
Physiological and Biochemical Dissection Reveals a Trade-Off between Antioxidant Capacity and Heat Tolerance in Bread Wheat (Triticum aestivum L.)

 ,
, 
 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Stress Treatment
2.2. Determination of Chlorophyll, Carotenoid and Anthocyanin Content
2.3. Leaf Temperature Depression (LTD)
2.4. Membrane Injury Index
2.5. Methylglyoxal
2.6. Membrane Lipid Peroxidation and H2O2 Content
2.7. Proline and Glycine Betaine
2.8. Ascorbate and Glutathione
2.9. Soluble Protein
2.10. Assays for Antioxidant and Glyoxalase Enzyme Activity
2.11. Statistical Analysis
3. Results
3.1. Effect of Variety and Heat Stress on the Studied Traits
3.2. Seedling Length and Dry Matter
3.3. Chlorophyll, Carotenoid and Anthocyanin Content
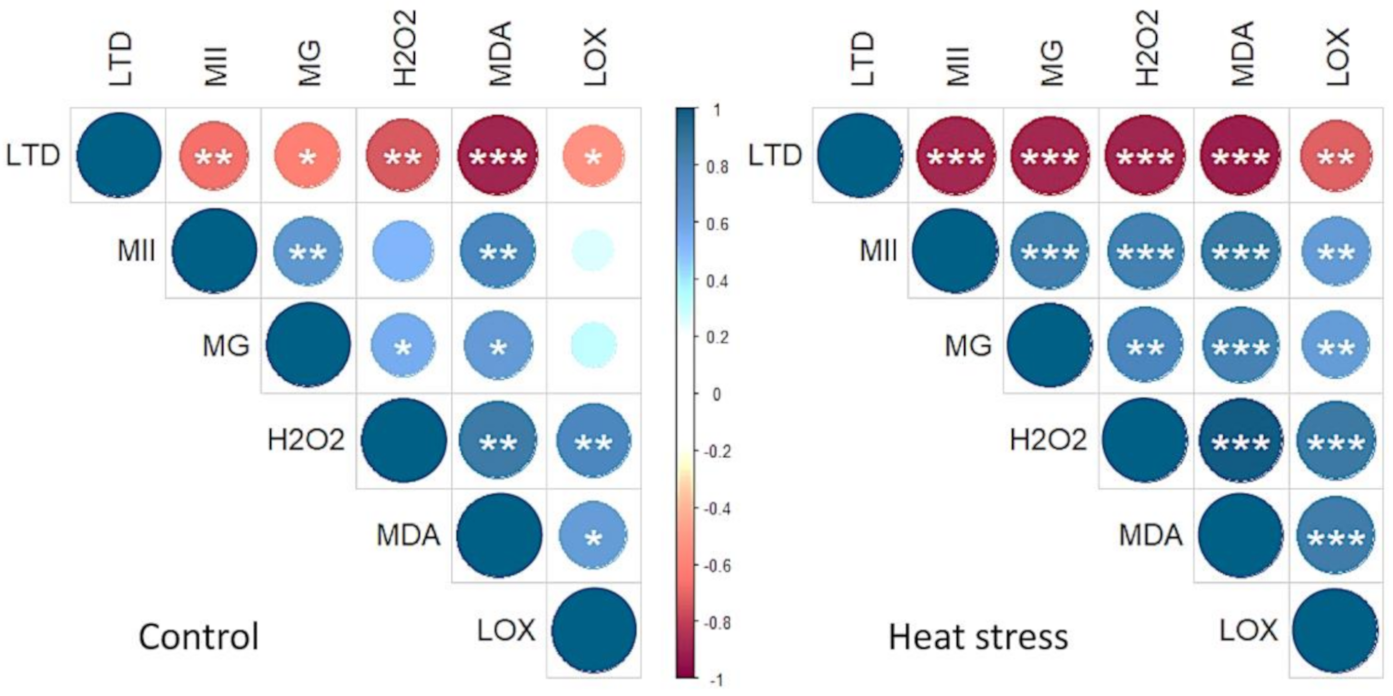
3.4. Leaf Temperature Depression and Membrane Injury Index
3.5. Membrane Lipid Peroxidation, LOX Activity and H2O2 Level
3.6. Methylglyoxal Level and Detoxifying Enzymes
3.7. Activity of Osmolytes and Non-Enzymatic Antioxidants
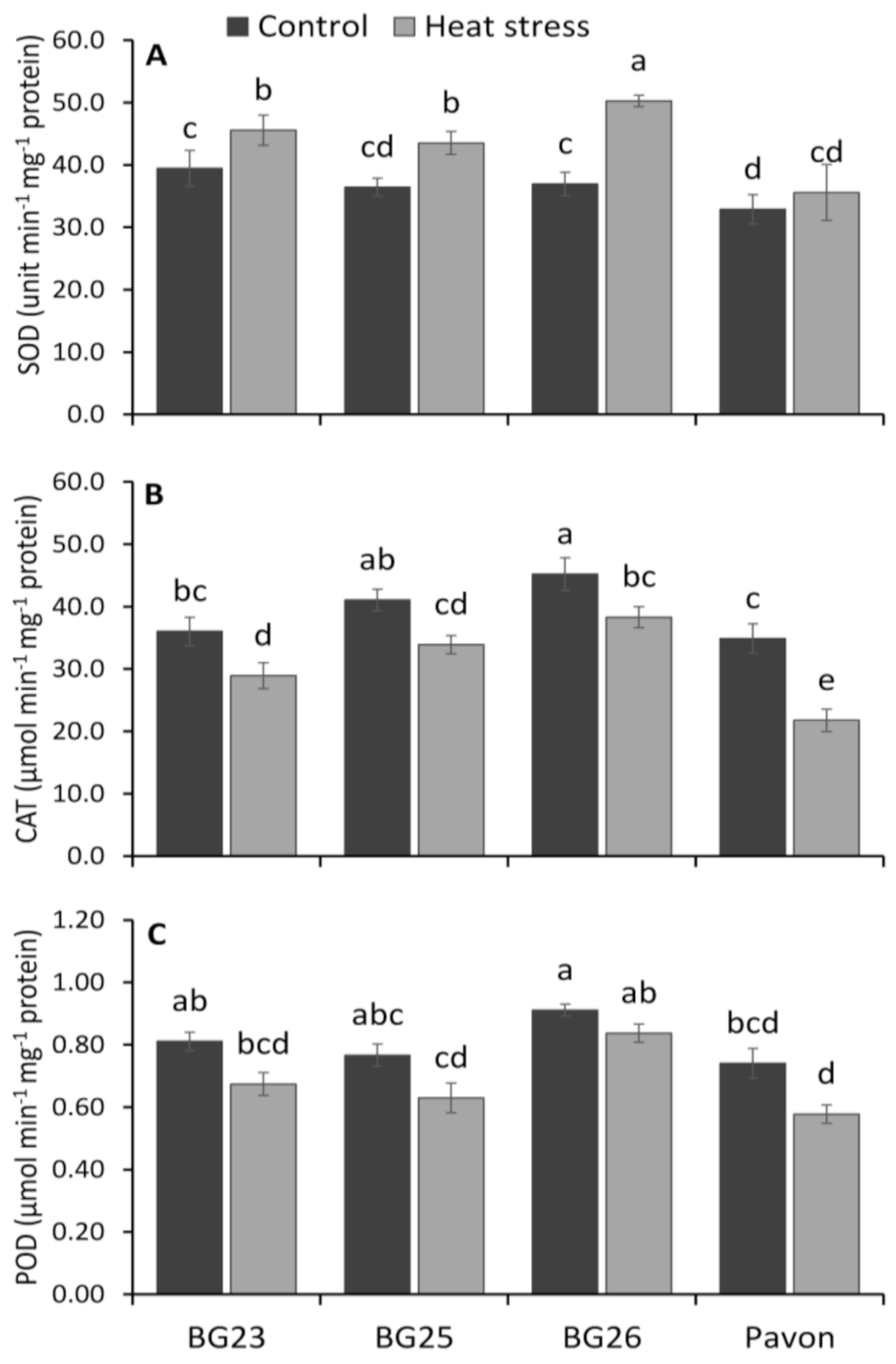
3.8. Reactive Oxigen Species (ROS) Scavenging Enzymes
3.9. AsA-GSH Cycle Enzymes
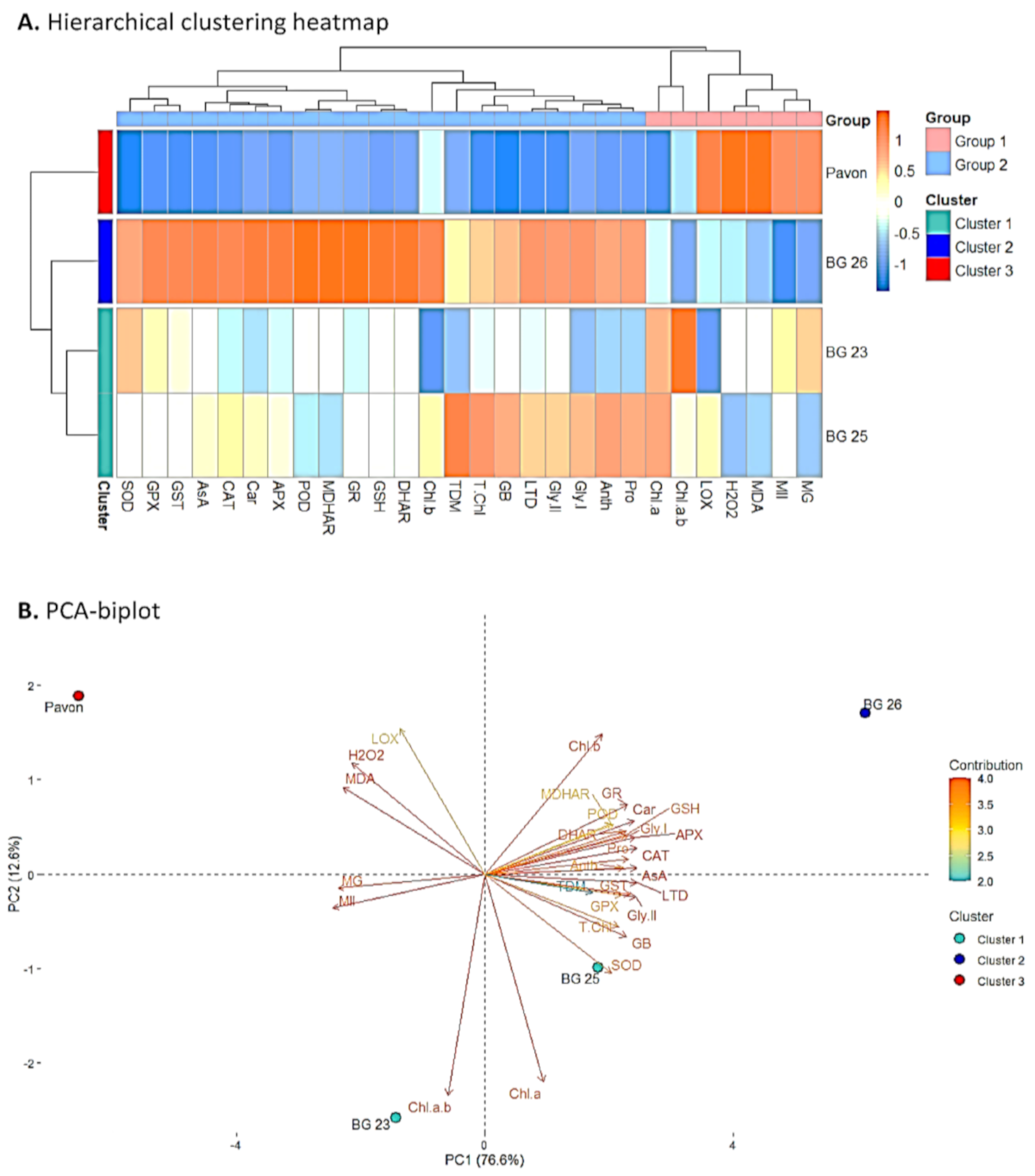
3.10. Assessment of the Association between Treatment, Varieties and Variables Using Hierarchical Clustering and Principal Component Analysis (PCA)
4. Discussion
4.1. Pigment Stability Confers Judicious Light Absorption under Heat Stress
4.2. Leaf Temperature Depression, Membrane Disruption and Cellular Toxicity Correlated with Oxidative Damage
4.3. Enhanced Osmolytes Accumulation Alleviate Heat-Induced Oxidative Stress in Wheat
4.4. Stable Antioxidant Enzyme System is Crucial for Heat Tolerance
4.5. Efficient Operation of Ascorbate-Glutathione Cycle is Pivotal for Heat Tolerance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations:
| AGE | Advanced glycation end product |
| Anth | Anthocyanins |
| AO | Ascorbate oxidase |
| APX | Ascorbate peroxidase |
| AsA | Ascorbate |
| BARI | Bangladesh Agricultural Research Institute |
| BSA | Bovine serum albumin |
| Car | Carotenoids |
| CAT | Catalase |
| CDNB | 1-chloro-2,4-dinitrobenzene |
| Chl | Chlorophyll |
| DHA | Dehydroascorbate |
| DHAR | Dehydroascorbate reductase |
| DTNB | 5,5′-dithiobis-(2-nitrobenzoic acid) |
| EDTA | Ethylenediaminetetraacetic acid |
| GB | Glycine betaine |
| Gly I | Glyoxalase-I |
| Gly II | Glyoxalase-II |
| GPX | Glutathione peroxidase |
| GR | Glutathione reductase |
| GSH | Glutathione (reduced) |
| GSSG | Glutathione (oxidized) |
| GST | Glutathione s-transferase |
| IPCC | The Intergovernmental Panel on Climate Change |
| LOX | Lipoxygenase |
| LTD | Leaf temperature depression |
| MDA | Malondialdehyde |
| MDHA | Monodehydroascorbate |
| MDHAR | Monodehydroascorbate reductase |
| MG | Methylglyoxal |
| MII | Membrane injury index |
| NADPH | Nicotinamide adenine dinucleotide phosphate (reduced) |
| NBT | Nitroblue tetrazolium |
| POD | Guaiacol peroxidases |
| Pro | Proline |
| PUFA | Polyunsaturated fatty acid |
| RDM | Root dry matter |
| RH | Relative humidity |
| RL | Root length |
| ROS | Reactive oxygen species |
| SDM | Shoot dry matter |
| SL | Shoot length |
| SLG | S-D-lactoylglutathione |
| SOD | Superoxide dismutase |
| STI | Stress tolerance index |
| TBA | Thiobarbituric acid |
| TCA | Trichloroacetic acid |
| TDM | Total dry matter |
References
- Paradiso, A.; Domingo, G.; Blanco, E.; Buscaglia, A.; Fortunato, S.; Marsoni, M.; Scarcia, P.; Caretto, S.; Vannini, C.; de Pinto, M.C. Cyclic AMP mediates heat stress response by the control of redox homeostasis and ubiquitin-proteasome system. Plant Cell Environ. 2020, 43, 2727–2742. [Google Scholar] [CrossRef]
- IPCC. Summary for policymakers. In Global Warming of 1.5 °C.; Masson-Delmotte, V., Zhai, P., Pörtner, H.O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; World Meteorological Organization: Geneva, Switzerland, 2014; p. 32. [Google Scholar]
- Aiqing, S.; Somayanda, I.; Sebastian, S.V.; Singh, K.; Gill, K.; Prasad, P.V.; Jagadish, S.V.K. Heat stress during flowering affects time of day of flowering, seed set, and grain quality in spring wheat. Crop Sci. 2018, 58, 380–392. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Al Mahmud, J.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asseng, S.; Ewert, F.; Martre, P.; Rötter, R.P.; Lobell, D.B.; Cammarano, D.; Kimball, B.A.; Ottman, M.J.; Wall, G.W.; White, J.W.; et al. Rising temperatures reduce global wheat production. Nat. Clim. 2015, 5, 143–147. [Google Scholar] [CrossRef]
- Tack, J.; Barkley, A.; Nalley, L.L. Effect of warming temperatures on US wheat yields. Proc. Natl. Acad. Sci. USA 2015, 112, 6931–6936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zandalinas, S.I.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A.; Inupakutika, M.A.; Mittler, R. ABA is required for the accumulation of APX1 and MBF1c during a combination of water deficit and heat stress. J. Exp. Bot. 2016, 67, 5381–5390. [Google Scholar] [CrossRef] [Green Version]
- Prasad, P.V.V.; Bheemanahalli, R.; Jagadish, S.V.K. Field crops and the fear of heat stress: Opportunities, challenges and future directions. Field Crop. Res. 2017, 200, 114–121. [Google Scholar] [CrossRef] [Green Version]
- Jagadish, S.V.K. Heat stress during flowering in cereals—Effects and adaptation strategies. New Phytol. 2020, 226, 1567–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mufti, M.U. Screening Wheat Seedlings for Heat and Drought Tolerance by In Vivo Chlorophyll Fluorescence. Master’s Thesis, The University of Adelaide, Adelaide, Australia, 2005. Available online: http://hdl.handle.net/2440/22299 (accessed on 13 February 2021).
- Barma, N.C.D.; Hossain, A.; Hakim, M.A.; Mottaleb, K.A.; Alam, M.A.; Reza, M.M.A.; Rohman, M.M. Progress and Challenges of Wheat Production in the Era of Climate Change: A Bangladesh Perspective. In Wheat Production in Changing Environments; Hasanuzzaman, M., Nahar, K., Hossain, M.A., Eds.; Springer Nature: Singapore, 2019; p. 633. [Google Scholar] [CrossRef]
- Farhad, M.; Tripathi, S.B.; Singh, R.P.; Joshi, A.K.; Bhati, P.K.; Mondal, S.; Kumar, U. Phenological variations in early planting of wheat breeding lines and identification of markers by GWAS. In Proceedings of the 2020 BGRI Technical Workshop (Virtual), Itacha, NY, USA, 7–9 October 2020; Available online: https://bgri.cornell.edu/wp-content/uploads/2020/10/poster-farhad.pdf (accessed on 18 February 2021).
- Kumar, S.; Kumari, P.; Kumar, U.; Grover, M.; Singh, A.K.; Singh, R.; Sengar, R.S. Molecular approaches for designing heat tolerant wheat. J. Plant Biochem. Biotechnol. 2013, 22, 359–371. [Google Scholar] [CrossRef]
- Öztürk, A.; Taşkesenligil, B.; Haliloğlu, K.; Aydin, M.; Çağlar, Ö. Evaluation of bread wheat genotypes for early drought resistance via germination under osmotic stress, cell membrane damage, and paraquat tolerance. Turk. J. Agric. For. 2016, 40, 146–159. [Google Scholar] [CrossRef]
- ElBasyoni, I.; Saadalla, M.; Baenziger, S.; Bockelman, H.; Morsy, S. Cell membrane stability and association mapping for drought and heat tolerance in a worldwide wheat collection. Sustainability 2017, 9, 1606. [Google Scholar] [CrossRef] [Green Version]
- Almeselmani, M.; Deshmukh, P.S.; Sairam, R.K. High temperature stress tolerance in wheat genotypes: Role of antioxidant defense enzymes. Acta Agron. Hung. 2009, 57, 1–14. [Google Scholar] [CrossRef]
- Rehman, S.U.; Bilal, M.; Rana, R.M.; Tahir, M.N.; Shah, M.K.N.; Ayalew, H.; Yan, G. Cell membrane stability and chlorophyll content variation in wheat (Triticum aestivum) genotypes under conditions of heat and drought. Crop Pasture Sci. 2016, 67, 712–718. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Nitric oxide modulates antioxidant defense and the methylglyoxal detoxification system and reduces salinity-induced damage of wheat seedlings. Plant Biotechnol. Rep. 2011, 5, 353–365. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Selenium-induced up-regulation of the antioxidant defense and methylglyoxal detoxification system reduces salinity-induced damage in rapeseed seedlings. Biol. Trace Elem. Res. 2011, 143, 1704–1721. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Fujita, M. Selenium pretreatment upregulates the antioxidant defense and methylglyoxal detoxification system and confers enhanced tolerance to drought stress in rapeseed seedlings. Biol. Trace Elem. Res. 2011, 143, 1758–1776. [Google Scholar] [CrossRef]
- Yadav, S.K.; Singla-Pareek, S.L.; Ray, M.; Reddy, M.K.; Sopory, S.K. Methylglyoxal levels in plants under salinity stress are dependent on glyoxalase I and glutathione. Biochem. Biophys. Res. Commun. 2005, 337, 61–67. [Google Scholar] [CrossRef]
- Kaur, C.; Singla-Pareek, S.L.; Sopory, S.K. Glyoxalase and methylglyoxal as biomarkers for plant stress tolerance. Crit. Rev. Plant Sci. 2014, 33, 429–456. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Ghosh, A.; Li, Z.G.; Siddiqui, M.N.; Fujita, M.; Tran, L.S.P. Methylglyoxal—A signaling molecule in plant abiotic stress responses. Free Radic. Biol. Med. 2018, 122, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Hoque, M.A.; Uraji, M.; Banu, M.N.A.; Mori, I.C.; Nakamura, Y.; Murata, Y. The effects of methylglyoxal on glutathione S-transferase from Nicotiana tabacum. Biosci. Biotechnol. Biochem. 2010, 74, 2124–2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.A.; Hasanuzzaman, M.; Fujita, M. Up-regulation of antioxidant and glyoxalase systems by exogenous glycinebetaine and proline in mung bean confer tolerance to cadmium stress. Physiol. Mol. Biol. Plants 2010, 16, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.K.; Singla-Pareek, S.L.; Reddy, M.K.; Sopory, S.K. Methylglyoxal detoxification by glyoxalase system: A survival strategy during environmental stresses. Physiol. Mol. Biol. Plants 2005, 11, 1–11. [Google Scholar]
- Yadav, S.K.; Singla-Pareek, S.L.; Reddy, M.K.; Sopory, S.K. Transgenic tobacco plants overexpressing glyoxalase enzymes resist an increase in methylglyoxal and maintain higher reduced glutathione levels under salinity stress. FEBS Lett. 2005, 579, 6265–6271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.S.; Gomes, R.A.; Ferreira, A.E.N.; Freire, A.P.; Cordeiro, C. The glyoxalase pathway: The first hundred years… and beyond. Biochem. J. 2013, 453, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, H.; Chen, Q.M.; Yi, M.F. Effects of short-term heat stress on oxidative damage and responses of antioxidant system in Lilium longiflorum. Plant Growth Regul. 2008, 54, 45–54. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Li, J.; Zhang, X.; Wei, H.; Cui, L. Effects of heat acclimation pretreatment on changes of membrane lipid peroxidation, antioxidant metabolites, and ultrastructure of chloroplasts in two cool-season turfgrass species under heat stress. Environ. Exp. Bot. 2006, 56, 274–285. [Google Scholar] [CrossRef]
- Rio, D.D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [PubMed]
- Yamauchi, Y.; Furutera, A.; Sugimoto, Y. Malondialdehyde generated from peroxidized linolenic acid causes protein modification in heat-stressed plants. Plant Physiol. Biochem. 2008, 46, 786–793. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Kong, L.; Wu, Z.; Rosenqvist, E.; Wang, Y.; Zhao, L.; Zhao, T.; Ottosen, C.O. Physiological response of tomatoes at drought, heat and their combination followed by recovery. Physiol. Plant. 2019, 165, 144–154. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Foyer, C.H. The roles of reactive oxygen metabolism in drought: Not so cut and dried. Plant Physiol. 2014, 164, 1636–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Exogenous silicon attenuates cadmium-induced oxidative stress in Brassica napus L. by modulating AsA-GSH pathway and glyoxalase system. Front. Plant Sci. 2017, 8, 1061. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef]
- Eggersdorfer, M.; Wyss, A. Carotenoids in human nutrition and health. Arch. Biochem. Biophys. 2018, 652, 18–26. [Google Scholar] [CrossRef]
- Savicka, M.; Škute, N. Effects of high temperature on malondialdehyde content, superoxide production and growth changes in wheat seedlings (Triticum aestivum L.). Ekologija 2010, 56, 26–33. [Google Scholar] [CrossRef]
- Hossain, A.; Teixeira da Silva, J.A. Wheat production in Bangladesh: Its future in the light of global warming. AoB Plants 2013, 5, pls042. [Google Scholar] [CrossRef] [Green Version]
- Khatun, S.; Ahmed, J.U.; Mohi-Ud-Din, M. Variation of wheat cultivars in their relationship between seed reserve utilization and leaf temperature under elevated temperature. J. Crop Sci. Biotechnol. 2015, 18, 97–101. [Google Scholar] [CrossRef]
- Arnon, D. Copper enzymes isolated chloroplasts, polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Lachman, J.; Hamouz, K.; Hejtmankova, A.; Dudjak, J.; Orsak, M.; Pivec, V. Effect of white fleece on the selected quality parameters of potato (Solanum tuberosum L.) tubers. Plant Soil Environ. 2003, 49, 370–377. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Mukta, J.A.; Sabir, A.A.; Gupta, D.R.; Mohi-Ud-Din, M.; Hasanuzzaman, M.; Miah, M.G.; Rahman, M.; Islam, M.T. Chitosan biopolymer promotes yield and stimulates accumulation of antioxidants in strawberry fruit. PLoS ONE 2018, 13, e0203769. [Google Scholar]
- Hughes, N.M.; Smith, W.K. Attenuation of incident light in Galaxurceolata (Diapensiaceae): Concerted influence of adaxial and abaxial anthocyanic layers on photoprotection. Am. J. Bot. 2007, 94, 784–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sairam, R.K.; Deshmukh, P.S.; Shukla, D.S. Tolerance of drought and temperature stress in relation to increased antioxidant enzyme activity in wheat. J. Agron. Crop Sci. 1997, 178, 171–178. [Google Scholar] [CrossRef]
- Fischer, R.A.; Rees, D.; Sayre, K.D.; Lu, Z.M.; Condon, A.G.; Saavedra, L.A. Wheat yield progress associated with higher stomatal conductance and photosynthetic rate, and cooler canopies. Crop Sci. 1998, 38, 1467–1475. [Google Scholar] [CrossRef]
- Deshmukh, P.S.; Sairam, R.K.; Shukla, D.S. Measurement of ion leakage as a screening technique for drought resistance in wheat genotypes. Indian J. Plant Physiol. 1991, 34, 89–91. [Google Scholar]
- Wild, R.; Ooi, L.; Srikanth, V.; Munch, G. A quick, convenient and economical method for the reliable determination of methylglyoxal in millimolar concentrations: The N-acetyl-L-cysteine assay. Anal. Bioanal. Chem. 2012, 403, 2577–2581. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplast. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Yu, C.W.; Murphy, T.M.; Lin, C.H. Hydrogen peroxide-induces chilling tolerance in mung beans mediated through ABA independent glutathione accumulation. Funct. Plant Biol. 2003, 30, 955–963. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teari, D. Rapid determination of free proline for water stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Valadez-Bustos, M.G.; Aguado-Santacruz, G.A.; Tiessen-Favier, A.; Robledo-Paz, A.; Munoz-Orozco, A.; Rascon-Cruz, Q.; Santacruz-Varela, A. A reliable method for spectrophotometric determination of glycine betaine in cell suspension and other systems. Anal. Biochem. 2016, 498, 47–52. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Hasanuzzaman, M.; Nahar, K.; Fujita, M. Exogenous proline and glycine betaine mediated upregulation of antioxidant defense and glyoxalase systems provides better protection against salt-induced oxidative stress in two rice (Oryza sativa L.) varieties. Biomed. Res. Int. 2014, 2014, 757219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and susceptible method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Spitz, D.R.; Oberley, L.W. An assay for superoxide dismutase activity in mammalian tissue homogenates. Annal. Biochem. 1989, 179, 8–18. [Google Scholar] [CrossRef]
- Doderer, A.; Kokkelink, I.; van der Veen, S.; Valk, B.; Schram, A.; Douma, A. Purification and characterization of two lipoxygenase isoenzymes from germinating barley. Biochim. Biophys. Acta 1992, 112, 97–104. [Google Scholar] [CrossRef]
- Castillo, F.I.; Penel, I.; Greppin, H. Peroxidase release induced by ozone in sedum album leaves. Plant Physiol. 1984, 74, 846–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elia, A.C.; Galarini, R.; Taticchi, M.I.; Dörr, A.J.; Mantilacci, L. Antioxidant responses and bioaccumulation in Ictalurus melas under mercury exposure. Ecotoxicol. Environ. Saf. 2003, 55, 162–167. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H.; Dicky, D.A. Principles and Procedures of Statistics, A Biometrical Approach, 3rd ed.; McGraw Hill Inc.: New York, NY, USA, 1997; pp. 352–358. [Google Scholar]
- Fernandez, G.C.J. Effective selection criteria for assessing plant stress tolerance. In Proceedings of the International Symposium on Adaptation of Vegetables and Other Food Crops to Temperature Water Stress, Shanhua, Taiwan, 13‒16 August 1992; Food and Agriculture Organization: Shanhua, Taiwan, 1992; pp. 257–270. [Google Scholar]
- Kolde, R. pheatmap: Pretty Heatmaps. R Package Version 1.0.12. 2012. Available online: https://rdrr.io/cran/pheatmap/ (accessed on 13 February 2021).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Asada, K. Production of active oxygen species in photosynthetic tissue. In Causes of Photooxidative Stress and Amelioration of Defense Systems in Plants; Foyer, C.H., Mullineaux, P.M., Eds.; CRC Press: Boca Roton, FL, USA, 1994; pp. 77–104. [Google Scholar]
- Singh, A.K.; Singhal, G.S. Effect of irradiance on the thermal stability of thylakoid membrane isolated from acclimated wheat leaves. Photosynthetica 2001, 39, 23–27. [Google Scholar] [CrossRef]
- Efeoglu, B.; Terzioglu, S. Photosynthetic responses of two wheat varieties to high temperature. Eur. Asian J. Biosci. 2009, 3, 97–106. [Google Scholar] [CrossRef]
- Aien, A.; Khetarpal, S.; Pal, M. Photosynthetic characteristics of potato cultivars grown under high temperature. Am. Eurasian J. Agric. Environ. Sci. 2011, 11, 633–639. [Google Scholar]
- Reda, F.; Mandoura, H.M.H. Response of enzymes activities, photosynthetic pigments, proline to low or high temperature stressed wheat plant (Triticum aestivum L.) in the presence or absence of exogenous proline or cysteine. Int. J. Acad. Res. 2011, 3, 108–115. [Google Scholar]
- Almeselmani, M.; Deshmukh, P.S.; Sairam, R.K.; Kushwaha, S.R.; Singh, T.P. Protective role of antioxidant enzymes under high temperature stress. Plant Sci. 2006, 171, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Yüzbaşioğlu, E.; Dalyan, E.; Akpinar, I. Changes in photosynthetic pigments, anthocyanin content and antioxidant enzyme activities of maize (Zea mays L.) seedlings under high temperature stress conditions. Trak. Univ. J. Nat. Sci. 2017, 18, 97–104. [Google Scholar]
- Mishra, V.; Srivastava, G.; Prasad, S.M.; Abraham, G. Growth, photosynthetic pigments and photosynthetic activity during seedling stage of cowpea (Vigna unguiculata) in response to UV-B and dimethoate. Pestic. Biochem. Phys. 2008, 92, 30–37. [Google Scholar] [CrossRef]
- Shao, L.; Shu, Z.; Sun, S.L.; Peng, C.L.; Wang, X.J.; Lin, Z.F. Antioxidation of anthocyanins in photosynthesis under high temperature stress. J. Integr. Plant Biol. 2007, 49, 1341–1351. [Google Scholar] [CrossRef]
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamental sand production of heat stress-tolerant crops. Front. Plant Sci. 2013, 273, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, M.P.; Ortiz-Monasterio, J.I.; McNab, A. Application of Physiology in Wheat Breeding; CIMMYT: El Batan, Mexico, 2001. [Google Scholar]
- Balla, K.; Karsai, I.; Bónis, P.; Kiss, T.; Berki, Z.; Horváth, Á.; Mayer, M.; Bencze, S.; Veisz, O. Heat stress responses in a large set of winter wheat cultivars (Triticum aestivum L.) depend on the timing and duration of stress. PLoS ONE 2019, 14, e0222639. [Google Scholar] [CrossRef]
- Deva, C.R.; Urban, M.O.; Challinor, A.J.; Falloon, P.; Svitákova, L. Enhanced leaf cooling is a pathway to heat tolerance in common bean. Front. Plant Sci. 2020, 11, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajji, M.; Kinet, J.M.; Lutts, S. The use of electrolyte leakage method for assessing cell membrane stability as water stress tolerance test in durum wheat. Plant Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Hemantaranjan, A. Heat stress responses and thermotolerance. Adv. Plants Agric. Res. 2014, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Pastori, G.M.; Trippi, V.S. Oxidative stress induces high rate of glutathione reductase synthesis in a drought resistant maize strain. Plant Cell Physiol. 1992, 33, 957–961. [Google Scholar]
- Mishra, A.; Jha, B.; Dubey, R.S. Arsenite treatment induces oxidative stress, upregulates antioxidant system, and causes phytochelatin synthesis in rice seedlings. Protoplasma 2011, 248, 565–577. [Google Scholar] [CrossRef]
- Alché, J.D. A concise appraisal of lipid oxidation and lipoxidation in higher plants. Redox Biol. 2019, 23, 101136. [Google Scholar] [CrossRef]
- Garg, N.; Manchanda, G. ROS generation in plants: Boon or bane? Plant Biosyst. 2009, 143, 81–96. [Google Scholar] [CrossRef]
- Lim, C.W.; Han, S.W.; Hwang, I.S.; Kim, D.S.; Hwang, B.K.; Lee, S.C. The pepper lipoxygenase Ca LOX1 plays a role in osmotic, drought and high salinity stress response. Plant Cell Physiol. 2015, 56, 930–942. [Google Scholar] [CrossRef] [Green Version]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majláth, I.; Szalai, G.; Soós, V.; Sebestyén, E.; Balázs, E.; Vanková, R.; Dobrev, P.I.; Tari, I.; Tandori, J.; Janda, T. Effect of light on the gene expression and hormonal status of winter and spring wheat plants during cold hardening. Physiol. Plant. 2012, 145, 296–314. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, W.; Wang, W.; Zhang, G.; Liu, Y.; Wang, Y.; Wang, W. Wheat F-box protein gene TaFBA1 is involved in plant tolerance to heat stress. Front. Plant Sci. 2018, 9, 521. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.U.; Din, J.U.; Qayyum, A.; Jan, N.E.; Jenks, M.A. Heat tolerance indicators in Pakistani wheat (Triticum aestivum L.) genotypes. Acta Bot. Croat. 2015, 74, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, N.; Umar, S.; Khan, N.A.; Corpas, F.J. Nitric Oxide and Hydrogen Sulfide Coordinately Reduce Glucose Sensitivity and Decrease Oxidative Stress via Ascorbate-Glutathione Cycle in Heat-Stressed Wheat (Triticum aestivum L.) Plants. Antioxidants 2021, 10, 108. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.P.; Zhang, X.Y.; Li, F.; Luo, Y.; Wang, W. Overaccumulation of glycine betaine enhances tolerance to drought and heat stress in wheat leaves in the protection of photosynthesis. Photosynthetica 2010, 48, 117–126. [Google Scholar] [CrossRef]
- Sarafraz-Ardakani, M.R.; Khavari-Nejad, R.A.; Moradi, F.; Najafi, F. Abscisic acid and cytokinin-induced osmotic and antioxidant regulation in two drought-tolerant and drought-sensitive cultivars of wheat during grain filling under water deficit in field conditions. Not. Sci. Biol. 2014, 6, 354–362. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.K.; Agarwal, S.; Agarwal, V.P.; Nathawat, N.S.; Gupta, S.; Singh, G. Effect of short-term heat stress on growth, physiology and antioxidative defense system in wheat seedlings. Acta Physiol. Plant. 2013, 35, 1837–1842. [Google Scholar] [CrossRef]
- Sharma, P.; Mehta, G.; Muthusamy, S.K.; Singh, S.K.; Singh, G.P. Development and validation of heat-responsive candidate gene and miRNA gene based SSR markers to analysis genetic diversity in wheat for heat tolerance breeding. Mol. Biol. Rep. 2021, 48, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Huang, B. Effects of calcium on antioxidant activities and water relations associated with heat tolerance in two cool-season grasses. J. Exp. Bot. 2001, 52, 341–349. [Google Scholar] [CrossRef]
- Anjali, A.; Shantha, N.; Pathak, P.C. Effect of high temperature on hydrogen peroxide scavenging enzymes during reproductive phase in aromatic rice cultivars. Indian J. Plant Physiol. 2006, 11, 427–431. [Google Scholar]
- Lu, P.; Sang, W.G.; Ma, K.P. Activity of stress-related antioxidative enzymes in the invasive plant crofton weed (Eupatorium adenophorum). J. Integr. Plant Biol. 2007, 49, 1555–1564. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Yang, Y.M.; Cao, L.; Hao, Y.F.; Huang, J.; Li, J.P.; Yao, D.X.; Wang, Z.M. Effect of high temperature on photosynthetic capability and antioxidant enzyme activity of flag leaf and non-leaf organs in wheat. Acta Agron. Sin. 2015, 41, 136–144. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef] [Green Version]
- Noctor, G.; Gomez, L.; Vanacker, H.; Foyer, C.H. Interactions between biosynthesis, compartmentation and transport in the control of glutathione homeostasis and signaling. J. Exp. Bot. 2002, 53, 1283–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kühn, H.; Borchert, A. Regulation of enzymatic lipid peroxidation: The interplay of peroxidizing and peroxide reducing enzymes. Free Radic. Biol. Med. 2002, 33, 154–172. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Fujita, M. Exogenous nitric oxide alleviates high temperature induced oxidative stress in wheat (Triticum aestivum L.) seedlings by modulating the antioxidant defense and glyoxalase system. Aust. J. Crop Sci. 2012, 6, 1314–1323. [Google Scholar]
- Dixon, D.P.; Skipsey, M.; Edwards, R. Roles for glutathione transferases in plant secondary metabolism. Phytochemistry 2010, 71, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, Y.K.; Yadav, S.K. Effect of high-temperature stress on ascorbate–glutathione cycle in maize. Agric. Res. 2019, 9, 179–187. [Google Scholar] [CrossRef]
- Kocsy, G.; Szalai, G.; Galiba, G. Effect of heat stress on glutathione biosynthesis in wheat. Acta Biol. Szeged. 2002, 46, 71–72. [Google Scholar]
- Pandey, S.; Fartyal, D.; Agarwal, A.; Shukla, T.; James, D.; Kaul, T.; Negi, Y.K.; Arora, S.; Reddy, M.K. Abiotic stress tolerance in plants: Myriad roles of ascorbate peroxidase. Front. Plant Sci. 2017, 8, 581. [Google Scholar] [CrossRef] [Green Version]
- Dash, S.; Mohanty, N. Response of seedlings to heat-stress in cultivars of wheat: Growth temperature-dependent differential modulation of photosystem 1 and 2 activity, and foliar antioxidant defense capacity. Plant Physiol. 2002, 159, 49–59. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivero, R.M.; Ruiz, J.M.; Romero, L. Oxidative metabolism in tomato plants subjected to heat stress. J. Hortic. Sci. Biotechnol. 2004, 79, 560–564. [Google Scholar] [CrossRef]
- Eltayeb, A.E.; Kawano, N.; Badawi, G.H.; Kaminaka, H.; Sanekata, T.; Morishima, I.; Shibahara, T.; Inanaga, S.; Tanaka, K. Enhanced tolerance to ozone and drought stresses in transgenic tobacco overexpressing dehydroascorbate reductase in cytosol. Physiol. Plant. 2006, 127, 57–65. [Google Scholar] [CrossRef]
- Eltayeb, A.L.; Kawano, N.; Badawi, G.H.; Kaminaka, H.; Sanekata, T.; Shibahara, T.; Inanaga, S.; Tanaka, K. Overexpression of monodehydroascorbate reductase in transgenic tobacco confers enhanced tolerance to ozone, salt and polyethylene glycol stresses. Planta 2007, 225, 1255–1264. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Growing Condition | Chl a (mg g−1 FW) | Chl b (mg g−1 FW) | Total Chl (mg g−1 FW) | Chl a/b Ratio | Carotenoid (mg g−1 FW) ǂ | Anthocyanin (µg g−1 FW)¶ |
|---|---|---|---|---|---|---|---|
| BG23 | Control | 0.49 ± 0.01 a,b | 0.26 ± 0.01 c,e | 0.75 ± 0.01 a–c | 1.88 ± 0.13 a | 0.09 ± 0.008 b–d | 71.51 ± 0.55 b |
| Heat stress | 0.42 ± 0.01 c | 0.23 ± 0.01 e | 0.65 ± 0.01 d,e | 1.83 ± 0.12 a,b | 0.07 ± 0.012 c–e | 55.73 ± 0.47 d | |
| (85.6 ± 1.45) | (88.8 ± 1.58) | (86.7 ±1.24) | (77.8 ± 5.88) | (78.0 ± 1.25) | |||
| BG25 | Control | 0.50 ± 0.02 a | 0.32 ± 0.02 a,b | 0.83 ± 0.02 a | 1.57 ± 0.12 a–c | 0.13 ± 0.015 a,b | 80.40 ± 0.67 a |
| Heat stress | 0.42 ± 0.01 c | 0.28 ± 0.01 b–e | 0.70 ± 0.02 c,d | 1.50 ± 0.01 b,c | 0.11 ± 0.012 b,c | 67.40 ± 0.50 c | |
| (83.5 ± 0.83) | (87.2 ± 5.61) | (84.7 ± 1.67) | (84.6 ± 1.04) | (83.9 ± 1.26) | |||
| BG26 | Control | 0.44 ± 0.01 b,c | 0.35 ± 0.01 a | 0.79 ± 0.01 a,b | 1.26 ± 0.03 c | 0.17 ± 0.009 a | 79.25 ± 0.83 a |
| Heat stress | 0.38 ± 0.01 c,d | 0.31 ± 0.01 a–c | 0.70 ± 0.01 c,d | 1.23 ± 0.08 c | 0.15 ± 0.014 a,b | 67.56 ± 0.59 c | |
| (87.2 ± 2.58) | (89.5 ± 2.61) | (88.2 ± 0.35) | (88.2 ± 8.48) | (85.3 ± 0.77) | |||
| Pavon | Control | 0.43 ± 0.01 b,c | 0.30 ± 0.01 a–d | 0.73 ± 0.02 b,c | 1.42 ± 0.05 c | 0.05 ± 0.012 d,e | 71.21 ± 0.74 b |
| Heat stress | 0.32 ± 0.02 d | 0.25 ± 0.01 d,e | 0.58 ± 0.02 e | 1.28 ± 0.10 c | 0.03 ± 0.009 e | 50.74 ± 0.65 e | |
| (75.3 ± 4.41) | (83.6 ± 4.88) | (78.6 ± 1.62) | (60.0 ± 8.45) | (71.3 ± 0.91) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohi-Ud-Din, M.; Siddiqui, N.; Rohman, M.; Jagadish, S.V.K.; Ahmed, J.U.; Hassan, M.M.; Hossain, A.; Islam, T. Physiological and Biochemical Dissection Reveals a Trade-Off between Antioxidant Capacity and Heat Tolerance in Bread Wheat (Triticum aestivum L.). Antioxidants 2021, 10, 351. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10030351
Mohi-Ud-Din M, Siddiqui N, Rohman M, Jagadish SVK, Ahmed JU, Hassan MM, Hossain A, Islam T. Physiological and Biochemical Dissection Reveals a Trade-Off between Antioxidant Capacity and Heat Tolerance in Bread Wheat (Triticum aestivum L.). Antioxidants. 2021; 10(3):351. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10030351
Chicago/Turabian StyleMohi-Ud-Din, Mohammed, Nurealam Siddiqui, Motiar Rohman, S. V. Krishna Jagadish, Jalal Uddin Ahmed, Mohamed M. Hassan, Akbar Hossain, and Tofazzal Islam. 2021. "Physiological and Biochemical Dissection Reveals a Trade-Off between Antioxidant Capacity and Heat Tolerance in Bread Wheat (Triticum aestivum L.)" Antioxidants 10, no. 3: 351. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10030351