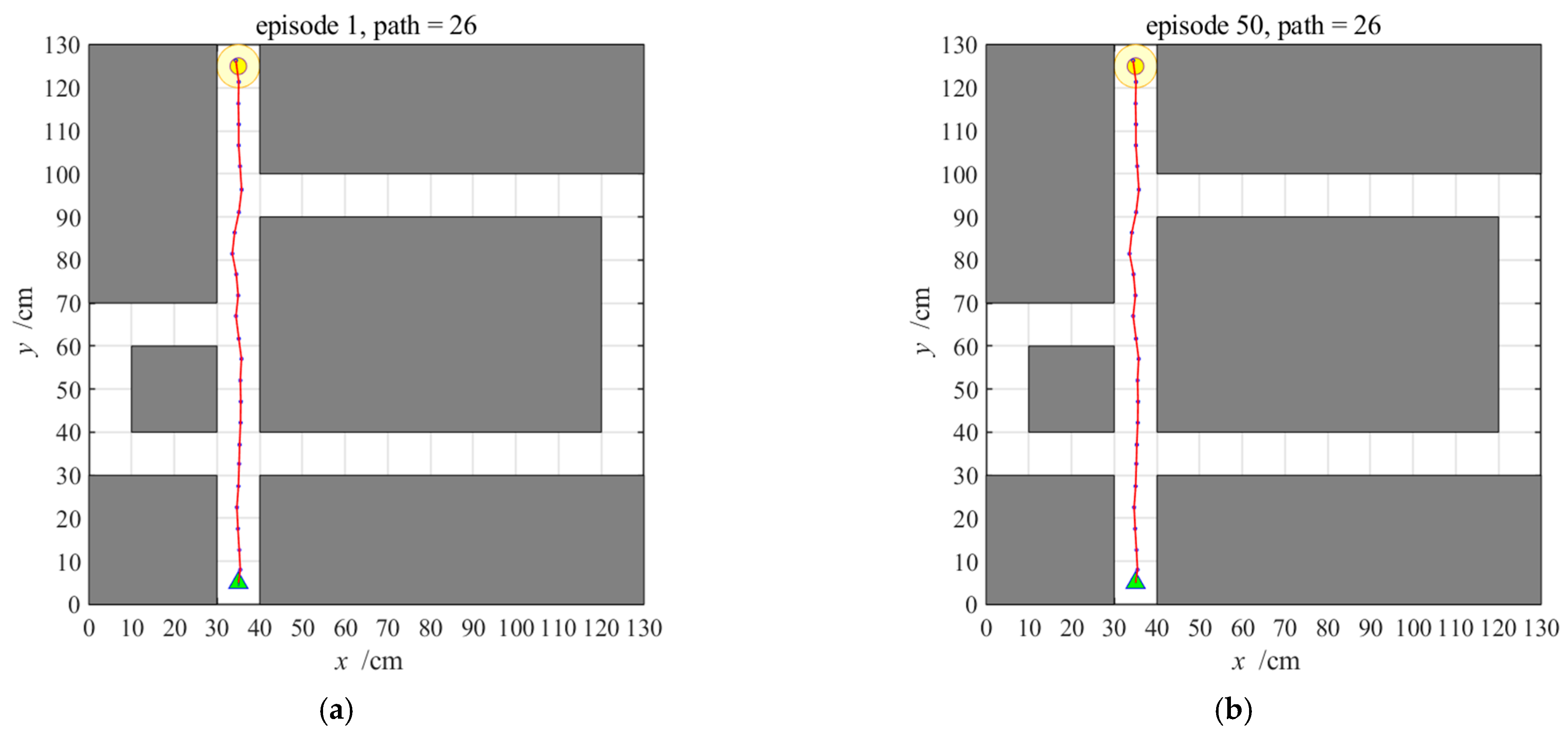
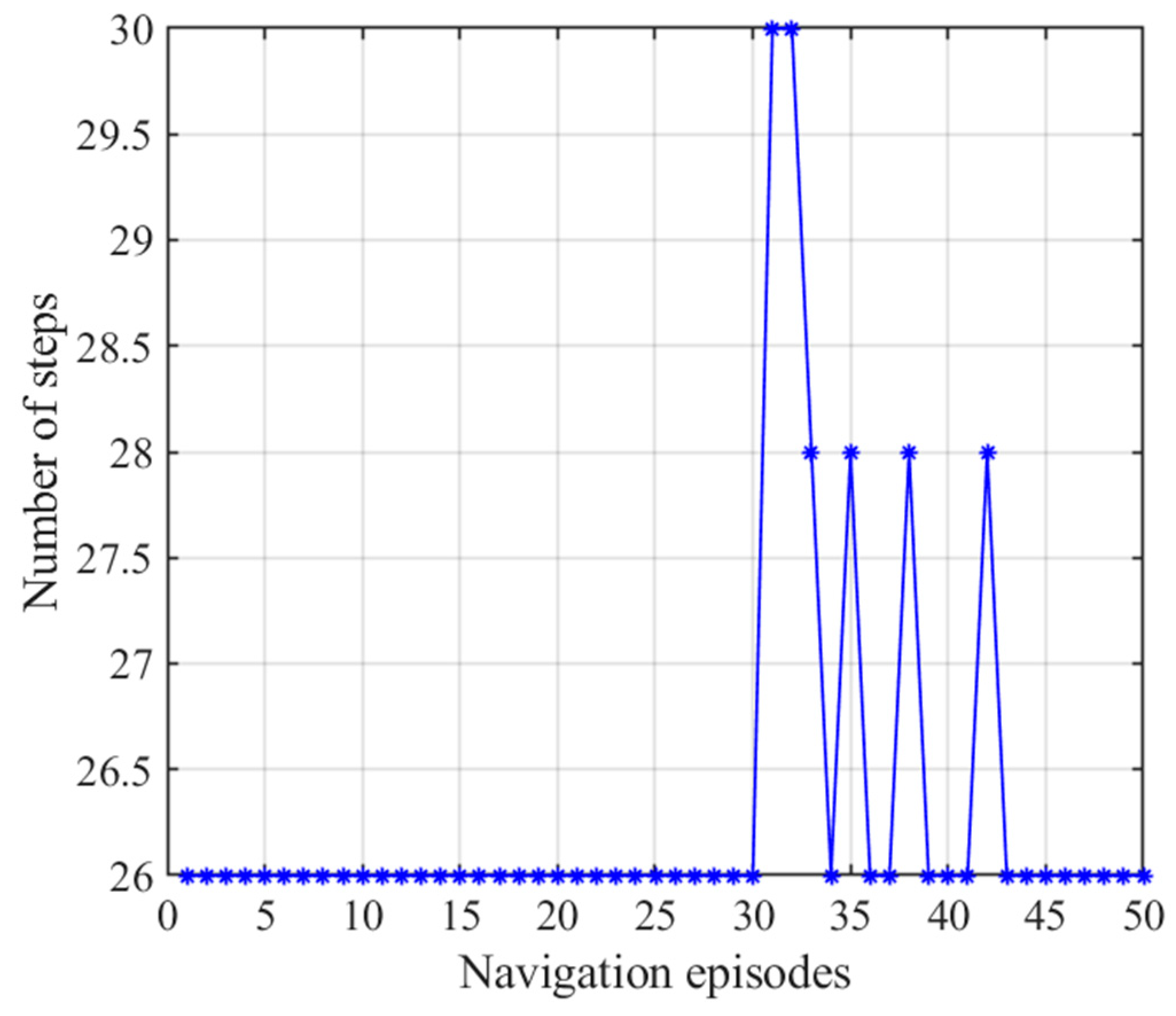
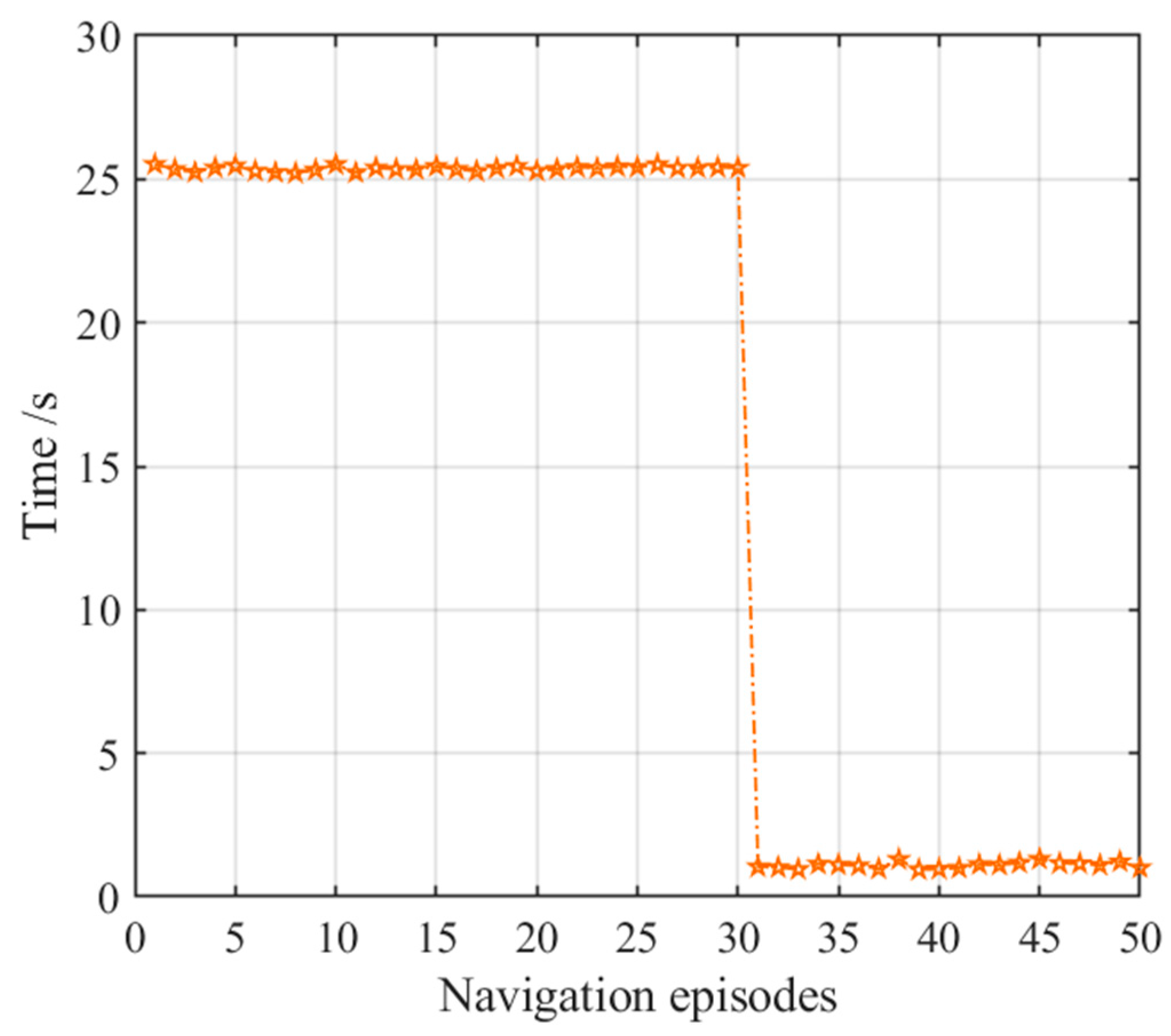
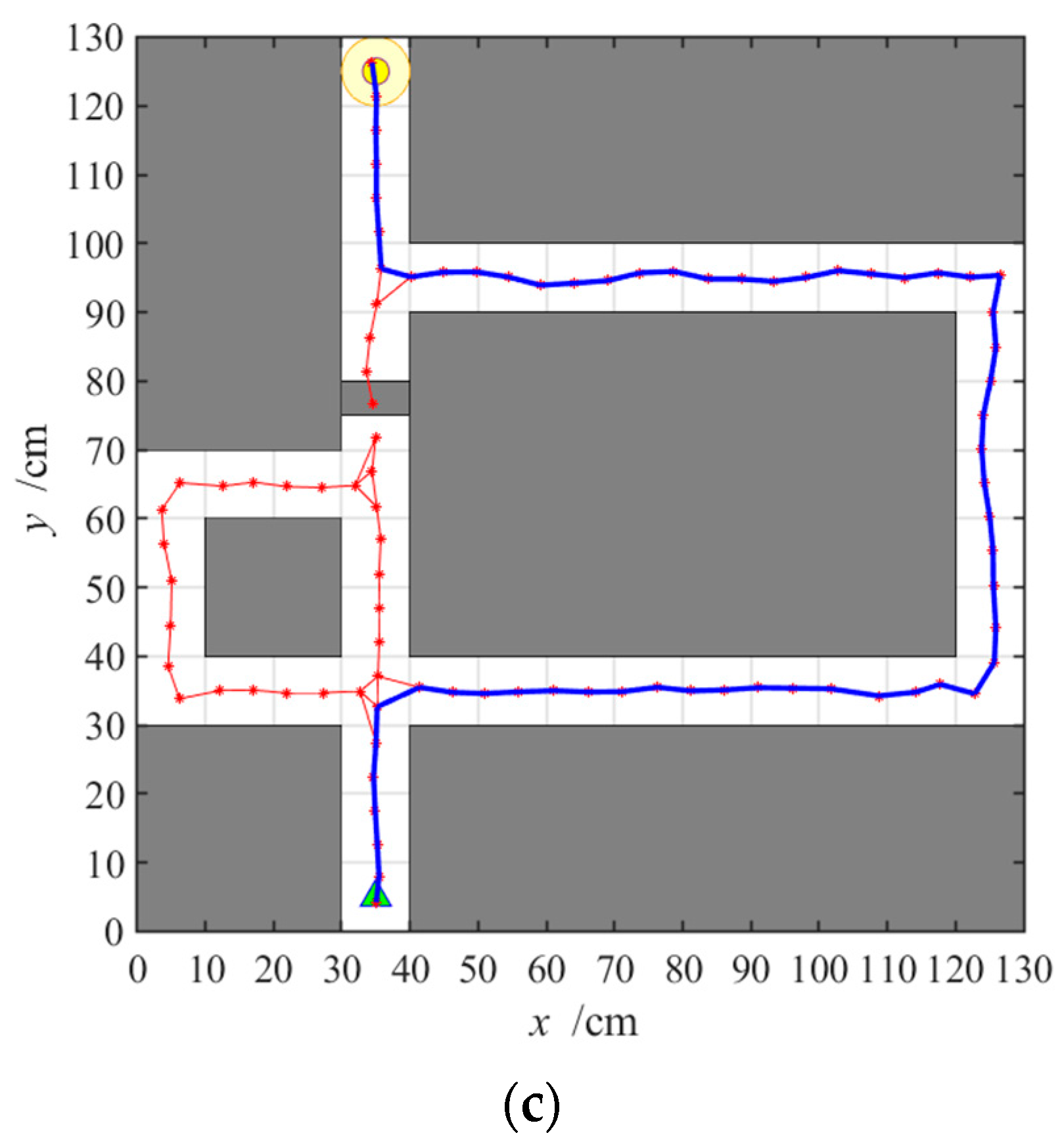
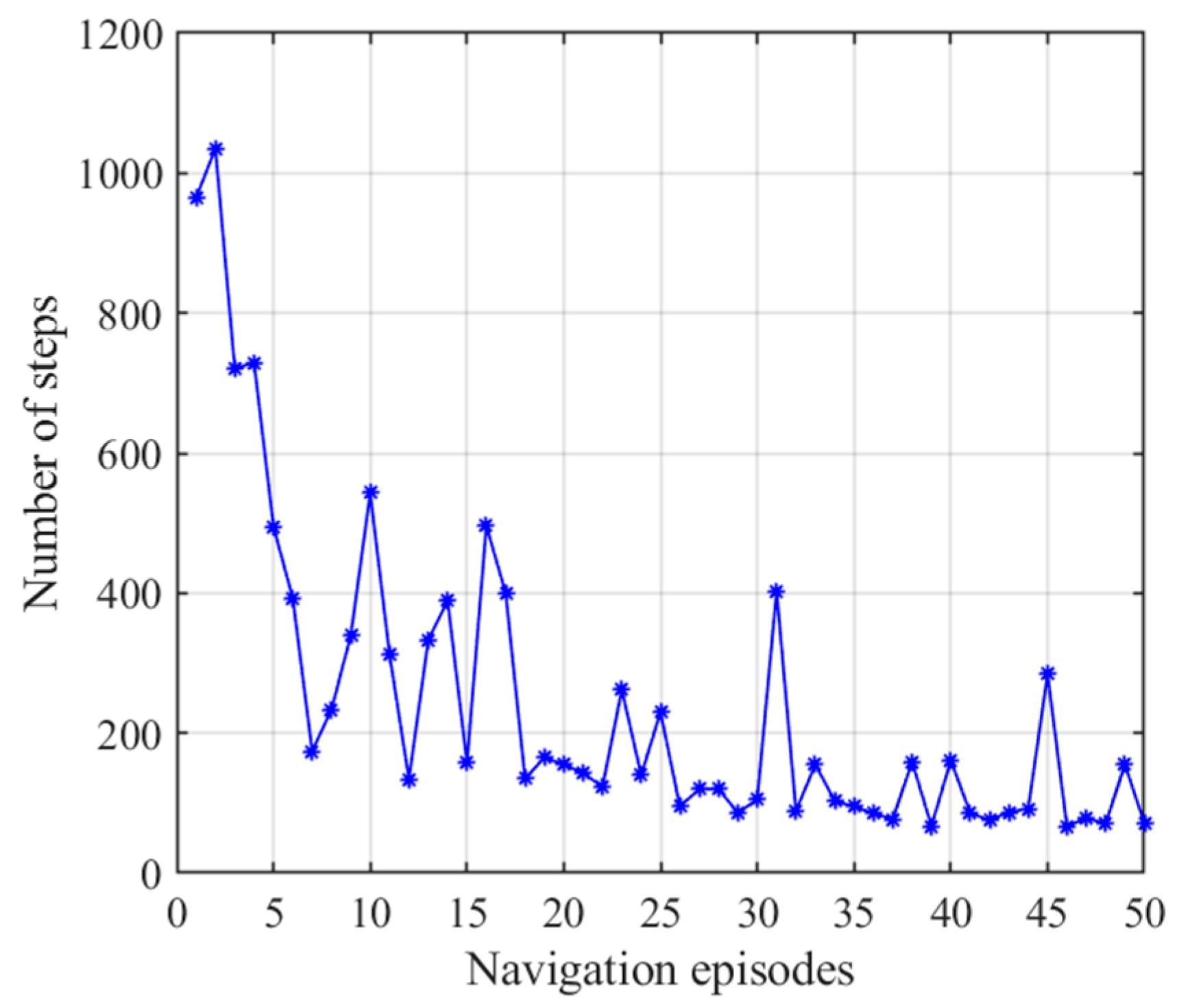
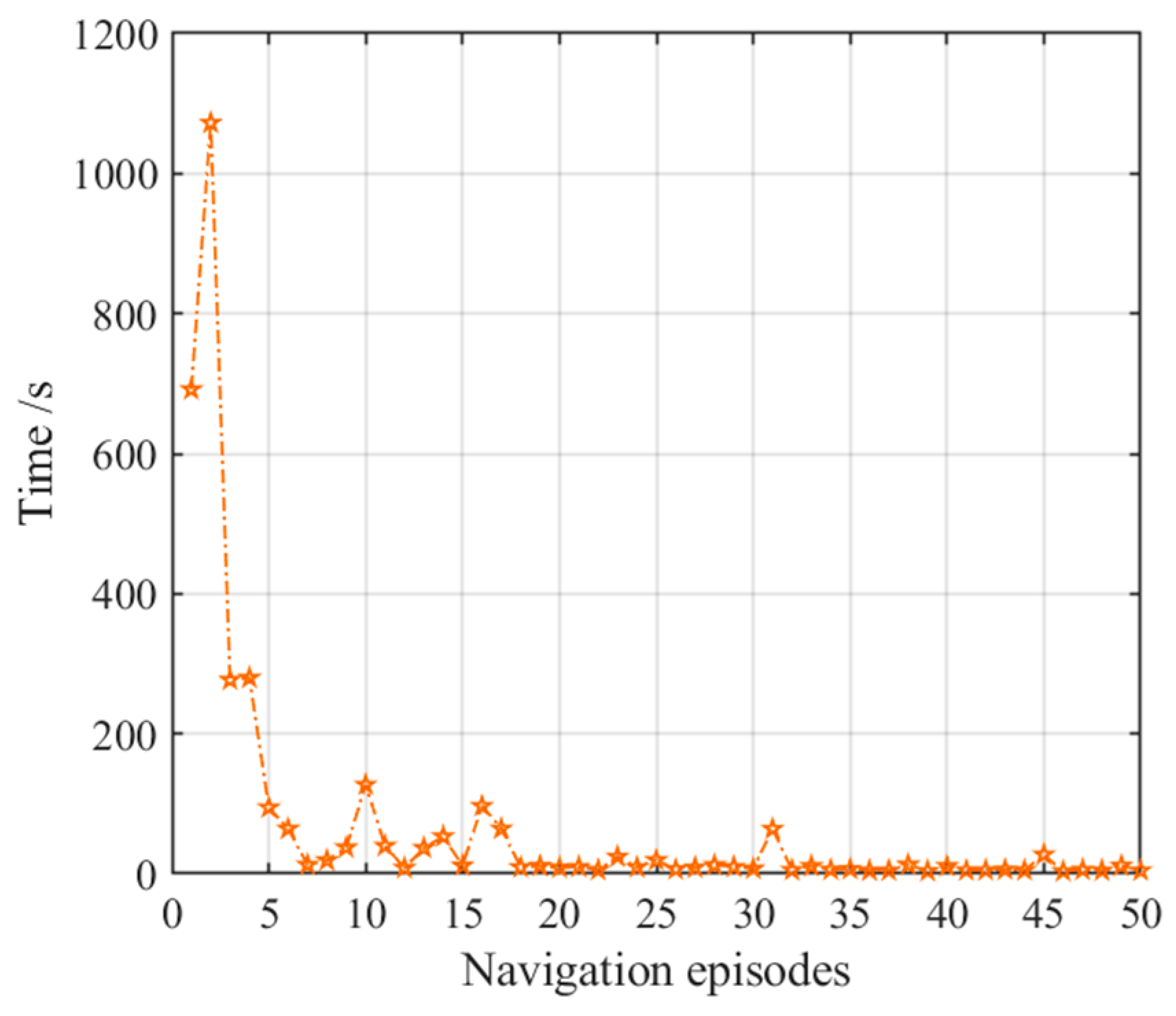
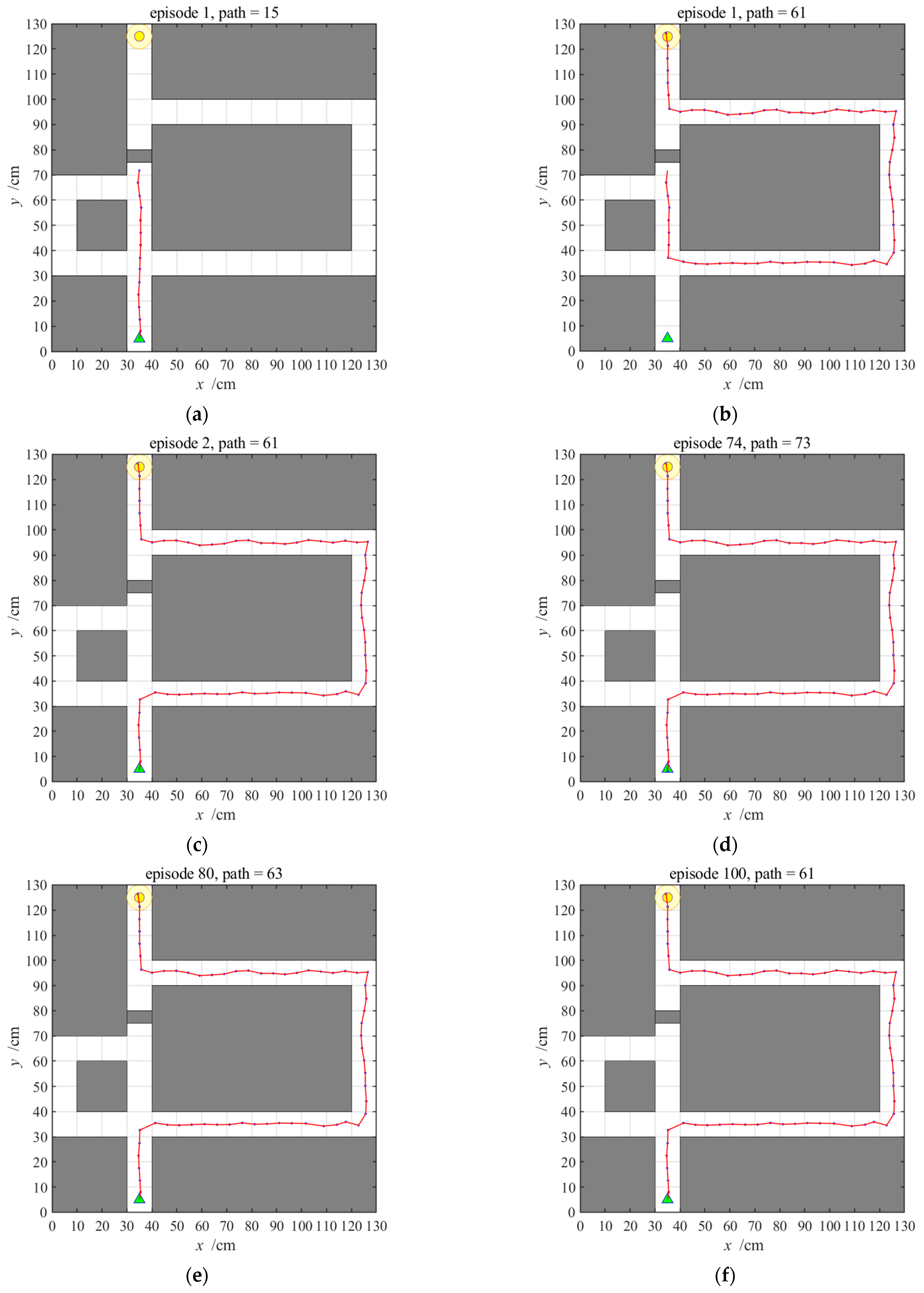
3.2.3. Navigation with the NLM-HS
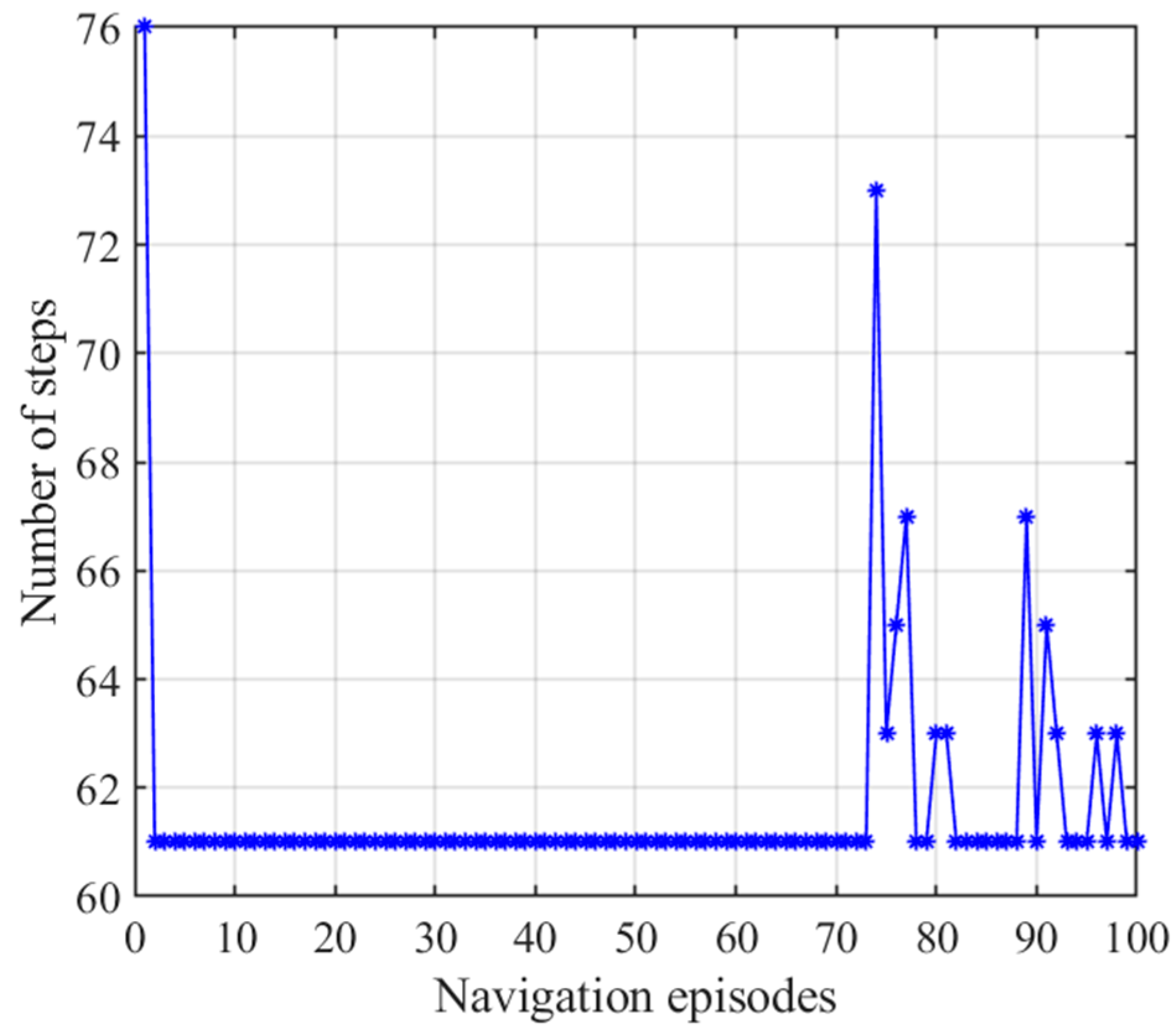
3.3. Adaptive Navigation Experiments
3.4. Results Comparison and Analysis
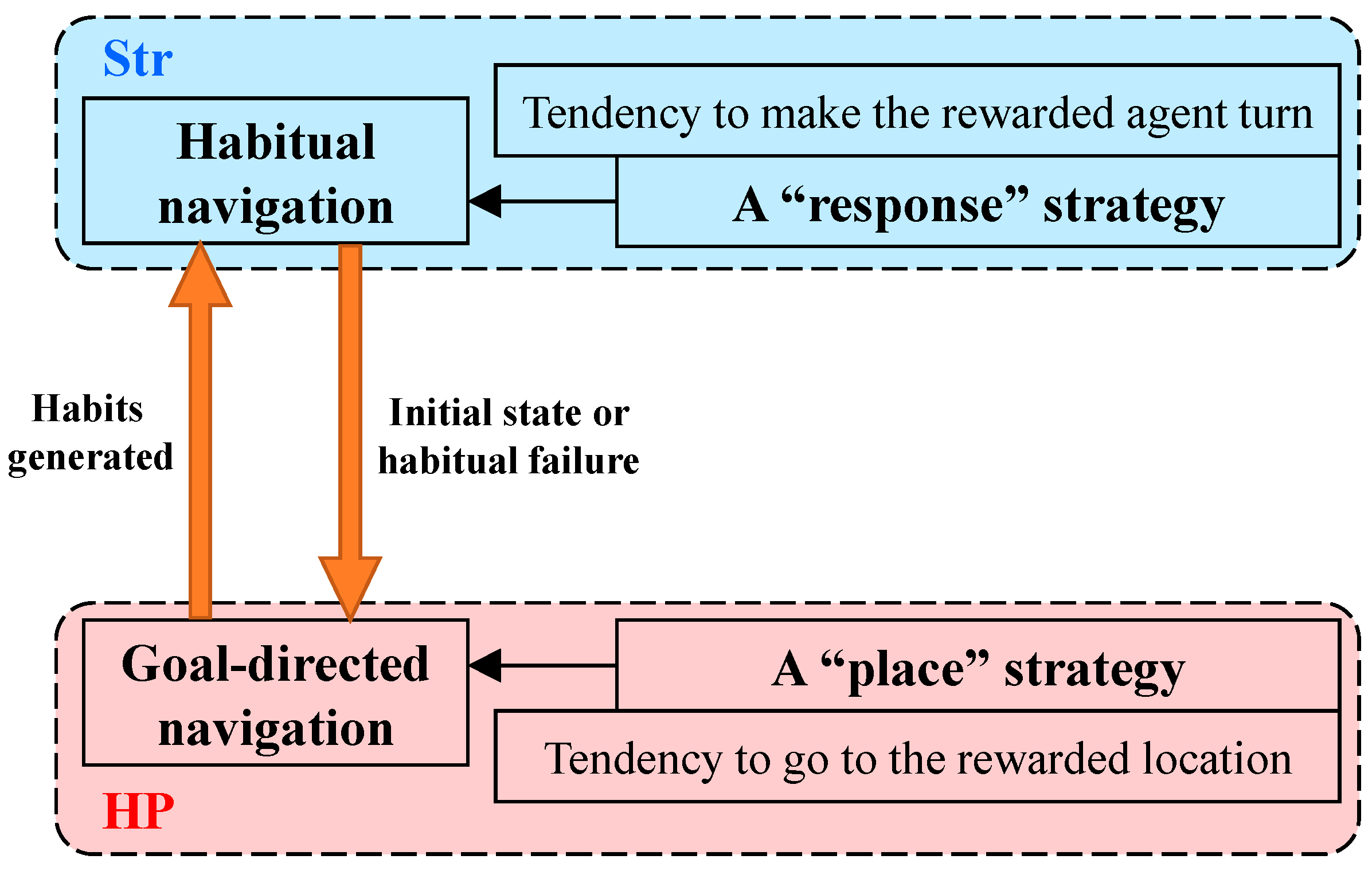
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ni, J.; Wu, L.; Fan, X.; Yang, S.X. Bioinspired intelligent algorithm and its applications for mobile robot control. Comput. Intel. Neurosc. 2016, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Barrera, A.; Weitzenfeld, A. Biologically-inspired Robot Spatial Cognition based on Rat Neurophysiological Studies. Auton. Robot. 2008, 25, 147–169. [Google Scholar] [CrossRef]
- Wyeth, G.; Milford, M. Spatial cognition for robots. IEEE Robot. Autom. Mag. 2009, 16, 24–32. [Google Scholar] [CrossRef]
- Geva-Sagiv, M.; Las, L.; Yovel, Y.; Ulanovsky, N. Spatial cognition in bats and rats: From sensory acquisition to multiscale maps and navigation. Nat. Rev. Neurosci. 2015, 16, 94–108. [Google Scholar] [CrossRef]
- Epstein, R.A.; Patai, E.Z.; Julian, J.B.; Spiers, H.J. The cognitive map in humans: Spatial navigation and beyond. Nat. Neurosci. 2017, 20, 1504–1513. [Google Scholar] [CrossRef]
- Maffeia, G.; Santos-Pataa, D.; Marcosa, E.; Sánchez-Fiblaa, M.; Verschure, P.F.M.J. An embodied biologically constrained model of foraging: From classical and operant conditioning to adaptive real-world behavior in DAC-X. Neural Netw. 2015, 72, 88–108. [Google Scholar] [CrossRef] [PubMed]
- Pennartz, C.M.A.; Ito, R.; Verschure, P.F.M.J.; Battaglia, F.P.; Robbins, T.W. The hippocampal-striatal axis in learning, prediction and goal-directed behavior. Trends Neurosci. 2011, 34, 548–559. [Google Scholar] [CrossRef] [PubMed]
- Chersi, F.; Pezzulo, G. Using hippocampal-striatal loops for spatial navigation and goal-directed decision-making. Cogn. Process. 2012, 13, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Peer, M.; Brunec, I.K.; Newcombe, N.S.; Epstein, R.A. Structuring Knowledge with Cognitive Maps and Cognitive Graphs. Trends Cogn. Sci. 2020, 25, 1–18. [Google Scholar] [CrossRef]
- Tolman, E.C. Cognitive maps in rats and men. Psychol. Rev. 1948, 55, 189–208. [Google Scholar] [CrossRef] [Green Version]
- O’Keefe, J.; Dostrovsky, J. The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 1971, 34, 171–175. [Google Scholar] [CrossRef]
- Mehta, M.R. From synaptic plasticity to spatial maps and sequence learning. Hippocampus 2015, 25, 756–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeiffer, B.E.; Foster, D.J. Hippocampal place-cell sequences depict future paths to remembered goals. Nature 2013, 497, 74–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milford, M. Principles of goal-directed spatial robot navigation in biomimetic models. Philos. Trans. R. Soc. B 2014, 36, 315–318. [Google Scholar] [CrossRef] [Green Version]
- Babayan, B.M.; Watilliaux, A.; Viejo, G.; Paradis, A.L.; Girard, B.; Rondi-Reig, L. A hippocampo-cerebellar centred network for the learning and execution of sequence-based navigation. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef]
- Lansink, C.S.; Goltstein, P.M.; Lankelma, J.V.; McNaughton, B.L.; Pennartz, C.M.A. Hippocampus leads ventral striatum in replay of place-reward information. PLoS Biol. 2009, 7, e1000173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moussa, R.; Poucet, B.; Amalric, M.; Sargolini, F. Contributions of dorsal striatal subregions to spatial alternation behavior. Learn. Mem. 2011, 18, 444–451. [Google Scholar] [CrossRef] [Green Version]
- Ashby, F.G.; Turner, B.O.; Horvitz, J.C. Cortical and basal ganglia contributions to habit learning and automaticity. Trends Cogn. Sci. 2010, 14, 208–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bornstein, A.M.; Daw, N.D. Multiplicity of control in the basal ganglia: Computational roles of striatal subregions. Curr. Opin. Neurobiol. 2011, 21, 374–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maniadakis, M.; Trahanias, P.; Tani, J. Self-organizing high-order cognitive functions in artificial agents: Implications for possible prefrontal cortex mechanisms. Neural Netw. 2012, 33, 76–87. [Google Scholar] [CrossRef]
- Srinivasa, N.; Chelian, S.E. Executive control of cognitive agents using a biologically inspired model architecture of the prefrontal cortex. Biol. Inspir. Cogn. Arc. 2012, 2, 13–24. [Google Scholar] [CrossRef]
- Doeller, C.F.; King, J.A.; Burgess, N. Parallel striatal and hippocampal systems for landmarks and boundaries in spatial memory. Proc. Natl. Acad. Sci. USA 2008, 105, 5915–5920. [Google Scholar] [CrossRef] [Green Version]
- Barraclough, D.J.; Conroy, M.L.; Lee, D. Prefrontal cortex and decision making in a mixed-strategy game. Nat. Neurosci. 2004, 7, 404–410. [Google Scholar] [CrossRef]
- Stachenfeld, K.L.; Botvinick, M.M.; Gershman, S.J. The hippocampus as a predictive map. Nat. Neurosci. 2017, 20, 1643. [Google Scholar] [CrossRef]
- Yu, N.; Zhai, Y.; Yuan, Y.; Wang, Z. A Bionic Robot Navigation Algorithm Based on Cognitive Mechanism of Hippocampus. IEEE Trans. Autom. Sci. Eng. 2019, 16, 1640–1652. [Google Scholar] [CrossRef]
- Zhao, F.; Zeng, Y.; Wang, G.; Bai, J.; Xu, B. A Brain-Inspired Decision Making Model Based on Top-Down Biasing of Prefrontal Cortex to Basal Ganglia and Its Application in Autonomous UAV Explorations. Cogn. Comput. 2018, 10, 296–306. [Google Scholar] [CrossRef]
- McDonald, R.J.; Hong, N.S.; Devan, B.D. The challenges of understanding mammalian cognition and memory-based behaviours: An interactive learning and memory systems approach. Neurosci. Biobehav. Rev. 2005, 28, 719–745. [Google Scholar] [CrossRef]
- Pezzulo, G.; Rigoli, F.; Chersi, F. The Mixed Instrumental Controller: Using Value of Information to Combine Habitual Choice and Mental Simulation. Front. Psychol. 2013, 4, 92–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chersi, F.; Burgess, N. The Cognitive Architecture of Spatial Navigation: Hippocampal and Striatal Contributions. Neuron 2015, 88, 64–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, X.; Chai, J.; Wu, Y.; Zhang, X.; Huang, J. Cognitive map construction and navigation based on hippocampal place cells. Acta Autom. Sin. 2021, 47, 666–677. [Google Scholar] [CrossRef]
- Lee, M.G.; Jun, G.; Choi, H.S.; Jang, H.S.; Bae, Y.C.; Suk, K.; Jang, I.S.; Choi, B.J. Operant conditioning of rat navigation using electrical stimulation for directional cues and rewards. Behav. Process 2010, 84, 715–720. [Google Scholar] [CrossRef]
- Alvernhe, A.; Save, E.; Poucet, B. Local remapping of place cell firing in the Tolman detour task. Eur. J. Neurosci. 2011, 33, 1696–1705. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.I.; Ross, R.S.; Tobyne, S.M.; Stern, C.E. Cooperative interactions between hippocampal and striatal systems support flexible navigation. Neuroimage 2012, 60, 1316–1330. [Google Scholar] [CrossRef] [Green Version]
- Banquet, J.P.; Hanoune, S.; Gaussier, P.; Quoy, M. From Cognitive to Habit Behavior during Navigation, through Cortical-Basal Ganglia Loops. In Artificial Neural Networks and Machine Learning—ICANN 2016; Villa, A., Masulli, P., Pons Rivero, A., Eds.; Lecture Notes in Computer Science; Springer: Cham, Switzerland, 2016; Volume 9886, pp. 238–247. [Google Scholar] [CrossRef]
- Killcross, S.; Coutureau, E. Coordination of Actions and Habits in the Medial Prefrontal Cortex of Rats. Cereb. Cortex 2003, 13, 400–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domenech, P.; Rheims, S.; Koechlin, E. Neural mechanisms resolving exploitation-exploration dilemmas in the medial prefrontal cortex. Science 2020, 369, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Graham, S.; Phua, E.; Soon, C.S.; Oh, T.; Au, C.; Shuter, B.; Wang, S.C.; Yeh, I.B. Role of medial cortical, hippocampal and striatal interactions during cognitive set-shifting. Neuroimage 2009, 45, 1359–1367. [Google Scholar] [CrossRef]
- Dahmani, L.; Bohbot, V.D. Dissociable contributions of the prefrontal cortex to hippocampus- and caudate nucleus-dependent virtual navigation strategies. Neurobiol. Learn. Mem. 2015, 117, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Daw, N.D.; Niv, Y.; Dayan, P. Uncertainty-based competition between prefrontal and dorsolateral striatal systems for behavioral control. Nat. Neurosci. 2005, 8, 1704–1711. [Google Scholar] [CrossRef]
- Seger, C.A.; Spiering, B.J. A Critical Review of Habit Learning and the Basal Ganglia. Front. Syst. Neurosci. 2011, 66. [Google Scholar] [CrossRef] [Green Version]
- Cazin, N.; Alonso, M.L.; Chiodi, P.S.; Pelc, T.; Harland, B.; Weitzenfeld, A.; Fellous, J.M.; Dominey, P.F. Reservoir computing model of prefrontal cortex creates novel combinations of previous navigation sequences from hippocampal place-cell replay with spatial reward propagation. PLoS Comput. Biol. 2019, 15, e1006624. [Google Scholar] [CrossRef] [Green Version]





























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Initial Period | After Behavioral Habit Is Formed | After Environment Is Changed | After New Behavioral Habit Is Formed | |
|---|---|---|---|---|
| 0 | 0 | |||
| >0 | <0 | >0 | <0 | |
| Choose strategy from | HP | Str | HP | Str |
| Line No. | Integrated Navigation Algorithm |
|---|---|
| 1 | Input: agent’s start and goal position, number of iterations , |
| 2 | Output: navigation strategies at each activated place cell. |
| 3 | Initialization: ; growing threshold ; the expected discounted return ; Assign start position to current position. |
| 4 | For navigation episode , loop |
| 5 | (1) Obtain the activated place cell corresponding to the current position; |
| 6 | (2) Choose action. |
| 7 | Agent produces strategy in HP according to a place cell sequence planning algorithm. |
| 8 | Agent produces strategy in Str according to the improved algorithm in the matrix. |
| 9 | Calculate the confidence and . |
| 10 | Calculate the confidence in the prefrontal cortex; |
| 11 | Agent chooses actions according to . |
| 12 | (3) Record or update. |
| 13 | (3.1) update in HP. |
| 14 | if changes in environment are detected, then |
| 15 | update the cognitive map according to the dynamic pruning mechanism; |
| 16 | end if. |
| 17 | Return to (2). |
| 18 | (3.2) Update the expected discounted return in Str. |
| 19 | (4) End judgment. If the goal is reached, then end the navigation episode; else, repeat step (2)~(4). |
| 20 | End loop |
| Models | HP Model | Str Model | NLM-HS |
|---|---|---|---|
| Average steps of first navigation steps | 26 | 517.40 | 26 |
| Average steps of stabilized navigation steps | 26 | 27.76 | 26.32 |
| Average time of first navigation time | 24.54 s | 208.01 s | 25.91 s |
| Average time of stabilized navigation time | 24.54 s | 2.80 s | 1.83 s |
| Average steps of first navigation after door B is closed | 76 | 1378.6 | 76 |
| Average steps of stabilized navigation after door B is closed | 61 | 85.68 | 61.48 |
| Average time of first navigation after door B is closed | 74.99 s | 777.82 s | 64.25 s |
| Average time of stabilized navigation after door B is closed | 60.30 s | 12.76 s | 3.63 s |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chai, J.; Ruan, X.; Huang, J. NLM-HS: Navigation Learning Model Based on a Hippocampal–Striatal Circuit for Explaining Navigation Mechanisms in Animal Brains. Brain Sci. 2021, 11, 803. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11060803
Chai J, Ruan X, Huang J. NLM-HS: Navigation Learning Model Based on a Hippocampal–Striatal Circuit for Explaining Navigation Mechanisms in Animal Brains. Brain Sciences. 2021; 11(6):803. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11060803
Chicago/Turabian StyleChai, Jie, Xiaogang Ruan, and Jing Huang. 2021. "NLM-HS: Navigation Learning Model Based on a Hippocampal–Striatal Circuit for Explaining Navigation Mechanisms in Animal Brains" Brain Sciences 11, no. 6: 803. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11060803