

Identification and Characterization of Elevated Expression of Transferrin and Its Receptor TfR1 in Mouse Models of Depression
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
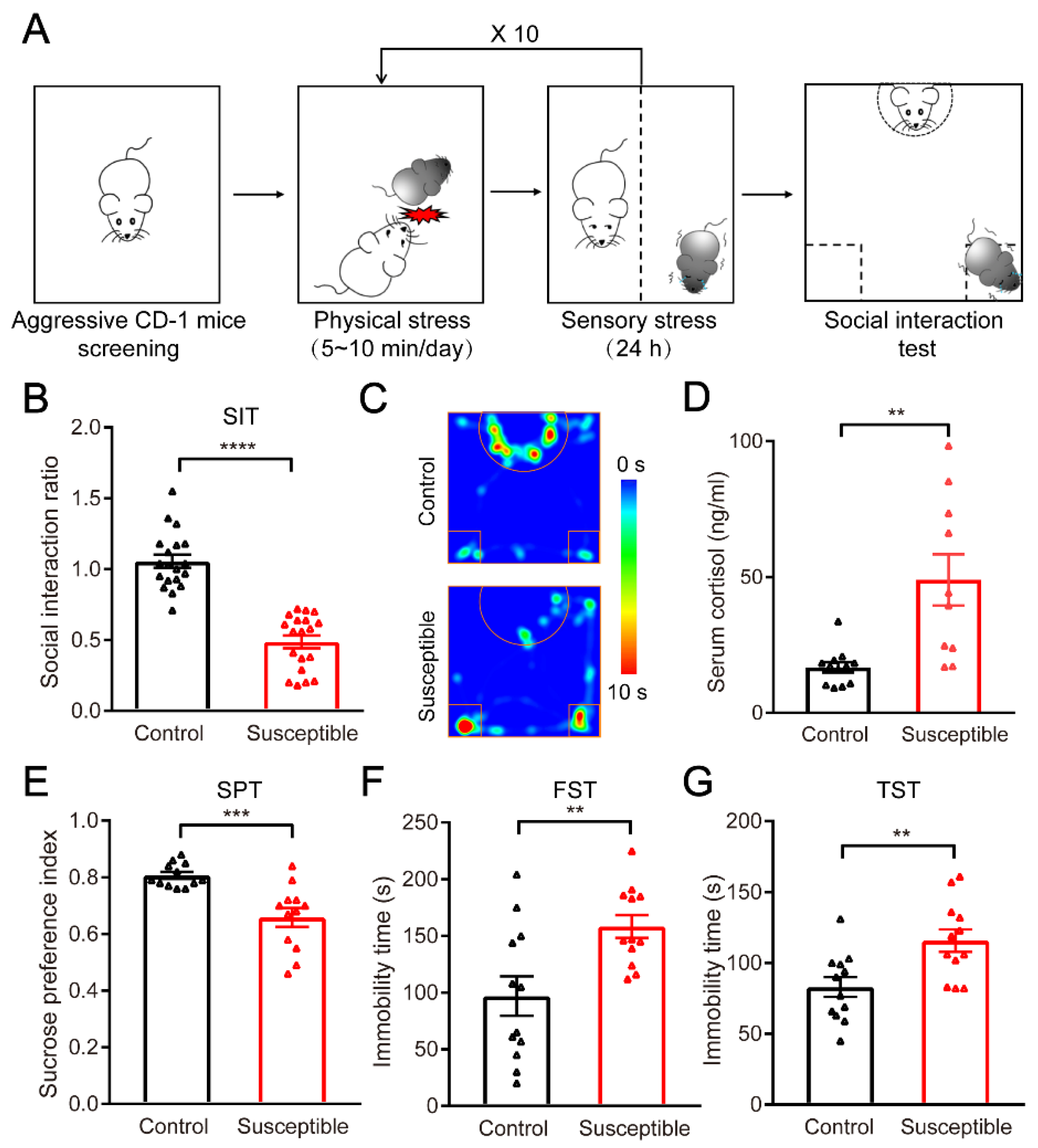
2.1. Chronic Social Defeat Stress Mice Model
2.2. Behavioral Experiments
2.2.1. Social Interaction Test
2.2.2. Sucrose Preference Test
2.2.3. Forced Swimming Test
2.2.4. Tail Suspension Test
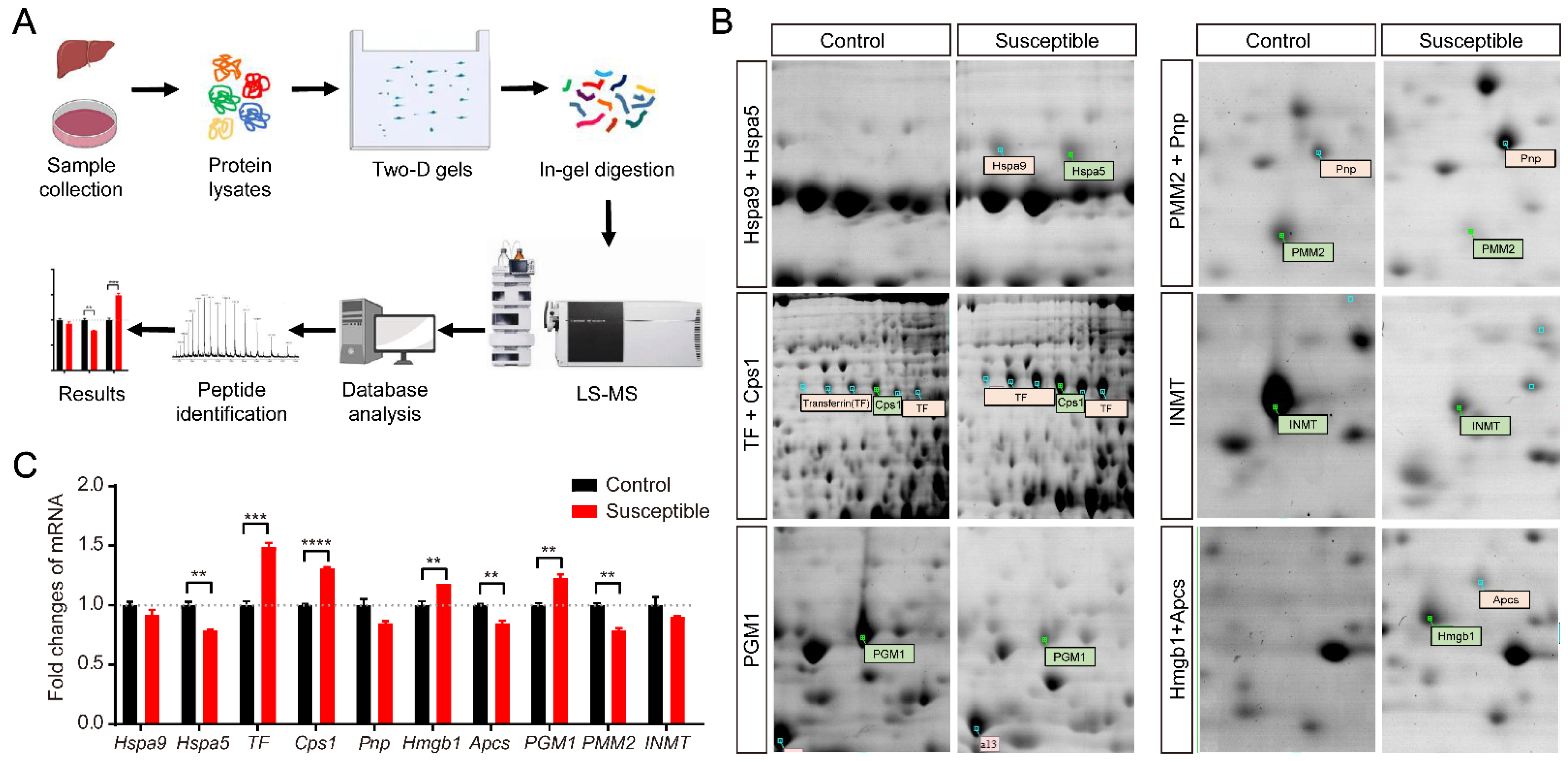
2.3. Two-Dimensional Electrophoresis and Mass Spectrometry
2.4. Quantitative Reverse Transcription PCR
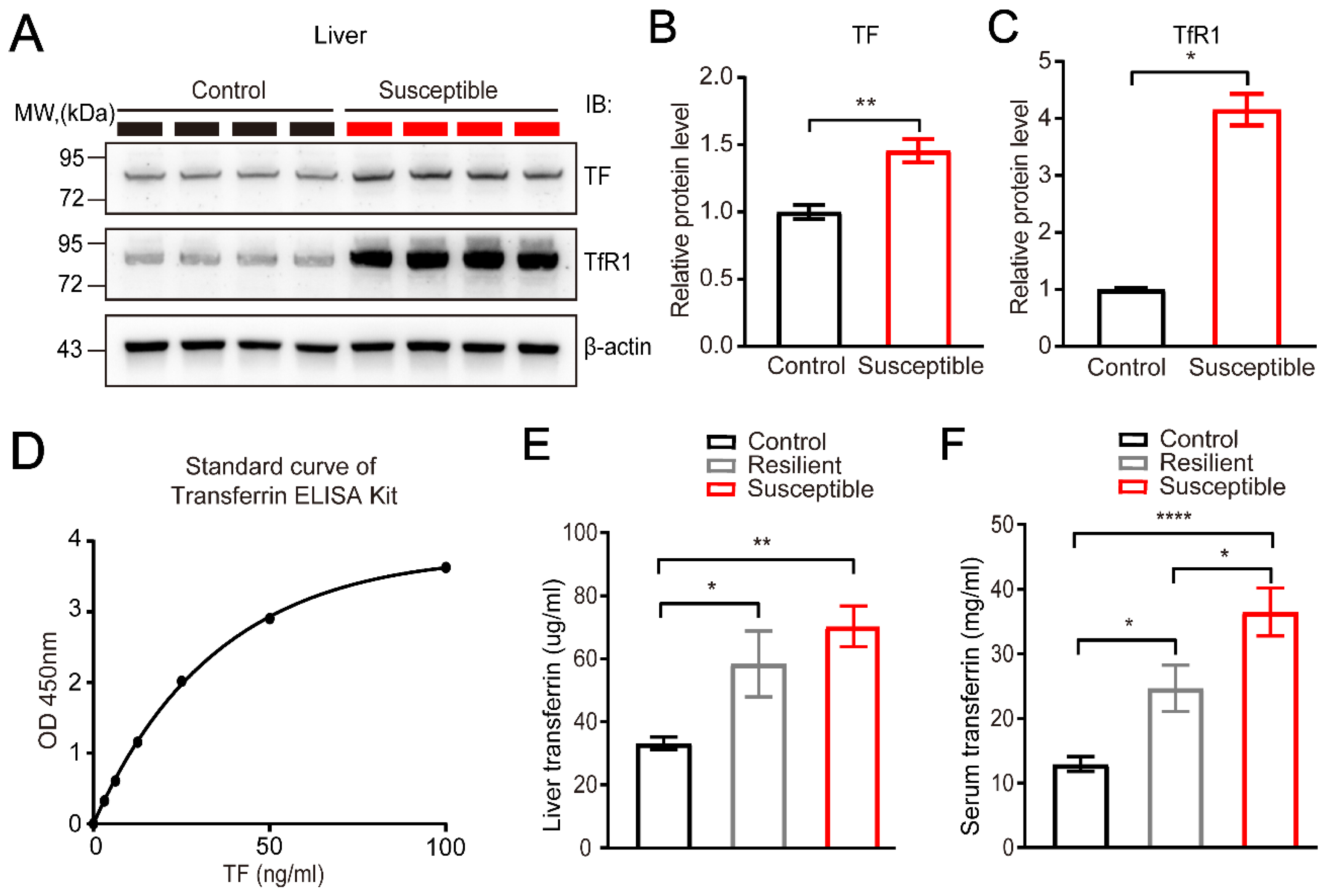
2.5. Protein Extraction and Western Blots
2.6. Enzyme Linked Immunosorbent Assay
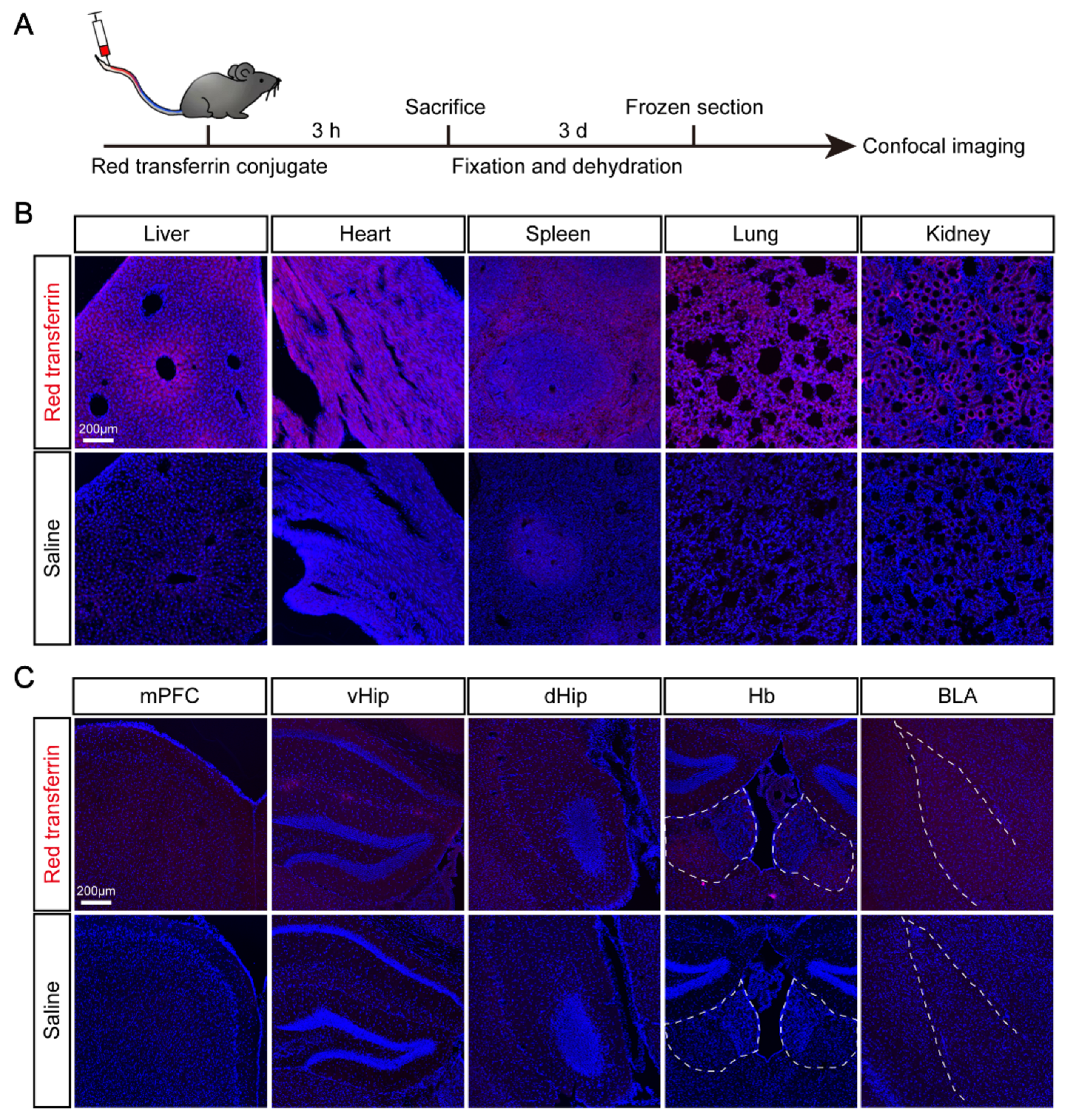
2.7. Red transferrin Conjugate Injection
2.8. Acquisition and Analysis of RNA Sequencing Data
2.9. Statistical Analysis
3. Results
3.1. Establishment and Evaluation of the CSDS Mouse Model
3.2. Identification of Differentially Expressed Proteins in the Stressed CSDS Liver by Two-Dimensional Electrophoresis and Mass Spectrometry
3.3. Characterization of the Elevated Expression of Transferrin in the Liver and Blood Serum Induced by CSDS
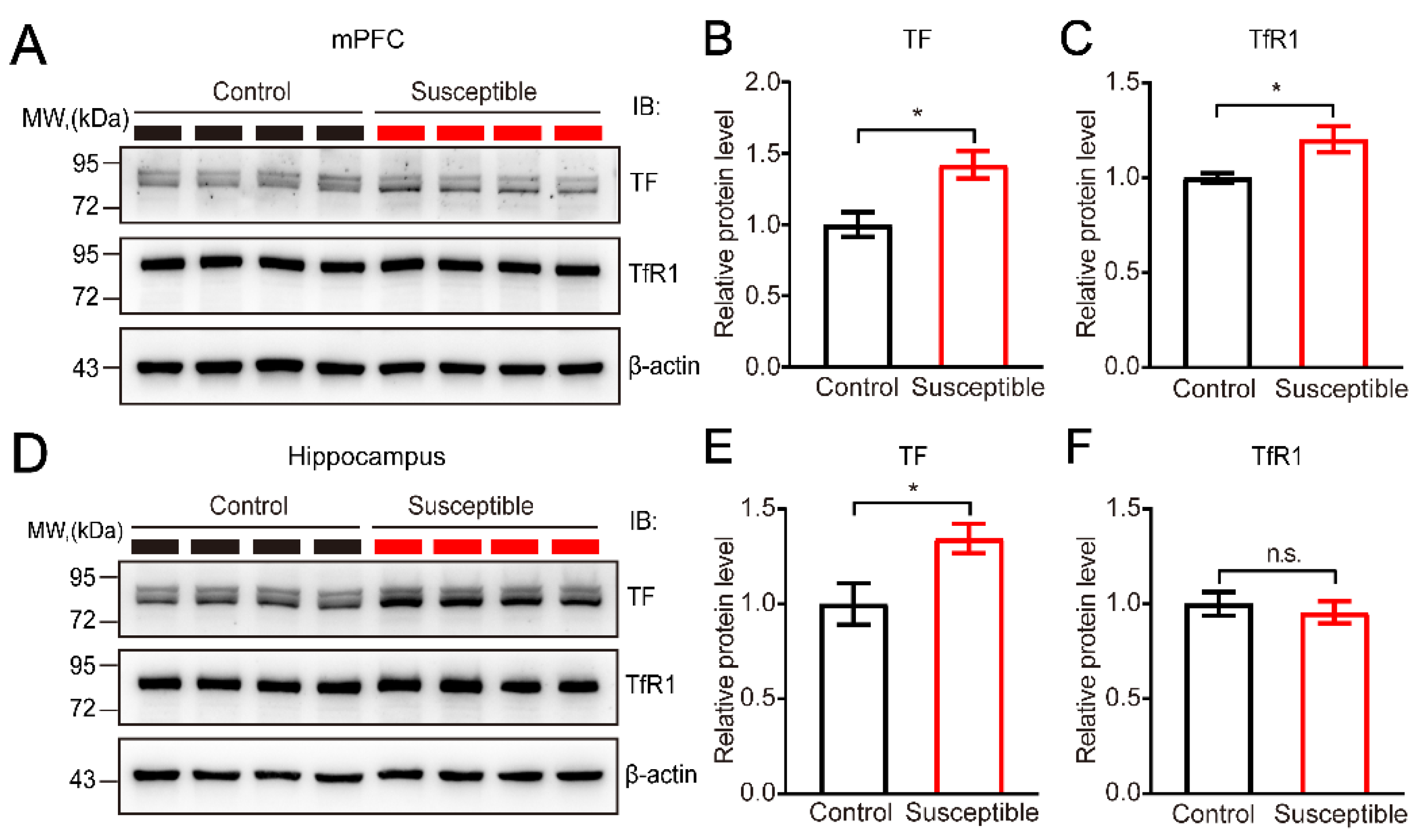
3.4. Elevated Expression of TF and Its Receptor TfR1 in Brain Regions after CSDS
3.5. The expression of TF and TfR1 in Stressed Animals from the Gene Expression Omnibus (GEO) Database
4. Discussion
5. Conclusions
6. Limitations and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flint, J.; Kendler, K.S. The Genetics of Major Depression. Neuron 2014, 81, 484–503. [Google Scholar] [CrossRef] [PubMed]
- Cassano, P.; Fava, M. Depression and public health: An overview. J. Psychosom. Res. 2002, 53, 849–857. [Google Scholar] [CrossRef]
- Tanaka, M.; Vecsei, L. Editorial of Special Issue “Crosstalk between Depression, Anxiety, and Dementia: Comorbidity in Behavioral Neurology and Neuropsychiatry”. Biomedicines 2021, 9, 517. [Google Scholar] [CrossRef]
- Mathers, C.D.; Loncar, D. Projections of global mortality and burden of disease from 2002 to 2030. PLoS Med. 2006, 3, e442. [Google Scholar] [CrossRef]
- Weisz, J.R.; McCarty, C.A.; Valeri, S.M. Effects of psychotherapy for depression in children and adolescents: A meta-analysis. Psychol. Bull. 2006, 132, 132–149. [Google Scholar]
- Weisz, J.R.; Kuppens, S.; Eckshtain, D.; Ugueto, A.M.; Hawley, K.M.; Jensen-Doss, A. Performance of evidence-based youth psychotherapies compared with usual clinical care: A multilevel meta-analysis. JAMA Psychiatry 2013, 70, 750–761. [Google Scholar] [CrossRef]
- Blazer, D.G. Depression in late life: Review and commentary. The journals of gerontology. Ser. A Biol. Sci. Med. Sci. 2003, 58, 249–265. [Google Scholar]
- Howard, D.M.; Adams, M.J.; Clarke, T.K.; Hafferty, J.D.; Gibson, J.; Shirali, M.; Coleman, J.R.I.; Hagenaars, S.P.; Ward, J.; Wigmore, E.M.; et al. Genome-wide meta-analysis of depression identifies 102 independent variants and highlights the importance of the prefrontal brain regions. Nat. Neurosci. 2019, 22, 343–352. [Google Scholar]
- Wingo, T.S.; Liu, Y.; Gerasimov, E.S.; Gockley, J.; Logsdon, B.A.; Duong, D.M.; Dammer, E.B.; Lori, A.; Kim, P.J.; Ressler, K.J.; et al. Brain proteome-wide association study implicates novel proteins in depression pathogenesis. Nat. Neurosci. 2021, 24, 810–817. [Google Scholar] [CrossRef]
- Castren, E.; Monteggia, L.M. Brain-Derived Neurotrophic Factor Signaling in Depression and Antidepressant Action. Biol. Psychiatry 2021, 90, 128–136. [Google Scholar] [CrossRef]
- Faugere, M.; Micoulaud-Franchi, J.A.; Faget-Agius, C.; Lancon, C.; Cermolacce, M.; Richieri, R. Quality of life is associated with chronic inflammation in depression: A cross-sectional study. J. Affect. Disord. 2018, 227, 494–497. [Google Scholar] [CrossRef]
- Ancelin, M.L.; Scali, J.; Norton, J.; Ritchie, K.; Dupuy, A.M.; Chaudieu, I.; Ryan, J. Heterogeneity in HPA axis dysregulation and serotonergic vulnerability to depression. Psychoneuroendocrinology 2017, 77, 90–94. [Google Scholar] [CrossRef]
- Tanaka, M.; Szabo, A.; Spekker, E.; Polyak, H.; Toth, F.; Vecsei, L. Mitochondrial Impairment: A Common Motif in Neuropsychiatric Presentation? The Link to the Tryptophan-Kynurenine Metabolic System. Cells 2022, 11, 2607. [Google Scholar] [CrossRef]
- Smagin, D.A.; Kovalenko, I.L.; Galyamina, A.G.; Belozertseva, I.V.; Tamkovich, N.V.; Baranov, K.O.; Kudryavtseva, N.N. Chronic Lithium Treatment Affects Anxious Behaviors and theExpression of Serotonergic Genes in Midbrain Raphe Nuclei of Defeated Male Mice. Biomedicines 2021, 9, 1293. [Google Scholar] [CrossRef]
- Zhang, F.F.; Peng, W.; Sweeney, J.A.; Jia, Z.Y.; Gong, Q.Y. Brain structure alterations in depression: Psychoradiological evidence. CNS Neurosci. Ther. 2018, 24, 994–1003. [Google Scholar] [CrossRef]
- Peng, W.; Chen, Z.; Yin, L.; Jia, Z.; Gong, Q. Essential brain structural alterations in major depressive disorder: A voxel-wise meta-analysis on first episode, medication-naive patients. J. Affect. Disord. 2016, 199, 114–123. [Google Scholar] [CrossRef]
- Ma, M.; Chang, X.; Wu, H. Animal models of stress and stress-related neurocircuits: A comprehensive review. Stress Brain 2021, 1, 108–127. [Google Scholar] [CrossRef]
- Battaglia, S.; Thayer, J.F. Functional interplay between central and autonomic nervous systems in human fear conditioning. Trends Neurosci. 2022, 45, 504–506. [Google Scholar] [CrossRef]
- Zhang, X.; Lei, B.; Yuan, Y.; Zhang, L.; Hu, L.; Jin, S.; Kang, B.; Liao, X.; Sun, W.; Xu, F.; et al. Brain control of humoral immune responses amenable to behavioural modulation. Nature 2020, 581, 204–208. [Google Scholar] [CrossRef]
- Suh, J.I.; Sakong, J.K.; Lee, K.; Lee, Y.K.; Park, J.B.; Kim, D.J.; Seo, Y.S.; Lee, J.D.; Ko, S.Y.; Lee, B.S.; et al. Anxiety and depression propensities in patients with acute toxic liver injury. World J. Gastroenterol. 2013, 19, 9069–9076. [Google Scholar] [CrossRef] [PubMed]
- Kortam, M.A.; Ali, B.M.; Fathy, N. The deleterious effect of stress-induced depression on rat liver: Protective role of resveratrol and dimethyl fumarate via inhibiting the MAPK/ERK/JNK pathway. J. Biochem. Mol. Toxicol. 2021, 35, e22627. [Google Scholar] [CrossRef]
- Chen, X.; Dong, J.; Wang, T.; Zhao, L. Liver fire invading the lung: Theoretical basis, identification and treatment. J. Tradit. Chin. Med. Sci. 2017, 4, 16–19. [Google Scholar] [CrossRef]
- Yan, C.; Xu, Z.W. Exploration on central neurobiological mechanisms of Gan in taking charge of dispersion and regulating emotion. Zhongguo Zhong Xi Yi Jie He Za Zhi 2005, 25, 459–462. [Google Scholar]
- Huang, X.; Liu, X.; Yu, Y. Depression and Chronic Liver Diseases: Are There Shared Underlying Mechanisms? Front. Mol. Neurosci. 2017, 10, 134. [Google Scholar] [CrossRef] [Green Version]
- Kudryavtseva, N.N.; Bakshtanovskaya, I.V.; Koryakina, L.A. Social model of depression in mice of C57BL/6J strain. Pharmacol. Biochem. Behav. 1991, 38, 315–320. [Google Scholar] [CrossRef]
- Menard, C.; Pfau, M.L.; Hodes, G.E.; Kana, V.; Wang, V.X.; Bouchard, S.; Takahashi, A.; Flanigan, M.E.; Aleyasin, H.; LeClair, K.B.; et al. Social stress induces neurovascular pathology promoting depression. Nat. Neurosci. 2017, 20, 1752–1760. [Google Scholar] [CrossRef]
- Harris, A.Z.; Atsak, P.; Bretton, Z.H.; Holt, E.S.; Alam, R.; Morton, M.P.; Abbas, A.I.; Leonardo, E.D.; Bolkan, S.S.; Hen, R.; et al. A Novel Method for Chronic Social Defeat Stress in Female Mice. Neuropsychopharmacology 2018, 43, 1276–1283. [Google Scholar] [CrossRef]
- Krishnan, V.; Han, M.H.; Graham, D.L.; Berton, O.; Renthal, W.; Russo, S.J.; Laplant, Q.; Graham, A.; Lutter, M.; Lagace, D.C.; et al. Molecular adaptations underlying susceptibility and resistance to social defeat in brain reward regions. Cell 2007, 131, 391–404. [Google Scholar] [CrossRef]
- Rogers, A.B.; Houghton, J. Helicobacter-based mouse models of digestive system carcinogenesis. Methods Mol. Biol. 2009, 511, 267–295. [Google Scholar]
- Golden, S.A.; Covington, H.E., 3rd; Berton, O.; Russo, S.J. A standardized protocol for repeated social defeat stress in mice. Nat. Protoc. 2011, 6, 1183–1191. [Google Scholar] [CrossRef]
- Yin, C.Y.; Li, L.D.; Xu, C.; Du, Z.W.; Wu, J.M.; Chen, X.; Xia, T.; Huang, S.Y.; Meng, F.; Zhang, J.; et al. A novel method for automatic pharmacological evaluation of sucrose preference change in depression mice. Pharmacol. Res. 2021, 168, 105601. [Google Scholar] [CrossRef] [PubMed]
- Iñiguez, S.D.; Riggs, L.M.; Nieto, S.J.; Dayrit, G.; Zamora, N.N.; Shawhan, K.L.; Cruz, B.; Warren, B.L. Social defeat stress induces a depression-like phenotype in adolescent male c57BL/6 mice. Stress 2014, 17, 247–255. [Google Scholar] [CrossRef]
- Castagné, V.; Moser, P.; Roux, S.; Porsolt, R.D. Rodent models of depression: Forced swim and tail suspension behavioral despair tests in rats and mice. Curr. Protoc. Neurosci. 2010, 49, 5–8. [Google Scholar] [CrossRef]
- Herbert, B.; Galvani, M.; Hamdan, M.; Olivieri, E.; MacCarthy, J.; Pedersen, S.; Righetti, P.G. Reduction and alkylation of proteins in preparation of two-dimensional map analysis: Why, when, and how? Electrophoresis 2001, 22, 2046–2057. [Google Scholar] [CrossRef]
- Langen, H.; Röder, D.; Juranville, J.F.; Fountoulakis, M. Effect of protein application mode and acrylamide concentration on the resolution of protein spots separated by two-dimensional gel electrophoresis. Electrophoresis 1997, 18, 2085–2090. [Google Scholar] [CrossRef]
- Fernandez, J.; Gharahdaghi, F.; Mische, S.M. Routine identification of proteins from sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gels or polyvinyl difluoride membranes using matrix assisted laser desorption/ionization-time of flight-mass spectrometry (MALDI-TOF-MS). Electrophoresis 1998, 19, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Wook Koo, J.; Labonté, B.; Engmann, O.; Calipari, E.S.; Juarez, B.; Lorsch, Z.; Walsh, J.J.; Friedman, A.K.; Yorgason, J.T.; Han, M.H.; et al. Essential Role of Mesolimbic Brain-Derived Neurotrophic Factor in Chronic Social Stress-Induced Depressive Behaviors. Biol. Psychiatry 2016, 80, 469–478. [Google Scholar] [CrossRef]
- Gomme, P.T.; McCann, K.B.; Bertolini, J. Transferrin: Structure, function and potential therapeutic actions. Drug Discov. Today 2005, 10, 267–273. [Google Scholar] [CrossRef]
- Moos, T.; Morgan, E.H. Transferrin and transferrin receptor function in brain barrier systems. Cell. Mol. Neurobiol. 2000, 20, 77–95. [Google Scholar] [CrossRef] [PubMed]
- Skarlatos, S.; Yoshikawa, T.; Pardridge, W.M. Transport of [125I]transferrin through the rat blood-brain barrier. Brain Res. 1995, 683, 164–171. [Google Scholar] [CrossRef]
- Takahashi, A.; Flanigan, M.E.; McEwen, B.S.; Russo, S.J. Aggression, Social Stress, and the Immune System in Humans and Animal Models. Front. Behav. Neurosci. 2018, 12, 56. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, N. Prolonged Stress Leads to Serious Health Problems: Preventive Approaches. Int. J. Health Sci. 2016, 10, V–VI. [Google Scholar]
- Ewert, A.; Chang, Y. Levels of Nature and Stress Response. Behav. Sci. 2018, 8, 49. [Google Scholar] [CrossRef]
- Tampa, M.; Sarbu, M.I.; Mitran, M.I.; Mitran, C.I.; Matei, C.; Georgescu, S.R. The Pathophysiological Mechanisms and the Quest for Biomarkers in Psoriasis, a Stress-Related Skin Disease. Dis. Markers 2018, 2018, 5823684. [Google Scholar] [CrossRef] [Green Version]
- McEwen, B.S. Biomarkers for assessing population and individual health and disease related to stress and adaptation. Metabolism 2015, 64, S2–S10. [Google Scholar] [CrossRef]
- Dhama, K.; Latheef, S.K.; Dadar, M.; Samad, H.A.; Munjal, A.; Khandia, R.; Karthik, K.; Tiwari, R.; Yatoo, M.I.; Bhatt, P.; et al. Biomarkers in Stress Related Diseases/Disorders: Diagnostic, Prognostic, and Therapeutic Values. Front. Mol. Biosci 2019, 6, 91. [Google Scholar] [CrossRef]
- Pratt, R.; Stapelberg, N.J.C. Early warning biomarkers in major depressive disorder: A strategic approach to a testing question. Biomarkers 2018, 23, 563–572. [Google Scholar] [CrossRef]
- Gadad, B.S.; Jha, M.K.; Czysz, A.; Furman, J.L.; Mayes, T.L.; Emslie, M.P.; Trivedi, M.H. Peripheral biomarkers of major depression and antidepressant treatment response: Current knowledge and future outlooks. J. Affect. Disord. 2018, 233, 3–14. [Google Scholar] [CrossRef]
- Domenici, E.; Muglia, P. The search for peripheral disease markers in psychiatry by genomic and proteomic approaches. Expert Opin. Med. Diagn. 2007, 1, 235–251. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Lu, S.; Zhang, C.; Zhu, L.; Li, Y.; Bai, M.; Xu, E. Quantitative proteomic analysis of the liver reveals antidepressant potential protein targets of Sinisan in a mouse CUMS model of depression. Biomed. Pharmacother. 2020, 130, 110565. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhao, M.; Magnussen, C.G.; Veeranki, S.P.; Xi, B. Psychological distress and mortality among US adults: Prospective cohort study of 330 367 individuals. J. Epidemiol. Community Health 2020, 74, 384–390. [Google Scholar] [CrossRef]
- Kaufling, J. Alterations and adaptation of ventral tegmental area dopaminergic neurons in animal models of depression. Cell Tissue Res. 2019, 377, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Boyer, J.L. Psychological stress is associated with relapse in type 1 autoimmune hepatitis. Liver Int. Off. J. Int. Assoc. Study Liver 2010, 30, 1439–1447. [Google Scholar] [CrossRef]
- Park, M.J.; D’Alecy, L.G.; Anderson, M.A.; Basrur, V.; Feng, Y.; Brady, G.F.; Kim, D.I.; Wu, J.; Nesvizhskii, A.I.; Lahann, J.; et al. Constitutive release of CPS1 in bile and its role as a protective cytokine during acute liver injury. Proc. Natl. Acad. Sci. USA 2019, 116, 9125–9134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, H.E.; Raucci, A. Alarmin(g) news about danger: Workshop on innate danger signals and HMGB1. EMBO Rep. 2006, 7, 774–778. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.E. DAMPs, PAMPs and alarmins: All we need to know about danger. J. Leukoc. Biol. 2007, 81, 1–5. [Google Scholar] [CrossRef]
- Ge, X.; Antoine, D.J.; Lu, Y.; Arriazu, E.; Leung, T.M.; Klepper, A.L.; Branch, A.D.; Fiel, M.I.; Nieto, N. High mobility group box-1 (HMGB1) participates in the pathogenesis of alcoholic liver disease (ALD). J. Biol. Chem. 2014, 289, 22672–22691. [Google Scholar] [CrossRef]
- Yamamoto, T.; Tajima, Y. HMGB1 is a promising therapeutic target for acute liver failure. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 673–682. [Google Scholar] [CrossRef]
- Matthijs, G.; Schollen, E.; Pardon, E.; Veiga-Da-Cunha, M.; Jaeken, J.; Cassiman, J.J.; Van Schaftingen, E. Mutations in PMM2, a phosphomannomutase gene on chromosome 16p13, in carbohydrate-deficient glycoprotein type I syndrome (Jaeken syndrome). Nat. Genet. 1997, 16, 88–92. [Google Scholar] [CrossRef] [PubMed]
- de Lonlay, P.; Seta, N.; Barrot, S.; Chabrol, B.; Drouin, V.; Gabriel, B.M.; Journel, H.; Kretz, M.; Laurent, J.; Le Merrer, M.; et al. A broad spectrum of clinical presentations in congenital disorders of glycosylation I: A series of 26 cases. J. Med. Genet. 2001, 38, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Dudek, K.A.; Dion-Albert, L.; Lebel, M.; LeClair, K.; Labrecque, S.; Tuck, E.; Ferrer Perez, C.; Golden, S.A.; Tamminga, C.; Turecki, G.; et al. Molecular adaptations of the blood-brain barrier promote stress resilience vs. depression. Proc. Natl. Acad. Sci. USA 2020, 117, 3326–3336. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Van de Vyvere, J.; Vandoolaeghe, E.; Bril, T.; Demedts, P.; Wauters, A.; Neels, H. Alterations in iron metabolism and the erythron in major depression: Further evidence for a chronic inflammatory process. J. Affect. Disord. 1996, 40, 23–33. [Google Scholar] [CrossRef]
- Berthou, C.; Iliou, J.P.; Barba, D. Iron, neuro-bioavailability and depression. EJHaem 2022, 3, 263–275. [Google Scholar] [CrossRef]
- Wang, Z.; Zeng, Y.N.; Yang, P.; Jin, L.Q.; Xiong, W.C.; Zhu, M.Z.; Zhang, J.Z.; He, X.; Zhu, X.H. Axonal iron transport in the brain modulates anxiety-related behaviors. Nat. Chem. Biol. 2019, 15, 1214–1222. [Google Scholar] [CrossRef]
- Liang, S.; Lu, Y.; Li, Z.; Li, S.; Chen, B.; Zhang, M.; Chen, B.; Ji, M.; Gong, W.; Xia, M.; et al. Iron Aggravates the Depressive Phenotype of Stressed Mice by Compromising the Glymphatic System. Neurosci. Bull. 2020, 36, 1542–1546. [Google Scholar] [CrossRef]
- Mehrpouya, S.; Nahavandi, A.; Khojasteh, F.; Soleimani, M.; Ahmadi, M.; Barati, M. Iron administration prevents BDNF decrease and depressive-like behavior following chronic stress. Brain Res. 2015, 1596, 79–87. [Google Scholar] [CrossRef]
- Ferreira, A.; Neves, P.; Gozzelino, R. Multilevel Impacts of Iron in the Brain: The Cross Talk between Neurophysiological Mechanisms, Cognition, and Social Behavior. Pharmaceuticals 2019, 12, 126. [Google Scholar] [CrossRef]
- Maes, M.; Meltzer, H.Y.; Buckley, P.; Bosmans, E. Plasma-soluble interleukin-2 and transferrin receptor in schizophrenia and major depression. Eur. Arch. Psychiatry Clin. Neurosci. 1995, 244, 325–329. [Google Scholar] [CrossRef]
- Tsai, S.Y.; Lee, H.C.; Chen, C.C.; Lee, C.H. Plasma levels of soluble transferrin receptors and Clara cell protein (CC16) during bipolar mania and subsequent remission. J. Psychiatr. Res. 2003, 37, 229–235. [Google Scholar] [CrossRef]
- Zecca, L.; Casella, L.; Albertini, A.; Bellei, C.; Zucca, F.A.; Engelen, M.; Zadlo, A.; Szewczyk, G.; Zareba, M.; Sarna, T. Neuromelanin can protect against iron-mediated oxidative damage in system modeling iron overload of brain aging and Parkinson’s disease. J. Neurochem. 2008, 106, 1866–1875. [Google Scholar]
- Xu, W.; Zhi, Y.; Yuan, Y.; Zhang, B.; Shen, Y.; Zhang, H.; Zhang, K.; Xu, Y. Correlations between abnormal iron metabolism and non-motor symptoms in Parkinson’s disease. J. Neural Transm. 2018, 125, 1027–1032. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequences (5′ to 3′) | Product Size |
|---|---|---|
| Hspa9 | F: ATGGCTGGAATGGCCTTAGC | 101 bp |
| R: ACCCAAATCAATACCAACCACTG | ||
| Hspa5 | F: ACTTGGGGACCACCTATTCCT | 134 bp |
| R: ATCGCCAATCAGACGCTCC | ||
| Tf | F: TGGGGGTTGGGTGTACGAT | 103 bp |
| R: AGCGTAGTAGTAGGTCTGTGG | ||
| Cps1 | F: ACATGGTGACCAAGATTCCTCG | 119 bp |
| R: TTCCTCAAAGGTGCGACCAAT | ||
| Pnp | F: ATCTGTGGTTCCGGCTTAGGA | 194 bp |
| R: TGGGGAAAGTTGGGTATCTCAT | ||
| Hmgb1 | F: GGCGAGCATCCTGGCTTATC | 86 bp |
| R: GGCTGCTTGTCATCTGCTG | ||
| Apcs | F: AGACAGACCTCAAGAGGAAAGT | 117 bp |
| R: AGGTTCGGAAACACAGTGTAAAA | ||
| PGM1 | F: CAGAACCCTTTAACCTCTGAGTC | 137 bp |
| R: CGAGAAATCCCTGCTCCCATAG | ||
| PMM2 | F: TGGTAGGTGGGTCAGATTTTGA | 93 bp |
| R: CCAAGCCATTCTCTGGAAACA | ||
| INMT | F: GCAGAGCAGGAAATCGTAAAGT | 167 bp |
| R: GGGGTGTAGTCAGTGACAATGAT | ||
| Actb1 | F: GTCCCAGACATCAGGGAGTAA | 102 bp |
| R: TCGGATACTTCAGCGTCAGGA |
| Name | UniProt Code | ProtScore | Mass | Matches | Sequences |
|---|---|---|---|---|---|
| Hspa9 | P38647 | 1817 | 73,701 | 45 | 19 |
| Hspa5 | P20029 | 711 | 72,492 | 24 | 14 |
| TF | Q921l1 | 7030 | 78,841 | 204 | 34 |
| Cps1 | Q8C196 | 2995 | 165,711 | 85 | 43 |
| PGM1 | Q9D0F9 | 1366 | 61,665 | 34 | 19 |
| PMM2 | Q9Z2M7 | 1265 | 27,981 | 38 | 10 |
| Pnp | P23492 | 8749 | 32,541 | 223 | 16 |
| INMT | P40936 | 1401 | 30,068 | 41 | 9 |
| Hmgb1 | P63158 | 933 | 25,049 | 20 | 7 |
| Apcs | P12246 | 1122 | 26,401 | 30 | 5 |
| GEO Accession | Models | Tissues | logFC | p Value |
|---|---|---|---|---|
| GSE151807 | CMS | amygdala | −0.273 | 0.0010 |
| GSE151807 | CMS | hippocampus | 0.296 | 0.0013 |
| GSE151807 | CMS | PFC | −0.099 | 0.0035 |
| GSE151807 | CMS | cerebral cortex | 0.419 | 0.0016 |
| GSE100236 | Handling | dorsal hippocampus | 0.060 | 0.4546 |
| GSE100236 | Handling | ventral hippocampus | 0.144 | 0.0305 |
| GSE100236 | Restraint | dorsal hippocampus | 0.177 | 0.0555 |
| GSE100236 | Restraint | ventral hippocampus | 0.071 | 0.1506 |
| GSE100236 | FS | dorsal hippocampus | 0.246 | 0.0006 |
| GSE100236 | FS | ventral hippocampus | 0.033 | 0.5636 |
| GSE172451 | CSI | ventral hippocampus | 0.051 | 0.0418 |
| GSE172451 | CSI + RES | ventral hippocampus | 0.103 | 0.0042 |
| GSE180055 | ELS | mPFC | 0.039 | 0.1343 |
| GSE68077 | PTSD | blood | 0.781 | 0.6360 |
| GSE68077 | CSDS | blood | 2.812 | 0.0025 |
| GSE68077 | PTSD | spleen | 2.095 | 0.0044 |
| GSE68077 | CSDS | spleen | 1.339 | 0.1120 |
| GSE68077 | PTSD | heart | 0.598 | 0.2926 |
| GSE68077 | CSDS | heart | −0.417 | 0.2320 |
| GEO Accession | Models | Tissue | logFC | p Value |
|---|---|---|---|---|
| GSE151807 | CMS | amygdala | −0.523 | 0.0001 |
| GSE151807 | CMS | hippocampus | 0.660 | <0.0001 |
| GSE151807 | CMS | PFC | 0.326 | <0.0001 |
| GSE151807 | CMS | cerebral cortex | 0.845 | <0.0001 |
| GSE100236 | Handling | dorsal hippocampus | −0.144 | 0.13243 |
| GSE100236 | Handling | ventral hippocampus | −0.243 | 8.04 × 10−5 |
| GSE100236 | Restraint | dorsal hippocampus | −0.146 | 0.035729 |
| GSE100236 | Restraint | ventral hippocampus | −0.246 | 0.000285 |
| GSE100236 | FS | dorsal hippocampus | 0.070 | 0.444395 |
| GSE100236 | FS | ventral hippocampus | −0.241 | 0.000132 |
| GSE172451 | CSI | ventral hippocampus | 0.103 | 0.0115 |
| GSE172451 | CSI + RES | ventral hippocampus | 0.131 | 0.0062 |
| GSE180055 | ELS | mPFC | 0.062 | 0.0292 |
| GSE68077 | PTSD | blood | 0.179 | 0.667 |
| GSE68077 | CSDS | blood | 0.787 | 0.063435 |
| GSE68077 | PTSD | spleen | −0.388 | 0.225383 |
| GSE68077 | CSDS | spleen | 0.392 | 0.615 |
| GSE68077 | PTSD | heart | −1.008 | 0.098956 |
| GSE68077 | CSDS | heart | −1.038 | 0.0052 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, X.; Ma, M.; Chen, L.; Song, Z.; Zhao, Z.; Shen, W.; Jiang, H.; Wu, Y.; Fan, M.; Wu, H. Identification and Characterization of Elevated Expression of Transferrin and Its Receptor TfR1 in Mouse Models of Depression. Brain Sci. 2022, 12, 1267. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12101267
Chang X, Ma M, Chen L, Song Z, Zhao Z, Shen W, Jiang H, Wu Y, Fan M, Wu H. Identification and Characterization of Elevated Expression of Transferrin and Its Receptor TfR1 in Mouse Models of Depression. Brain Sciences. 2022; 12(10):1267. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12101267
Chicago/Turabian StyleChang, Xin, Mengxin Ma, Liping Chen, Zhihong Song, Zhe Zhao, Wei Shen, Huihui Jiang, Yan Wu, Ming Fan, and Haitao Wu. 2022. "Identification and Characterization of Elevated Expression of Transferrin and Its Receptor TfR1 in Mouse Models of Depression" Brain Sciences 12, no. 10: 1267. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12101267