Local and Relayed Effects of Deep Brain Stimulation of the Pedunculopontine Nucleus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Pedunculopontine Nucleus
2.1. Anatomy
2.2. Physiology
2.3. Intrinsic Properties
2.4. Circuit Effects
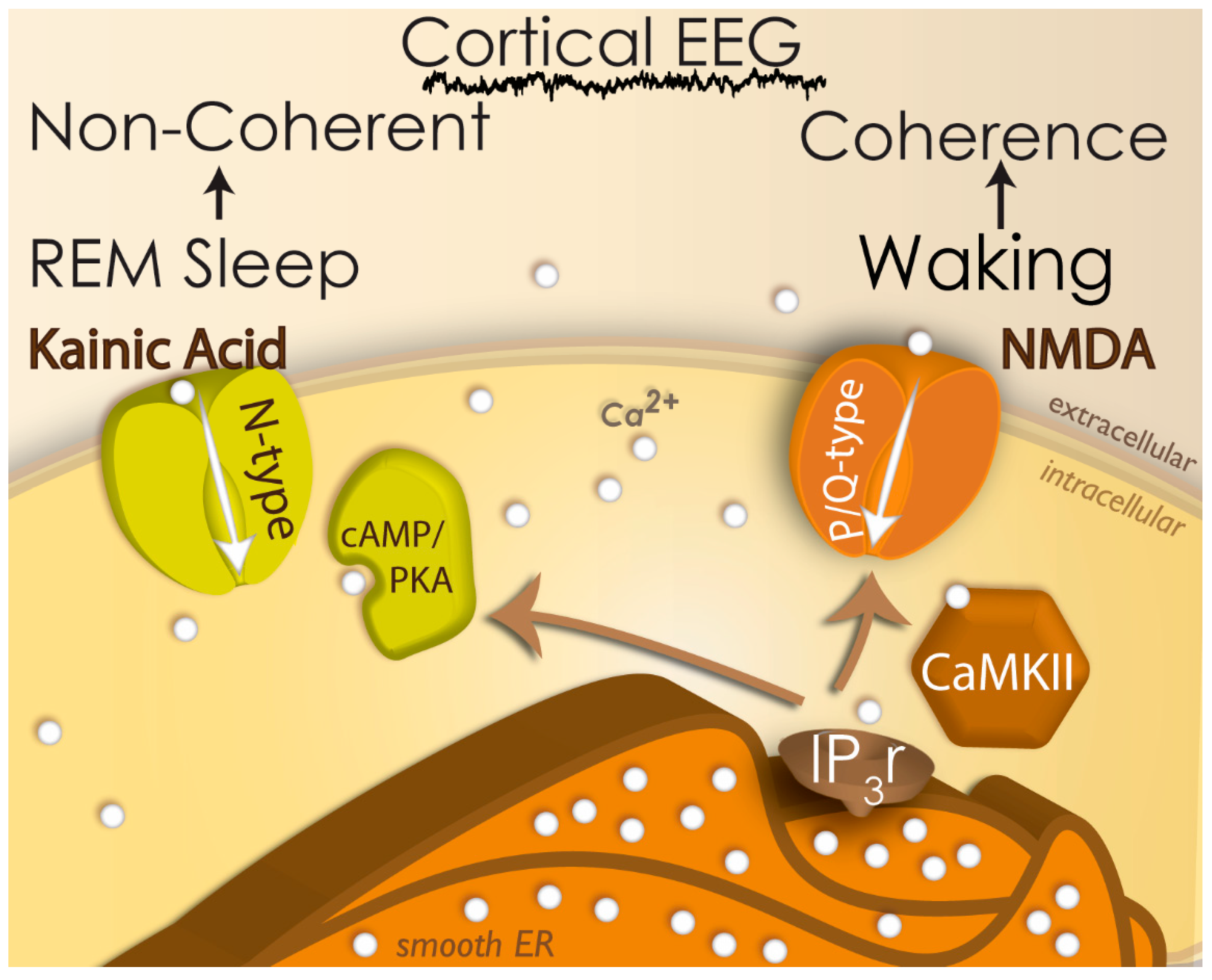
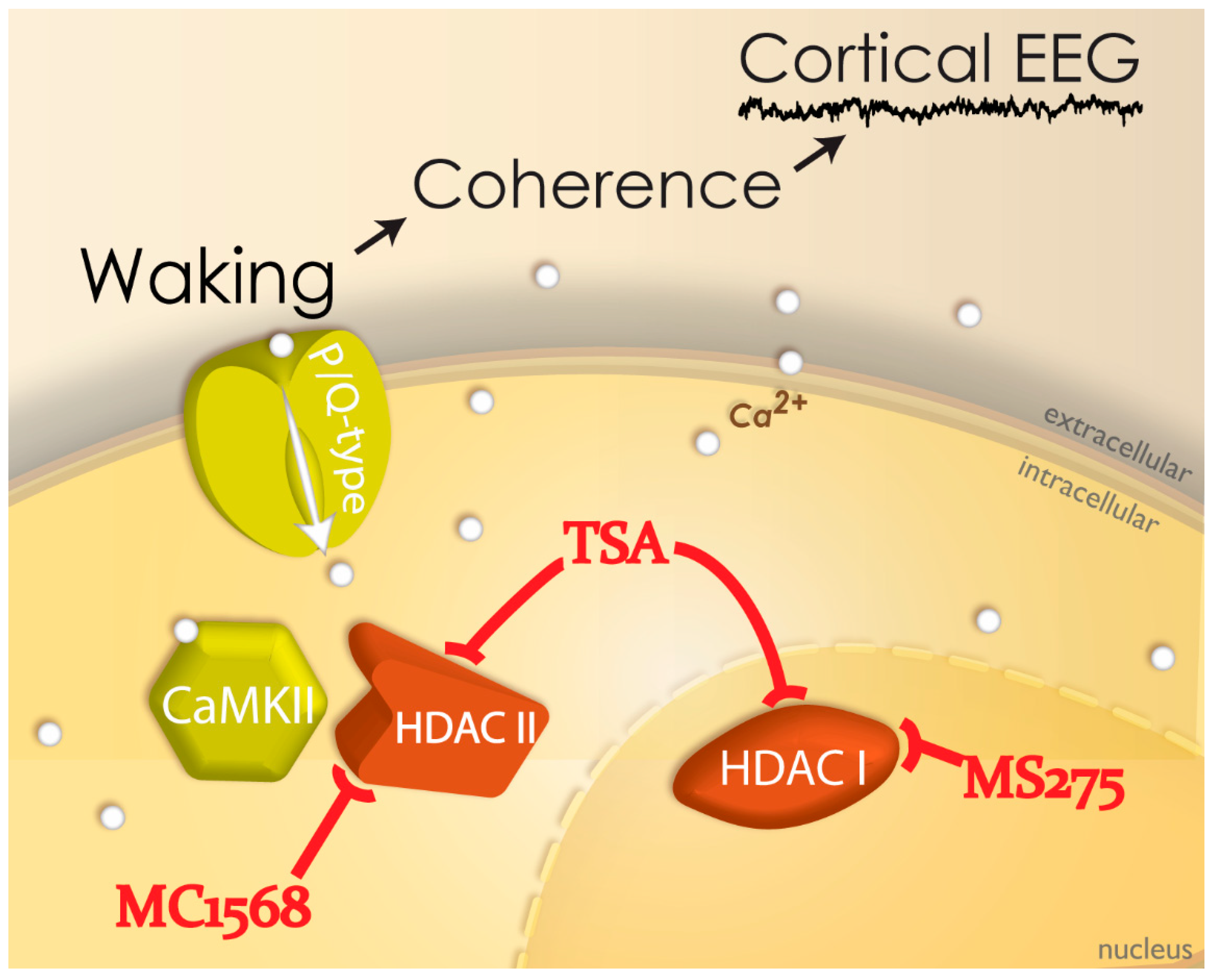
2.5. Neuroepigenetics of PPN Activity
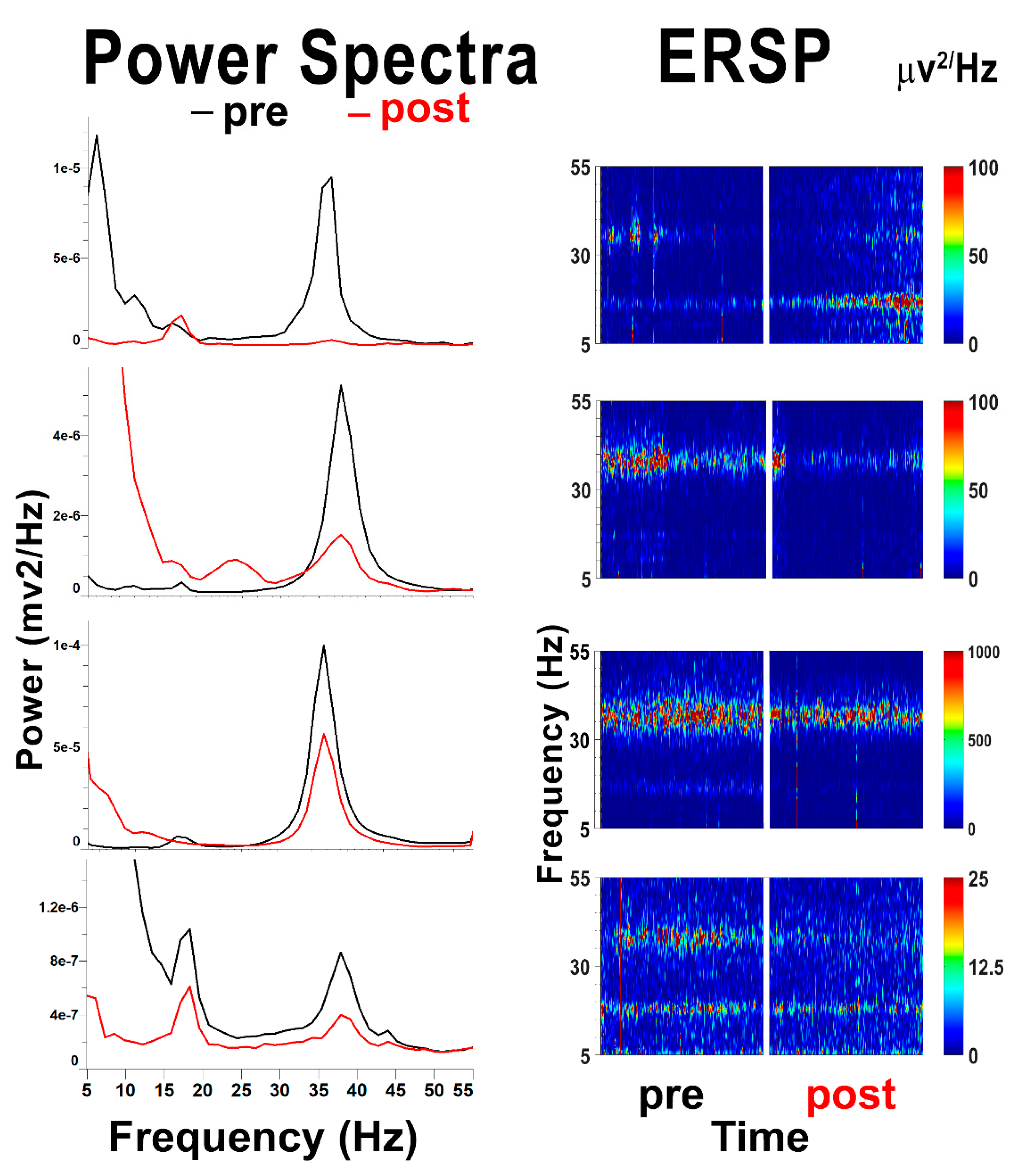
2.6. The Wrong Place for Optogenetics
3. Clinical Implications
3.1. Stimulus Parameters
3.2. PPN DBS
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Orr, H. Contribution to the embryology of the lizard; with especial reference to the central nervous system and some organs of the head; together with observations on the origin of the vertebrates. J. Morphol. 1887, 1, 311–372. [Google Scholar] [CrossRef]
- Jenkinson, N.; Nandi, D.; Muthusamy, K.; Ray, N.J.; Gregory, R.; Stein, J.F.; Aziz, T. Anatomy, Physiology, and Pathophysiology of the Pedunculopontine Nucleus. Mov. Disord. 2009, 24, 319–328. [Google Scholar] [CrossRef]
- Nauta, W.J.; Mehler, W.R. Projections of the lentiform nucleus in the monkey. Brain Res. 1966, 1, 3–42. [Google Scholar] [CrossRef]
- Kim, R.; Nakano, K.; Jayaraman, A.; Carpenter, M.B. Projections of the globus pallidus and adjacent structures: An autoradiographic study in the monkey. J. Comp. Neurol. 1976, 169, 263–290. [Google Scholar] [CrossRef]
- Dahlstrom, A.; Fuxe, K. Evidence for the existence of monoamine-containing neurons in the central nervous system. I. Demonstration of monoamines in the cell bodies of brain stem neurons. Acta Physiol. Scand. 1964, 62 (Suppl. 232), 5–55. [Google Scholar]
- Schute, C.C.D.; Lewis, P.R. The ascending cholinergic reticular system: Neocortical, olfactory and subcortical projections. Brain 1967, 90, 497–520. [Google Scholar] [CrossRef]
- Rye, D.B.; Saper, C.B.; Lee, H.J.; Wainer, B.H. Pedunculopontine tegmental nucleus of the rat: Cytoarchitecture, cytochemistry, and some extrapyramidal connections of the mesopontine tegmentum. J. Comp. Neurol. 1987, 259, 483–528. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rill, E.; Skinner, R.D.; Gilmore, S.A. Pallidal projections to the mesencephalic locomotor region. Am. J. Anat. 1981, 161, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rill, E.; Skinner, R.D.; Jackson, M.B.; Smith, M.M. Connections of the mesencephalic locomotor region (MLR). I. Substantia Nigra afferents. Brain Res. Bull. 1983, 10, 57–62. [Google Scholar] [CrossRef]
- Garcia-Rill, E.; Skinner, R.D.; Gilmore, S.A.; Owings, R. Connections of the mesencephalic locomotor region (MLR). II. Afferents and efferents. Brain Res. Bull. 1983, 10, 63–71. [Google Scholar] [CrossRef]
- Wang, H.L.; Morales, M. Pedunculopontine and laterodorsal tegmental nuclei contan distinct populations of cholinergic, glutamatergic and GABAergic neurons in the rat. Eur. J. Neurosci. 2009, 29, 340–358. [Google Scholar] [CrossRef] [PubMed]
- Reese, N.B.; Garcia-Rill, E.; Skinner, R.D. The pedunculopontine nucleus-auditory input, arousal and pathophysiology. Prog. Neurobiol. 1995, 47, 105–133. [Google Scholar] [CrossRef]
- Garcia-Rill, E. Sleep and arousal states: Reticular activating system. In New Encyclopedia of Neuroscience; Squire, L.R., Bloom, F., Spitzer, N., Gage, F., Albright, T., Eds.; Elsevier: Oxford, UK, 2009; Volume 8, pp. 137–143. [Google Scholar]
- Boucetta, S.; Cisse, Y.; Mainville, L.; Morales, M.; Jones, B.E. Discharge profiles across the sleep-waking cycle of identified cholinergic, GABAergic, and glutamatergic neurons in the pontomesencephalic tegmentum of the rat. J. Neurosci. 2014, 34, 4708–4727. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Siwek, D.F. Single cell activity patterns of pedunculopontine tegmentum neurons across the sleep-wake cycle in the freely moving rats. J. Neurosci. Res. 2002, 70, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K. Some anatomical and physiological properties of ponto-mesencephalic tegmental neurons with special reference to the PGO waves and postural Atonia during paradoxical sleep in the cat. In The Reticular Formation Revisited; Hobson, J.A., Brazier, M.A.B., Eds.; Raven: New York, NY, USA, 1980; pp. 427–447. [Google Scholar]
- Steriade, M.; Paré, D.; Datta, S.; Oakson, D.; Curro Dossi, R. Different cellular types in mesopontine cholinergic nuclei related to ponto-geniculo-occipital waves. J. Neurosci. 1990, 10, 2560–2579. [Google Scholar] [CrossRef] [PubMed]
- Datta, S. Evidence that REM sleep is controlled by the activation of brain stem pedunculopontine tegmental kainate receptor. J. Neurophysiol. 2002, 87, 1790–1798. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Siwek, D.F. Excitation of the brain stem pedunculopontine tegmentum cholinergic cells induces wakefulness and rem sleep. J. Neurophysiol. 1997, 77, 2975–2988. [Google Scholar] [CrossRef]
- Datta, S.; Spoley, E.E.; Patterson, E.H. Microinjection of glutamate into the pedunculopontine tegmentum induces REM sleep and wakefulness in the rat. Am. J. Physiol. Reg. Integr. Comp. Physiol. 2001, 280, R752–R759. [Google Scholar] [CrossRef]
- Datta, S.; Patterson, E.H.; Spoley, E.E. Excitation of pedunculopontine tegmental NMDA receptors induces wakefulness and cortical activation in the rat. J. Neurosci. Res. 2001, 66, 109–116. [Google Scholar] [CrossRef]
- Datta, S.; Desarnaud, F. Protein kinase A in the pedunculopontine tegmental nucleus of rat contributes to regulation of rapid eye movement sleep. J. Neurosci. 2010, 30, 12263–12273. [Google Scholar] [CrossRef]
- Datta, S.; O’Malley, M.O.; Patterson, E.H. Calcium/calmodulin kinase II in the pedunculopontine tegmental nucleus modulates the initiation and maintenance of wakefulness. J. Neurosci. 2011, 23, 17007–17016. [Google Scholar] [CrossRef]
- Kayama, Y.; Ohta, M.; Jodo, E. Firing of ‘possibly’ cholinergic neurons in the rat laterodorsal tegmental nucleus during sleep and wakefulness. Brain Res. 1992, 569, 210–220. [Google Scholar] [CrossRef]
- Sakai, K.; El Mansari, M.; Jouvet, M. Inhibition by carbachol microinjections of presumptive cholinergic PGO-on neurons in freely moving cats. Brain Res. 1990, 527, 213–223. [Google Scholar] [CrossRef]
- Lindsley, D.B.; Bowden, J.W.; Magoun, H.W. Effect upon the EEG of acute injury to the brainstem activating system. Electroencephalogr. Clin. Neurophysiol. 1949, 1, 475–486. [Google Scholar] [CrossRef]
- Moruzzi, G. The sleep-waking cycle. Ergeb. Physiol. 1972, 64, 1–165. [Google Scholar]
- Moruzzi, G.; Magoun, H.W. Brain stem reticular formation and activation of the EEG. Electroencephalogr. Clin. Neurophysiol. 1949, 1, 455–473. [Google Scholar] [CrossRef]
- Steriade, M.; Constantinescu, E.; Apostol, V. Correlations between alterations of the cortical transaminase activity and EEG patterns of sleep and wakefulness induced by brainstem transections. Brain Res. 1969, 13, 177–180. [Google Scholar] [CrossRef]
- Steriade, M.; Curro Dossi, R.; Paré, D.; Oakson, G. Fast oscillations (20–40 Hz) in thalamocortical systems and their potentiation by mesopontine cholinergic nuclei in the cat. Proc. Natl. Acad. Sci. USA 1991, 88, 4396–4400. [Google Scholar] [CrossRef]
- Brown, R.E.; Winston, S.; Basheer, R.; Thakkar, M.M.; McCarley, R.W. Electrophysiological characterization of neurons in the dorsolateral pontine REM sleep induction zone of the rat: Intrinsic membrane properties and responses to carbachol and orexins. Neuroscience 2006, 143, 739–755. [Google Scholar] [CrossRef]
- Goetz, L.; Piallat, B.; Bhattacharjee, M.; Mathieu, H.; David, O.; Chabardès, S. The primate pedunculopontine nucleus region: Towards a dual role in locomotion and waking state. J. Neural Transm. 2016, 123, 667–678. [Google Scholar] [CrossRef]
- Fraix, V.; Bastin, J.; David, O.; Goetz, L.; Ferraye, M.; Benabid, A.L.; Chabardes, S.; Pollak, P.; Debu, B. Pedunculopontine nucleus area oscillations during stance, stepping and freezing in Parkinson’s disease. PLoS ONE 2013, 8, e83919. [Google Scholar] [CrossRef] [PubMed]
- Mena-Segovia, J.; Sims, H.M.; Magill, P.J.; Bolam, J.P. Cholinergic brainstem neurons modulate cortical gamma activity during slow oscillations. J. Physiol. 2008, 586, 2947–2960. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.; Kezunovic, N.; Ye, M.; Hyde, J.R.; Hayar, A.; Williams, D.K.; Garcia-Rill, E. Gamma band unit activity and population responses in the pedunculopontine nucleus. J. Neurophysiol. 2010, 104, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Kezunovic, N.; Urbano, F.J.; Simon, C.; Hyde, J.; Smith, K.; Garcia-Rill, E. Mechanism behind gamma band activity in the pedunculopontine nucleus (PPN). Eur. J. Neurosci. 2011, 34, 404–415. [Google Scholar] [CrossRef] [PubMed]
- Kezunovic, N.; Hyde, J.; Goitia, B.; Bisagno, V.; Urbano, F.J.; Garcia-Rill, E. Muscarinic modulation of high frequency oscillations in pedunculopontine neurons. Front. Neurol. 2013, 4, 176. [Google Scholar] [CrossRef] [PubMed]
- Kezunovic, N.; Hyde, J.; Simon, C.; Urbano, F.J.; Williams, D.K.; Garcia-Rill, E. Gamma band activity in the developing parafascicular nucleus (Pf). J. Neurophysiol. 2012, 107, 772–784. [Google Scholar] [CrossRef]
- Hyde, J.; Kezunovic, N.; Urbano, F.J.; Garcia-Rill, E. Visualization of fast calcium oscillations in the parafascicular nucleus. J. Eur. Physiol. 2013, 465, 1327–1340. [Google Scholar] [CrossRef]
- Hyde, J.; Kezunovic, N.; Urbano, F.J.; Garcia-Rill, E. Spatiotemporal properties of high speed calcium oscillations in the pedunculopontine nucleus. J. Appl. Physiol. 2013, 115, 1402–1414. [Google Scholar] [CrossRef]
- D’Onofrio, S.; Kezunovic, N.; Hyde, J.R.; Luster, B.; Messias, E.; Urbano, F.J.; Garcia-Rill, E. Modulation of gamma oscillations in the pedunculopontine nucleus (PPN) by neuronal calcium sensor protein-1 (NCS-1): Relevance to schizophrenia and bipolar disorder. J. Neurophysiol. 2015, 113, 709–719. [Google Scholar] [CrossRef]
- Luster, B.; D’Onofrio, S.; Urbano, F.J.; Garcia-Rill, E. High-Threshold Ca2+ channels behind gamma band activity in the pedunculopontine nucleus (PPN). Physiol. Rep. 2015, 3, e12431. [Google Scholar] [CrossRef]
- Garcia-Rill, E.; Heister, D.S.; Ye, M.; Charlesworth, A.; Hayar, A. Electrical coupling: Novel mechanism for sleep-wake control. Sleep 2007, 30, 1405–1414. [Google Scholar] [CrossRef]
- Urbano, F.J.; Leznik, E.; Llinas, R. Modafinil enhances thalamocortical activity by increasing neuronal electrotonic coupling. Proc. Natl. Acad. Sci. USA 2007, 104, 12554–12559. [Google Scholar] [CrossRef]
- Garcia-Rill, E.; Mahaffey, S.; Hyde, J.R.; Urbano, F.J. Bottom-up gamma maintenance in various disorders. Neurobiol. Dis. 2018. [Google Scholar] [CrossRef]
- Urbano, F.J.; Kezunovic, N.; Hyde, J.; Simon, C.; Beck, P.; Garcia-Rill, E. Gamma band activity in the reticular activating system (RAS). Front. Neurol. Sleep Chronobiol. 2012, 3, 6. [Google Scholar]
- Luster, B.; Urbano, F.J.; Garcia-Rill, E. Intracellular mechanisms modulating gamma band activity in the pedunculopontine nucleus (PPN). Physiol. Rep. 2016, 4, e12787. [Google Scholar] [CrossRef]
- Garcia-Rill, E.; Kezunovic, N.; D’Onofrio, S.; Luster, B.; Hyde, J.; Bisagno, V.; Urbano, F.J. Gamma band activity in the RAS- intracellular mechanisms. Exp. Brain Res. 2014, 232, 1509–1522. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rill, E.; Hyde, J.; Kezunovic, N.; Urbano, F.J.; Petersen, E. The physiology of the pedunculopontine nucleus- implications for deep brain stimulation. J. Neural Transm. 2014, 122, 225–235. [Google Scholar] [CrossRef]
- Shik, M.L.; Severin, F.V.; Orlovskii, G.N. Control of walking and running by means of electric stimulation of the midbrain. Biofizika 1966, 11, 659–666. [Google Scholar]
- Garcia-Rill, E.; Charlesworth, A.; Heister, D.; Ye, M.; Hayar, A. The developmental decrease in REM sleep: The role of transmitters and electrical coupling. Sleep 2008, 31, 673–690. [Google Scholar] [CrossRef]
- Garcia-Rill, E.; D’Onofrio, S.; Mahaffey, S. Bottom-up Gamma: The Pedunculopontine Nucleus and Reticular Activating System. Transl. Brain Rhythm. 2016, 1, 49–53. [Google Scholar] [PubMed]
- Bastos, A.M.; Vezoli, J.; Bosman, C.A.; Schoffelen, J.M.; Oostenveld, R.; Dowdall, J.R.; De Weerd, P.; Kennedy, H.; Fries, P. Visual areas exert feedforward and feedback through distinct frequency channels. Neuron 2015, 85, 390–401. [Google Scholar] [CrossRef]
- Garcia-Rill, E.; Skinner, R.D.; Miyazato, H.; Homma, Y. Pedunculopontine stimulation induces prolonged activation of pontine reticular neurons. Neuroscience 2001, 104, 455–465. [Google Scholar] [CrossRef]
- Mamiya, N.; Bay, K.; Skinner, R.D.; Garcia-Rill, E. Induction of long-lasting depolarization in medioventral medulla (MED) neurons by cholinergic input from the pedunculopontine nucleus (PPN). J. Appl. Physiol. 2005, 99, 1127–1137. [Google Scholar] [CrossRef]
- Urbano, F.J.; Bisagno, V.; Mahaffey, S.; Lee, S.; Garcia-Rill, E. Class II histone deacetylases require P/Q-type Ca2+ channels and CaMKII to maintain gamma oscillations in the pedunculopontine nucleus. Sci. Rep. 2018, 8, 13156. [Google Scholar] [CrossRef]
- Fenno, L.; Yizhar, O.; Deisseroth, K. The development and application of optogenetics. Ann. Rev. Neurosci. 2001, 34, 389–412. [Google Scholar] [CrossRef]
- Cheng, K.P.; Kiernan, E.A.; Eliceiri, K.W.; Williams, J.C.; Watters, J.J. Blue Light Modulates Murine Microglial Gene Expression in the Absence of Optogenetic Protein Expression. Sci. Rep. 2016, 6, 21172. [Google Scholar] [CrossRef]
- Brady, J.D.; Rich, T.C.; Le, X.; Stafford, K.; Fowler, C.J.; Lynch, L.; Karpen, J.W.; Brown, R.L.; Martens, J.R. Functional role of lipid raft microdomains in cyclic nucleotide-gated channel activation. Mol. Pharmacol. 2004, 65, 503–511. [Google Scholar] [CrossRef]
- Azzopardi, E.; Louttit, A.G.; DeOliveira, C.; Laviolette, S.R.; Scmid, S. The role of cholinergic midbrain neurons in startle and prepulse inhibition. J. Neurosci. 2018, 38, 8798–8808. [Google Scholar] [CrossRef]
- Scammell, T.; Arrigoni, E.; Lipton, L. Neural circuitry of wakefulness and sleep. Neuron 2018, 93, 747–765. [Google Scholar] [CrossRef]
- Gueguinou, M.; Gambade, A.; Felix, R.; Chantome, A.; Fourbon, Y.; Bougnoux, P.; Weber, G.; Potier-Cartereau, M.; Vandier, C. Lipid rafts, KCa/ClCa/Ca2+ channel complexes and EGFR signaling: Novel targets to reduce tumor development by lipids? Biochim. Biophys. Acta 2015, 1848, 2603–2620. [Google Scholar] [CrossRef]
- Irnich, W. The chronaxie time and its practical implications. Pacing Clin. Electrophysiol. 1980, 3, 292–301. [Google Scholar] [CrossRef]
- Nowak, L.G.; Bullier, J. Axons, but not cell bodies, are activated by electrical stimulation in cortical gray matter. Exp. Brain Res. 1998, 118, 477–488. [Google Scholar] [CrossRef]
- Coles, C.H.; Bradke, F. Coordinating neuronal actin-microtubule dynamics. Curr. Biol. 2015, 25, R677–R691. [Google Scholar] [CrossRef]
- Shen, K.; Teruel, M.N.; Subramanian, K.; Meyer, T. CaMKIIbeta functions as an F-actin targeting module that localizes CaMKIIalpha/beta heterooligomers to dendritic spines. Neuron 1998, 21, 593–606. [Google Scholar] [CrossRef]
- Shen, K.; Meyer, T. Dynamic control of CaMKII translocation and localization in hippocampal neurons by NMDA receptor stimulation. Science 1999, 284, 162–166. [Google Scholar] [CrossRef]
- Mazzone, P.; Lozano, A.; Stanzione, P.; Galati, S.; Scarnati, E.; Peppe, A.; Stefani, A. Implantation of human pedunculopontine nucleus: A safe and clinically relevant target in Parkinson’s disease. Neuroreport 2005, 16, 1877–1891. [Google Scholar] [CrossRef]
- Plaha, P.; Gill, S.S. Bilateral deep brain stimulation of the pedunculopontine nucleus for Parkinson’s disease. Neuroreport 2005, 16, 1883–1887. [Google Scholar] [CrossRef]
- Mazzone, P.; Sposato, S.; Insola, A.; Dilazzaro, V.; Scarnati, E. Stereotaxic surgery of the nucleus tegmenti pedunculopontinus. Br. J. Neurosurg. 2008, 22, S33–S40. [Google Scholar] [CrossRef]
- Mazzone, P.; Insola, A.; Sposato, S.; Scarnati, E. The deep brain stimulation of the pedunculopontine tegmental nucleus. Neuromodulation 2009, 12, 191–204. [Google Scholar] [CrossRef]
- Zrinzo, L.; Zrinzo, L.V.; Tisch, S.; Limousin, P.D.; Yousry, T.A.; Afshar, F.; Hariz, M.I. Stereotactic localization of the human pedunculopontine nucleus: Atlas-based coordinates and validation of a magnetic resonance imaging protocol for direct localization. Brain 2008, 131, 1588–1598. [Google Scholar] [CrossRef]
- Weinberger, M.; Hamani, C.; Hutchison, W.D.; Moro, E.; Lozano, A.M.; Dostrovsky, J.O. Pedunculopontine nucleus microelectrode recordings in movement disorder patients. Exp. Brain Res. 2008, 188, 165–174. [Google Scholar] [CrossRef]
- Karachi, C.; Grabli, D.; Bernard, F.A.; Tandé, D.; Wattiez, N.; Belaid, H.; Bardinet, E.; Prigent, A.; Nothacker, H.P.; Hunot, S.; et al. Cholinergic mesencephalic neurons are involved in gait and postural disorders in Parkinson disease. J. Clin. Investig. 2010, 120, 2745–2754. [Google Scholar] [CrossRef]
- Tattersall, T.L.; Stratton, P.G.; Coyne, T.J.; Cook, R.; Silberstein, P.; Silburn, P.A.; Windels, F.; Sah, P. Imagined gait modulates neuronal network dynamics in the human pedunculopontine nucleus. Nat. Neurosci. 2014, 17, 449–454. [Google Scholar] [CrossRef]
- Ferraye, M.U.; Debu, B.; Fraix, V.; Goetz, L.; Ardouin, C.; Yelnik, J.; Henry-Lagrange, C.; Seigneuret, E.; Piallat, B.; Krack, P.; et al. Effects of pedunculopontine nucleus area stimulation on gait disorders in Parkinson’s disease. Brain 2010, 133, 205–214. [Google Scholar] [CrossRef]
- Moro, E.; Hamani, C.; Poon, Y.Y.; Al-Khairallah, T.; Dostrovsky, J.O.; Hutchison, W.D.; Lozano, A.M. Unilateral pedunculopontine stimulation improves falls in Parkinson’s disease. Brain 2010, 133, 215–224. [Google Scholar] [CrossRef]
- Stefani, A.; Lozano, A.M.; Peppe, A.; Stanzione, P.; Galati, S.; Troppei, D.; Pierantozzi, M.; Brusa, L.; Scarnati, E.; Mazzone, P. Bilateral deep brain stimulation of the pedunculopontine and subthalamic nuclei in severe Parkinson’s disease. Brain 2007, 130, 1596–1607. [Google Scholar] [CrossRef]
- Stefani, A.; Peppe, A.; Galati, S.; Stampanoni, A.; Bassi, M.; D’Angelo, V.; Pierantozzi, M. The serendipity case of the pedunculopontine nucleus low-frequency brain stimulation: Chasing a gait response, finding sleep, and cognitive improvement. Front. Neurol. 2013, 4, 68. [Google Scholar] [CrossRef]
- Alessandro, S.; Ceravolo, R.; Brusa, L.; Pierantozzi, M.; Costa, A.; Galati, S.; Placidi, F.; Romigi, A.; Iani, C.; Marzetti, F.; et al. Non-motor functions in parkinsonian patients implanted in the pedunculopontine nucleus: Focus on sleep and cognitive problems. J. Neurol. Sci. 2010, 289, 44–48. [Google Scholar] [CrossRef]
- Thevathasan, W.; Cole, M.H.; Grapel, C.L.; Hyam, J.A.; Jenkinson, N.; Brittain, J.S.; Coyne, T.J.; Silburn, P.A.; Aziz, T.Z.; Kerr, G.; Brown, P.; et al. A spatiotemporal analysis of gait freezing and the impact of pedunculopontine nucleus stimulation. Brain 2012, 135, 1446–1454. [Google Scholar] [CrossRef]
- Thevathasan, W.; Silburn, P.A.; Brooker, H.; Coyne, T.J.; Kahn, S.; Gill, S.S.; Aziz, T.Z.; Brown, P. The impact of low-frequency stimulation of the pedunculopontine nucleus region on reaction time in Parkinsonism. J. Neurol. Neurosurg. Psychiatry 2010, 81, 1099–1104. [Google Scholar] [CrossRef]
- Hamani, C.; Aziz, T.; Bloem, B.R.; Brown, P.; Chabardes, S.; Coyne, T.; Foote, K.; Garcia-Rill, E.; Hirsch, E.C.; Lozano, A.M.; et al. Pedunculopontine Nucleus Region Deep Brain Stimulation in Parkinson Disease: Surgical Anatomy and Terminology. Stereotact. Funct. Neurosurg. 2016, 94, 298–306. [Google Scholar] [CrossRef]
- Hamani, C.; Lozano, A.M.; Mazzone, P.A.; Moro, E.; Hutchison, W.; Silburn, P.A.; Zrinzo, L.; Alam, M.; Goetz, L.; Pereira, E.; et al. Pedunculopontine Nucleus Region Deep Brain Stimulation in Parkinson Disease: Surgical Techniques, Side Effects, and Postoperative Imaging. Stereotact. Funct. Neurosurg. 2016, 94, 307–319. [Google Scholar] [CrossRef]
- Goetz, L.; Bhattacharjee, M.; Ferraye, M.U.; Fraix, V.; Maineri, C.; Nosko, D.; Fenoy, A.J.; Piallat, B.; Torres, N.; Krainik, A.; et al. Deep brain stimulation of the pedunculopontine nucleus area in Parkinson’s disease: MRI-based anatomical correlations and optimal target. Neurosurgery 2018, 84, 506–518. [Google Scholar] [CrossRef]
- Buhmann, C.; Huckhagel, T.; Engel, K.; Gulberti, A.; Hidding, U.; Poetter-Nerger, M.; Goerendt, I.; Ludewig, P.; Braass, H.; Choe, C.U.; et al. Adverse events in deep brain stimulation: A retrspective long-term analysis of neurological, psychiatric and other occurrences. PLoS ONE 2017, 21, e0178984. [Google Scholar] [CrossRef]
- Garcia-Rill, E.; D’Onofrio, S.; Luster, B.; Mahaffey, S.; Urbano, F.J.; Phillips, C. The 10 Hz Frequency: A fulcrum for transitional brain states. Transl. Brain Rhythm. 2016, 1, 7–13. [Google Scholar]
- Little, S.; Beudel, M.; Zrinzo, L.; Foltynie, T.; Limousin, P.; Hariz, M.; Neal, S.; Cheeran, B.; Cagnan, H.; Gratwicke, J.; et al. Bilateral adaptive deep brain stimulation is effective in Parkinson’s disease. Mov. Disord. 2016, 87, 717–721. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Rill, E.; Tackett, A.J.; Byrum, S.D.; Lan, R.S.; Mackintosh, S.G.; Hyde, J.R.; Bisagno, V.; Urbano, F.J. Local and Relayed Effects of Deep Brain Stimulation of the Pedunculopontine Nucleus. Brain Sci. 2019, 9, 64. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9030064
Garcia-Rill E, Tackett AJ, Byrum SD, Lan RS, Mackintosh SG, Hyde JR, Bisagno V, Urbano FJ. Local and Relayed Effects of Deep Brain Stimulation of the Pedunculopontine Nucleus. Brain Sciences. 2019; 9(3):64. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9030064
Chicago/Turabian StyleGarcia-Rill, Edgar, Alan J. Tackett, Stephanie D. Byrum, Renny S. Lan, Samuel G. Mackintosh, James R. Hyde, Veronica Bisagno, and Francisco J. Urbano. 2019. "Local and Relayed Effects of Deep Brain Stimulation of the Pedunculopontine Nucleus" Brain Sciences 9, no. 3: 64. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9030064