
Dopaminergic System in Promoting Recovery from General Anesthesia

Department of Anesthesiology, Beijing Chao-Yang Hospital, Capital Medical University, Beijing 100020, China
*
Author to whom correspondence should be addressed.
Brain Sci. 2023, 13(4), 538; https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci13040538
Submission received: 25 February 2023
/
Revised: 22 March 2023
/
Accepted: 22 March 2023
/
Published: 24 March 2023
(This article belongs to the Special Issue Role of Neuroreceptors in Health and Disease)
Abstract
:Dopamine is an important neurotransmitter that plays a biological role by binding to dopamine receptors. The dopaminergic system regulates neural activities, such as reward and punishment, memory, motor control, emotion, and sleep–wake. Numerous studies have confirmed that the dopaminergic system has the function of maintaining wakefulness in the body. In recent years, there has been increasing evidence that the sleep–wake cycle in the brain has similar neurobrain network mechanisms to those associated with the loss and recovery of consciousness induced by general anesthesia. With the continuous development and innovation of neurobiological techniques, the dopaminergic system has now been proved to be involved in the emergence from general anesthesia through the modulation of neuronal activity. This article is an overview of the dopaminergic system and the research progress into its role in wakefulness and general anesthesia recovery. It provides a theoretical basis for interpreting the mechanisms regulating consciousness during general anesthesia.

1. Introduction
General anesthesia is a reversible state induced by anesthetics, including loss of consciousness, analgesia, and muscle relaxation [1]. The mechanism of general anesthesia has gone through several parts, including lipid bilayer theory, protein target, and specific receptor target [2,3]. Understanding the mechanisms of reversible alterations of consciousness in the organism during general anesthesia has long been a popular area of neuroscience research. Lately, the mechanism of action of the neural circuit in the regulation of consciousness during general anesthesia has received increasing attention [3,4]. There is increasing evidence that the reversible loss and recovery of consciousness under general anesthesia result from the interaction and modulation of many neural circuits [5]. To ensure surgical safety and advance anesthesia science, it is essential to investigate the mechanisms by which general anesthesia induces reversible changes in consciousness.
Dopaminergic neurons are involved in a wide range of functions in the central nervous system (CNS), including movement, memory, arousal, and cognition [6,7,8,9]. Dopamine (DA) is a major neurotransmitter that exerts biological functions by binding to dopamine receptors [10]. In mammals, the substantia nigra pars compacta (SNc) and the ventral tegmental area (VTA) are areas of the brain rich in dopaminergic neurons, both located in the ventral part of the midbrain [11,12]. Activation of dopaminergic nuclei has been shown to promote arousal, including the VTA, ventral periaqueductal grey matter (vPAG), and dorsal raphe nucleus (DRN) [11,13,14]. The study found that dopaminergic neurons in the VTA and SNc project to prominent pro-arousal brain regions, including the thalamus, locus coeruleus, basal forebrain, and laterodorsal tegmental area [11]. These projections suggest that dopaminergic neurons in the VTA and SNc are intimately involved in maintaining wakefulness. As the brain’s arousal system is inhibited in general anesthesia, and the neural activity of general anesthesia resembles that of sleep, it is suggested that reversible loss and recovery of consciousness caused by general anesthesia may have a similar mechanism to the activity changes of sleep–wake-related nuclei [15,16]. In this review, we provide an overview of dopaminergic neurons, the neural projections they are involved in, and their role in wakefulness and general anesthesia recovery.
2. Dopaminergic System
2.1. Dopaminergic Neurons
DA is the most abundant monoamine neurotransmitter in CNS and regulates a diverse range of neural activities in the brain. Dopaminergic neurons catalyze tyrosine uptake into dopa in the cytoplasm via tyrosine hydroxylase (TH) [17]. Dopamine decarboxylase (DDC) then acts on this tyrosine to produce dopamine, which is stored in vesicles [18]. Subsequently, dopaminergic neurons release the vesicles into the synaptic terminal, where DA in the synaptic cleft acts on dopamine receptors in the postsynaptic membrane to exert neurotransmitter effects [19,20]. Dopaminergic neurons are anatomically and functionally heterogeneous cell populations widely distributed in the diencephalon, midbrain, and olfactory bulb. Although dopaminergic neurons make up only about 1% of the total number of neurons in the brain, they play an essential role in the regulation of the basic functions of the brain. Most dopaminergic neuron populations are located in the ventral part of the midbrain. They account for approximately 90% of all dopaminergic neurons in the brain, with various physiological functions to accommodate motor regulation and mental activity [21]. Dopaminergic neurons are engaged in three main types of physiological activity: first, the coordination of extrapyramidal movements, including involvement in the pathogenesis of Parkinson’s disease [22]; second, the regulation of mental activity, including functions, such as learning, memory, cognition, mood, depression and the sleep–wake cycle [9,23,24,25]; and third, the modulation of endocrine system functions, such as tuberoinfundibular dopaminergic neurons that regulate the secretion of prolactin [26].
2.2. Dopaminergic Neuron Projections
The dopaminergic nervous system is divided into the ascending dopaminergic neural projection system, the descending dopaminergic neural projection system, and the local dopaminergic projection system in the aqueduct of Sylvius and the periventricular region [27]. Dopaminergic neurons in the brain are concentrated in the SNc, the VTA, the hypothalamus, and the periventricular nucleus. Projections from midbrain dopaminergic neurons are received and modulated by numerous brain regions. There are four main dopaminergic pathways in the mammalian CNS: the substantia nigra–striatal pathway, the midbrain–limbic system pathway, the mesolimbic–cortical pathway, and the tubero–infundibular pathway. The substantia nigra–striatal pathway, the dopaminergic pathway from the SNc to the striatum, is a crucial component of the basal ganglia, which is primarily implicated in the regulation of movement and whose deterioration causes Parkinson’s syndrome [28]. The midbrain–limbic system pathway, which connects the VTA to the nucleus accumbens (NAc), hippocampus, and amygdala, is associated with reward, reinforcement, and emotion [29]. On the other hand, the mesolimbic–cortical pathway is a projection of cells from the medial VTA to the prefrontal, cingulate, and peripheral cortices, complementing the function of the mesolimbic pathway and contributing to cognition [30]. There is a considerable degree of overlap of dopaminergic neurons between VTA neurons projecting to different targets. Therefore, the two systems involved in the VTA are usually referred to as the mesocortical–limbic system [31]. The nodal funnel bundle arises from the hypothalamus’s arcuate nucleus, projects to the hypothalamus’s midline, and regulates prolactin release [32]. In addition to the four main pathways mentioned above, there is another mesolimbic dopaminergic cluster in the posterior dorsal hypothalamus called area A11 that is involved in perception [33]. There are also some short-range dopamine projections, such as DA amacrine cells (DACs) with long axon-like synapses that extend to the inner nuclear layer, ganglion cell layer, and sometimes to the outer plexiform layer, which overlap and branch to form a dense dendritic network [34]. Depending on their distance and dense network, DACs have the potential to influence activity at different levels in the retina [35].
2.3. Dopamine Receptors
Dopamine receptors (DRs) belong to the G protein-coupled receptor family, have a typical seven-transmembrane structure, signal via a G protein-dependent and independent mechanism, and are widely expressed in the CNS [36]. DA exerts effects by binding to respective membrane receptors. Based on their different binding to ligands and pharmacological properties and signaling pathways, DRs are classified into D1-like and D2-like categories [37]. D1-like DRs contain D1R and D5R, while D2-like DRs contain D2R, D3R, and D4R, of which D1R and D2R are widely distributed in the CNS, whereas the other subtypes are small and restricted in distribution [38,39]. D1-like receptors are found mainly in the caudate nucleus, NAc, substantia nigra pars reticulate, amygdala, frontal cortex, and olfactory bulb [40]. D2-like receptors are mainly expressed in the striatum, VTA, NAc, hypothalamus, hippocampus, and cortex [41]. DA binding to D1-like receptors activates adenylate cyclase, leading to increased cAMP levels and increased cAMP-dependent protein kinase (PKA) activity, resulting in physiological consequences [42]. However, DA binds to D2-like receptors to inhibit adenylate cyclase, decreasing cAMP content [43]. Although cAMP signaling is most relevant to the activation of DRs, some studies have shown that DRs exert biological effects through alternative signaling pathways that are not dependent on cAMP. For example, activation of both D1-like and D2-like receptors can activate protein lipase C-β to induce inositol triphosphate-mediated intracellular calcium flux [44]. Both D1R and D2R trans-activate brain-derived neurotrophic factor receptors in neurons [45]. In addition, DRs regulate the internalization of calcium ions through protein-to-protein interactions [46]. Direct interaction of D1 and D2 receptors with Na+-K+-ATPase has also been demonstrated [47]. Thus, dopamine action depends on the classical adenylate cyclase/cAMP/PKA pathway but is also influenced by second messenger responses, postsynaptic plasma membrane ion channels, and protein expression profiles [36].
3. Dopaminergic System and Arousal
The natural sleep–wake regulation system is composed of several neural clusters that form an interactive neural network. Dopaminergic-related neural circuits play a vital role in natural wakefulness [48]. The ascending dopaminergic system’s SNc, VTA, striatum, and pallidum receive innervation from sleep–wake regulatory nuclei, forming an interacting neural network that regulates wake-related behavior [49,50]. Studies have shown that the release of DA in the cerebral cortex is closely aligned with arousal levels [51]. The string firing pattern of dopaminergic neurons may be relevant to the generation of arousal EEG. Dopaminergic neurons in the SNc and VTA evidenced prominent string firing during arousal [52]. The cell membranes of dopamine neurons contain proteins that facilitate dopamine reuptake, known as dopamine transporters. Treatment with the dopamine transporter inhibitor GBR12909 results in a dose-dependent increase in arousal levels [13]. In addition, damage to dopaminergic neurons can induce a coma-like state [53].
There are numerous arousal-promoting nuclei in the CNS, of which dopaminergic projections from the periaqueductal grey (PAG), VTA, and NAc are closely associated with arousal. The PAG receives input from the ascending spinal pathway, amygdala, and hypothalamus and projects to a wide range of brain regions, including the ventral lateral medulla and hypothalamus [54]. The ventral periaqueductal plenum (vPAG) is associated with sleep, and selective chemical damage to dopaminergic neurons in the vPAG significantly increases sleep duration [13]. Chemogenetic activation of dopaminergic neurons of locus coeruleus-vPAG or direct activation of vPAG-DA neurons promotes arousal [55]. The role of vPAG-DA neurons in sleep–wake is associated with their projections to multiple pro-arousal brain regions, including the prefrontal cortex (PFC), ventral lateral preoptic area, and locus coeruleus [13,56,57]. The vPAG-DA neurons coordinate orexinergic, cholinergic, GABAergic, and noradrenergic neurons to impact sleep–wake processes. [13,58]. The VTA is a major concentration of DA neurons, and multiple studies have confirmed its role in promoting arousal. EEG spectral analysis revealed that activation of the VTA-DA to the prelimbic cortex (PrL) pathway during sevoflurane anesthesia decreased sleep-related low-frequency activity and increased wake-related high-frequency activity [59]. In addition, extracellular DA levels in the prefrontal cortex were increased, and arousal was enhanced by injecting orexin-A into the VTA [60]. NAc, a critical brain region for the projection of dopaminergic nuclei, is also implicated in sleep–wake regulation. A recent study showed that optogenetics activation of the neural circuits projected by VTA dopaminergic neurons to NAc promoted arousal in mice [61]. Extracellular DA levels in the NAc decrease during non-rapid eye movement sleep and increase significantly during wakefulness and rapid eye movement sleep [62]. The study found that the DRs of the NAc are involved in regulating the sleep arousal mechanism [63]. Moreover, age-related downregulation of D1R in the NAc shell results in decreased function of D1R in promoting arousal [64]. The ventral pallidum (VP) is a major component of the basal ganglia, which receives intensive input from the NAc and a large number of VTA dopaminergic projections [65,66]. Studies have found that NAc D1R neurons regulate wakefulness, while NAc D2R neurons projecting to the VP regulate sleep [67,68]. It was shown that in vivo stimulation of VP-GABAergic neurons increased VTA-dopaminergic neuronal activity, which in turn induced arousal, whereas pretreatment with dopaminergic antagonists completely blocked arousal induced by VP-GABAergic neuronal activation [65]. This suggests that VTA-dopaminergic neurons mediate arousal through activation elicited by VP-GABAergic neurons.
4. Dopaminergic System for General Anesthesia Recovery
General anesthesia is widespread in clinical practice, but the mechanisms by which general anesthesia causes a reversible loss and recovery of consciousness still need to be well elucidated. It is generally accepted that the CNS is interconnected by neuronal projections between nuclei forming a complex and diverse neural network achieved by the interaction of multiple neurotransmitters and neuromodulators [69,70]. Dopaminergic neurons are integral to neural network regulation and perform critical functions in cognition, memory, sleep, and general anesthesia [7,9,11,71]. Identifying the mechanisms and patterns of emergence regulation in anesthesia is central to improving the quality of recovery from general anesthesia. A growing number of studies have documented that central dopaminergic system activation is associated with the emergence of general anesthesia [59,72]. Current research into the dopaminergic system regulating consciousness in general anesthesia has focused on the dopaminergic nuclei, associated neural circuits, and dopamine receptors (Table 1).
4.1. Nervous Nuclei and General Anesthesia
4.1.1. VTA
Dopaminergic neurons in the VTA are thought to be essential in altering consciousness induced by general anesthesia. As an integral part of the upstream activation system, projections from the VTA to the basal forebrain, NAc, and prefrontal cortex contribute to cortical and behavioral activation [73]. Bilateral VTA lesions significantly prolong the emergence time in propofol anesthesia but exert no impact on induction time. However, isoflurane and ketamine anesthesia are insensitive to VTA lesions [85]. This may be due to different mechanisms of action between the different anesthetic drugs. Dopamine transporter (DAT) is a transmembrane protein that is specifically distributed on the surface of DA neurons and regulates extracellular dopamine concentrations by transporting extracellular DA into the cell [74]. A recent study revealed that DAT inhibition in the VTA enhances PFC neuronal activity, thereby facilitating emergence in rats under propofol anesthesia [75]. Furthermore, electrical stimulation of VTA-DA neurons induced rapid arousal under isoflurane or propofol anesthesia. Consciousness and EEG change consistent with arousal during general anesthesia were restored with electrical stimulation of the VTA brain region but not observed with SNc stimulation [76]. Optogenetic manipulation of the VTA-DA under isoflurane anesthesia has been reported to reduce the time to wakefulness [77]. Li et al. found that microinjection of orexin-A into the VTA of rats reduced the burst suppression ratio during the maintenance of isoflurane anesthesia and promoted emergence from isoflurane anesthesia. In isolated brain slices, orexin-A also directly increased action potential frequency of dopaminergic neurons in VTA with 1 MAC isoflurane, implying that projections from orexinergic neurons to VTA-DA neurons may be related to the emergence of general anesthesia by affecting the neuronal activity of DA neurons [86]. Selective activation of VTA DA neurons during sevoflurane anesthesia leads to prolonged induction and shortened recovery time associated with sevoflurane anesthesia [62]. These results suggest that VTA-DA neurons are involved in the induction and emergence of general anesthesia and that there are differences in the alterations in consciousness under different anesthetics. Further research is necessary to elucidate the mechanisms of anesthetics in different states of anesthesia.
4.1.2. NAc
The NAc is a significant component of the ventral striatum and is constituted by medium-spiny GABAergic neurons [87]. Medium spiny neurons (MSNs) in the NAc shell receive projections primarily from dopaminergic neurons in the VTA, and the core of the NAc receives projections primarily from the substantia nigra [88]. Recent studies have demonstrated that the dopaminergic system is associated with the modulation of consciousness in the NAc during general anesthesia. Using the microdialysis technique to observe changes in DA concentrations in the NAc of rats under propofol anesthesia, it was found that low doses (9 mg/kg) decreased DA concentrations, while medium doses (60 mg/kg) and high doses (100 mg/kg) increased DA concentrations. This suggests that there may be variations in the activity of DA neurons in NAc at different anesthetic doses. [89]. Gui et al. reported that DA in the NAc diminished with the loss of consciousness induced by sevoflurane anesthesia, remained stable during anesthesia, and increased in the transition from anesthesia to arousal [62]. NAc is a potential target for the involvement of the dopaminergic system in the regulation of consciousness by general anesthesia due to its high expression of D1R and D2R, as well as its complex fiber connections with cortical and midbrain dopaminergic nuclei [78]. Fiber-optic recording of changes in calcium-signaling NAc activity during propofol anesthesia revealed that the neuronal excitation of rat NAc neurons decreased during the induction and restored during anesthesia emergence [72]. Other results using the same fiber-optic calcium-sensing technology showed reduced neuron activity in NAc expressing D1R during induction and maintenance of sevoflurane and enhanced neuron activity during wakefulness. NAc-D1R neurons induce cortical activation when activated by optogenetic stimulation, implicating NAc-D1R neurons as promising targets for the modulation of consciousness [90]. By studying specific subtypes of neurons in more detail, it is possible to identify the specific mechanisms that influence general anesthesia arousal, which also provides a new direction for future mechanistic research.
4.1.3. vPAG
The vPAG-DA is concerned with regulating consciousness by general anesthesia through projections to several nuclei, notably the basal forebrain, locus coeruleus, and ventral lateral preoptic area [91,92,93]; 6-OHDA is a potent dopamine neurotoxicant extensively applied as a selective catecholaminergic neurotoxic agent in cellular or animal models of Parkinson’s disease [79,94]. Damaging dopaminergic neurons in vPAG by 6-OHDA increased EEG δ wave, shortened induction time, and prolonged recovery time of propofol anesthesia, indicating that vPAG-DA neurons are related to the anesthetic mechanism of propofol [80]. In addition, propofol increased the sensitivity of postsynaptic GABAA receptors and the release of the presynaptic inhibitory neurotransmitter GABA in vPAG-DA neurons, ultimately inhibiting the activity of vPAG dopamine neurons. Microinjection of a GABAAR antagonist abolished the suppressive efficacy on vPAG-DA neurons, suggesting that GABAA receptors are engaged in isoflurane anesthesia through the regulation of vPAG-DA [80]. Liu et al. discovered that isoflurane anesthesia caused a decrease in DA neuronal excitation in the vPAG. In contrast, the activity of the whole population of vPAG neurons increased dramatically during the emergence of isoflurane anesthesia. The induction time is shortened, and the recovery time is lengthened by unilateral ablation of vPAG-DA neurons. Furthermore, isoflurane significantly increased presynaptic inhibitory neurotransmitter GABA release from vPAG-DA neurons by increasing spontaneous inhibitory postsynaptic current (sIPSC) frequency and decay time [71].
4.2. Neural Circuit and General Anesthesia
The dopaminergic system in general anesthesia not only influences the anesthetic state of the body by altering DA levels in the corresponding nuclei but also participates in the regulation of consciousness through neurocircuitry (Figure 1). Orexin injections into the VTA were found to elevate extracellular DA levels in the PFC and prolong wakefulness [60]. These findings suggest that there are dopaminergic projections in the VTA-PFC and that the projections of orexinergic neurons in the VTA may correlate with levels of DA release in the PFC and are involved in the effects on consciousness. In general anesthesia, this has been well demonstrated. Consciousness modulation by sevoflurane anesthesia is associated with dopaminergic projections from VTA to PrL, part of the PFC. Chemogenetics and optogenetics suggest that the VTA-PrL dopaminergic pathway participates in the mechanism of sevoflurane anesthesia and that activation of this pathway prolongs the induction of anesthesia and enhances anesthesia emergence [59]. Using DA neurotransmitter probes to record changes in DA in the neural nuclei, dexmedetomidine was found to increase DA concentrations in the medial prefrontal cortex and NAc in the dopaminergic neuronal projection area through activation of VTA dopaminergic neurons, which explains the rapid awakening after dexmedetomidine sedation and reflects the vital role of the VTA-NAc/mPFC dopaminergic pathway in promoting awakening during general anesthesia [81]. In addition, Gui et al. found that stimulation of the VTA-NAc dopaminergic pathway extended the induction of sevoflurane anesthesia in mice and at the same time shortened the wake time, accompanied by a decrease in δ waves and an increase in γ waves, whereas inhibition of the pathway had the opposite effect [62]. Activation of orexin neural projection from the periaqueductal lateral hypothalamic area (PeFLH) to the VTA promotes emergence from isoflurane anesthesia by modulation of dopaminergic neurons [86]. The dopaminergic system has been identified as an eminent contributor to the arousal-promoting properties of general anesthesia. As research continues, it is becoming clear that the regulation of consciousness during general anesthesia is achieved through the interaction and co-regulation of multiple types of neurons.
4.3. Dopamine Receptor and General Anesthesia
Depending on the type of neuronal terminals they receive and the type of neurons to which they project, dopamine receptor neurons play different roles in regulating arousal in different regions of the brain. Over the past years, dopamine receptors have been implicated in the regulation of consciousness during general anesthesia. Systemic administration of D1R agonists or methyl benzene significantly accelerated recovery in isoflurane-anesthetized rats [82,95]. The D1R agonist choro-APB shortened the time to wakefulness after isoflurane general anesthesia, and the D1R antagonist SCH-23390 inhibited this effect. In contrast, awakening after isoflurane general anesthesia was not induced by the D2R agonist quinpirole [95]. D-amphetamine reversed dexmedetomidine-induced unconsciousness by promoting dopamine release and activating D1 and D5 receptors in the brain. Furthermore, D-amphetamine induced recovery from propofol and sevoflurane anesthesia. This effect is not seen in ketamine anesthesia, implying that ketamine may have a different pharmacological mechanism than other general anesthetics [83,84]. These studies are consistent with the involvement of expressing DR neurons in recovering consciousness after anesthesia.
In the course of the research, DRs have been targeted in specific nuclei, and it has been confirmed that dopamine receptors in various nuclei engage in the mechanism of action of general anesthesia. Optogenetic activation of the VTA-DA under isoflurane anesthesia has been reported to shorten the time to wakefulness, which can be inhibited by intraperitoneal injection of D1R blockers [77]. To accurately determine the role of DRs during general anesthesia, DR agonists or antagonists are used on specific nerve clusters. Both intraperitoneal injection and PrL microinjection of D1R agonist Choro-APB promoted recovery from sevoflurane anesthesia, while SCH23390, a D1R antagonist, enhanced anesthesia, suggesting the D1R in PrL is an essential regulatory site of the dopaminergic system in sevoflurane anesthesia [59]. NAc dopaminergic receptors exert maintaining arousal. Systemic activation of D1R, but not D2R, shortens the recovery time from isoflurane anesthesia, suggesting that D1R mediates arousal in general anesthesia [68,95]. Moreover, microinjection of D1R agonists into the NAc shell accelerated recovery from isoflurane anesthesia in mice, whereas D1R antagonists delayed wakefulness [64]. Zhang et al. found that D1R receptor regulation of MSNs activity of NAc is essential for the recovery of propofol-induced loss of consciousness. D1R is involved in the increased frequency and prolonged decay time of sIPSCs and miniature inhibitory postsynaptic current (mIPSCs) of the MSNs induced by propofol anesthesia [72]. Optogenetics activation of D1R neurons in NAc induces cortical activation and recovery during sevoflurane anesthesia, which is also reflected in burst suppression oscillations induced by deep sevoflurane anesthesia [90]. Aging may attenuate anesthetic arousal by downregulating D1R expression in the NAc shell [64]. Therefore, changes in the dopaminergic system in response to age-related factors, including DA neuronal damage, reduced DA synthesis, and downregulation of DR, all influence the regulation of consciousness by general anesthesia. In addition, the olfactory nodule (OT) is intensively innervated by dopaminergic neurons of VTA, and its main neuronal components are neurons expressing D1R and D2R. Injection of D1R or D2R agonists into the OT significantly accelerates isoflurane-induced wakefulness, whereas D1R or D2R antagonists significantly delay wakefulness [96].
5. Conclusions
In summary, DA neurons are involved in the regulation of both wakefulness and recovery from general anesthesia. The dopaminergic system plays a vital role in the regulation of consciousness during general anesthesia. It is involved in the mechanism of action through nuclei, associated neural circuits, and dopamine receptors. Nevertheless, the mechanisms underlying the regulation of consciousness by general anesthetics remain poorly understood. With the development and innovation of scientific techniques, such as optogenetics, chemogenetics, neurotransmitter probes, electroencephalography, and so on, it is possible to explore in greater depth the mechanisms by which general anesthesia regulates consciousness. These techniques allow us to explore the dynamics of neurotransmitters and neuropeptides in general anesthesia and how they are involved in the altered consciousness of general anesthesia, the nuclei and their interactions in general anesthesia, and even provide a clearer understanding of the specific pharmacological effects of different general anesthetics. Dopaminergic neurons are involved in facilitating recovery of consciousness from general anesthesia, and it is expected that improvements in the clinical problem of delayed awakening will be achieved. In this review, dopaminergic neurons and their role in arousal and recovery under general anesthesia are summarized, providing a theoretical basis for the pharmacological mechanism of general anesthetics in regulating consciousness.
Author Contributions
Conceptualization, J.W., A.W. and C.W.; data curation, X.M., Y.S. and S.L.; writing-original draft, J.W.; writing-review and editing, J.W., X.M., Y.S., S.L., A.W. and C.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research no external funding.
Institutional Review Board Statement
No applicable.
Informed Consent Statement
No applicable.
Data Availability Statement
No applicable.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Brown, E.N.; Purdon, P.L.; Van Dort, C.J. General Anesthesia and Altered States of Arousal: A Systems Neuroscience Analysis. Annu. Rev. Neurosci. 2011, 34, 601–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antkowiak, B. How Do General Anaesthetics Work? Naturwissenschaften 2001, 88, 201–213. [Google Scholar] [CrossRef]
- Franks, N.P. General Anaesthesia: From Molecular Targets to Neuronal Pathways of Sleep and Arousal. Nat. Rev. Neurosci. 2008, 9, 370–386. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Pan, J.; Yu, Y. Regulation of Neural Circuitry under General Anesthesia: New Methods and Findings. Biomolecules 2022, 12, 898. [Google Scholar] [CrossRef] [PubMed]
- Moody, O.A.; Zhang, E.R.; Vincent, K.F.; Kato, R.; Melonakos, E.D.; Nehs, C.J.; Solt, K. The Neural Circuits Underlying General Anesthesia and Sleep. Anesth. Analg. 2021, 132, 1254–1264. [Google Scholar] [CrossRef]
- Pérez-Fernández, J.; Barandela, M.; Jiménez-López, C. The Dopaminergic Control of Movement-Evolutionary Considerations. Int. J. Mol. Sci. 2021, 22, 11284. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Liang, J.; Wang, R.; Yan, T.; Zhou, Y.; Liu, Y.; Feng, Q.; Sun, F.; Li, Y.; Li, A.; et al. The Raphe Dopamine System Controls the Expression of Incentive Memory. Neuron 2020, 106, 498–514.e498. [Google Scholar] [CrossRef]
- Dong, H.; Wang, J.; Yang, Y.F.; Shen, Y.; Qu, W.M.; Huang, Z.L. Dorsal Striatum Dopamine Levels Fluctuate across the Sleep-Wake Cycle and Respond to Salient Stimuli in Mice. Front. Neurosci. 2019, 13, 242. [Google Scholar] [CrossRef] [PubMed]
- Engelhard, B.; Finkelstein, J.; Cox, J.; Fleming, W.; Jang, H.J.; Ornelas, S.; Koay, S.A.; Thiberge, S.Y.; Daw, N.D.; Tank, D.W.; et al. Specialized Coding of Sensory, Motor and Cognitive Variables in Vta Dopamine Neurons. Nature 2019, 570, 509–513. [Google Scholar] [CrossRef]
- Berke, J.D. What Does Dopamine Mean? Nat. Neurosci. 2018, 21, 787–793. [Google Scholar] [CrossRef]
- Monti, J.M.; Monti, D. The Involvement of Dopamine in the Modulation of Sleep and Waking. Sleep Med. Rev. 2007, 11, 113–133. [Google Scholar] [CrossRef] [PubMed]
- Islam, K.U.S.; Meli, N.; Blaess, S. The Development of the Mesoprefrontal Dopaminergic System in Health and Disease. Front. Neural Circuits 2021, 15, 746582. [Google Scholar] [CrossRef]
- Lu, J.; Jhou, T.C.; Saper, C.B. Identification of Wake-Active Dopaminergic Neurons in the Ventral Periaqueductal Gray Matter. J. Neurosci. 2006, 26, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Monti, J.M. The Neurotransmitters of Sleep and Wake, a Physiological Reviews Series. Sleep Med. Rev. 2013, 17, 313–315. [Google Scholar] [CrossRef]
- Leung, L.S.; Luo, T.; Ma, J.; Herrick, I. Brain Areas That Influence General Anesthesia. Prog. Neurobiol. 2014, 122, 24–44. [Google Scholar] [CrossRef]
- Wang, D.; Guo, Q.; Zhou, Y.; Xu, Z.; Hu, S.W.; Kong, X.X.; Yu, Y.M.; Yang, J.X.; Zhang, H.; Ding, H.L.; et al. Gabaergic Neurons in the Dorsal-Intermediate Lateral Septum Regulate Sleep-Wakefulness and Anesthesia in Mice. Anesthesiology 2021, 135, 463–481. [Google Scholar] [CrossRef] [PubMed]
- Daubner, S.C.; Le, T.; Wang, S. Tyrosine Hydroxylase and Regulation of Dopamine Synthesis. Arch. Biochem. Biophys. 2011, 508, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Christenson, J.G.; Dairman, W.; Udenfriend, S. Preparation and Properties of a Homogeneous Aromatic L-Amino Acid Decarboxylase from Hog Kidney. Arch. Biochem. Biophys. 1970, 141, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Garris, P.A.; Ciolkowski, E.L.; Pastore, P.; Wightman, R.M. Efflux of Dopamine from the Synaptic Cleft in the Nucleus Accumbens of the Rat Brain. J. Neurosci. 1994, 14, 6084–6093. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, J.; Jørgensen, T.N.; Gether, U. Regulation of Dopamine Transporter Function by Protein-Protein Interactions: New Discoveries and Methodological Challenges. J. Neurochem. 2010, 113, 27–41. [Google Scholar] [CrossRef]
- Chinta, S.J.; Andersen, J.K. Dopaminergic Neurons. Int. J. Biochem. Cell Biol. 2005, 37, 942–946. [Google Scholar] [CrossRef]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson Disease. Nat. Rev. Dis. Prim. 2017, 3, 17013. [Google Scholar] [CrossRef] [PubMed]
- Kempadoo, K.A.; Mosharov, E.V.; Choi, S.J.; Sulzer, D.; Kandel, E.R. Dopamine Release from the Locus Coeruleus to the Dorsal Hippocampus Promotes Spatial Learning and Memory. Proc. Natl. Acad. Sci. USA 2016, 113, 14835–14840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Condon, M.D.; Platt, N.J.; Zhang, Y.F.; Roberts, B.M.; Clements, M.A.; Vietti-Michelina, S.; Tseu, M.Y.; Brimblecombe, K.R.; Threlfell, S.; Mann, E.O.; et al. Plasticity in Striatal Dopamine Release Is Governed by Release-Independent Depression and the Dopamine Transporter. Nat. Commun. 2019, 10, 4263. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.R.; Treweek, J.B.; Robinson, J.E.; Xiao, C.; Bremner, L.R.; Greenbaum, A.; Gradinaru, V. Dorsal Raphe Dopamine Neurons Modulate Arousal and Promote Wakefulness by Salient Stimuli. Neuron 2017, 94, 1205–1219.e1208. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wei, J.; Ma, F.; Shan, Q.; Gao, D.; Jin, Y.; Sun, P. Melatonin Modulates Lactation by Regulating Prolactin Secretion Via Tuberoinfundibular Dopaminergic Neurons in the Hypothalamus- Pituitary System. Curr. Protein Pept. Sci. 2020, 21, 744–750. [Google Scholar] [CrossRef] [PubMed]
- Björklund, A.; Dunnett, S.B. Dopamine Neuron Systems in the Brain: An Update. Trends Neurosci. 2007, 30, 194–202. [Google Scholar] [CrossRef]
- Liu, Y.L.; Qin, L.; Wilson, B.; Wu, X.; Qian, L.; Granholm, A.C.; Crews, F.T.; Hong, J.S. Endotoxin Induces a Delayed Loss of Th-Ir Neurons in Substantia Nigra and Motor Behavioral Deficits. Neurotoxicology 2008, 29, 864–870. [Google Scholar] [CrossRef] [Green Version]
- Fields, H.L.; Hjelmstad, G.O.; Margolis, E.B.; Nicola, S.M. Ventral Tegmental Area Neurons in Learned Appetitive Behavior and Positive Reinforcement. Annu. Rev. Neurosci. 2007, 30, 289–316. [Google Scholar] [CrossRef] [Green Version]
- Obeso, J.A.; Marin, C.; Rodriguez-Oroz, C.; Blesa, J.; Benitez-Temiño, B.; Mena-Segovia, J.; Rodríguez, M.; Olanow, C.W. The Basal Ganglia in Parkinson’s Disease: Current Concepts and Unexplained Observations. Ann. Neurol. 2008, 64 (Suppl. S2), S30–S46. [Google Scholar] [CrossRef]
- Wise, R.A. Dopamine, Learning and Motivation. Nat. Rev. Neurosci. 2004, 5, 483–494. [Google Scholar] [CrossRef]
- Smargiassi, A.; Mutti, A. Peripheral Biomarkers and Exposure to Manganese. Neurotoxicology 1999, 20, 401–406. [Google Scholar]
- Klein, M.O.; Battagello, D.S.; Cardoso, A.R.; Hauser, D.N.; Bittencourt, J.C.; Correa, R.G. Dopamine: Functions, Signaling, and Association with Neurological Diseases. Cell Mol. Neurobiol. 2019, 39, 31–59. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Field, G.D. Dopaminergic Modulation of Retinal Processing from Starlight to Sunlight. J. Pharmacol. Sci. 2019, 140, 86–93. [Google Scholar] [CrossRef]
- Indrieri, A.; Pizzarelli, R.; Franco, B.; De Leonibus, E. Dopamine, Alpha-Synuclein, and Mitochondrial Dysfunctions in Parkinsonian Eyes. Front. Neurosci. 2020, 14, 567129. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, J.M.; Espinoza, S.; Gainetdinov, R.R. Dopamine Receptors—Iuphar Review 13. Br. J. Pharmacol. 2015, 172, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Kato, M.; Takano, H.; Arakawa, R.; Okumura, M.; Otsuka, T.; Kodaka, F.; Hayashi, M.; Okubo, Y.; Ito, H.; et al. Differential Contributions of Prefrontal and Hippocampal Dopamine D(1) and D(2) Receptors in Human Cognitive Functions. J. Neurosci. 2008, 28, 12032–12038. [Google Scholar] [CrossRef] [Green Version]
- Gingrich, J.A.; Caron, M.G. Recent Advances in the Molecular Biology of Dopamine Receptors. Annu. Rev. Neurosci. 1993, 16, 299–321. [Google Scholar] [CrossRef]
- Gurevich, E.V.; Gainetdinov, R.R.; Gurevich, V.V. G Protein-Coupled Receptor Kinases as Regulators of Dopamine Receptor Functions. Pharmacol. Res. 2016, 111, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyson, S.J.; McGonigle, P.; Molinoff, P.B. Quantitative Autoradiographic Localization of the D1 and D2 Subtypes of Dopamine Receptors in Rat Brain. J. Neurosci. 1986, 6, 3177–3188. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, C.; Okamura, H.; Nakajima, T.; Taguchi, J.; Ibata, Y. Autoradiographic Distribution of [3h]Ym-09151-2, a High-Affinity and Selective Antagonist Ligand for the Dopamine D2 Receptor Group, in the Rat Brain and Spinal Cord. J. Comp. Neurol. 1994, 344, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Boyd, K.N.; Mailman, R.B. Dopamine Receptor Signaling and Current and Future Antipsychotic Drugs. Handb. Exp. Pharmacol. 2012, 212, 53–86. [Google Scholar]
- Albert, P.R.; Neve, K.A.; Bunzow, J.R.; Civelli, O. Coupling of a Cloned Rat Dopamine-D2 Receptor to Inhibition of Adenylyl Cyclase and Prolactin Secretion. J. Biol. Chem. 1990, 265, 2098–2104. [Google Scholar] [CrossRef] [PubMed]
- Channer, B.; Matt, S.M.; Nickoloff-Bybel, E.A.; Pappa, V.; Agarwal, Y.; Wickman, J.; Gaskill, P.J. Dopamine, Immunity, and Disease. Pharmacol. Rev. 2023, 75, 62–158. [Google Scholar] [CrossRef] [PubMed]
- Swift, J.L.; Godin, A.G.; Doré, K.; Freland, L.; Bouchard, N.; Nimmo, C.; Sergeev, M.; De Koninck, Y.; Wiseman, P.W.; Beaulieu, J.M. Quantification of Receptor Tyrosine Kinase Transactivation through Direct Dimerization and Surface Density Measurements in Single Cells. Proc. Natl. Acad. Sci. USA 2011, 108, 7016–7021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kisilevsky, A.E.; Zamponi, G.W. D2 Dopamine Receptors Interact Directly with N-Type Calcium Channels and Regulate Channel Surface Expression Levels. Channels 2008, 2, 269–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazelwood, L.A.; Free, R.B.; Cabrera, D.M.; Skinbjerg, M.; Sibley, D.R. Reciprocal Modulation of Function between the D1 and D2 Dopamine Receptors and the Na+,K+-Atpase. J. Biol. Chem. 2008, 283, 36441–36453. [Google Scholar] [CrossRef] [Green Version]
- Noseda, R.; Borsook, D.; Burstein, R. Neuropeptides and Neurotransmitters That Modulate Thalamo-Cortical Pathways Relevant to Migraine Headache. Headache 2017, 57 (Suppl. 2), 97–111. [Google Scholar] [CrossRef] [Green Version]
- Yanagihara, S.; Ikebuchi, M.; Mori, C.; Tachibana, R.O.; Okanoya, K. Arousal State-Dependent Alterations in Neural Activity in the Zebra Finch Vta/Snc. Front. Neurosci. 2020, 14, 897. [Google Scholar] [CrossRef]
- Qiu, M.H.; Zhong, Z.G.; Chen, M.C.; Lu, J. Nigrostriatal and Mesolimbic Control of Sleep-Wake Behavior in Rat. Brain. Struct. Funct. 2019, 224, 2525–2535. [Google Scholar] [CrossRef]
- Wisor, J.P.; Nishino, S.; Sora, I.; Uhl, G.H.; Mignot, E.; Edgar, D.M. Dopaminergic Role in Stimulant-Induced Wakefulness. J. Neurosci. 2001, 21, 1787–1794. [Google Scholar] [CrossRef]
- Rye, D.B.; Jankovic, J. Emerging Views of Dopamine in Modulating Sleep/Wake State from an Unlikely Source: Pd. Neurology 2002, 58, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.E.; Bobillier, P.; Pin, C.; Jouvet, M. The Effect of Lesions of Catecholamine-Containing Neurons Upon Monoamine Content of the Brain and Eeg and Behavioral Waking in the Cat. Brain Res. 1973, 58, 157–177. [Google Scholar] [CrossRef] [PubMed]
- Behbehani, M.M. Functional Characteristics of the Midbrain Periaqueductal Gray. Prog. Neurobiol. 1995, 46, 575–605. [Google Scholar] [CrossRef] [PubMed]
- Porter-Stransky, K.A.; Centanni, S.W.; Karne, S.L.; Odil, L.M.; Fekir, S.; Wong, J.C.; Jerome, C.; Mitchell, H.A.; Escayg, A.; Pedersen, N.P.; et al. Noradrenergic Transmission at Alpha1-Adrenergic Receptors in the Ventral Periaqueductal Gray Modulates Arousal. Biol. Psychiatry 2019, 85, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C.; Bjorkum, A.A.; Gaus, S.E.; Lu, J.; Scammell, T.E.; Saper, C.B. Afferents to the Ventrolateral Preoptic Nucleus. J. Neurosci. 2002, 22, 977–990. [Google Scholar] [CrossRef] [Green Version]
- Verret, L.; Fort, P.; Gervasoni, D.; Léger, L.; Luppi, P.H. Localization of the Neurons Active During Paradoxical (Rem) Sleep and Projecting to the Locus Coeruleus Noradrenergic Neurons in the Rat. J. Comp. Neurol. 2006, 495, 573–586. [Google Scholar] [CrossRef]
- Omelchenko, N.; Sesack, S.R. Periaqueductal Gray Afferents Synapse onto Dopamine and Gaba Neurons in the Rat Ventral Tegmental Area. J. Neurosci. Res. 2010, 88, 981–991. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Chu, R.; Cao, F.; Wang, Y.; Liu, Y.; Cao, J.; Guo, Y.; Mi, W.; Tong, L. Dopaminergic Neurons in the Ventral Tegmental-Prelimbic Pathway Promote the Emergence of Rats from Sevoflurane Anesthesia. Neurosci. Bull. 2022, 38, 417–428. [Google Scholar] [CrossRef]
- Vittoz, N.M.; Berridge, C.W. Hypocretin/Orexin Selectively Increases Dopamine Efflux within the Prefrontal Cortex: Involvement of the Ventral Tegmental Area. Neuropsychopharmacology 2006, 31, 384–395. [Google Scholar] [CrossRef] [Green Version]
- Eban-Rothschild, A.; Rothschild, G.; Giardino, W.J.; Jones, J.R.; de Lecea, L. Vta Dopaminergic Neurons Regulate Ethologically Relevant Sleep-Wake Behaviors. Nat. Neurosci. 2016, 19, 1356–1366. [Google Scholar] [CrossRef] [PubMed]
- Gui, H.; Liu, C.; He, H.; Zhang, J.; Chen, H.; Zhang, Y. Dopaminergic Projections from the Ventral Tegmental Area to the Nucleus Accumbens Modulate Sevoflurane Anesthesia in Mice. Front. Cell Neurosci. 2021, 15, 671473. [Google Scholar] [CrossRef] [PubMed]
- Monti, J.M.; Jantos, H. The Effects of Local Microinjection of Selective Dopamine D1 and D2 Receptor Agonists and Antagonists into the Dorsal Raphe Nucleus on Sleep and Wakefulness in the Rat. Behav. Brain Res. 2018, 339, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gui, H.; Hu, L.; Li, C.; Zhang, J.; Liang, X. Dopamine D1 Receptor in the Nac Shell Is Involved in Delayed Emergence from Isoflurane Anesthesia in Aged Mice. Brain Behav. 2021, 11, e01913. [Google Scholar] [CrossRef]
- Li, Y.D.; Luo, Y.J.; Xu, W.; Ge, J.; Cherasse, Y.; Wang, Y.Q.; Lazarus, M.; Qu, W.M.; Huang, Z.L. Ventral Pallidal Gabaergic Neurons Control Wakefulness Associated with Motivation through the Ventral Tegmental Pathway. Mol. Psychiatry 2021, 26, 2912–2928. [Google Scholar] [CrossRef]
- Clark, M.; Bracci, E. Dichotomous Dopaminergic Control of Ventral Pallidum Neurons. Front. Cell Neurosci. 2018, 12, 260. [Google Scholar] [CrossRef]
- Oishi, Y.; Xu, Q.; Wang, L.; Zhang, B.J.; Takahashi, K.; Takata, Y.; Luo, Y.J.; Cherasse, Y.; Schiffmann, S.N.; de Kerchove, A.; et al. Slow-Wave Sleep Is Controlled by a Subset of Nucleus Accumbens Core Neurons in Mice. Nat. Commun. 2017, 8, 734. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.J.; Li, Y.D.; Wang, L.; Yang, S.R.; Yuan, X.S.; Wang, J.; Cherasse, Y.; Lazarus, M.; Chen, J.F.; Qu, W.M.; et al. Nucleus Accumbens Controls Wakefulness by a Subpopulation of Neurons Expressing Dopamine D(1) Receptors. Nat. Commun. 2018, 9, 1576. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Liu, T.; Dong, Y.; Kondoh, K.; Lu, Z. Trans-Synaptic Neural Circuit-Tracing with Neurotropic Viruses. Neurosci. Bull. 2019, 35, 909–920. [Google Scholar] [CrossRef]
- Li, J.; Liu, Y.; Yuan, L.; Zhang, B.; Bishop, E.S.; Wang, K.; Tang, J.; Zheng, Y.Q.; Xu, W.; Niu, S.; et al. A Tissue-Like Neurotransmitter Sensor for the Brain and Gut. Nature 2022, 606, 94–101. [Google Scholar] [CrossRef]
- Liu, C.; Zhou, X.; Zhu, Q.; Fu, B.; Cao, S.; Zhang, Y.; Zhang, L.; Zhang, Y.; Yu, T. Dopamine Neurons in the Ventral Periaqueductal Gray Modulate Isoflurane Anesthesia in Rats. CNS Neurosci. Ther. 2020, 26, 1121–1133. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gui, H.; Duan, Z.; Yu, T.; Zhang, J.; Liang, X.; Liu, C. Dopamine D1 Receptor in the Nucleus Accumbens Modulates the Emergence from Propofol Anesthesia in Rat. Neurochem. Res. 2021, 46, 1435–1446. [Google Scholar] [CrossRef] [PubMed]
- Swanson, L.W. The Projections of the Ventral Tegmental Area Adjacent Regions: A Combined Fluorescent Retrograde Tracer and Immunofluorescence Study in the Rat. Brain Res. Bull. 1982, 9, 321–353. [Google Scholar] [CrossRef]
- Efimova, E.V.; Gainetdinov, R.R.; Budygin, E.A.; Sotnikova, T.D. Dopamine Transporter Mutant Animals: A Translational Perspective. J. Neurogenet. 2016, 30, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Xu, K.; Yin, J.W.; Zhang, H.; Yin, J.T.; Li, Y. Dopamine Transporter in the Ventral Tegmental Area Modulates Recovery from Propofol Anesthesia in Rats. J. Chem. Neuroanat. 2022, 121, 102083. [Google Scholar] [CrossRef] [PubMed]
- Solt, K.; Van Dort, C.J.; Chemali, J.J.; Taylor, N.E.; Kenny, J.D.; Brown, E.N. Electrical Stimulation of the Ventral Tegmental Area Induces Reanimation from General Anesthesia. Anesthesiology 2014, 121, 311–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, N.E.; Van Dort, C.J.; Kenny, J.D.; Pei, J.; Guidera, J.A.; Vlasov, K.Y.; Lee, J.T.; Boyden, E.S.; Brown, E.N.; Solt, K. Optogenetic Activation of Dopamine Neurons in the Ventral Tegmental Area Induces Reanimation from General Anesthesia. Proc. Natl. Acad. Sci. USA 2016, 113, 12826–12831. [Google Scholar] [CrossRef] [Green Version]
- Soares-Cunha, C.; Coimbra, B.; David-Pereira, A.; Borges, S.; Pinto, L.; Costa, P.; Sousa, N.; Rodrigues, A.J. Activation of D2 Dopamine Receptor-Expressing Neurons in the Nucleus Accumbens Increases Motivation. Nat. Commun. 2016, 7, 11829. [Google Scholar] [CrossRef] [Green Version]
- Kostrzewa, R.M. Neonatal 6-Hydroxydopamine Lesioning of Rats Dopaminergic Neurotoxicity: Proposed Animal Model of Parkinson’s Disease. J. Neural Transm. 2022, 129, 445–461. [Google Scholar] [CrossRef]
- Li, J.; Yu, T.; Shi, F.; Zhang, Y.; Duan, Z.; Fu, B.; Zhang, Y. Involvement of Ventral Periaqueductal Gray Dopaminergic Neurons in Propofol Anesthesia. Neurochem. Res. 2018, 43, 838–847. [Google Scholar] [CrossRef]
- Qiu, G.; Wu, Y.; Yang, Z.; Li, L.; Zhu, X.; Wang, Y.; Sun, W.; Dong, H.; Li, Y.; Hu, J. Dexmedetomidine Activation of Dopamine Neurons in the Ventral Tegmental Area Attenuates the Depth of Sedation in Mice. Anesthesiology 2020, 133, 377–392. [Google Scholar] [CrossRef]
- Solt, K.; Cotten, J.F.; Cimenser, A.; Wong, K.F.; Chemali, J.J.; Brown, E.N. Methylphenidate Actively Induces Emergence from General Anesthesia. Anesthesiology 2011, 115, 791–803. [Google Scholar] [CrossRef] [Green Version]
- Kenny, J.D.; Taylor, N.E.; Brown, E.N.; Solt, K. Dextroamphetamine (but Not Atomoxetine) Induces Reanimation from General Anesthesia: Implications for the Roles of Dopamine and Norepinephrine in Active Emergence. PLoS ONE 2015, 10, e0131914. [Google Scholar] [CrossRef]
- Kato, R.; Zhang, E.R.; Mallari, O.G.; Moody, O.A.; Vincent, K.F.; Melonakos, E.D.; Siegmann, M.J.; Nehs, C.J.; Houle, T.T.; Akeju, O.; et al. D-Amphetamine Rapidly Reverses Dexmedetomidine-Induced Unconsciousness in Rats. Front. Pharmacol. 2021, 12, 668285. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, Y.; Zhang, C.; Wang, M.; Zhang, M.; Yu, L.; Yan, M. The Role of Dopaminergic Vta Neurons in General Anesthesia. PLoS ONE 2015, 10, e0138187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Li, H.; Wang, D.; Guo, Y.; Zhang, X.; Ran, M.; Yang, C.; Yang, Q.; Dong, H. Orexin Activated Emergence from Isoflurane Anaesthesia Involves Excitation of Ventral Tegmental Area Dopaminergic Neurones in Rats. Br. J. Anaesth. 2019, 123, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Gangarossa, G.; Espallergues, J.; de Kerchove, A.; El Mestikawy, S.; Gerfen, C.R.; Hervé, D.; Girault, J.A.; Valjent, E. Distribution and Compartmental Organization of Gabaergic Medium-Sized Spiny Neurons in the Mouse Nucleus Accumbens. Front. Neural Circuits 2013, 7, 22. [Google Scholar] [CrossRef] [Green Version]
- Salgado, S.; Kaplitt, M.G. The Nucleus Accumbens: A Comprehensive Review. Stereotact. Funct. Neurosurg. 2015, 93, 75–93. [Google Scholar] [CrossRef]
- Pain, L.; Gobaille, S.; Schleef, C.; Aunis, D.; Oberling, P. In Vivo Dopamine Measurements in the Nucleus Accumbens after Nonanesthetic and Anesthetic Doses of Propofol in Rats. Anesth. Analg. 2002, 95, 915–919. [Google Scholar] [CrossRef] [Green Version]
- Bao, W.W.; Xu, W.; Pan, G.J.; Wang, T.X.; Han, Y.; Qu, W.M.; Li, W.X.; Huang, Z.L. Nucleus Accumbens Neurons Expressing Dopamine D1 Receptors Modulate States of Consciousness in Sevoflurane Anesthesia. Curr. Biol. 2021, 31, 1893–1902.e1895. [Google Scholar] [CrossRef]
- Zhang, L.N.; Li, Z.J.; Tong, L.; Guo, C.; Niu, J.Y.; Hou, W.G.; Dong, H.L. Orexin-a Facilitates Emergence from Propofol Anesthesia in the Rat. Anesth. Analg. 2012, 115, 789–796. [Google Scholar] [CrossRef]
- Yuan, J.; Luo, Z.; Zhang, Y.; Zhang, Y.; Wang, Y.; Cao, S.; Fu, B.; Yang, H.; Zhang, L.; Zhou, W.; et al. Gabaergic Ventrolateral Pre-Optic Nucleus Neurons Are Involved in the Mediation of the Anesthetic Hypnosis Induced by Propofol. Mol. Med. Rep. 2017, 16, 3179–3186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, Y.; Qian, K.; Zhang, L.; Yu, T. Histaminergic H1 and H2 Receptors Mediate the Effects of Propofol on the Noradrenalin-Inhibited Neurons in Rat Ventrolateral Preoptic Nucleus. Neurochem. Res. 2017, 42, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Li, D.D.; Wang, G.Q.; Wu, Q.; Shi, J.S.; Zhang, F. Dendrobium Nobile Lindl Alkaloid Attenuates 6-Ohda-Induced Dopamine Neurotoxicity. Biotechnol. Appl. Biochem. 2021, 68, 1501–1507. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.E.; Chemali, J.J.; Brown, E.N.; Solt, K. Activation of D1 Dopamine Receptors Induces Emergence from Isoflurane General Anesthesia. Anesthesiology 2013, 118, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Ao, Y.; Liu, Y.; Zhang, X.; Li, Y.; Tang, F.; Xu, H. Activation of Dopamine Signals in the Olfactory Tubercle Facilitates Emergence from Isoflurane Anesthesia in Mice. Neurochem. Res. 2021, 46, 1487–1501. [Google Scholar] [CrossRef] [PubMed]
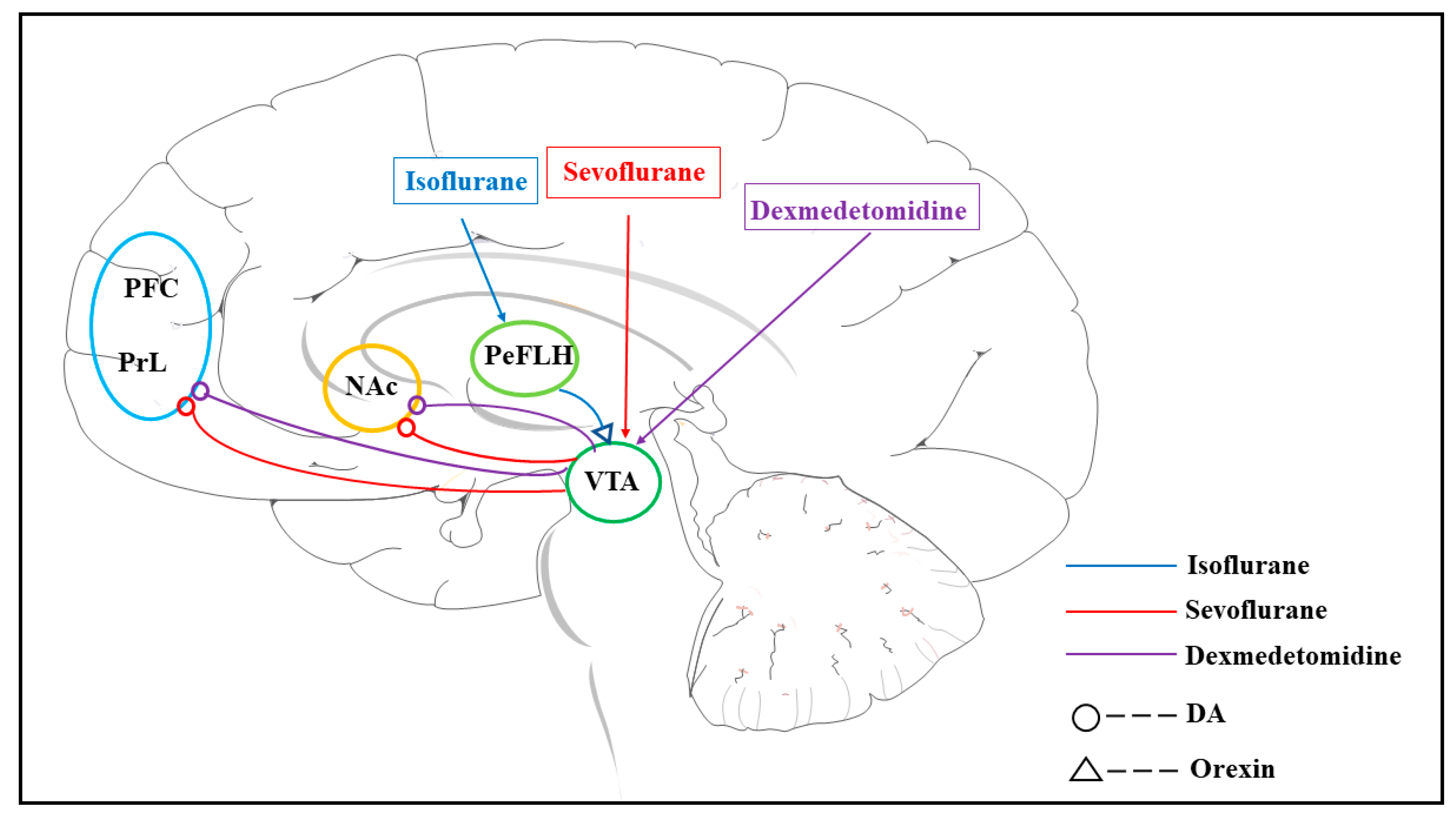
Figure 1.
Neural circuit of the dopaminergic system in general anesthesia. Neural circuits are involved in the dopamine system under different anesthetics on recovery from general anesthesia. Circles represent dopaminergic neural projections, and the triangle represents orexin neural projections. The arrows indicate that the anesthetics act on the corresponding nucleus, and the colors are used to identify the different anesthetics. The neural circuits under the same general anesthetic have the same color line; abbreviations: the prefrontal cortex (PFC), prelimbic cortex (PrL), nucleus accumbens (NAc), periaqueductal lateral hypothalamic area (PeFLH), ventral tegmental area (VTA), Dexmedetomidine (Dex), and dopamine (DA).
Figure 1.
Neural circuit of the dopaminergic system in general anesthesia. Neural circuits are involved in the dopamine system under different anesthetics on recovery from general anesthesia. Circles represent dopaminergic neural projections, and the triangle represents orexin neural projections. The arrows indicate that the anesthetics act on the corresponding nucleus, and the colors are used to identify the different anesthetics. The neural circuits under the same general anesthetic have the same color line; abbreviations: the prefrontal cortex (PFC), prelimbic cortex (PrL), nucleus accumbens (NAc), periaqueductal lateral hypothalamic area (PeFLH), ventral tegmental area (VTA), Dexmedetomidine (Dex), and dopamine (DA).


{kind=link}
{kind=link}
Table 1.
The dopaminergic system is involved in regulating consciousness during general anesthesia; abbreviations: Sprague Dawley (SD), the ventral tegmental area (VTA), dopamine (DA), the dopamine receptor (DR), substantia nigra (SN), the dopamine transporter (DAT), nucleus accumbens (NAc), medium spiny neurons (MSNs), ventral periaqueductal grey matter (vPAG), 6-hydroxydopamine (6-OHDA), the prelimbic cortex (PrL), the medial prefrontal cortex (mPFC), dexmedetomidine (Dex), the periaqueductal lateral hypothalamic area (PeFLH), tyrosine hydroxylase (TH), and the olfactory tubercle (OT).
Table 1.
The dopaminergic system is involved in regulating consciousness during general anesthesia; abbreviations: Sprague Dawley (SD), the ventral tegmental area (VTA), dopamine (DA), the dopamine receptor (DR), substantia nigra (SN), the dopamine transporter (DAT), nucleus accumbens (NAc), medium spiny neurons (MSNs), ventral periaqueductal grey matter (vPAG), 6-hydroxydopamine (6-OHDA), the prelimbic cortex (PrL), the medial prefrontal cortex (mPFC), dexmedetomidine (Dex), the periaqueductal lateral hypothalamic area (PeFLH), tyrosine hydroxylase (TH), and the olfactory tubercle (OT).
| Animal | Nervous Nuclei/ Neural Circuit | General Anesthetics | Intervention /Methods | Findings in Consciousness during General Anesthesia | Reference |
|---|---|---|---|---|---|
| SD rats | VTA-PrL | Sevoflurane | D1R agonist and antagonist were intraperitoneally injected or microinjected into PrL. Chemogenetics and Optogenetics | The dopaminergic pathway of VTA-PrL promotes the emergence of sevoflurane anesthesia. | [58] |
| DAT-cre mice | VTA, VTA-NAc | Sevoflurane, isoflurane | Chemogenetics, Optogenetics, and DA sensor based genetically encoded | Activation of the VTA-NAc dopaminergic pathway delayed the induction and promoted the emergence from general anesthesia | [61] |
| SD rats | vPAG | Isoflurane | Microinjection of 6-OHDA into vPAG. Whole-cell patch clamp recording | Lesion to the vPAG-DA neurons shortens the induction time and prolongs the recovery time in isoflurane anesthesia by activating the GABAA receptor | [70] |
| SD rats | NAc | Propofol | Microinjection of DR agonists and antagonists into NAc. Whole-cell patch clamp recording | The D1R of the NAc MSNs is involved in regulating the emergence of propofol-induced unconsciousness. | [71] |
| SD rats | VTA | Isoflurane, propofol, ketamine | 6-OHDA lesioned Bilateral VTA-DA neurons | VTA-DA neurons may be involved in the emergence of propofol | [73] |
| SD rats | VTA | Propofol | Knockdown DAT | DAT inhibition in VTA enhances PFC neurons activity and promotes recovery after propofol anesthesia. | [74] |
| SD rats | VTA, SN | Isoflurane, propofol | Electrostimulation | Electrical stimulation of VTA, but not SN, induced recovery from anesthesia | [75] |
| DAT-cre mice | VTA | Isoflurane | Optogenetics activation of VTA-DA. DR antagonists were used to intervening | Selective stimulation of VTA-DA neurons induced emergence from general anesthesia | [76] |
| Hcrt-cre rats | VTA PeFLH- VTA | Isoflurane | Microinjection of Orexins into VTA. Identification of Orexin receptors and DA neurons in VTA by immunofluorescence. Optogenetics | Orexin promotes the emergence of isoflurane anesthesia by activating DA neurons in VTA. | [77] |
| D1R-cre mice | NAc | Sevoflurane | Chemogenetics and Optogenetics | Activation of NAc D1R neurons induced cortical activation and behavioral emergence during sevoflurane anesthesia | [78] |
| SD rats | vPAG | Propofol | Microinjection of 6-OHDA into vPAG. Whole-cell patch clamp recording | Lesion to the vPAG-DA neurons shortens the induction time and prolongs the recovery time during propofol anesthesia | [79] |
| DAT-cre mice | VTA-NAc, VTA-mPFC | Dex | DA sensor based genetically encoded. Chemogenomics | Dex activates DA neurons in the VTA and increases DA concentration in the NAc and mPFC. | [80] |
| SD rats | / | Isoflurane | Intraperitoneal injection of DR agonists and antagonists | Activation of D1R mediates emergence from isoflurane | [81] |
| SD rats | / | Sevoflurane, propofol | Dextroamphetamine or atomoxetine is injected intraperitoneally during anesthesia. | Dextroamphetamine induced recovery from general anesthesia, whereas atomoxetine did not | [82] |
| SD rats | / | Dex, ketamine | Dextroamphetamine is administered intravenously during anesthesia | Dextroamphetamine induced recovery after anesthesia with Dex via D1R or D5R, but not with ketamine | [83] |
| TH-IRES-cre mice | OT | Isoflurane | Microinjection of DR agonists and antagonists into OT. Optogenetics | The dopaminergic pathway in OT accelerated emergence from isoflurane anesthesia | [84] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, J.; Miao, X.; Sun, Y.; Li, S.; Wu, A.; Wei, C. Dopaminergic System in Promoting Recovery from General Anesthesia. Brain Sci. 2023, 13, 538. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci13040538
AMA Style
Wang J, Miao X, Sun Y, Li S, Wu A, Wei C. Dopaminergic System in Promoting Recovery from General Anesthesia. Brain Sciences. 2023; 13(4):538. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci13040538
Chicago/Turabian StyleWang, Jinxu, Xiaolei Miao, Yi Sun, Sijie Li, Anshi Wu, and Changwei Wei. 2023. "Dopaminergic System in Promoting Recovery from General Anesthesia" Brain Sciences 13, no. 4: 538. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci13040538
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.