
Early Detection of Male-Predominant Phenotypes in the Pattern of Ultrasonic Vocalizations Emitted by Autism Spectrum Disorder Model (Crmp4-Knockout) Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
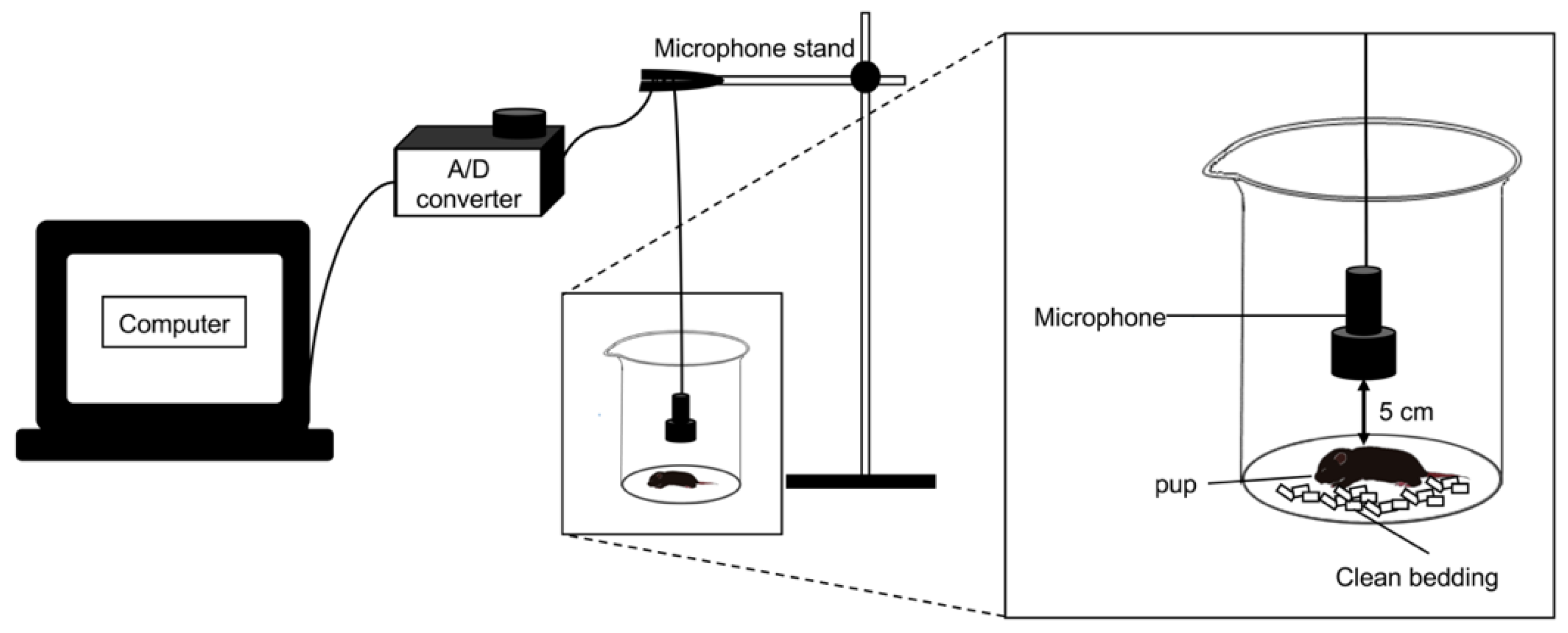
2.2. USVs
2.3. Statistical Analysis
3. Results
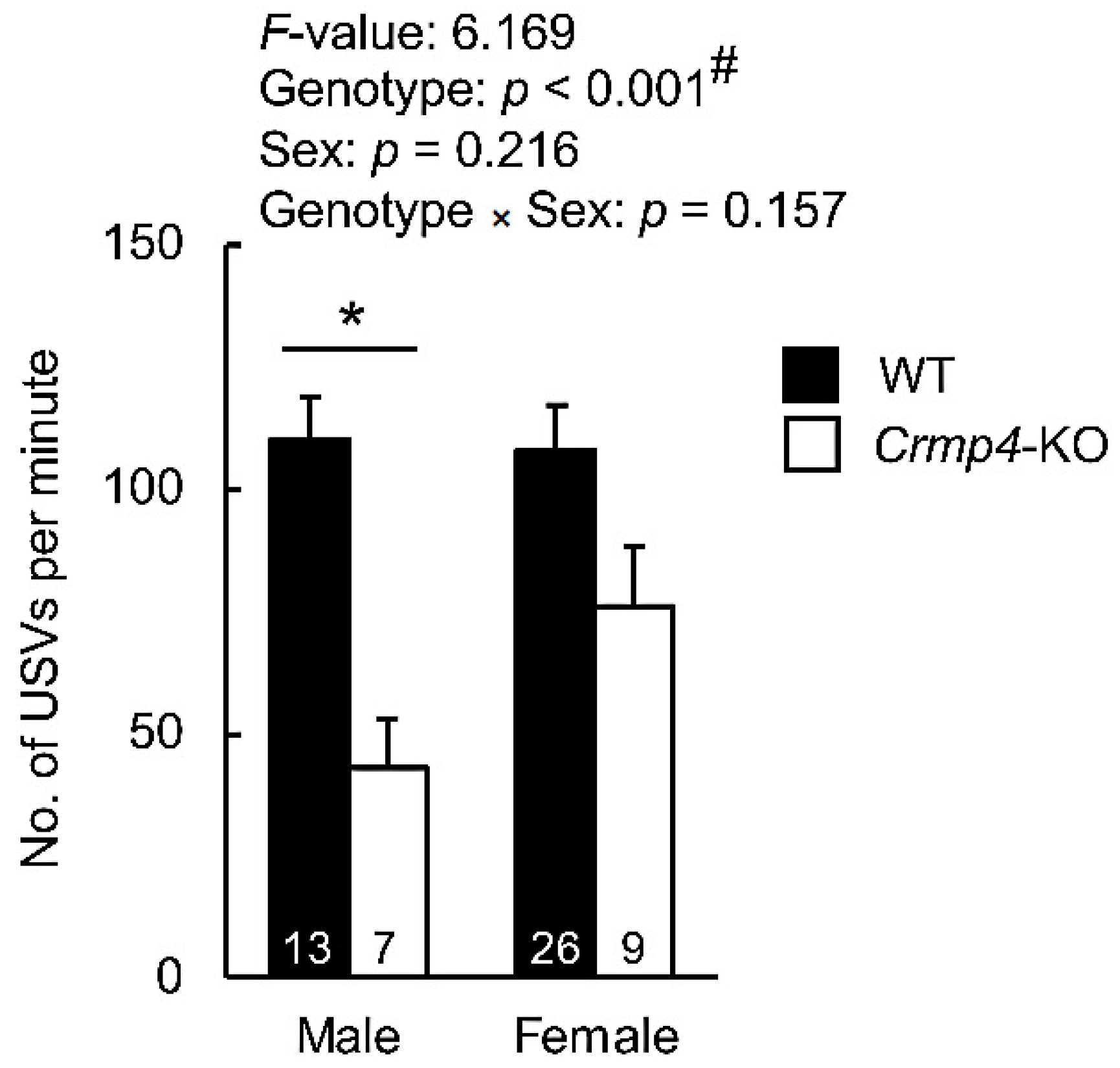
3.1. Total Number of USVs Was Decreased in Male Crmp4-KO Pups
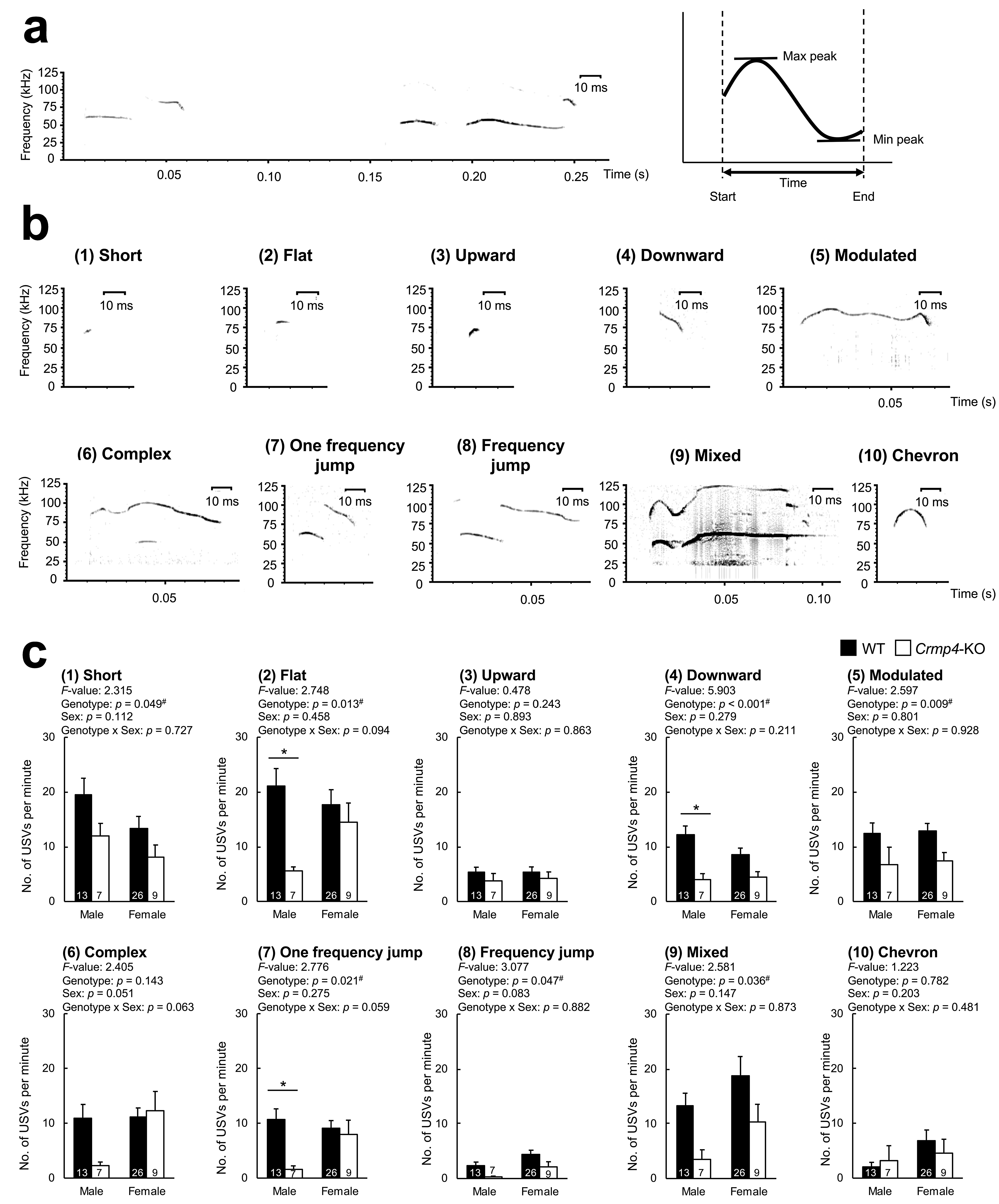
3.2. Crmp4-KO Decreased Specific USV Types in a Sex-Dependent Manner
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; (DSM-5); American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar]
- Hus, Y.; Segal, O. Challenges surrounding the diagnosis of autism in children. Neuropsychiatr. Dis. Treat 2021, 17, 3509–3529. [Google Scholar] [CrossRef] [PubMed]
- Maenner, M.J.; Shaw, K.A.; Bakian, A.V.; Bilder, D.A.; Durkin, M.S.; Esler, A.; Furnier, S.M.; Hallas, L.; Hall-Lande, J.; Hudson, A.; et al. Prevalence and characteristics of autism spectrum disorder among children aged 8 years—Autism and developmental disabilities monitoring network, 11 sites, United States, 2018. MMWR Surveill. Summ. 2021, 70, 1–16. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). ASD Fact Sheet Newsroom. Available online: https://www.who.int/news-room/fact-sheets/detail/autism-spectrum-disorders (accessed on 16 March 2022).
- Enriquez, K.D.; Gupta, A.R.; Hoffman, E.J. Signaling pathways and sex differential processes in autism spectrum disorder. Front. Psychiatry 2021, 12, 716673. [Google Scholar] [CrossRef] [PubMed]
- Schaafsma, S.M.; Pfaff, D.W. Etiologies underlying sex differences in autism spectrum disorders. Front. Neuroendocrinol. 2014, 35, 255–271. [Google Scholar] [CrossRef]
- Bonsi, P.; De Jaco, A.; Fasano, L.; Gubellini, P. Postsynaptic autism spectrum disorder genes and synaptic dysfunction. Neurobiol. Dis. 2022, 162, 105564. [Google Scholar] [CrossRef]
- Ohtani-Kaneko, R. Crmp4-KO mice as an animal model for investigating certain phenotypes of autism spectrum disorders. Int. J. Mol. Sci. 2019, 20, 2485. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, J.C.; Reim, D.; Boeckers, T.M.; Schmeisser, M.J. Genetic animal models for autism spectrum disorder. Curr. Top. Behav. Neurosci. 2017, 30, 311–324. [Google Scholar] [CrossRef]
- Silverman, J.L.; Thurm, A.; Ethridge, S.B.; Soller, M.M.; Petkova, S.P.; Abel, T.; Bauman, M.D.; Brodkin, E.S.; Harony-Nicolas, H.; Wöhr, M.; et al. Reconsidering animal models used to study autism spectrum disorder: Current state and optimizing future. Genes Brain Behav. 2022, e12803. [Google Scholar] [CrossRef]
- Sledziowska, M.; Galloway, J.; Baudouin, S.J. Evidence for a contribution of the Nlgn3/Cyfip1/Fmr1 pathway in the pathophysiology of autism spectrum disorders. Neuroscience 2020, 445, 31–41. [Google Scholar] [CrossRef]
- Gonçalves, J.; Violante, I.R.; Sereno, J.; Leitão, R.A.; Cai, Y.; Abrunhosa, A.; Silva, A.P.; Silva, A.J.; Castelo-Branco, M. Testing the excitation/inhibition imbalance hypothesis in a mouse model of the autism spectrum disorder: In vivo neurospectroscopy and molecular evidence for regional phenotypes. Mol. Autism 2017, 8, 47. [Google Scholar] [CrossRef]
- Sohal, V.S.; Rubenstein, J.L.R. Excitation-inhibition balance as a framework for investigating mechanisms in neuropsychiatric disorders. Mol. Psychiatry 2019, 24, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Ishii, K.; Nagaoka, A.; Kishida, Y.; Okazaki, H.; Yagishita, S.; Ucar, H.; Takahashi, N.; Saito, N.; Kasai, H. In vivo volume dynamics of dendritic spines in the neocortex of wild-type and Fmr1 KO mice. Eur. Neurol. 2018, 5, ENEURO.0282-18.2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathania, M.; Davenport, E.C.; Muir, J.; Sheehan, D.F.; López-Doménech, G.; Kittler, J.T. The autism and schizophrenia associated gene CYFIP1 is critical for the maintenance of dendritic complexity and the stabilization of mature spines. Transl. Psychiatry 2014, 4, e374. [Google Scholar] [CrossRef] [PubMed]
- Sarn, N.; Jaini, R.; Thacker, S.; Lee, H.; Dutta, R.; Eng, C. Cytoplasmic-predominant Pten increases microglial activation and synaptic pruning in a murine model with autism-like phenotype. Mol. Psychiatry 2021, 26, 1458–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsutiya, A.; Nakano, Y.; Hansen-Kiss, E.; Kelly, B.; Nishihara, M.; Goshima, Y.; Corsmeier, D.; White, P.; Herman, G.E.; Ohtani-Kaneko, R. Human CRMP4 mutation and disrupted Crmp4 expression in mice are associated with ASD characteristics and sexual dimorphism. Sci. Rep. 2017, 7, 16812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaff, D.W.; Rapin, I.; Goldman, S. Male predominance in autism: Neuroendocrine influences on arousal and social anxiety. Autism Res. 2011, 4, 163–176. [Google Scholar] [CrossRef]
- Schaafsma, S.M.; Gagnidze, K.; Reyes, A.; Norstedt, N.; Månsson, K.; Francis, K.; Pfaff, D.W. Sex-specific gene-environment interactions underlying ASD-like behaviors. Proc. Natl. Acad. Sci. USA 2017, 114, 1383–1388. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, H.; Sousa, A.C.; Sereno, J.; Martins, J.; Castelo-Branco, M.; Gonçalves, J. Sex-Dependent Social and Repetitive Behavior and Neurochemical Profile in Mouse Model of Autism Spectrum Disorder. Metabolites 2022, 12, 71. [Google Scholar] [CrossRef]
- Jeon, S.J.; Gonzales, E.L.; Mabunga, D.F.N.; Valencia, S.T.; Kim, D.G.; Kim, Y.; Adil, K.J.L.; Shin, D.; Park, D.; Shin, C.Y. Sex-specific behavioral features of rodent models of autism spectrum disorder. Exp. Neurobiol. 2018, 27, 321–343. [Google Scholar] [CrossRef]
- Iwakura, T.; Sakoh, M.; Tsutiya, A.; Yamashita, N.; Ohtani, A.; Tsuda, M.C.; Ogawa, S.; Tsukahara, S.; Nishihara, M.; Shiga, T.; et al. Collapsin response mediator protein 4 affects the number of tyrosine hydroxylase-immunoreactive neurons in the sexually dimorphic nucleus in female mice. Dev. Neurobiol. 2013, 73, 502–517. [Google Scholar] [CrossRef]
- Tsutiya, A.; Nishihara, M.; Goshima, Y.; Ohtani-Kaneko, R. Mouse pups lacking collapsin response mediator protein 4 manifest impaired olfactory function and hyperactivity in the olfactory bulb. Eur. J. Neurosci. 2015, 42, 2335–2345. [Google Scholar] [CrossRef] [PubMed]
- Tsutiya, A.; Watanabe, H.; Nakano, Y.; Nishihara, M.; Goshima, Y.; Ohtani-Kaneko, R. Deletion of collapsin response mediator protein 4 results in abnormal layer thickness and elongation of mitral cell apical dendrites in the neonatal olfactory bulb. J. Anat. 2016, 228, 792–804. [Google Scholar] [CrossRef] [PubMed]
- Mosienko, V.; Beis, D.; Alenina, N.; Wöhr, M. Reduced isolation-induced pup ultrasonic communication in mouse pups lacking brain serotonin. Mol. Autism 2015, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wöhr, M.; Roullet, F.I.; Hung, A.Y.; Sheng, M.; Crawley, J.N. Communication impairments in mice lacking Shank1: Reduced levels of ultrasonic vocalizations and scent marking behavior. PLoS ONE 2011, 6, e20631. [Google Scholar] [CrossRef] [Green Version]
- Niisato, E.; Nagai, J.; Yamashita, N.; Abe, T.; Kiyonari, H.; Goshima, Y.; Ohshima, T. CRMP4 suppresses apical dendrite bifurcation of CA1 pyramidal neurons in the mouse hippocampus. Dev. Neurobiol. 2012, 72, 1447–1457. [Google Scholar] [CrossRef]
- Kikusui, T.; Nakanishi, K.; Nakagawa, R.; Nagasawa, M.; Mogi, K.; Okanoya, K. Cross fostering experiments suggest that mice songs are innate. PLoS ONE 2011, 6, e17721. [Google Scholar] [CrossRef]
- Ey, E.; Torquet, N.; Le Sourd, A.M.; Leblond, C.S.; Boeckers, T.M.; Faure, P.; Bourgeron, T. The autism ProSAP1/Shank2 mouse model displays quantitative and structural abnormalities in ultrasonic vocalisations. Behav. Brain Res. 2013, 256, 677–689. [Google Scholar] [CrossRef]
- Scattoni, M.L.; Gandhy, S.U.; Ricceri, L.; Crawley, J.N. Unusual repertoire of vocalizations in the BTBR T+tf/J mouse model of autism. PLoS ONE 2008, 3, e3067. [Google Scholar] [CrossRef] [Green Version]
- Fish, E.W.; Sekinda, M.; Ferrari, P.F.; Dirks, A.; Miczek, K.A. Distress vocalizations in maternally separated mouse pups: Modulation via 5-HT(1A), 5-HT(1B) and GABA(A) receptors. Psychopharmacology 2000, 149, 277–285. [Google Scholar] [CrossRef]
- Lemasson, M.; Delbé, C.; Gheusi, G.; Vincent, J.D.; Lledo, P.M. Use of ultrasonic vocalizations to assess olfactory detection in mouse pups treated with 3-methylindole. Behav. Processes 2005, 68, 13–23. [Google Scholar] [CrossRef]
- Scattoni, M.L.; Martire, A.; Cartocci, G.; Ferrante, A.; Ricceri, L. Reduced social interaction, behavioural flexibility and BDNF signalling in the BTBR T+ tf/J strain, a mouse model of autism. Behav. Brain Res. 2013, 251, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Wöhr, M. Ultrasonic vocalizations in Shank mouse models for autism spectrum disorders: Detailed spectrographic analyses and developmental profiles. Neurosci. Biobehav Rev. 2014, 43, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Caruso, A.; Ricceri, L.; Scattoni, M.L. Ultrasonic vocalizations as a fundamental tool for early and adult behavioral phenotyping of autism spectrum disorder rodent models. Neurosci. Biobehav. Rev. 2020, 116, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Holy, T.E.; Guo, Z. Ultrasonic songs of male mice. PLoS Biol. 2005, 3, e386. [Google Scholar] [CrossRef]
- Nyby, J. Ultrasonic vocalizations during sex behavior of male house mice (Mus musculus): A description. Behav. Neural. Biol. 1983, 39, 128–134. [Google Scholar] [CrossRef]
- Wright, J.M.; Gourdon, J.C.; Clarke, P.B. Identification of multiple call categories within the rich repertoire of adult rat 50-kHz ultrasonic vocalizations: Effects of amphetamine and social context. Psychopharmacology 2010, 211, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mogi, K.; Takakuda, A.; Tsukamoto, C.; Ooyama, R.; Okabe, S.; Koshida, N.; Nagasawa, M.; Kikusui, T. Mutual mother–infant recognition in mice: The role of pup ultrasonic vocalizations. Behav. Brain Res. 2017, 325, 138–146. [Google Scholar] [CrossRef]
- Okabe, S.; Nagasawa, M.; Mogi, K.; Kikusui, T. Importance of mother–infant communication for social bond formation in mammals. Anim. Sci. J. 2012, 83, 446–452. [Google Scholar] [CrossRef]
- Takahashi, T.; Okabe, S.; Broin, P.Ó.; Nishi, A.; Ye, K.; Beckert, M.V.; Izumi, T.; Machida, A.; Kang, G.; Abe, S.; et al. Structure and function of neonatal social communication in a genetic mouse model of autism. Mol. Psychiatry. 2016, 21, 1208–1214. [Google Scholar] [CrossRef] [Green Version]
- Binder, M.S.; Lugo, J.N. NS-Pten knockout mice show sex- and age-specific differences in ultrasonic vocalizations. Brain Behav. 2017, 7, e00857. [Google Scholar] [CrossRef] [Green Version]
- Nolan, S.O.; Hodges, S.L.; Lugo, J.N. High-throughput analysis of vocalizations reveals sex-specific changes in Fmr1 mutant pups. Genes Brain Behav. 2020, 19, e12611. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Categories | Criteria |
|---|---|
| Short | Time < 5 ms and frequency modulation < 6.25 kHz. |
| Flat | Time modulation > 5 ms, and frequency modulation is <6.25 kHz. |
| Upward | Frequency increases in one direction, and modulation is ≥6.25 kHz. |
| Downward | The frequency decreases in one direction, and modulation is ≥6.25 kHz. |
| Modulated | The frequency is modulated in two or more directions, and the modulation is ≥6.25 kHz. |
| Complex | One or more additional frequency components (overtone or non-linear phenomenon, but not concentrated) are added, and the frequency range is not fixed. |
| One frequency jump | There is no time difference between continuous frequencies, one jump in which the frequency changes instantaneously, and no noise. |
| Frequency jumps | There is no time difference between continuous frequencies, multiple jumps in which the frequency changes instantaneously, and no noise. |
| Mixed | A frequency that has one or more jumps and is noisy. |
| Chevron | The frequency decreases by ≥6.25 kHz and increases by ≥12.5 kHz, and is similar to the “inverse U type”. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shiono, S.; Tsutiya, A.; Ohtani-Kaneko, R. Early Detection of Male-Predominant Phenotypes in the Pattern of Ultrasonic Vocalizations Emitted by Autism Spectrum Disorder Model (Crmp4-Knockout) Mice. Brain Sci. 2022, 12, 666. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050666
Shiono S, Tsutiya A, Ohtani-Kaneko R. Early Detection of Male-Predominant Phenotypes in the Pattern of Ultrasonic Vocalizations Emitted by Autism Spectrum Disorder Model (Crmp4-Knockout) Mice. Brain Sciences. 2022; 12(5):666. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050666
Chicago/Turabian StyleShiono, Suzuka, Atsuhiro Tsutiya, and Ritsuko Ohtani-Kaneko. 2022. "Early Detection of Male-Predominant Phenotypes in the Pattern of Ultrasonic Vocalizations Emitted by Autism Spectrum Disorder Model (Crmp4-Knockout) Mice" Brain Sciences 12, no. 5: 666. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050666