
Success of Hand Movement Imagination Depends on Personality Traits, Brain Asymmetry, and Degree of Handedness

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Measures
2.3. Experimental Procedure
2.3.1. A Session of Control of BCI Based on Kinesthetic Imagination of Hand Movements
2.3.2. Motor Test
2.4. EEG Analysis
2.5. Statistical Analysis
3. Results
3.1. Differences between Accuracy of Classification of Brain Activity during Imagination of RH or LH Movements Depending on Personality Traits without Taking into Account Latent Left-Handedness
3.2. Differences between Accuracy of Classification of Brain Activity during Imagination of RH or LH Movements Depending on Personality Traits Taking into Account Latent Left-Handedness
3.3. Subjective Complexity of Real and Imaging Movements Correlated with the Accuracy of Classification of the Brain States
4. Discussion
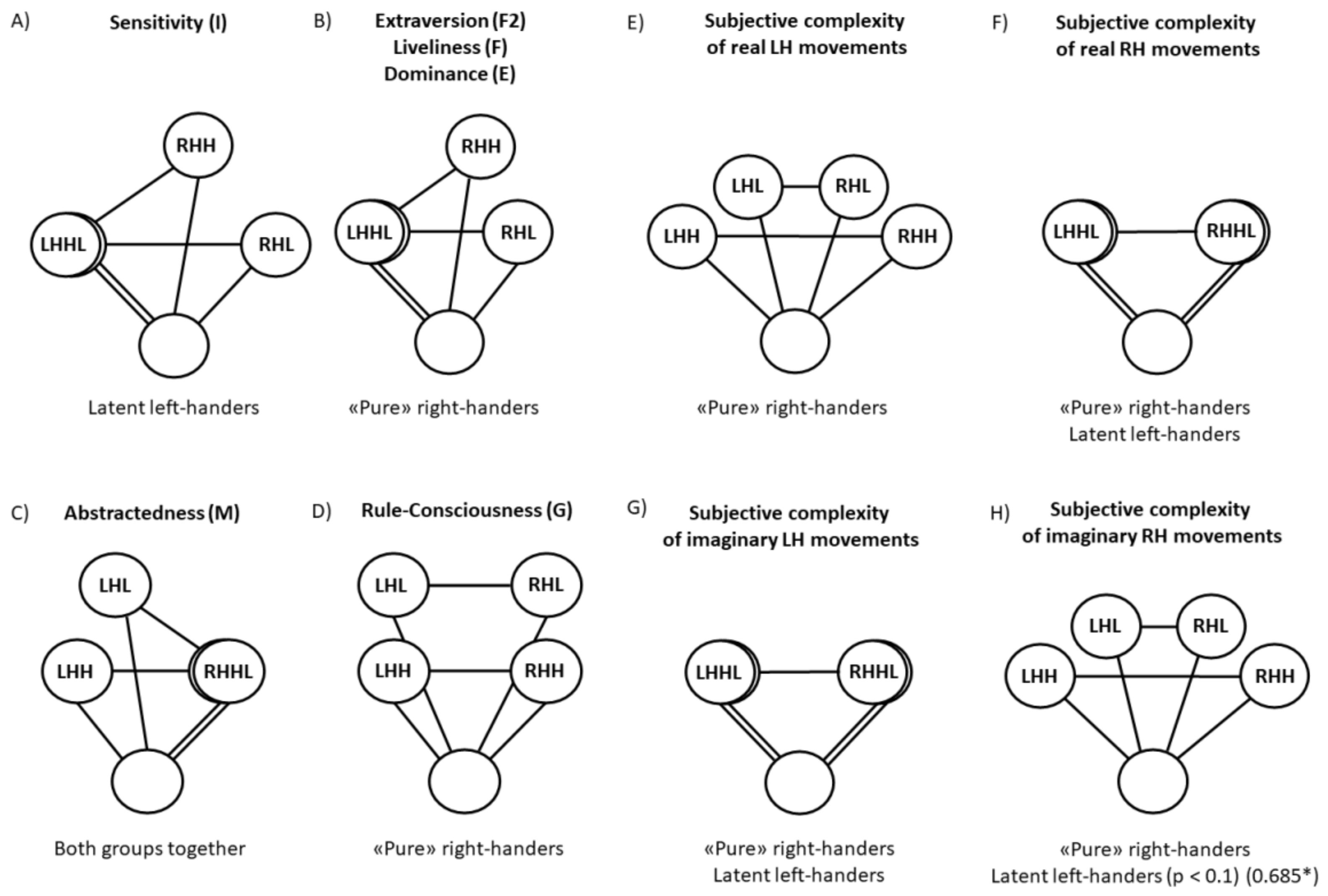
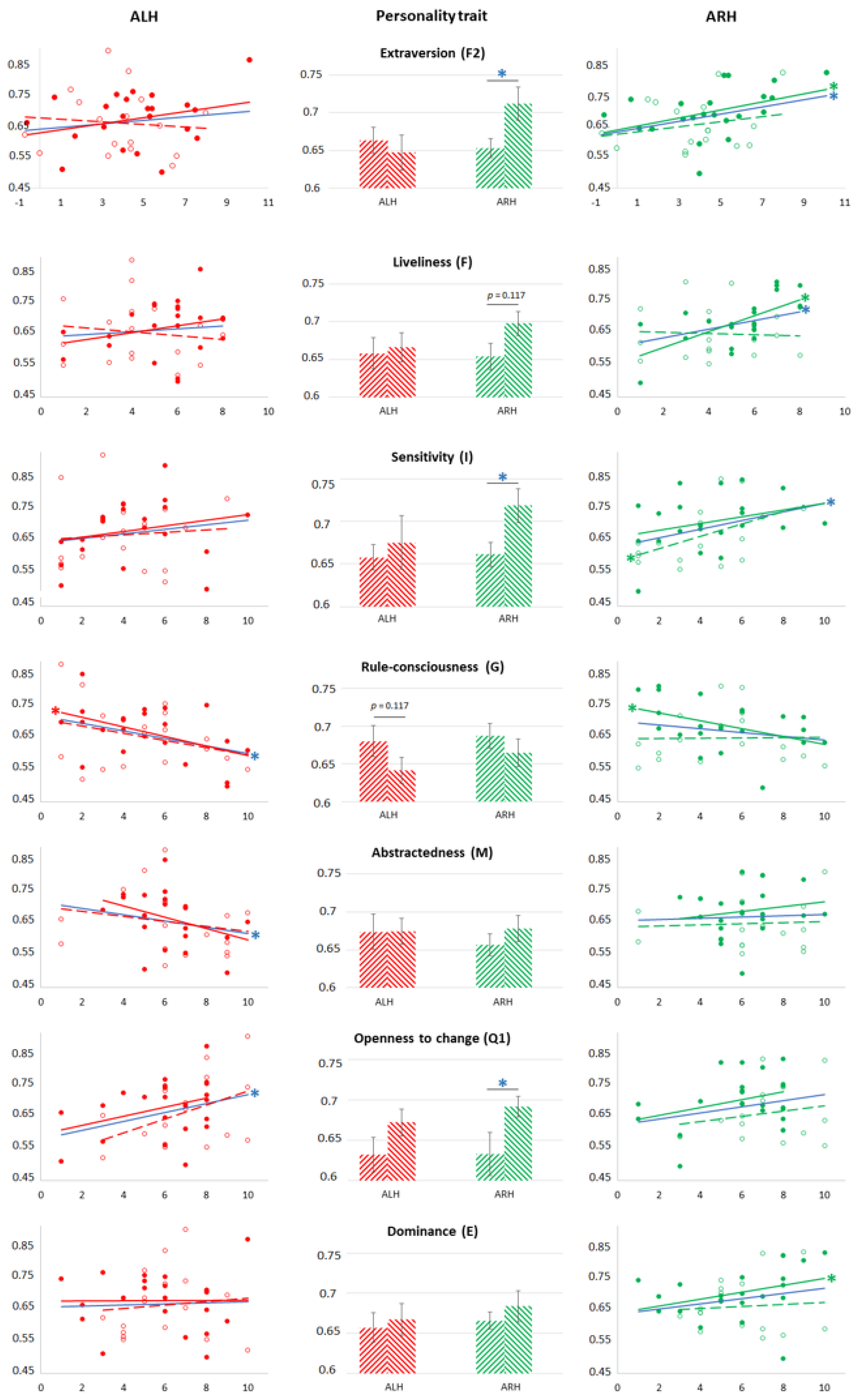
4.1. Why Are Lively Dominant Extroverts Higher in ARH?
4.2. Why Does the Factor Sensitivity (I) in Latent Left-Handers (But Not in “Pure” Right-Handers) Affect ARH (Not ALH)?
4.3. Why Does Factor Abstractedness (M) Affect ALH?
4.4. What Is the Reason for the Differences in the Subjective Complexity of Realizing Real Movements of the RH and LH and Their Imagination?
4.5. How Can We Imagine the Relationship between the Results Described above?
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- Buch, E.; Weber, C.; Cohen, L.G.; Braun, C.; Dimyan, M.A.; Ard, T.; Mellinger, J.; Caria, A.; Soekadar, S.; Fourkas, A.; et al. Think to Move: A Neuromagnetic Brain-Computer Interface (BCI) System for Chronic Stroke. Stroke 2008, 39, 910–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, G.; Herman, P.; Coyle, D.; McDonough, S.; Crosbie, J. Applying a Brain-Computer Interface to Support Motor Imagery Practice in People with Stroke for Upper Limb Recovery: A Feasibility Study. J. Neuroeng. Rehabil. 2010, 7, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mokienko, O.A.; Chervyakov, A.V.; Kulikova, S.N.; Bobrov, P.D.; Chernikova, L.A.; Frolov, A.A.; Piradov, M.A. Increased motor cortex excitability during motor imagery in brain-computer interface trained subjects. Front. Comput. Neurosci. 2013, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebedev, M. Brain-Machine Interfaces: An Overview. Transl. Neurosci. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Lebedev, M.A.; Nicolelis, M.A.L. Brain-Machine Interfaces: From Basic Science to Neuroprostheses and Neurorehabilitation. Physiol. Rev. 2017, 97, 767–837. [Google Scholar] [CrossRef] [PubMed]
- Soekadar, S.R.; Birbaumer, N.; Slutzky, M.W.; Cohen, L.G. Brain–machine interfaces in neurorehabilitation of stroke. Neurobiol. Dis. 2015, 83, 172–179. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.; Kim, S.; Lee, B. Effects of Action Observational Training Plus Brain-Computer Interface-Based Functional Electrical Stimulation on Paretic Arm Motor Recovery in Patient with Stroke: A Randomized Controlled. Trial. Occup. Ther. Int. 2016, 23, 39–47. [Google Scholar] [CrossRef]
- Donati, A.R.C.; Shokur, S.; Morya, E.; Campos, D.S.F.; Moioli, R.C.; Gitti, C.M.; Augusto, P.B.; Tripodi, S.; Pires, C.G.; Pereira, G.A.; et al. Long-Term Training with a Brain-Machine Interface-Based Gait Protocol Induces Partial Neurological Recovery in Paraplegic Patients. Sci. Rep. 2016, 6, 30383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frolov, A.A.; Mokienko, O.; Lyukmanov, R.; Biryukova, E.; Kotov, S.; Turbina, L.; Nadareyshvily, G.; Bushkova, Y. Post-stroke Rehabilitation Training with a Motor-Imagery-Based Brain-Computer Interface (BCI)-Controlled Hand Exoskeleton: A Randomized Controlled Multicenter Trial. Front. Neurosci. 2017, 11. [Google Scholar] [CrossRef] [Green Version]
- Frolov, A.A.; Húsek, D.; Biryukova, E.V.; Bobrov, P.D.; Mokienko, O.A.; Alexandrov, A.V. Principles of Motor Recovery in Post-Stroke Patients Using Hand Exoskeleton Controlled by the Brain-Computer Interface Based on Motor Imagery. Neural Netw. World 2017, 27, 107–137. [Google Scholar] [CrossRef] [Green Version]
- Frolov, A.A.; Bobrov, P.D.; Biryukova, E.V.; Silchenko, A.V.; Kondur, A.A.; Dzhalagoniya, I.Z.; Massion, J. Electrical, Hemodynamic, and Motor Activity in BCI Post-stroke Rehabilitation: Clinical Case Study. Front. Neurol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Bobrova, E.V.; Frolov, A.A.; Reshetnikova, V.V. Methods and Approaches to Optimizing Control Using a Brain–Computer Interface System by Healthy Subjects and Patients with Motor Disorders. Neurosci. Behav. Physiol. 2018, 48, 1041–1052. [Google Scholar] [CrossRef]
- Jeunet, C.; Glize, B.; McGonigal, A.; Batail, J.-M.; Micoulaud-Franchi, J.-A. Using EEG-based brain computer interface and neurofeedback targeting sensorimotor rhythms to improve motor skills: Theoretical background, applications and prospects. Neurophysiol. Clin. 2019, 49, 125–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, E.; Lee, B.-H.; Hwang, S. Therapeutic effects of brain-computer interface-controlled functional electrical stimulation training on balance and gait performance for stroke. Medicine 2020, 99, e22612. [Google Scholar] [CrossRef]
- Bobrova, E.V.; Reshetnikova, V.V.; Frolov, A.A.; Gerasimenko, Y.P. Use of Imaginary Lower Limb Movements to Control Brain–Computer Interface Systems. Neurosci. Behav. Physiol. 2020, 50, 585–592. [Google Scholar] [CrossRef]
- Khan, M.A.; Das, R.; Iversen, H.K.; Puthusserypady, S. Review on motor imagery based BCI systems for upper limb post-stroke neurorehabilitation: From designing to application. Comput. Biol. Med. 2020, 123, 103843. [Google Scholar] [CrossRef]
- Baniqued, P.D.E.; Stanyer, E.C.; Awais, M.; Alazmani, A.; Jackson, A.E.; Mon-Williams, M.A.; Mushtaq, F.; Holt, R.J. Brain–computer interface robotics for hand rehabilitation after stroke: A systematic review. J. Neuroeng. Rehabil. 2021, 18, 15. [Google Scholar] [CrossRef] [PubMed]
- Paszkiel, S.; Dobrakowski, P. Brain–Computer Technology-Based Training System in the Field of Motor Imagery. IET Sci. Meas. Technol. 2020, 14, 1014–1018. [Google Scholar] [CrossRef]
- King, C.E.; Wang, P.T.; Chui, L.A.; Do, A.H.; Nenadic, Z. Operation of a brain-computer interface walking simulator for individuals with spinal cord injury. J. Neuroeng. Rehabil. 2013, 10, 77. [Google Scholar] [CrossRef] [Green Version]
- Paszkiel, S. Augmented Reality of Technological Environment in Correlation with Brain Computer Interfaces for Control Processes. Recent Advances in Automation, Robotics and Measuring Techniques. In Advances in Intelligent Systems and Computing; Szewczyk, R., Zieliński, C., Kaliczyńska, M., Eds.; Springer: Cham, Switzerland, 2014; pp. 197–203. [Google Scholar] [CrossRef]
- Jeunet, C.; N’Kaoua, B.; Subramanian, S.; Hachet, M.; Lotte, F. Predicting Mental Imagery-Based BCI Performance from Personality, Cognitive Profile and Neurophysiological Patterns. PLoS ONE 2015, 10, e0143962. [Google Scholar] [CrossRef] [PubMed]
- Blankertz, B.; Sannelli, C.; Halder, S.; Hammer, E.M.; Kübler, A.; Müller, K.-R.; Curio, G.; Dickhaus, T. Neurophysiological predictor of SMR-based BCI performance. Neuroimage 2010, 51, 1303–1309. [Google Scholar] [CrossRef] [Green Version]
- Sannelli, C.; Vidaurre, C.; Müller, K.-R.; Blankertz, B. A large scale screening study with a SMR-based BCI: Categorization of BCI users and differences in their SMR activity. PLoS ONE 2019, 14, e0207351. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Xu, P.; Chen, R.; Li, F.; Guo, L.; Li, P.; Zhang, T.; Yao, D. Predicting Inter-session Performance of SMR-Based Brain–Computer Interface Using the Spectral Entropy of Resting-State EEG. Brain Topogr. 2015, 28, 680–690. [Google Scholar] [CrossRef]
- Ahn, M.; Ahn, S.; Hong, J.H.; Cho, H.; Kim, K.; Kim, B.S.; Chang, J.W.; Jun, S.C. Gamma band activity associated with BCI performance: Simultaneous MEG/EEG study. Front. Hum. Neurosci. 2013, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bamdadian, A.; Guan, C.; Ang, K.K.; Xu, J. The predictive role of pre-cue EEG rhythms on MI-based BCI classification performance. J. Neurosci. Methods 2014, 235, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Grosse-Wentrup, M.; Schölkopf, B. High gamma-power predicts performance in sensorimotor-rhythm brain–computer interfaces. J. Neural Eng. 2012, 9, 046001. [Google Scholar] [CrossRef] [PubMed]
- Jeunet, C.; N’Kaoua, B.; Lotte, F. Advances in user-training for mental-imagery-based BCI control. Prog. Brain Res. 2016, 228, 3–35. [Google Scholar] [PubMed]
- Hammer, E.M.; Kaufmann, T.; Kleih, S.C.; Blankertz, B.; Kübler, A. Visuo-motor coordination ability predicts performance with brain-computer interfaces controlled by modulation of sensorimotor rhythms (SMR). Front. Hum. Neurosci. 2014, 8. [Google Scholar] [CrossRef] [Green Version]
- Daum, I.; Rockstroh, B.; Birbaumer, N.; Elbert, T.; Canavan, A.; Lutzenberger, W. Behavioural treatment of slow cortical potentials in intractable epilepsy: Neuropsychological predictors of outcome. J. Neurol. Neurosurg. Psychiatry 1993, 56, 94–97. [Google Scholar] [CrossRef] [Green Version]
- Grosse-Wentrup, M.; Schölkopf, B.; Hill, J. Causal influence of gamma oscillations on the sensorimotor rhythm. Neuroimage 2011, 56, 837–842. [Google Scholar] [CrossRef]
- Hammer, E.M.; Halder, S.; Blankertz, B.; Sannelli, C.; Dickhaus, T.; Kleih, S.; Müller, K.-R.; Kübler, A. Psychological predictors of SMR-BCI performance. Biol. Psychol. 2012, 89, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Neumann, N. Predictors of successful self control during brain-computer communication. J. Neurol. Neurosurg. Psychiatry 2003, 74, 1117–1121. [Google Scholar] [CrossRef] [Green Version]
- Burde, W.; Blankertz, B. Is the locus of control of reinforcement a predictor of brain-computer interface performance? In Proceedings of the 3rd International Brain-Computer Interface Workshop and Training Course, Graz, Austria, 21–24 September 2006; Muller-Putz, G.R., Brunner, R., Leeb, R., Scherer, R., Shclogl, A., Wriessnegger, S., Pfurtscheller, G., Eds.; TU Graz: Graz, Austria, 2006; pp. 108–109. [Google Scholar]
- Nijboer, F. The influence of psychological state and motivation on brain-computer interface performance in patients with amyotrophic lateral sclerosis—A longitudinal study. Front. Neurosci. 2010. [Google Scholar] [CrossRef] [Green Version]
- Witte, M.; Kober, S.E.; Ninaus, M.; Neuper, C.; Wood, G. Control beliefs can predict the ability to up-regulate sensorimotor rhythm during neurofeedback training. Front. Hum. Neurosci. 2013, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleih, S.C.; Kaufmann, T.; Hammer, E.M.; Pisotta, I.; Picchiori, F.; Riccio, A.; Mattia, D.; Kubler, A. Motivation and SMR-BCI: Fear of Failure Affects BCI Performance. In Proceedings of the 5th International Brain-Computer Interface Meeting: Defining the Furture, Pacific Grove, CA, USA, 3–7 June 2013; Millan, J.d.R., Gao, S., Muller-Putz, G., Wolpaw, J.R., Huggins, J.E., Eds.; Graz University of Technology Publishing House: Graz, Austria, 2013; pp. 160–161. [Google Scholar]
- Felder, R.M.; Spurlin, J. Applications, reliability and validity of the Index of Learning Styles. Int. J. Eng. Educ. 2005, 21, 103–112. [Google Scholar]
- Pacheco, K.; Acuna, K.; Carranza, E.; Achanccaray, D.; Andreu-Perez, J. Performance predictors of motor imagery brain-computer interface based on spatial abilities for upper limb rehabilitation. In Proceedings of the 2017 39th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC) IEEE, Jeju, Korea, 11–15 July 2017; pp. 1014–1017. [Google Scholar]
- Vuckovic, A.; Osuagwu, B.A. Using a motor imagery questionnaire to estimate the performance of a Brain–Computer Interface based on object oriented motor imagery. Clin. Neurophysiol. 2013, 124, 1586–1595. [Google Scholar] [CrossRef] [Green Version]
- Nijboer, F.; Furdea, A.; Gunst, I.; Mellinger, J.; McFarland, D.J.; Birbaumer, N.; Kübler, A. An auditory brain–computer interface (BCI). J. Neurosci. Methods 2008, 167, 43–50. [Google Scholar] [CrossRef]
- Randolph, A.B. Not All Created Equal: Individual-Technology Fit of Brain-Computer Interfaces. In Proceedings of the 2012 45th Hawaii International Conference on System Sciences IEEE, Maui, HI, USA, 4–7 January 2012; pp. 572–578. [Google Scholar]
- Schmidtke, J.I.; Heller, W. Personality, affect and EEG: Predicting patterns of regional brain activity related to extraversion and neuroticism. Pers. Individ. Dif. 2004, 36, 717–732. [Google Scholar] [CrossRef]
- Minnix, J.A.; Kline, J.P. Neuroticism predicts resting frontal EEG asymmetry variability. Pers. Individ. Dif. 2004, 36, 823–832. [Google Scholar] [CrossRef]
- Gale, A.; Coles, M.; Kline, P.; Penfold, V. Extraversion-introversion, neuroticism and the EEG: Basal and response measures during habituation of the orienting response. Br. J. Psychol. 1971, 62, 533–543. [Google Scholar] [CrossRef]
- McCrae, R.R.; John, O.P. An Introduction to the Five-Factor Model and Its Applications. J. Pers. 1992, 60, 175–215. [Google Scholar] [CrossRef] [PubMed]
- McCrae, R.R.; Costa, P.T. The five-factor theory of personality. In Handbook of Personality: Theory and Research; John, O.P., Robins, R.W., Pervin, L.A., Eds.; The Guilford Press: New York, NY, USA, 2008; pp. 159–181. [Google Scholar]
- Nazre, A.R.; Mohd, N.T.; Sahrim, L.; Norizam, S.; Zunairah, H.M. Learners’ learning style correlated to agreeableness based on EEG. In Proceedings of the International Conference on Management and Education Innovation, Kuala Lumpur, Malasia, 5 May 2012; ACSIT Press: Singapore, 2012; pp. 123–127. [Google Scholar]
- Stough, C.; Donaldson, C.; Scarlata, B.; Ciorciari, J. Psychophysiological correlates of the NEO PI-R Openness, Agreeableness and Conscientiousness: Preliminary results. Int. J. Psychophysiol. 2001, 41, 87–91. [Google Scholar] [CrossRef]
- Stenberg, G. Personality and the EEG: Arousal and emotional arousability. Pers. Individ. Dif. 1992, 13, 1097–1113. [Google Scholar] [CrossRef] [Green Version]
- Pavlenko, V.B.; Chernyi, S.V.; Goubkina, D.G. EEG Correlates of Anxiety and Emotional Stability in Adult Healthy Subjects. Neurophysiology 2009, 41, 337–345. [Google Scholar] [CrossRef]
- Bobrova, E.V.; Reshetnikova, V.V.; Vershinina, E.A.; Grishin, A.A.; Frolov, A.A.; Gerasimenko, Y.P. Interhemispheric Asymmetry and Personality Traits of Brain–Computer Interface Users in Hand Movement Imagination. Dokl. Biol. Sci. 2020, 495, 265–267. [Google Scholar] [CrossRef]
- Sartarelli, M. Handedness, Earnings, Ability and Personality. Evidence from the Lab. PLoS ONE 2016, 11, e0164412. [Google Scholar] [CrossRef]
- Mascie-Taylor, C.G.N. Hand Preference and Personality Traits. Cortex 1981, 17, 319–322. [Google Scholar] [CrossRef]
- Coren, S. Personality Differences between Left- and Right-Handers: An Overlooked Minority Group? J. Res. Pers. 1994, 28, 214–229. [Google Scholar] [CrossRef]
- Lester, D. The relationship between handedness and personality traits (extraversion and neuroticism). Pers. Individ. Dif. 1987, 8, 437. [Google Scholar] [CrossRef]
- Grimshaw, G.M.; Wilson, M.S. A sinister plot? Facts, beliefs, and stereotypes about the left-handed personality. Laterality Asymmetries Body Brain Cogn. 2013, 18, 135–151. [Google Scholar] [CrossRef] [PubMed]
- Christman, S.D. Encyclopedia of Human Behavior, 2nd ed.; Ramachandran, V.S., Ed.; Academic Press: London, UK, 2012; ISBN 978-0-08-096180-4. [Google Scholar]
- Luria, A.R.; Critchley, M. Traumatic Aphasia; De Gruyter Mouton: The Hague, The Netherlands, 1970; ISBN 9789027907172. [Google Scholar]
- Tran, U.S.; Stieger, S.; Voracek, M. Handedness and sex roles: Mixed-handers are less sex-congruent stereotyped. Pers. Individ. Dif. 2014, 66, 10–13. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Bragina, N.N.; Dobrohotova, T.A. Human Functional Asymmetries, 2nd ed.; Makhotin, Y.V., Ed.; Medicine: Moscow, Russia, 1998. [Google Scholar]
- Goldberg, E.; Costa, L.D. Hemisphere differences in the acquisition and use of descriptive systems. Brain Lang. 1981, 14, 144–173. [Google Scholar] [CrossRef]
- Heilman, K.M.; Abell, T.V.D. Right hemisphere dominance for attention: The mechanism underlying hemispheric asymmetries of inattention (neglect). Neurology 1980, 30, 327. [Google Scholar] [CrossRef]
- Geschwind, N. Cerebral Lateralization. Arch. Neurol. 1985, 42, 428. [Google Scholar] [CrossRef] [PubMed]
- Hammond, G. Correlates of human handedness in primary motor cortex: A review and hypothesis. Neurosci. Biobehav. Rev. 2002, 26, 285–292. [Google Scholar] [CrossRef]
- Davidson, R.J.; Hugdahl, K. The Asymmetrical Brain, 1st ed.; The MIT Press: Cambridge, MA, USA, 2003. [Google Scholar]
- Toga, A.; Thompson, P. Mapping brain asymmetry. Nat. Rev. Neurosci. 2003, 4, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Hatta, T. Handedness and the Brain: A Review of Brain-imaging Techniques. Magn. Reson. Med. Sci. 2007, 6, 99–112. [Google Scholar] [CrossRef] [Green Version]
- Ocklenburg, S.; Güntürkün, O. Handedness and Other Behavioral Asymmetries. In The Lateralized Brain; Elsevier: Amsterdam, The Netherlands, 2018; pp. 123–158. [Google Scholar]
- Harrington, D.L.; Haaland, K.Y. Hemispheric specialization for motor sequencing: Abnormalities in levels of programming. Neuropsychologia 1991, 29, 147–163. [Google Scholar] [CrossRef]
- Haaland, K.Y.; Harrington, D.L. Hemispheric asymmetry of movement. Curr. Opin. Neurobiol. 1996, 6, 796–800. [Google Scholar] [CrossRef]
- Haaland, K.Y. Hemispheric asymmetries for kinematic and positional aspects of reaching. Brain 2004, 127, 1145–1158. [Google Scholar] [CrossRef] [Green Version]
- Sainburg, R.L.; Kalakanis, D. Differences in Control of Limb Dynamics During Dominant and Nondominant Arm Reaching. J. Neurophysiol. 2000, 83, 2661–2675. [Google Scholar] [CrossRef] [Green Version]
- Sainburg, R. Evidence for a dynamic-dominance hypothesis of handedness. Exp. Brain Res. 2002, 142, 241–258. [Google Scholar] [CrossRef] [PubMed]
- Sainburg, R.L. Handedness: Differential Specializations for Control of Trajectory and Position. Exerc. Sport Sci. Rev. 2005, 33, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Sainburg, R.L.; Duff, S.V. Does motor lateralization have implications for stroke rehabilitation? J. Rehabil. Res. Dev. 2006, 43, 311. [Google Scholar] [CrossRef]
- Duff, S.V.; Sainburg, R.L. Lateralization of motor adaptation reveals independence in control of trajectory and steady-state position. Exp. Brain Res. 2007, 179, 551–561. [Google Scholar] [CrossRef]
- Przybyla, A.; Good, D.C.; Sainburg, R.L. Dynamic dominance varies with handedness: Reduced interlimb asymmetries in left-handers. Exp. Brain Res. 2012, 216, 419–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutha, P.K.; Haaland, K.Y.; Sainburg, R.L. The Effects of Brain Lateralization on Motor Control and Adaptation. J. Mot. Behav. 2012, 44, 455–469. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.; Mutha, P.K.; Przybyla, A.; Haaland, K.Y.; Good, D.C.; Sainburg, R.L. Contralesional motor deficits after unilateral stroke reflect hemisphere-specific control mechanisms. Brain 2013, 136, 1288–1303. [Google Scholar] [CrossRef] [Green Version]
- Mutha, P.K.; Sainburg, R.L.; Haaland, K.Y. Critical neural substrates for correcting unexpected trajectory errors and learning from them. Brain 2011, 134, 3647–3661. [Google Scholar] [CrossRef] [PubMed]
- Bobrova, E.V.; Lyakhovetskii, V.A.; Bogacheva, I.N. Learning and Reproduction of Memorized Sequences of Right and Left Hand Movements. Neurosci. Behav. Physiol. 2016, 46, 762–769. [Google Scholar] [CrossRef]
- Bobrova, E.V.; Lyakhovetskii, V.A.; Borshchevskaya, E.R. The Role of “Prehistory” in the Reproduction of Sequential Movements of the Right and Left Hands: Encoding of Positions, Movements, and Sequence Structure. Neurosci. Behav. Physiol. 2013, 43, 56–62. [Google Scholar] [CrossRef]
- Lyakhovetskii, V.A.; Bobrova, E.V. Age-Related Changes in Memorizing Sequences of Movements with the Dominant and Subdominant Hands. Neurosci. Behav. Physiol. 2015, 45, 74–77. [Google Scholar] [CrossRef]
- Bobrova, E.V.; Bogacheva, I.N.; Lyakhovetskii, V.A.; Fabinskaja, A.A.; Fomina, E.V. Memorization of sequences of movements of the right and left hand by right- and left-handers. Hum. Physiol. 2015, 41, 629–635. [Google Scholar] [CrossRef]
- Bobrova, E.V.; Bogacheva, I.N.; Lyakhovetskii, V.A.; Fabinskaja, A.A.; Fomina, E.V. Memorization of Sequences of Movements of the Right or the Left Hand by Right- and Left-Handers: Vector Coding. Hum. Physiol. 2017, 43, 13–21. [Google Scholar] [CrossRef]
- Witelson, S.F. Hand and sex differences in the isthmus and genu of the human corpus callosum. Brain 1989, 112, 799–835. [Google Scholar] [CrossRef] [PubMed]
- Witelson, S.F.; Goldsmith, C.H. The relationship of hand preference to anatomy of the corpus callosum in men. Brain Res. 1991, 545, 175–182. [Google Scholar] [CrossRef]
- Witelson, S. The brain connection: The corpus callosum is larger in left-handers. Science 1985, 229, 665–668. [Google Scholar] [CrossRef]
- Leong, W.J.; Andrew Ng, C.R.A. Left-handedness detection. Int. J. Smart Sens. Intell. Syst. 2014, 7, 442–457. [Google Scholar] [CrossRef] [Green Version]
- Steinmetz, H.; Volkmann, J.; Jäncke, L.; Freund, H.-J. Anatomical left-right asymmetry of language-related temporal cortex is different in left- and right-handers. Ann. Neurol. 1991, 29, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Snyder, P. Cerebellar volume asymmetries are related to handedness: A quantitative MRI study. Neuropsychologia 1995, 33, 407–419. [Google Scholar] [CrossRef]
- Zetzsche, T.; Meisenzahl, E.M.; Preuss, U.W.; Holder, J.J.; Kathmann, N.; Leinsinger, G.; Hahn, K.; Hegerl, U.; Möller, H.-J. In-vivo analysis of the human planum temporale (PT): Does the definition of PT borders influence the results with regard to cerebral asymmetry and correlation with handedness? Psychiatry Res. Neuroimaging 2001, 107, 99–115. [Google Scholar] [CrossRef]
- Zhavoronkova, L.A. Dextrals and sinistrals (right-handers and left-handers): Specificity of interhemispheric brain asymmetry and EEG coherence parameters. Zhurnal Vysshei Nervnoi Deiatelnosti Imeni I P Pavlova 2007, 57, 645–662. [Google Scholar]
- Volkmann, J.; Schnitzler, A.; Witte, O.W.; Freund, H.-J. Handedness and Asymmetry of Hand Representation in Human Motor Cortex. J. Neurophysiol. 1998, 79, 2149–2154. [Google Scholar] [CrossRef] [Green Version]
- Grabowska, A.; Gut, M.; Binder, M.; Forsberg, L.; Rymarczyk, K.; Urbanik, A. Switching handedness: fMRI study of hand motor control in right-handers, left-handers and converted left-handers. Acta Neurobiol. Exp. 2012, 72, 439–451. [Google Scholar]
- Van den Berg, F.E.; Swinnen, S.P.; Wenderoth, N. Involvement of the Primary Motor Cortex in Controlling Movements Executed with the Ipsilateral Hand Differs between Left- and Right-handers. J. Cogn. Neurosci. 2011, 23, 3456–3469. [Google Scholar] [CrossRef] [PubMed]
- Goble, D.J.; Noble, B.C.; Brown, S.H. Proprioceptive target matching asymmetries in left-handed individuals. Exp. Brain Res. 2009, 197, 403–408. [Google Scholar] [CrossRef]
- Dassonville, P.; Zhu, X.-H.; Ugurbil, K.; Kim, S.-G.; Ashe, J. Functional activation in motor cortex reflects the direction and the degree of handedness. Proc. Natl. Acad. Sci. USA 1997, 94, 14015–14018. [Google Scholar] [CrossRef] [Green Version]
- Pool, E.-M.; Rehme, A.K.; Fink, G.R.; Eickhoff, S.B.; Grefkes, C. Handedness and effective connectivity of the motor system. Neuroimage 2014, 99, 451–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flowers, K. Handedness and controlled movement. Br. J. Psychol. 1975, 66, 39–52. [Google Scholar] [CrossRef]
- Verstynen, T.; Diedrichsen, J.; Albert, N.; Aparicio, P.; Ivry, R.B. Ipsilateral Motor Cortex Activity During Unimanual Hand Movements Relates to Task Complexity. J. Neurophysiol. 2005, 93, 1209–1222. [Google Scholar] [CrossRef]
- Adamo, D.E.; Martin, B.J. Position sense asymmetry. Exp. Brain Res. 2009, 192, 87–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamo, D.E.; Scotland, S.; Martin, B.J. Upper limb kinesthetic asymmetries: Gender and handedness effects. Neurosci. Lett. 2012, 516, 188–192. [Google Scholar] [CrossRef]
- Solodkin, A.; Hlustik, P.; Noll, D.C.; Small, S.L. Lateralization of motor circuits and handedness during finger movements. Eur. J. Neurol. 2001, 8, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Willems, R.M.; Van der Haegen, L.; Fisher, S.E.; Francks, C. On the other hand: Including left-handers in cognitive neuroscience and neurogenetics. Nat. Rev. Neurosci. 2014, 15, 193–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohr, C.; Landis, T.; Bracha, H.S.; Brugger, P. Opposite Turning Behavior in Right-Handers and Non-Right-Handers Suggests a Link Between Handedness and Cerebral Dopamine Asymmetries. Behav. Neurosci. 2003, 117, 1448–1452. [Google Scholar] [CrossRef] [Green Version]
- Glick, S.D.; Ross, D.A.; Hough, L.B. Lateral asymmetry of neurotransmitters in human brain. Brain Res. 1982, 234, 53–63. [Google Scholar] [CrossRef]
- Van Dyck, C.H.; Seibyl, J.P.; Malison, R.T.; Laruelle, M.; Zoghbi, S.S.; Baldwin, R.M.; Innis, R.B. Age-related decline in dopamine transporters: Analysis of striatal subregions, nonlinear effects, and hemispheric asymmetries. Am. J. Geriatr. Psychiatry 2002, 10, 36–43. [Google Scholar] [CrossRef]
- Wacker, J. Effects of positive emotion, extraversion, and dopamine on cognitive stability-flexibility and frontal EEG asymmetry. Psychophysiology 2018, 55, e12727. [Google Scholar] [CrossRef]
- Bernstein, N.A. Physiology of Movement and Activity, 1st ed.; Gasenko, O.G., Ed.; Nauka: Moscow, Russia, 1990. [Google Scholar]
- Bernstein’s Construction of Movements; Latash, M.L. (Ed.) Routledge: New York, NY, USA, 2020; ISBN 9780367816797. [Google Scholar]
- Profeta, V.L.S.; Turvey, M.T. Bernstein’s levels of movement construction: A contemporary perspective. Hum. Mov. Sci. 2018, 57, 111–133. [Google Scholar] [CrossRef]
- Mutha, P.K.; Haaland, K.Y.; Sainburg, R.L. Rethinking Motor Lateralization: Specialized but Complementary Mechanisms for Motor Control of Each Arm. PLoS ONE 2013, 8, e58582. [Google Scholar] [CrossRef]
- Gutwinski, S.; Löscher, A.; Mahler, L.; Kalbitzer, J.; Heinz, A.; Bermpohl, F. Understanding Left-Handedness. Dtsch. Aerzteblatt Online 2011. [Google Scholar] [CrossRef] [PubMed]
- Jager, G. On the hemispheric specialization for categorical and coordinate spatial relations: A review of the current evidence. Neuropsychologia 2003, 41, 504–515. [Google Scholar] [CrossRef]
- Alkadhi, H.; Brugger, P.; Boendermaker, S.H.; Crelier, G.; Curt, A.; Hepp-Reymond, M.-C.; Kollias, S.S. What Disconnection Tells about Motor Imagery: Evidence from Paraplegic Patients. Cereb. Cortex 2005, 15, 131–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Z.; Allison, B.Z.; Jin, J.; Zhang, Y.; Wang, X.; Li, W.; Cichocki, A. Optimized Motor Imagery Paradigm Based on Imagining Chinese Characters Writing Movement. IEEE Trans. Neural Syst. Rehabil. Eng. 2017, 25, 1009–1017. [Google Scholar] [CrossRef]
- Gustin, S.M.; Wrigley, P.J.; Gandevia, S.C.; Middleton, J.W.; Henderson, L.A.; Siddall, P.J. Movement imagery increases pain in people with neuropathic pain following complete thoracic spinal cord injury. Pain 2008, 137, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.; Chan, A.; Ng, A.; Luk, B. A preliminary analysis of movement times and subjective evaluations for a visually-controlled foot-tapping task on touch pad device. In Proceedings of the International MultiConference of Engineers and Computer Scientist, Hong Kong, China, 17–19 March 2010; Ao, S.I., Castillo, O., Douglas, C., Feng, D.D., Leaa, J.-A., Eds.; Newswood Limited: Hong Kong, China, 2010; pp. 1968–1970. [Google Scholar]
- Chan, A.H.S.; Hoffmann, E.R. Subjective Difficulty of Movements With Ongoing Visual Control. J. Mot. Behav. 2013, 45, 507–517. [Google Scholar] [CrossRef]
- Chan, A.H.S.; Hoffmann, E.R. Subjective Estimation of Task Time and Task Difficulty of Simple Movement Tasks. J. Mot. Behav. 2017, 49, 185–199. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
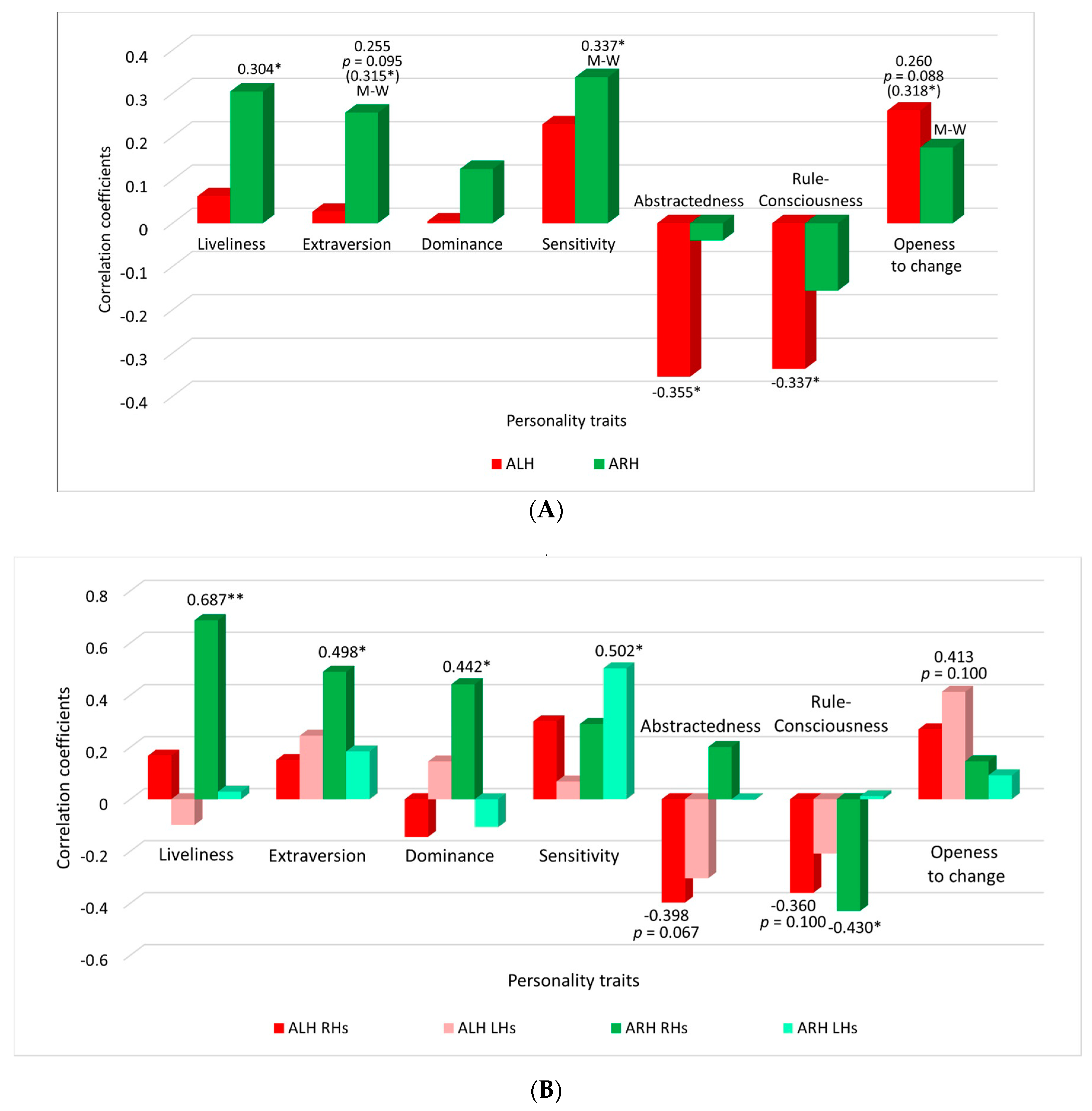
| Personality Trait (Cattell) | Accuracy of Classification of Brain States in Imagining Left Hand Movements and in Rest State (ALH) | Accuracy of Classification of Brain States in Imagining Right Hand Movements and in Rest State (ARH) | ||
|---|---|---|---|---|
| PCC | NCC | PCC | NCC | |
| Liveliness (F) | - | - | +0.346 * | +0.304 * |
| Extraversion (F2) | - | - | +0.315 * | +0.255 (p = 0.095) |
| Sensitivity (I) | - | - | +0.347 * | +0.337 * |
| Abstractedness (M) | −0.270 (p = 0,076) | −0.355 * | - | - |
| Rule- consciousness (G) | −0.362 * | −0.337 * | - | - |
| Openness to change (Q1) | +0.318 * | +0.260 (p = 0.088) | - | - |
| “Pure” Right-Handers | Latent Left-Handers | All Participants | |
|---|---|---|---|
| Accuracy of classification of the LH movements imagination comparative to rest (ALH) | 66.9 ± 8.5% | 66.5 ± 10.4% | 66.2 ± 9.1% |
| Accuracy of classification of the RH movements imagination comparative to rest (ARH) | 68.7 ± 8% | 65.6 ± 8.1% | 67.7 ± 8% |
| Accuracy of classification of the imagination of RH movements comparative to LH (ARLH) | 62.4 ± 7.1% | 63 ± 6.9% | 62.9 ± 7% |
| Mean | 66 ± 8.2% | 65 ± 8.5% | 65.6 ± 8.3% |
| Personality Trait (Cattell) | Accuracy of Classification of Brain States in Imagining Left Hand Movements and in Rest State (ALH) | Accuracy of Classification of Brain States in Imagining Right Hand Movements and in Rest State (ARH) | ||
|---|---|---|---|---|
| PCC | NCC | PCC | NCC | |
| Liveliness (F) | - | - | All +0.346 * | All +0.304 * |
| - | - | RHs +0.656 * | RHs +0.687 ** | |
| - | - | - | - | |
| Extraversion (F2) | - | - | All +0.315 * | All +0.255 (p = 0.095) |
| - | - | RHs +0.420 * | RHs +0.489 * | |
| - | - | - | - | |
| Dominance (E) | - | - | - | - |
| - | - | - | - | |
| - | - | - | RHs +0.442 * | |
| Sensitivity (I) | - | - | All +0.347 * | All +0.337 * |
| - | - | - | - | |
| - | - | LHs +0.518 * | LHs +0.502 * | |
| Abstractedness (M) | All −0.270 (p = 0.076) | All −0.355 * | - | - |
| - | - | - | - | |
| - | - | - | - | |
| Rule- consciousness (G) | All −0.362 * RHs −0.478 * - | All −0.337 * RHs −0.360 (p = 0.1) - | - RHs −0.433 * - | - RHs −0.430 * - |
| Openness to change (Q1) | All +0.318 * - - | All +0.260 (p = 0.088) - LHs 0.413 (p = 0.1) | - - - | - - - |
| LH | RH | |||
|---|---|---|---|---|
| Mean SC | Correlations | Mean SC | Correlations | |
| Real movement | All 1.88 | - | All 1.96 | - |
| RHs 1.80 | RHs 0.527 * | RHs 1.87 | - | |
| LHs 2.00 | - | LHs 2.11 | - | |
| Motor imagery | All 2.73 | - | All 2.54 | All 0.697 ** |
| RHs 2.77 | - | RHs 2.40 | RHs 0.664 ** | |
| LHs 2.67 | - | LHs 2.78 | LHs 0.604 (p = 0.085) (0.685 *) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bobrova, E.V.; Reshetnikova, V.V.; Vershinina, E.A.; Grishin, A.A.; Bobrov, P.D.; Frolov, A.A.; Gerasimenko, Y.P. Success of Hand Movement Imagination Depends on Personality Traits, Brain Asymmetry, and Degree of Handedness. Brain Sci. 2021, 11, 853. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11070853
Bobrova EV, Reshetnikova VV, Vershinina EA, Grishin AA, Bobrov PD, Frolov AA, Gerasimenko YP. Success of Hand Movement Imagination Depends on Personality Traits, Brain Asymmetry, and Degree of Handedness. Brain Sciences. 2021; 11(7):853. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11070853
Chicago/Turabian StyleBobrova, Elena V., Varvara V. Reshetnikova, Elena A. Vershinina, Alexander A. Grishin, Pavel D. Bobrov, Alexander A. Frolov, and Yury P. Gerasimenko. 2021. "Success of Hand Movement Imagination Depends on Personality Traits, Brain Asymmetry, and Degree of Handedness" Brain Sciences 11, no. 7: 853. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11070853