3.2. Yield and Yield Parameters
Tomato is one of the most consumed vegetables worldwide and represents one of the driving crops for many countries economy, due to its dual use as a fresh and processed product (e.g., pasta, sauce, peeled tomatoes, juice, ketchup) [
8,
10]. To date, world tomato production is estimated at 180 million tons, with China alone having a total production of approximately 63 million tons, followed by India (~19 million tons), Turkey (~13 million tons), the United States of America (~11 million tons), Egypt (~7 million tons), and Italy (~5 million tons) [
40]. However, it is well known that tomato yield is strongly influenced by environmental factors (such as humidity, temperature, and solar radiation), genotype, and pre-harvest factors (growing practices) [
19,
41].
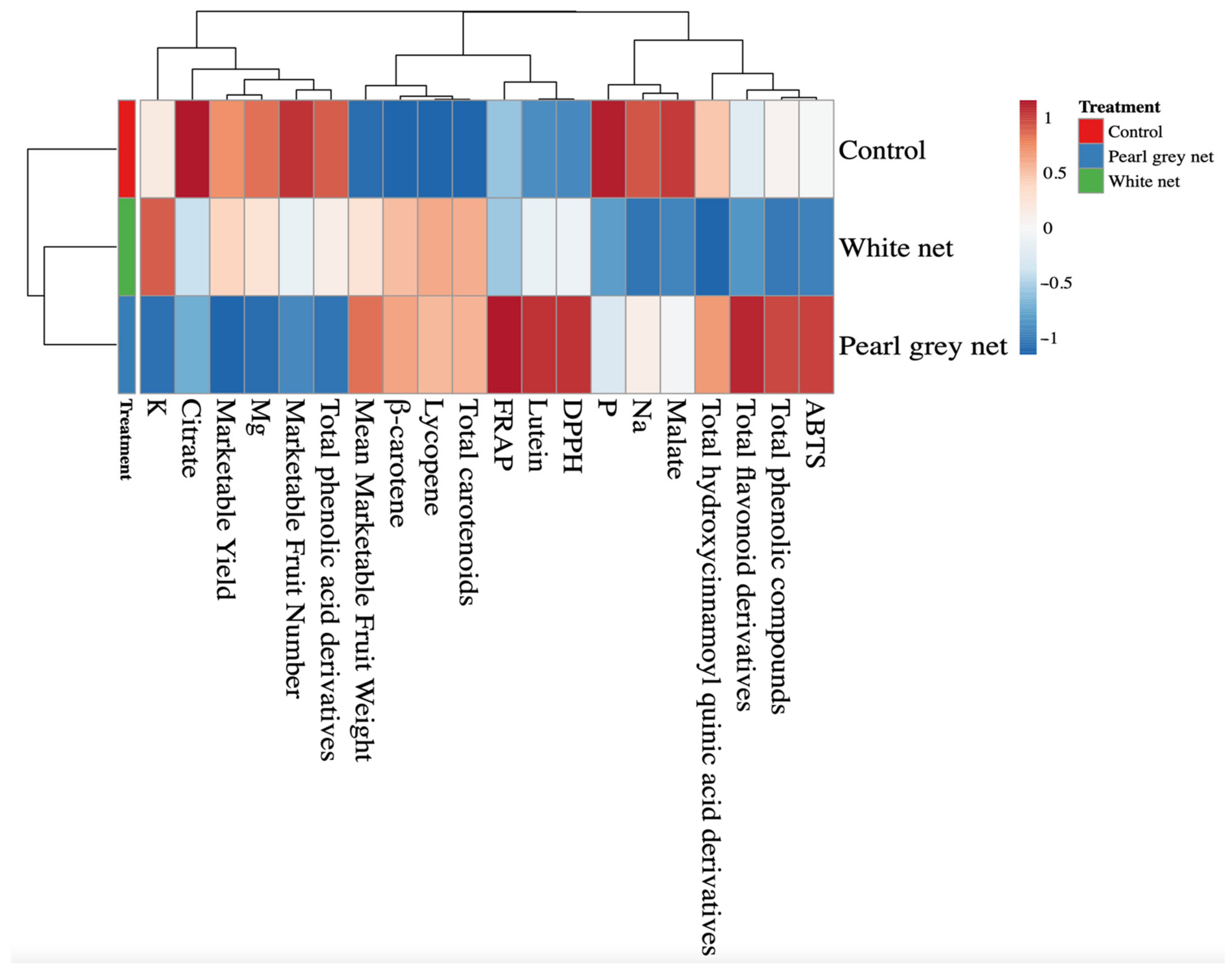
In our study, the number of total fruits per plant showed a significant decrease compared to the control when the plants were under shading (Control > White net > Pearl grey net;
Table 3). Probably, high light intensity conditions induced an eco-physiological response to mitigate stress, increasing the number of fruits but reducing the transpiring surface (diameter of the fruit) compared to shaded conditions. Consequently, the reduction in the fruit number induced by shading did not affect the weight of the fruit (
Table 3). Total, marketable, and unmarketable fruits weight (kg pl
−1) did not show significant differences in shaded plants compared to the control, in contrast to the findings of Angmo et al. [
42], who reported an increase in total marketable fruit weight in the open field compared to shaded conditions, which could be attributed to different environmental conditions, genetic material, and cultural practices [
43,
44,
45]. In the present experiment, we adopted a processing tomato cultivar that was neither tied nor defoliated, in contrast to the methods used by the authors mentioned above, and in part, could have determined different production responses. Shading nets resulted in a considerable increase in marketable fruit weight of 47.5%, compared to the control, justifying the non-significant difference in total fruit weight per plant (
Table 3).
3.3. Quality Attributes of Fruits
The growing interest in high-quality food products has forced growers to meet the changing needs of increasingly demanding consumers. In the past, the marketable quality of vegetables relied primarily on visible characteristics, but now sensory and organoleptic characteristics have become a primary parameter driving consumer choice [
2,
46]. In tomatoes, one of the sensory attributes that determine the organoleptic quality of the fruit is the content of soluble solids (glucose, fructose, and sucrose), which, combined with organic acids and amino acids, represents approximately 75% of dry matter [
46]. In the literature, it is known that fruit sweetness is strongly influenced by genetic material [
26]. Almeida et al. [
20] have studied the effects on the accumulation of total soluble solids (TSS) of five genotypes of tomatoes under different environmental conditions. The authors found that the TSS content ranged among the genotypes from 5.6 to 7.2 °Brix, and according to the environmental conditions from 3.8 to 8.9 °Brix. In our experiment, we found that light radiation affected this crucial qualitative parameter (
Table 4). Davies et al. [
47] highlighted the evidence of a direct relationship between solar radiation and sugar content in tomatoes. Our results confirm this correlation as fruits exposed to direct solar radiation (Control) showed the highest value of TSS (7.43 °Brix;
Table 4), confirming what was reported by Ilić et al. [
48] in a similar experiment. The higher TSS content in the control fruits was probably attributable to the lower water assimilation capacity of the fruits, which also justified the high dry matter content (8.71%;
Table 4) [
49]. However, it is worth considering that an increase in the amount of solar radiation received by the plant may increase photosynthesis, and thus carbohydrates in the fruit [
8]. On the other hand, the direct correlation between solar radiation and TSS is not univocal, as different results are found in the literature, again highlighting how genotype plays a crucial role in the adaptation to different environmental conditions [
10,
48].
Another qualitative aspect that can influence consumer choice is color, since a well-colored fruit is qualitatively superior. Practically, the color of the fruit depends on the physical and biochemical changes that occur naturally during the growth and ripening stages or after harvest [
50]. Among the CIELab colorimetric parameters, only the L (brightness) parameter varied significantly in response to shading, with the highest value obtained in fruits grown under white shading nets (
Table 4). For definition, L is “an approximate measure of brightness, which is the property according to which any color can be considered equivalent to a member of the greyscale, between black and white” [
51]. The increase in L under shaded conditions agrees with the findings of Messina et al. [
52]. However, the same authors also reported a decrease in a* values (less intense red color) and an increase in b* values (more intense yellow color) that we did not observe in our study (
Table 4).
Similar to TSS content and color, fruit size and shape are also essential quality traits. Although shape is primarily determined by genetic background, size also depends on the interaction of the latter with the environment [
2]. From a physiological point of view, the increase in fruit size depends on the enlargement of the pericarp due to the production of new cells during the anthesis process and the growth and expansion of cells that last until the fruit ripening [
2]. According to Angmo et al. [
42], compared to the control, we observed an average increase in the equatorial and polar diameter of fruits of 8.60 and 10.50%, respectively, when grown under shading nets (
Table 4). The larger fruit size recorded under shading nets accounted for the higher average marketable fruit weight (
Table 3), attributed to the higher water content in the fruit (lower dry matter) that resulted in a dilution effect on TSS (
Table 4).
3.4. Mineral Content of Fruits
Minerals, like other macromolecules (carbohydrates, proteins, and fats), are required to preserve some physical and biochemical processes essential for life [
53]. Currently, mineral deficiency in the human diet is a severe problem for industrial and developing countries [
54]. Given the high intake of tomatoes, the potential contribution of tomato fruit to the mineral intake of human diet is of high importance [
55]. It is well established that the most abundant mineral in tomato fruit is potassium [
54]. Potassium plays a crucial role in maintaining cellular homeostasis, nerve impulse conduction and muscle contraction, and the glycogenesis process [
53,
56]. In plants, potassium is an activator of enzymatic processes and contributes significantly to the photosynthetic process [
57]. Among the macronutrients reported in
Table 5, potassium was the most abundant mineral in the fruit and was affected by shading treatment, with the highest value recorded under white net. Although potassium is crucial for color determination, the change in its content was not coupled with a perceived change in color (a*) (
Table 4). This result could be partially related to the optimal potassium content (361.43–445.65 mg 100 g
−1 fw) [
57].
Phosphorus is the main component of bones, and is involved in many metabolic processes (kidney function and cell growth); it has a buffering action and is involved in the formation of high-energy compounds (adenosine triphosphate) and in phospholipid synthesis [
53,
58]. Similarly to potassium, the phosphorus content was significantly affected by growth conditions (
Table 5). The higher value (14.88 mg 100 g
−1 fw) recorded in fruits grown under unshaded conditions would help to better explain the higher TSS obtained from the same treatment. Indeed, Lavon et al. [
59] showed a positive correlation between this essential macroelement and TSS content in tomato fruits.
Although magnesium deficiency in the human diet is rarely a determinant of pathological states (WHO; [
60]), this mineral is crucial, as it is a component of bones and teeth and is an active component of different enzymatic systems [
53]. In our study, the magnesium content was not affected by shading (
Table 5). Although Milenković et al. [
19] observed a reduction in magnesium in tomato fruits exposed to direct solar radiation, our results do not show the same trend. This discordance could be attributable not only to the different genetic material, but also to the different light conditions.
Tomato acidity is a crucial component of the organoleptic quality of fruits [
61]. Interactions between reducing sugars and organic acids are essential to confer sweetness, tartness, and flavor intensity to fruits [
2,
3,
9]. The main organic acids in tomato fruits are malic and citric acids, but the perception of acidity is mainly due to the latter, which is the most abundant organic acid [
2,
61]. Shading treatments reduced the citrate content in fruits by 23.7%, compared to the control that showed the highest values (140.36 mg 100 g
−1 fw). The higher citrate content in the fruits of the control could probably be attributable to a higher source:sink ratio during the pre-ripening phase, increasing the respiration rate of the fruits. Therefore, a higher respiration rate could have promoted glycolysis and increased citrate production [
61]. However, it should be noted that changes in fruit water content were observed between treatments (
Table 3), which may have interfered with acidity due to a dilution/dehydration effect [
61].
3.5. Fruit Pigments
The relevance of the quanti-qualitative profile of carotenoids in tomato fruits is mainly attributable to their dual function. In fact, while these biomolecules determine the coloration of ripe fruits, they are crucial in the human diet due to their recognized antioxidant activity [
2,
62]. Furthermore, it is important to note that the body cannot synthesize these valuable pigments, making their intake through plant consumption mandatory [
63]. In plants, carotenoids are used to capture light and protect the photosynthetic apparatus from excessive solar radiation, attract pollinators, and facilitate seed dispersal [
2,
62]. The data reported in
Table 6 show that, compared to the control, shading treatments resulted in the highest biosynthesis of total carotenoids in fruits. Although there is a wide variability in carotenoid content in the literature, our results are in agreement with the findings of Flores et al. [
64] in red tomato fruits, which showed that the most abundant carotenoid was lycopene, followed by
β-carotene, and lutein. Compared to shaded conditions, the ~40.0% reduction in the lycopene content in control fruits confirms that excessive radiation exerts an inhibitory effect on the biosynthesis and accumulation of this critical pigment [
3,
65,
66]. Leyva et al. [
3] noted that the decrease in lycopene content which is found in our work as well, could be attributed to direct solar radiation and not air temperature, since Helyes et al. [
67] observed that fruit surface temperatures of 30 °C trigger the degradation of this pigment. These conditions could have occurred in our case under control unshaded conditions.
Lycopene is a crucial intermediate in the biosynthesis of many carotenoids such as
β-carotene and xanthophylls such as lutein [
63]. Consequently, it is not surprising that high solar radiation (Control) resulted in an average reduction in
β-carotene (−43.4%) content, compared to shaded conditions. Not least, it is interesting to note that although lycopene is responsible for the red color of tomatoes [
68], the significant differences in lycopene content between treatments did not affect the colorimetric parameter a* (
Table 4). This result could be related to the direct correlation between lycopene content and fruit size (equatorial diameter and polar diameter;
Table 4), which probably influenced the colorimetric analyses.
3.6. Phenolic Compounds and Antioxidant Activity of Fruits
In their natural habitats, plants are threatened by a large number of potential enemies, and to defend themselves, they produce a wide range of heterogeneous protection compounds (pigments, signaling molecules, and aromas) known as ‘secondary metabolites’, which play an important role in their survival [
69]. Secondary metabolites are classified on the basis of their chemical structure and biosynthetic pathways. They can be divided into three groups: terpenoids, phenolic compounds and flavonoids, and sulfur-containing compounds and nitrogen-containing alkaloids [
70]. Present in most fruits and vegetables, secondary metabolites show beneficial effects on human health [
71]. They have well-established anticancer, antiaging, anti-diabetic, and anti-obesity activity, in addition to their protection against Alzheimer’s and cardiovascular diseases [
70]. Synthesized through the shikimate biochemical pathway, phenolic compounds represent the most abundant type of secondary metabolites in plants [
72]. Their biosynthesis begins from non-oxidative deamination of phenylalanine, mediated by the enzyme phenylalanine ammonia-lyase (PAL), leading to the formation of cinnamic trans acid as a key intermediate at the base of secondary products derived from phenylpropanoid (flavonoids and isoflavonoids, coumarins, lignins, esters of hydroxycinnamic acid, and phenolic compounds) [
72]. The accumulation of these compounds varies between organisms, tissues, and growth stage, and can be influenced by environmental conditions, because gene expression levels that encode key enzymes in the phenylpropanoid biosynthesis pathway are affected by environmental stressors (light, temperature, and nutritional deficits) [
73].
The UHPLC analysis identified 20 phenolic compounds that could be classified into the following categories: phenolic acid derivatives, flavonoid derivatives, and hydroxycinnamoyl quinic acid derivatives (
Table 7). The different light intensity conditions that characterized the treatments in the present experiment influenced the total content of phenolic compounds, calculated as the sum of all the detected individual phenolic compounds. Specifically, the Pearl grey shade net resulted in the highest accumulation of total phenolic compounds (14,997 µg 100 g
−1 fw), followed by the control (12,377 µg 100 g
−1 fw) and the White shade net (9869 µg 100 g
−1 fw). Regardless of the treatment, the largest contribution to total phenolic compounds resulted from flavonoid derivatives (7776 µg 100 g
−1 fw, on average), as reported by Bertin and Génard [
2], followed by phenolic acid derivatives (4097 µg 100 g
−1 fw, on average) and lastly hydroxycinnamoyl quinic acid derivatives (541 µg 100 g
−1 fw, on average). Although flavonoids are ‘semi-essential’ compounds, having no well-defined nutritional function, they are crucial for protecting antioxidant compounds from oxidative degradation in humans and plants [
74].
As reported by Slimestad and Verheul [
8] and Abreu et al. [
75], regardless of treatments, rutin was the most abundant flavonoid (
Table 7). In contrast, Bertin and Génard [
2] reported that the most abundant flavonoid in tomatoes was naringenin, demonstrating how genotype, cultural practices, environmental conditions, and even analytical determination techniques can influence the content of these compounds. Rutin is considered one of the best natural antioxidants currently known that can exert important pharmacological activities, acting as antibacterial, anti-inflammatory, antiallergic, antiviral, antiprotozoal, and antitumor. It has also marked cytoprotective, vasoactive, antiplatelet, hypolipidemic, and antihypertensive activities [
76]. The highest rutin values were obtained using a Pearl grey shading net (4414 µg 100 g
−1 fw), similarly to the other flavonoid derivatives (kampferol-3-diglucoside > naringenin > rutin-O-pentoside > kaempferol-3-O-rutinoside > naringenin-C-diglycoside > apigenin-C-hexoside-hexoside > naringenin-C-hexoside > quercetin-O-dihexoside > genistin), except for rutin-O-hexoside, which showed the highest value under the White shading net and control (
Table 7). The reviewed literature shows that the increase in flavonoids (mainly rutin) is promoted by intense solar radiation [
77,
78], which is not in line with the results obtained in this experiment. The total flavonoid content of fruits grown under pearl grey net was 59.9% higher than that recorded in the unshaded control (
Table 7). The reason behind this can be partly explained by the fact that most of the reviewed works did not separate the effect of temperature from that of solar radiation, while in our study, the only significantly different parameter was PAR (
Table 1 and
Table 2). Furthermore, it should be considered that the response of the plant to a stressor depends not only on the genotype, intensity, and magnitude of the stressor, but also on the stage of development of the plant and the organs involved [
2]. For example, despite evidence in the literature that water stress can increase the levels of phenolic compounds, Atkinson et al. [
79] observed that the greatest accumulation of flavonoids was recorded in tomato leaves following water stress, while in fruits no changes were observed. Similarly, Abreu et al. [
75] showed, in line with our results, that shading increased the phenolic content in tomato fruits, compared to the control, that resulted in a greater accumulation of these compounds only in the leaves. This result was probably attributable to the fact that the leaves were directly exposed to the stressor (high direct solar radiation). Not least, the obtained up-regulation of flavonoids could result not only from the limitation of solar radiation recorded in August under the pearl grey net (
Table 1), but also from eco-physiological responses induced by the different microclimatic conditions that lead to the plant to modify its primary metabolism (fewer but larger fruits;
Table 3). As argued by Campa et al. [
80], low light intensity would have triggered the production of phenolic compounds with which plants would have counterbalanced the reduction in antioxidant enzyme activity. In contrast with flavonoids trend, the highest hydroxycinnamoyl quinic acid derivatives values were obtained under Pearl grey net (685 µg 100 g
−1 fw) and in the control (633 µg 100 g
−1 fw), while the lowest were obtained under White net (305 µg 100 g
−1 fw) (
Table 7). In particular, the values of tricaffeoylquinic and dicaffeoylquinic acids in the more shaded conditions (pearl grey) were 53.30 and 44.50% higher than the average of the other treatments.
In contrast to what was observed for flavonoid derivatives and hydroxycinnamoyl quinic acids, the content of phenolic acid derivatives was lower in fruits harvested under pearl grey shading net (
Table 7). As reported in the literature, chlorogenic acid is the main non-flavonoid phenolic compound found in tomato fruits that possess high antioxidant, antibacterial, anti-inflammatory, antiviral, antimicrobial, hepatoprotective, cardioprotective, and neuroprotective properties [
2,
8,
81,
82]. The highest value of this crucial antioxidant compound was found in the unshaded control (3363 µg 100 g
−1 fw) while the lowest was found in the Pearl grey net treatment (1,799 µg 100 g
−1 fw) (
Table 7). However, Botella et al. [
83] reported that homovanillic acid-O-hexoside was the primary compound in tomato fruits. In our case, homovanillic acid O-hexoside was found to be the second most prevalent compound, with the highest values (11.51 µg g
−1 fw) recorded in fruits grown under the white net. The lowest values of coumaric acid O-hexoside were found in the White net treatment, whereas the lowest ones of ferulic and caffeic acids were found in the control (
Table 7).
In contrast to what was observed with flavonoid and hydroxycinnamoyl quinic acid derivatives, the heterogeneity of phenolic acid derivatives among treatments emphasizes that irradiation and/or shading strongly influenced the biosynthesis of this class of phenolic compounds. Probably, different light conditions could have unequivocally influenced the assignment of phenolic substrates to individual branches of the phenylpropanoid pathway [
73].
Different spectrophotometric assays were carried out for the determination of antioxidant activity of tomato fruits. Specifically, we evaluated the free radical scavenging activity DPPH, the free radical scavenging activity by ABTS decolorization, and the ferric reducing antioxidant capacity FRAP (
Table 8). Our findings showed significant antioxidant activity in fruits grown under pearl grey net, probably related to the higher content of total phenolic compounds (
Table 7) [
75].
In detail, DPPH antioxidant activity increased as the degree of shading increased, with the highest (40.72 mmol Trolox eq. kg−1 dw) and lowest (32.21 mmol Trolox eq. kg−1 dw) values obtained in fruits shaded with the pearl grey net and in the control, respectively. On the contrary, the ABTS assay did not show the same trend, with the lowest value (35.33 mmol Trolox eq. kg−1 dw) obtained in fruits shaded with the white net and the highest value (43.70 mmol Trolox eq. kg−1 dw) obtained with the pearl grey net.
The FRAP activity showed the highest value (34.38 mmol Trolox eq. kg−1 dw) in fruits shaded with the pearl grey net, while no significant differences were observed between the unshaded control and the White net treatment.
Furthermore, the correlation coefficient between the content of total phenolic compounds and antioxidant activities was highly significant, especially for the FRAP (R
2 = 0.74) and ABTS (R
2 = 0.99) assays. The correlation coefficient between the DPPH assay and the total phenolic compounds (R
2 = 0.37) was less significant than that of the FRAP and ABTS assays (
Table 8). These discrepancies could be due to synergistic effects between phenolic compounds and other chemical components that can contribute to the total antioxidant activity or the type of assay used [
84,
85]. The lower value of DPPH found in control plants (
Table 8) could be attributed to the lower content of total carotenoids, especially lycopene (
Table 6).


 ,
, 



{kind=link}