
Exploiting the Freshwater Shrimp Neocaridina denticulata as Aquatic Invertebrate Model to Evaluate Nontargeted Pesticide Induced Toxicity by Investigating Physiologic and Biochemical Parameters

 , ,
, ,  ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
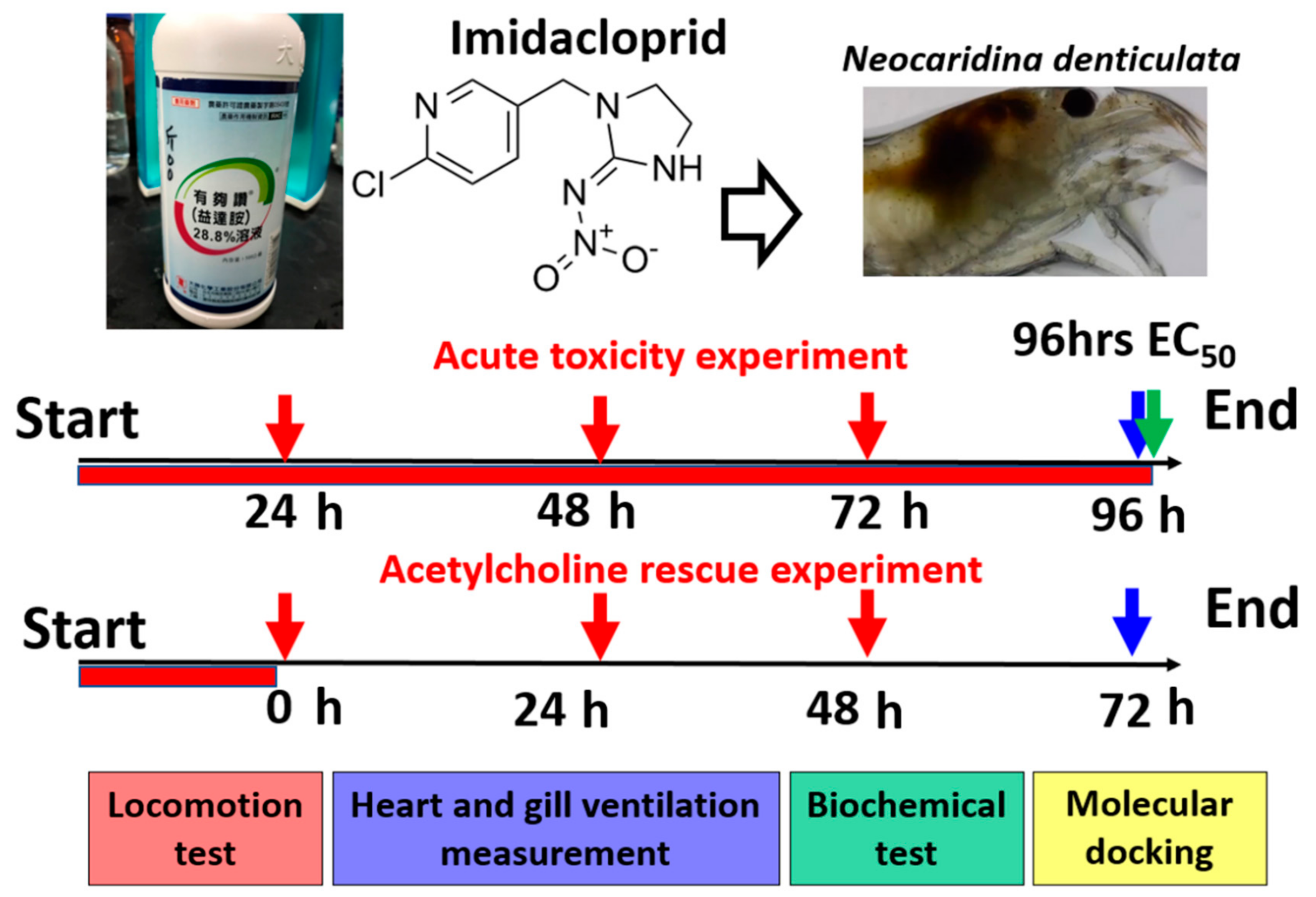
2.1. Overview of Experimental Design to Conduct Acute Toxicity Test in Shrimp
2.2. Animal Housing
2.3. Imidacloprid Treatment
2.4. EC50 Measurement for Immobilization
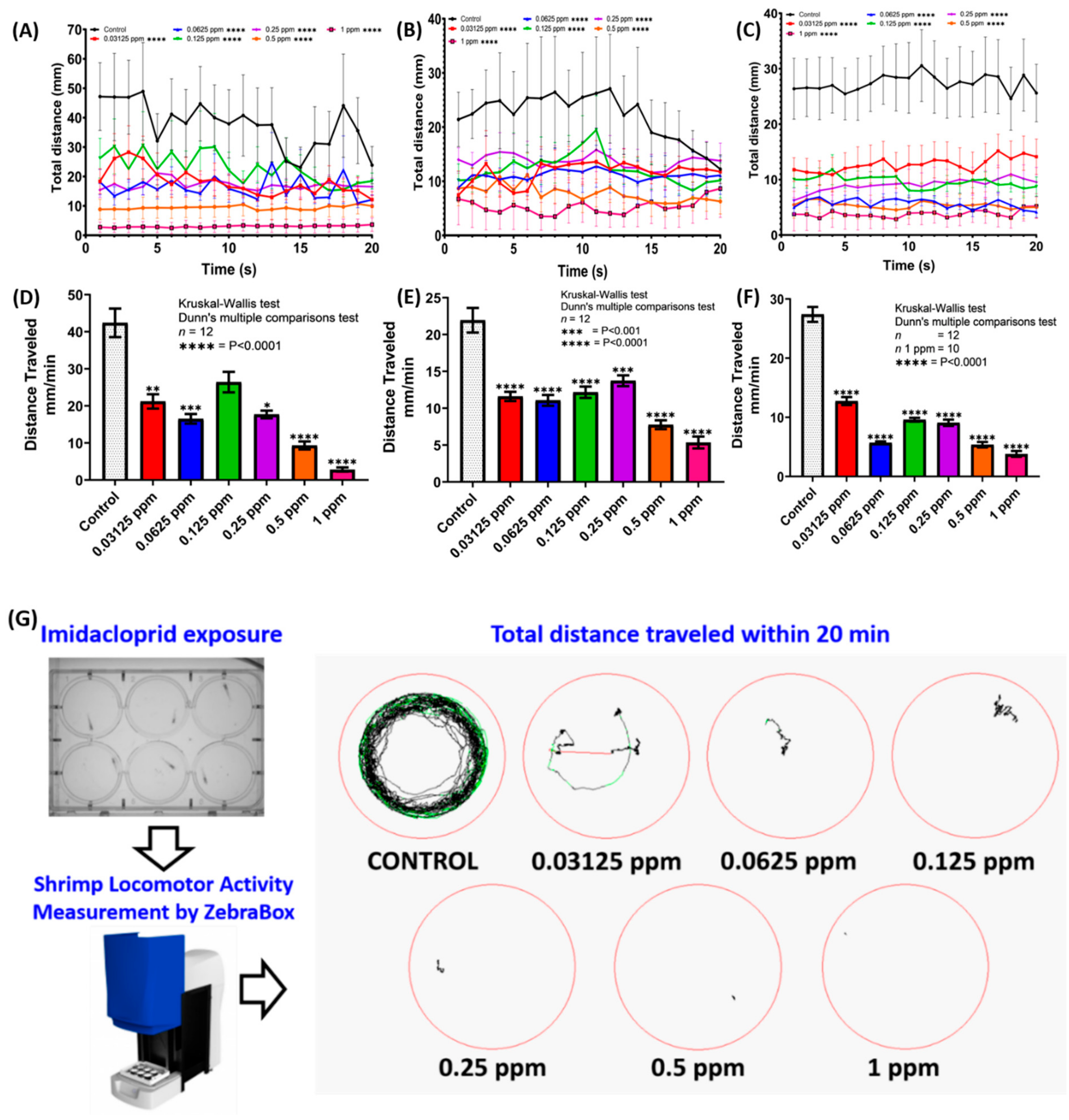
2.5. Locomotion Tracking and Quantification
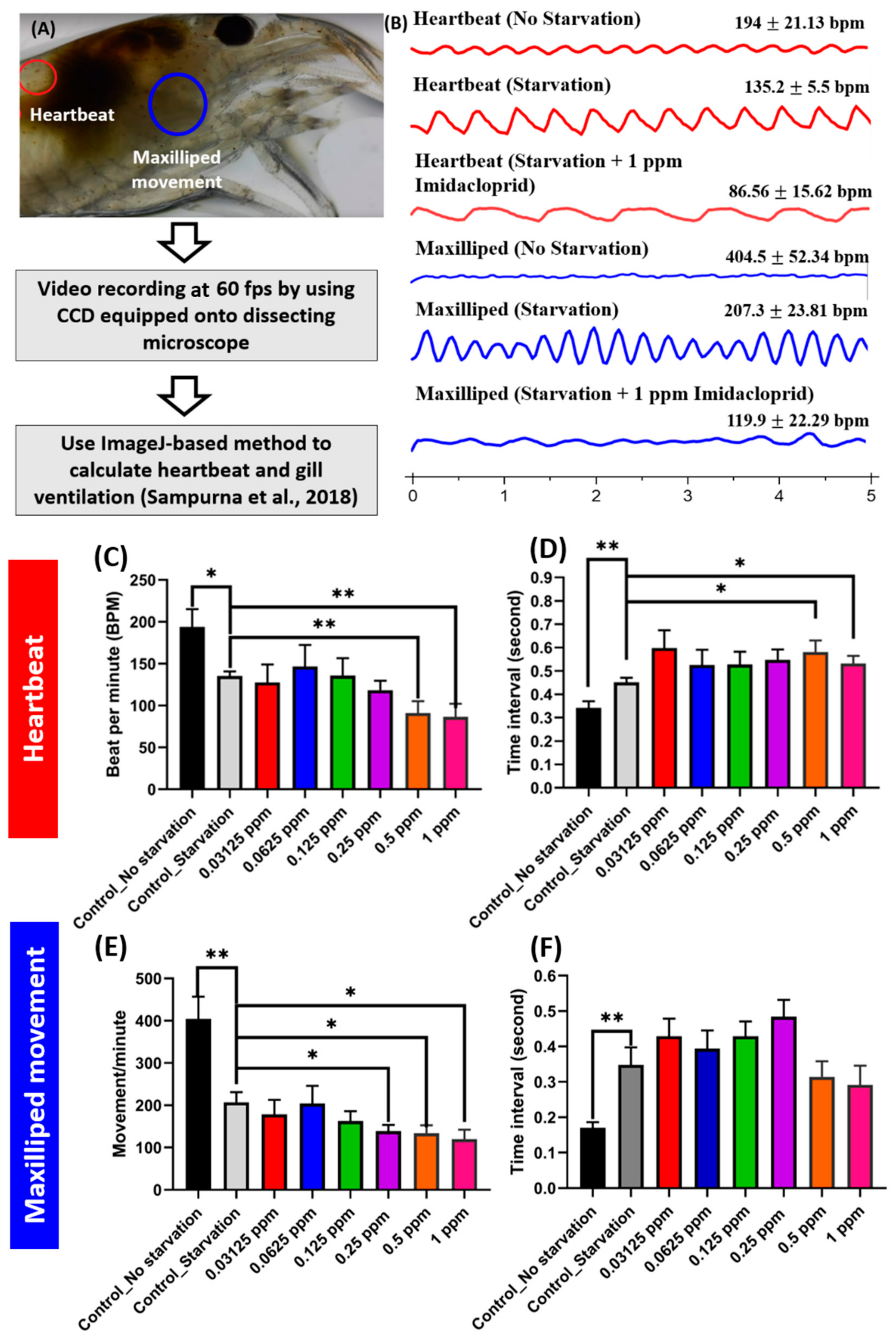
2.6. Heartbeat and Gill Ventilation (Based on Maxilliped Movement) Measurement
2.7. ELISA for Measuring Biomarker Expression
2.8. In Silico Methods
2.8.1. Protein Preparation
2.8.2. Ligand Preparation
2.8.3. Molecular Docking
2.9. Acetylcholine Rescue Experiment
2.10. Biostatistics
3. Results
3.1. Imidacloprid Exposure Impaired Locomotor Activity in Shrimp
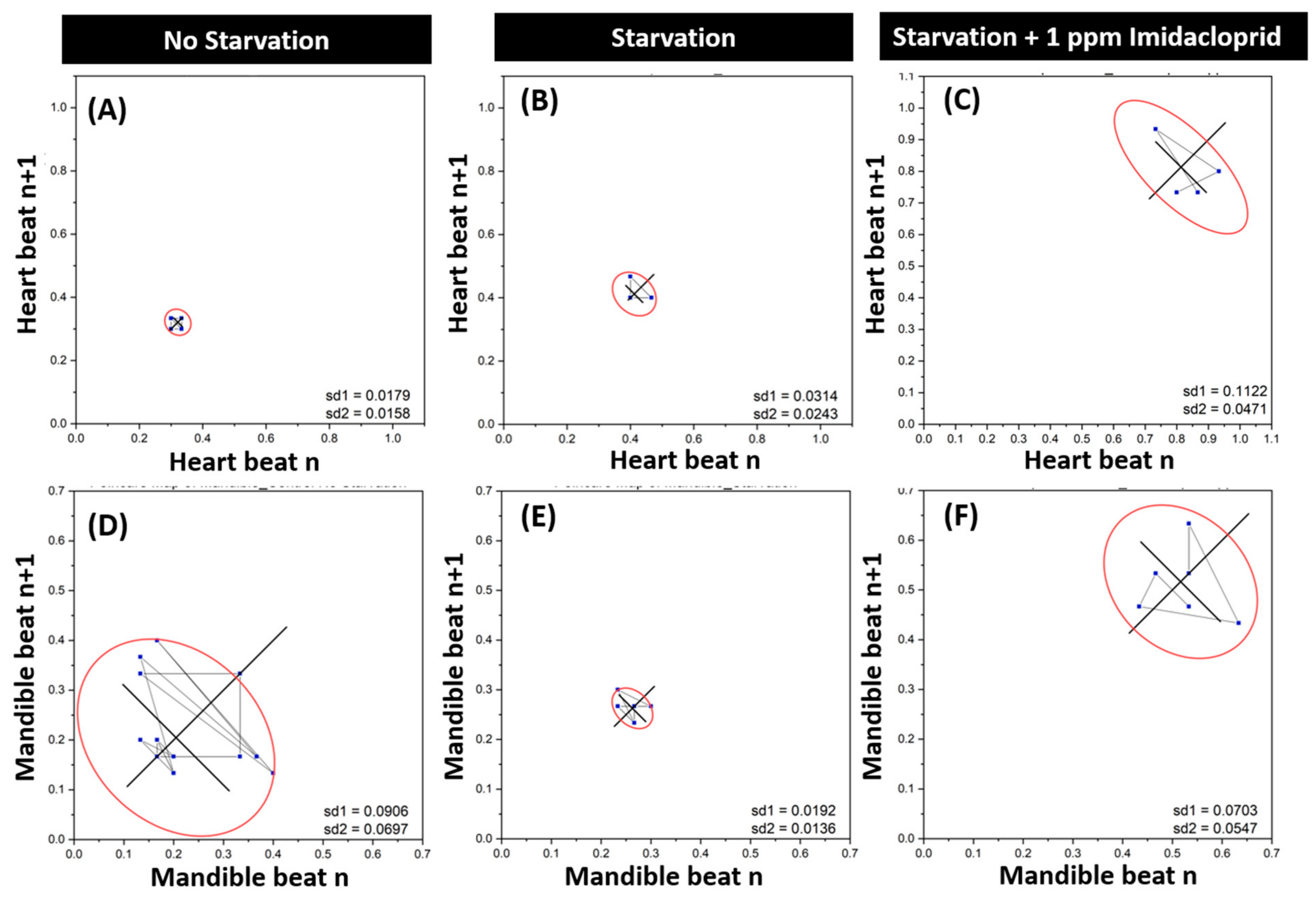
3.2. Imidacloprid Exposure Impaired Heartbeat and Gill Ventilation in Shrimp
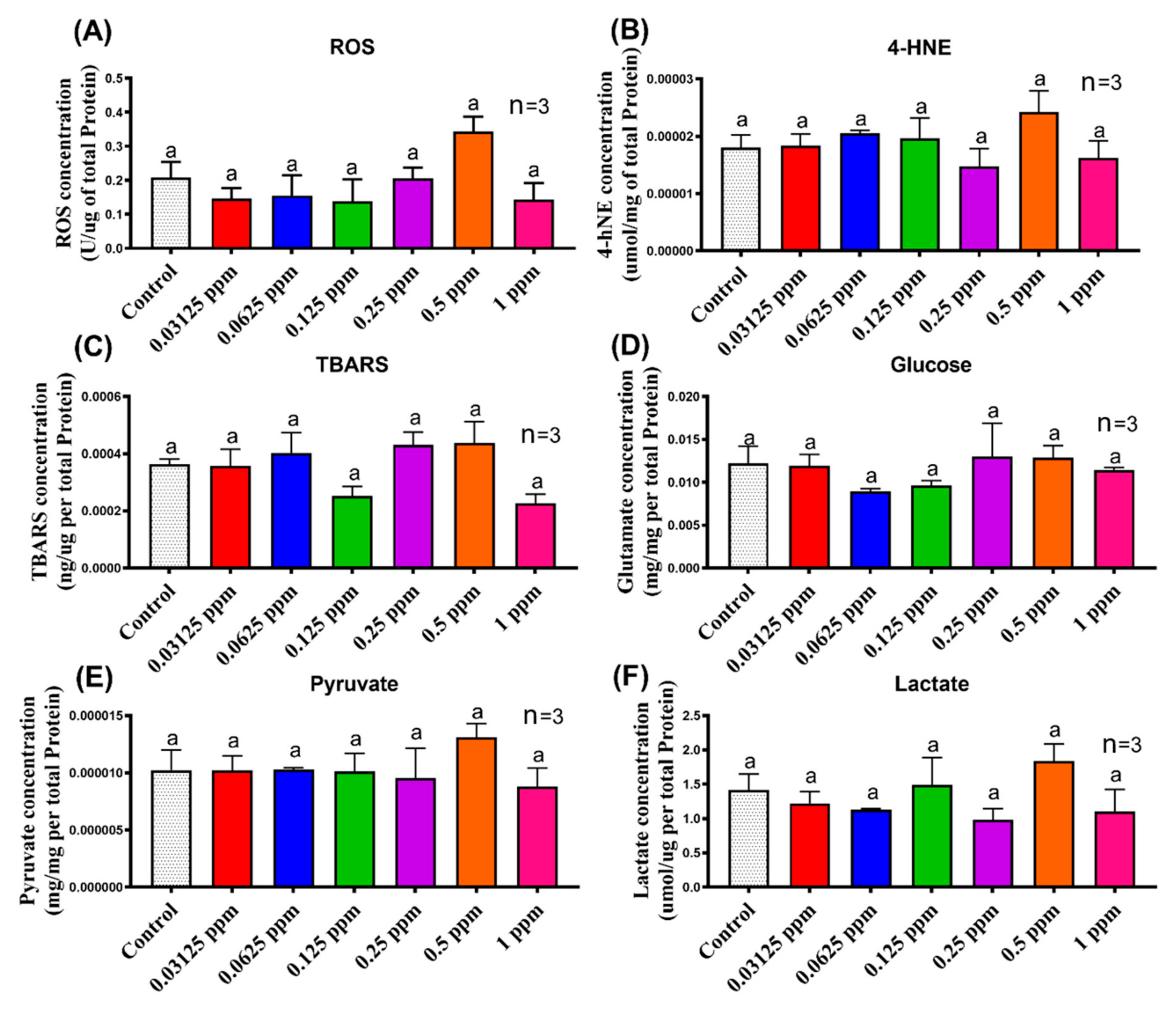
3.3. Biomarker Expression in Shrimp after Imidacloprid Administration
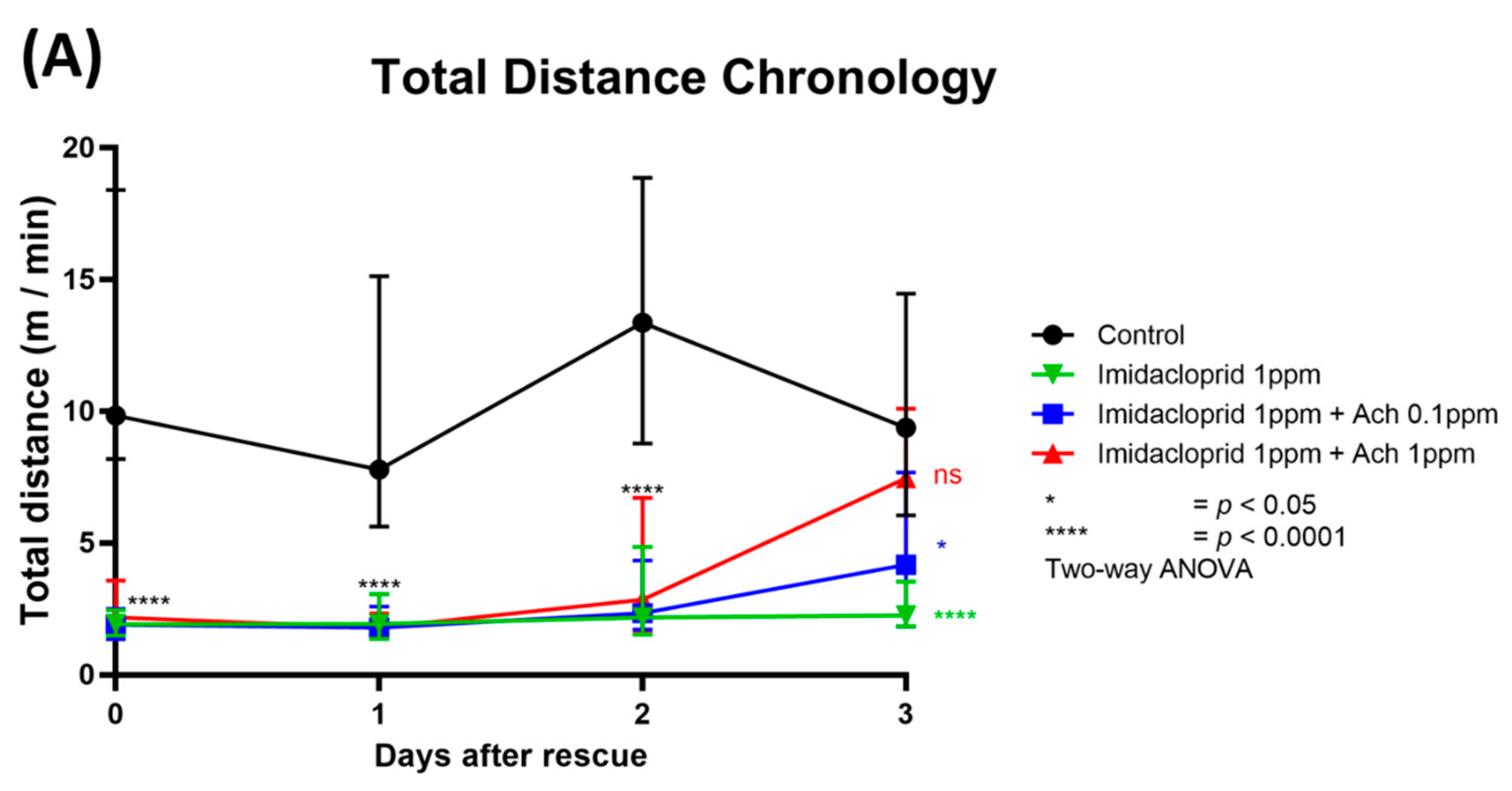
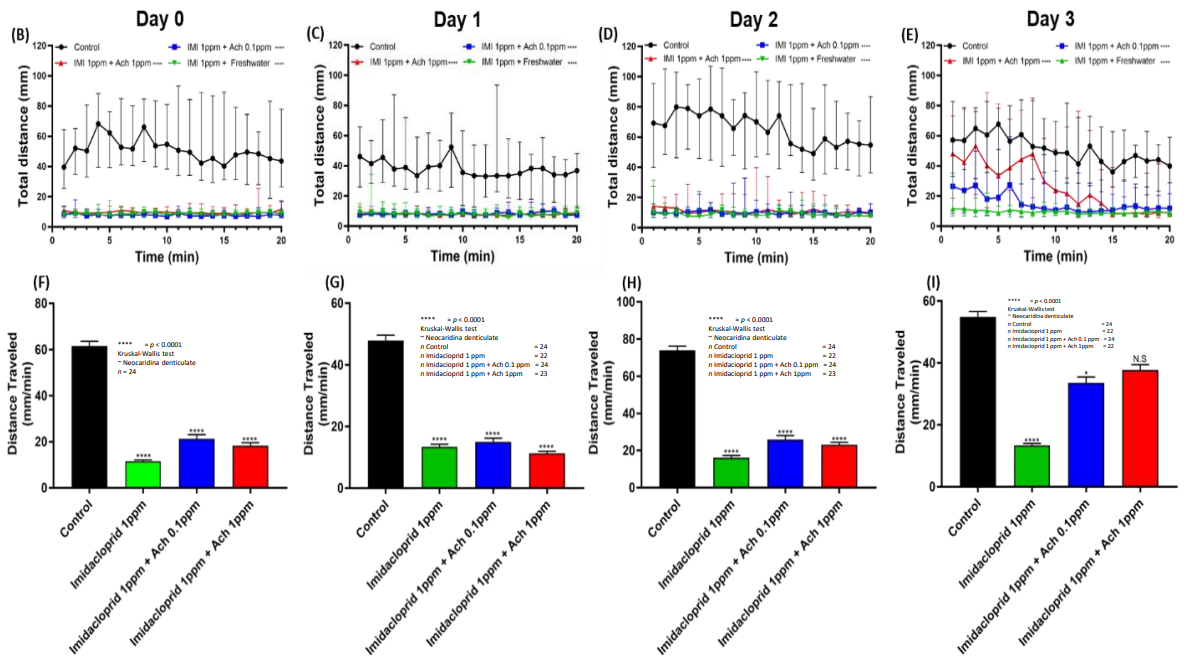
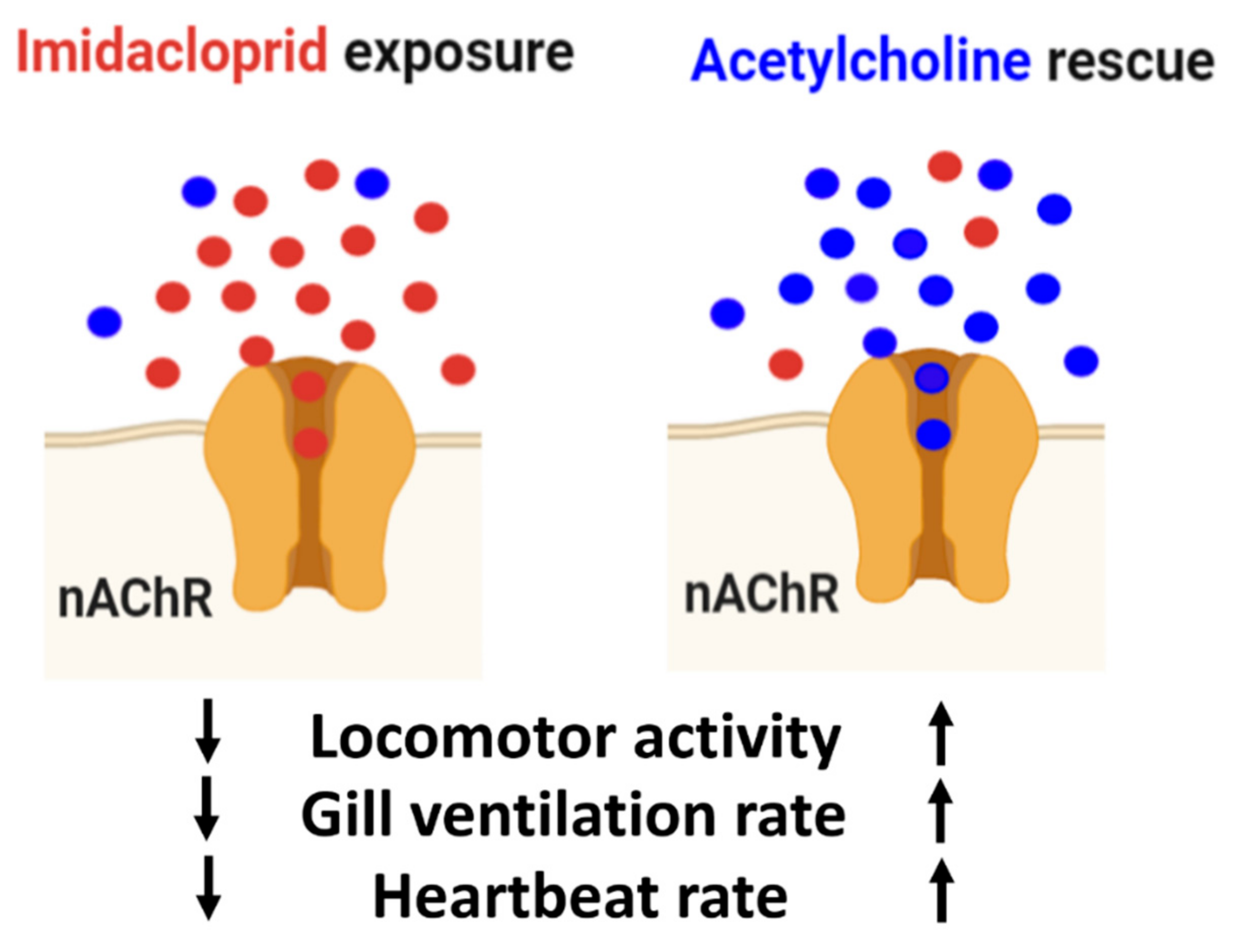
3.4. Locomotion Impairment Can Be Rescued by Acetylcholine Administration
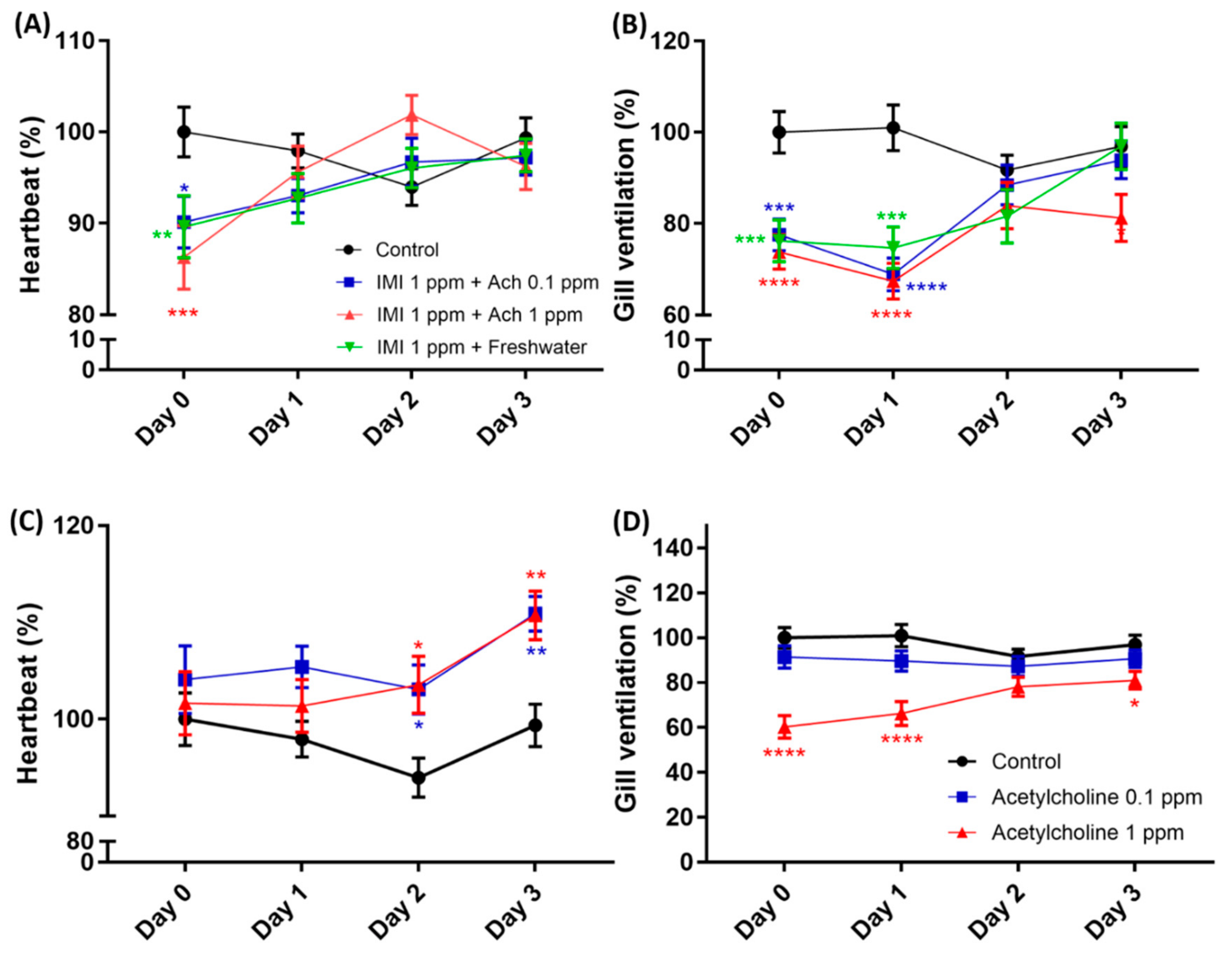
3.5. Heartbeat and Gill Ventilation Rescue by Acetylcholine Administration
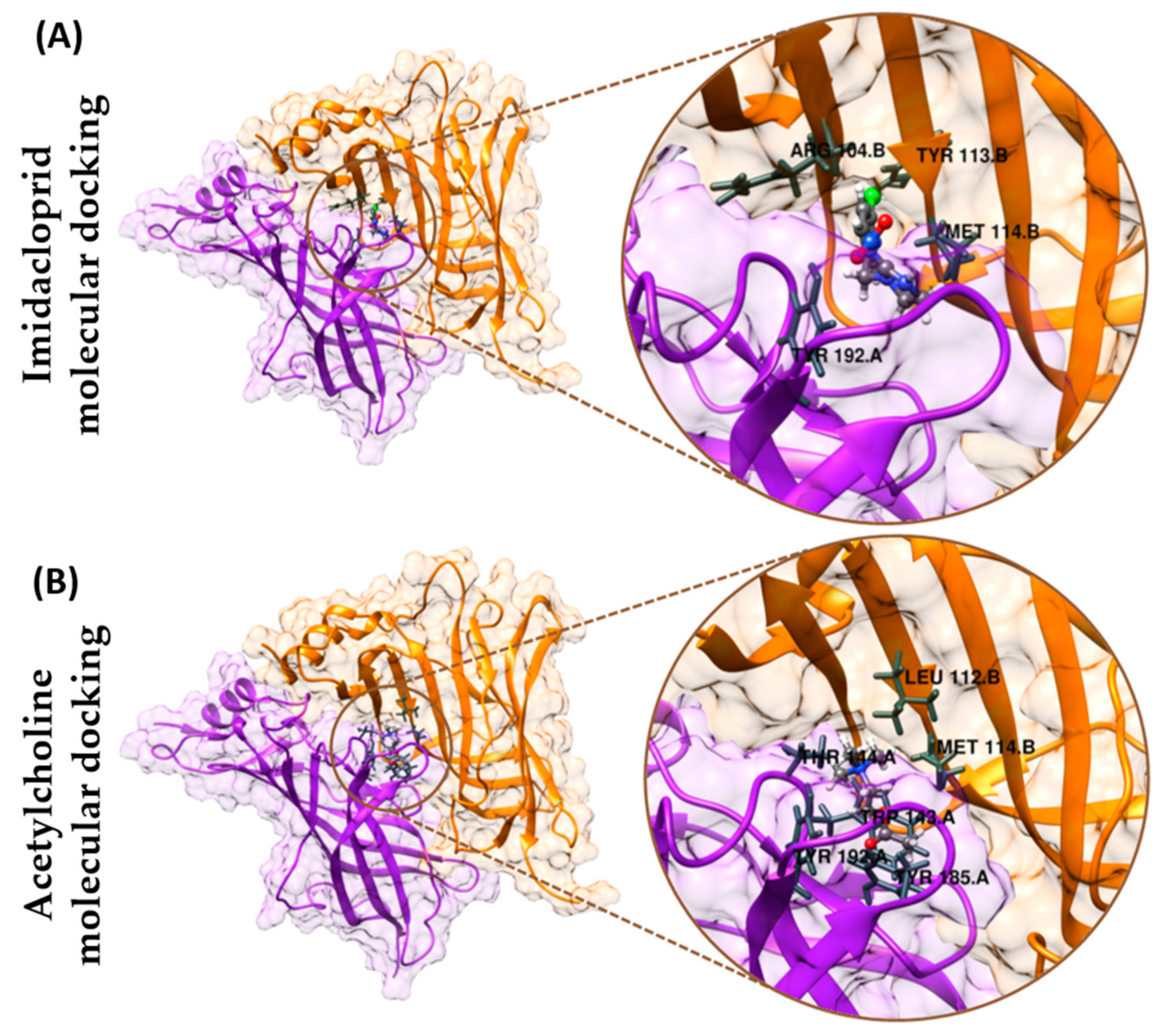
3.6. Molecular Docking
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Chaudhuri, K.; Selvaraj, S.; Pal, A. Studies on the genotoxicity of endosulfan in bacterial systems. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 1999, 439, 63–67. [Google Scholar] [CrossRef]
- Salameh, P.; Brochard, P. Respiratory symptoms in children and exposure to pesticides. Eur. Respir. J. 2003, 22, 507–512. [Google Scholar] [CrossRef] [Green Version]
- Paolini, M.; Sapone, A.; Gonzalez, F.J. Parkinson’s disease, pesticides and individual vulnerability. Trends Pharmacol. Sci. 2004, 25, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Flower, K.B.; Hoppin, J.A.; Lynch, C.F.; Blair, A.; Knott, C.; Shore, D.L.; Sandler, D.P. Cancer risk and parental pesticide application in children of Agricultural Health Study participants. Environ. Health Perspect. 2004, 112, 631–635. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S. Occupational exposure associated with reproductive dysfunction. J. Occup. Health 2004, 46, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Hook, S.E.; Doan, H.; Gonzago, D.; Musson, D.; Du, G.; Kookana, R.; Sellars, M.J.; Kummar, A. The impacts of modern-use pesticides on shrimp aquaculture: An assessment for north eastern Australia. Ecotoxicol. Environ. Saf. 2018, 148, 770–780. [Google Scholar] [CrossRef]
- Yan, S.H.; Wang, J.H.; Zhu, L.S.; Chen, A.M. Thiamethoxam induces oxidative stress and antioxidant response in zebrafish (Danio rerio) livers. Environ. Toxicol. 2016, 31, 2006–2015. [Google Scholar] [CrossRef]
- Azevedo-Pereira, H.; Lemos, M.; Soares, A.M. Effects of imidacloprid exposure on Chironomus riparius Meigen larvae: Linking acetylcholinesterase activity to behaviour. Ecotoxicol. Environ. Saf. 2011, 74, 1210–1215. [Google Scholar] [CrossRef] [PubMed]
- Jemec, A.; Tisler, T.; Drobne, D.; Sepcić, K.; Fournier, D.; Trebse, P. Comparative toxicity of imidacloprid, of its commercial liquid formulation and of diazinon to a non-target arthropod, the microcrustacean Daphnia magna. Chemosphere 2007, 68, 1408–1418. [Google Scholar] [CrossRef] [PubMed]
- Stoughton, S.J.; Liber, K.; Culp, J.; Cessna, A. Acute and chronic toxicity of imidacloprid to the aquatic invertebrates Chironomus tentans and Hyalella azteca under constant-and pulse-exposure conditions. Arch. Environ. Contam. Toxicol. 2008, 54, 662–673. [Google Scholar] [CrossRef]
- Dani, J.A.; De Biasi, M. Cellular mechanisms of nicotine addiction. Pharmacol. Biochem. Behav. 2001, 70, 439–446. [Google Scholar] [CrossRef]
- Matsuda, K.; Buckingham, S.D.; Freeman, J.C.; Squire, M.D.; Baylis, H.A.; Sattelle, D.B. Effects of the α subunit on imidacloprid sensitivity of recombinant nicotinic acetylcholine receptors. Br. J. Pharmacol. 1998, 123, 518–524. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, K.; Buckingham, S.D.; Kleier, D.; Rauh, J.J.; Grauso, M.; Sattelle, D.B. Neonicotinoids: Insecticides acting on insect nicotinic acetylcholine receptors. Trends Pharmacol. Sci. 2001, 22, 573–580. [Google Scholar] [CrossRef]
- Tomizawa, M.; Casida, J.E. Neonicotinoid insecticide toxicology: Mechanisms of selective action. Annu. Rev. Pharm. Toxicol 2005, 45, 247–268. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.-J.; Chen, H.-C. Effects of chlordane and lindane on testosterone and vitellogenin levels in green neon shrimp (Neocaridina denticulata). Int. J. Toxicol. 2004, 23, 91–95. [Google Scholar] [CrossRef]
- Sung, H.-H.; Lin, Y.-H.; Hsiao, C.-Y. Differential immune responses of the green neon shrimp (Neocaridina denticulate) to dipropyl phthalate. Fish Shellfish Immunol. 2011, 31, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, P.; Nauen, R.; Schindler, M.; Elbert, A. Overview of the status and global strategy for neonicotinoids. J. Agric. Food Chem. 2011, 59, 2897–2908. [Google Scholar] [CrossRef]
- Simon-Delso, N.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Chagnon, M.; Downs, C.; Furlan, L.; Gibbons, D.W.; Giorio, C.; Girolami, V.; et al. Systemic insecticides (neonicotinoids and fipronil): Trends, uses, mode of action and metabolites. Environ. Sci. Pollut. Res. 2015, 22, 5–34. [Google Scholar] [CrossRef]
- Sanchez-Bayo, F. Insecticides mode of action in relation to their toxicity to non-target organisms. J. Environ. Anal. Toxicol. 2012, 21, 973–992. [Google Scholar]
- Chao, S.L.; Casida, J.E. Interaction of imidacloprid metabolites and analogs with the nicotinic acetylcholine receptor of mouse brain in relation to toxicity. Pestic. Biochem. Physiol. 1997, 58, 77–88. [Google Scholar] [CrossRef]
- Shimomura, M.; Yokota, M.; Ihara, M.; Akamatsu, M.; Sattelle, D.B.; Matsuda, K. Role in the selectivity of neonicotinoids of insect-specific basic residues in loop D of the nicotinic acetylcholine receptor agonist binding site. Mol. Pharmacol. 2006, 70, 1255–1263. [Google Scholar] [CrossRef] [Green Version]
- Barrantes, F. Structural basis for lipid modulation of nicotinic acetylcholine receptor function. Brain Res. Rev. 2004, 47, 71–95. [Google Scholar] [CrossRef]
- Tiwari, P.; Dwivedi, S.; Singh, M.P.; Mishra, R.; Chandy, A. Basic and modern concepts on cholinergic receptor: A review. Asian Pac. J. Trop. Dis. 2013, 3, 413–420. [Google Scholar] [CrossRef]
- Beers, W.; Reich, E. Structure and activity of acetylcholine. Nature 1970, 228, 917–922. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landis, W.; Sofield, R.; Yu, M.-H.; Landis, W.G. Introduction to Environmental Toxicology: Impacts of Chemicals upon Ecological Systems; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Sampurna, B.P.; Audira, G.; Juniardi, S.; Lai, Y.-H.; Hsiao, C.-D. A simple imagej-based method to measure cardiac rhythm in zebrafish embryos. Inventions 2018, 3, 21. [Google Scholar] [CrossRef] [Green Version]
- Ihara, M.; Okajima, T.; Yamashita, A.; Oda, T.; Hirata, K.; Nishiwaki, H.; Morimoto, T.; Akamatsu, M.; Ashikawa, Y.; Kuroda, S.I. Crystal structures of Lymnaea stagnalis AChBP in complex with neonicotinoid insecticides imidacloprid and clothianidin. Invertebr. Neurosci 2008, 8, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, W.; Kollman, P.A.; Case, D.A. Automatic atom type and bond type perception in molecular mechanical calculations. J. Mol. Graph. Model. 2006, 25, 247–260. [Google Scholar] [CrossRef]
- Yang, J.; Roy, A.; Zhang, Y.J.B. Protein–ligand binding site recognition using complementary binding-specific substructure comparison and sequence profile alignment. Bioinformatics 2013, 29, 2588–2595. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, O.; Takeuchi, R. UMATracker: An intuitive image-based tracking platform. J. Exp. Biol. 2018, 221. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-K.; Guo, Q.; Jang, I.-K. Effect of biofloc on the survival and growth of the postlarvae of three penaeids (Litopenaeus vannamei, Fenneropenaeus chinensis, and Marsupenaeus japonicus) and their biofloc feeding efficiencies, as related to the morphological structure of the third maxilliped. J. Crustacean Biol. 2015, 35, 41–50. [Google Scholar]
- Estes, P.S. Cardiovascular and Respiratory Responses of the Ghost Shrimp, Callianassa Californiensis Dana, to the Pesticide Carbaryl and Its Hydrolytic Product 1-Naphthol. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 1986. [Google Scholar]
- Olima, C.; Pablo, F.; Lim, R. Comparative tolerance of three populations of the freshwater shrimp (Paratya australiensis) to the organophosphate pesticide, chlorpyrifos. Bull. Env.. Contam. Toxicol 1997, 59, 321–328. [Google Scholar] [CrossRef]
- Alexander, C.; Hindley, J.; Jones, S. Structure and function of the third maxillipeds of the banana prawn Penaeus merguiensis. Mar. Biol. 1980, 58, 245–249. [Google Scholar] [CrossRef]
- Hsiao, C.-D.; Wu, H.-H.; Malhotra, N.; Liu, Y.-C.; Wu, Y.-H.; Lin, Y.-N.; Saputra, F.; Santoso, F.; Chen, K.H.-C. Expression and Purification of Recombinant GHK Tripeptides Are Able to Protect against Acute Cardiotoxicity from Exposure to Waterborne-Copper in Zebrafish. Biomolecules 2020, 10, 1202. [Google Scholar] [CrossRef]
- Guzik, P.; Piskorski, J.; Krauze, T.; Wykretowicz, A. Heart rate asymmetry by Poincaré plots of RR intervals. Biomed. Tech. 2006, 51, 272–275. [Google Scholar] [CrossRef]
- Piskorski, J.; Guzik, P. Geometry of the Poincaré plot of RR intervals and its asymmetry in healthy adults. Physiol. Meas. 2007, 28, 287. [Google Scholar] [CrossRef] [Green Version]
- Slaninova, A.; Smutna, M.; Modra, H.; Svobodova, Z. REVIEWS Oxidative stress in fish induced by pesticides. Neuroendocr. Lett 2009, 30, 2. [Google Scholar]
- Martelli, F.; Zhongyuan, Z.; Wang, J.; Wong, C.-O.; Karagas, N.E.; Roessner, U.; Rupasinghe, T.; Venkatachalam, K.; Perry, T.; Bellen, H.J. Low doses of the neonicotinoid insecticide imidacloprid induce ROS triggering neurological and metabolic impairments in Drosophila. Proc. Natl. Acad. Sci. USA 2020, 117, 25840–25850. [Google Scholar] [CrossRef]
- Tsai, M.-C.; Huang, T.-L. Thiobarbituric acid reactive substances (TBARS) is a state biomarker of oxidative stress in bipolar patients in a manic phase. J. Affect. Disord. 2015, 173, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Kwack, S.J.; Yoon, K.S.; Choi, J.S.; Lee, B.-M. 4-Hydroxynonenal: A superior oxidative biomarker compared to malondialdehyde and carbonyl content induced by carbon tetrachloride in rats. J. Toxicol. Environ. Healthpart A 2015, 78, 1051–1062. [Google Scholar] [CrossRef]
- Liu, M.-Y.; Casida, J.E. High affinity binding of [3H] imidacloprid in the insect acetylcholine receptor. Pestic. Biochem. Physiol. 1993, 46, 40–46. [Google Scholar] [CrossRef]
- McGehee, D.S.; Role, L.W. Physiological diversity of nicotinic acetylcholine receptors expressed by vertebrate neurons. Annu. Rev. Physiol. 1995, 57, 521–546. [Google Scholar] [CrossRef]
- Levitt-Barmats, Y.A.; Yanai, Z.; Cohen, T.M.; Shenkar, N. Life-history traits and ecological characteristics of the ornamental shrimp Neocaridina denticulata (De Haan, 1844), recently introduced into the freshwater systems of Israel. Aquat. Invasions 2019, 14, 684–702. [Google Scholar] [CrossRef]
- Kenny, N.J.; Sin, Y.W.; Shen, X.; Zhe, Q.; Wang, W.; Chan, T.F.; Tobe, S.S.; Shimeld, S.M.; Chu, K.H.; Hui, J.H. Genomic sequence and experimental tractability of a new decapod shrimp model, Neocaridina denticulata. Mar. Drugs 2014, 12, 1419–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.-W.; Chu, P.-Y.; Wu, Y.-F.; Chan, W.-R.; Wang, Y.-H. Identification of Functional SSR Markers in Freshwater Ornamental Shrimps Neocaridina denticulata Using Transcriptome Sequencing. Mar. Biotechnol. 2020, 22, 772–785. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Jeon, E.; Jung, S.; Ahn, S.; Park, M.; Seo, J. Gene expression profiling and expression analysis of freshwater shrimp (Neocaridina denticulata denticulata) using expressed sequence tags and short-term exposure to copper. J. Environ. Biol. 2018, 39, 51–57. [Google Scholar] [CrossRef]
- Xing, K.; Liu, Y.; Yan, C.; Zhou, Y.; Sun, Y.; Su, N.; Yang, F.; Xie, S.; Zhang, J. Transcriptome analysis of Neocaridina denticulate sinensis under copper exposure. Gene 2020, 764, 145098. [Google Scholar] [CrossRef]
- Chou, C.-H.; Huang, H.-Y.; Huang, W.-C.; Hsu, S.-D.; Hsiao, C.-D.; Liu, C.-Y.; Chen, Y.-H.; Liu, Y.-C.; Huang, W.-Y.; Lee, M.-L. The aquatic animals’ transcriptome resource for comparative functional analysis. BMC Genom. 2018, 19, 161–170. [Google Scholar] [CrossRef]
- Mykles, D.L.; Hui, J.H. Neocaridina denticulata: A decapod crustacean model for functional genomics. Integr. Comp. Biol. 2015, 55, 891–897. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zang, L.; Liu, M.; Zhang, C.; Shen, G.; Du, W.; Sun, Z.; Fei, J.; Yang, L.; Wang, Y. Ecological risk assessment of the increasing use of the neonicotinoid insecticides along the east coast of China. Environ. Int 2019, 127, 550–557. [Google Scholar] [CrossRef]
- Pittendrigh, B.R.; Margam, V.M.; Sun, L.; Huesing, J.E. Resistance in the postgenomics age. Insect Resist. Manag. Biol. Econ. Predict. 2008, 2, 39–68. [Google Scholar]
- Dalefield, R. Veterinary Toxicology for Australia and New Zealand; Elsevier: Amsterdam, The Netherlands, 2017; Volume 1, Available online: https://0-www-elsevier-com.brum.beds.ac.uk/books/veterinary-toxicology-for-australia-and-new-zealand/dalefield/978-0-12-420227-6 (accessed on 21 January 2021).
- Gupta, R.C. Biomarkers in Toxicology; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Miles, J.C.; Hua, J.; Sepulveda, M.S.; Krupke, C.H.; Hoverman, J.T. Effects of clothianidin on aquatic communities: Evaluating the impacts of lethal and sublethal exposure to neonicotinoids. PLoS ONE 2017, 12, e0174171. [Google Scholar] [CrossRef]
- Vignet, C.; Cappello, T.; Fu, Q.; Lajoie, K.; De Marco, G.; Clérandeau, C.; Mottaz, H.; Maisano, M.; Hollender, J.; Schirmer, K. Imidacloprid induces adverse effects on fish early life stages that are more severe in Japanese medaka (Oryzias latipes) than in zebrafish (Danio rerio). Chemosphere 2019, 225, 470–478. [Google Scholar] [CrossRef]
- Vieira, C.E.D.; Pérez, M.R.; Acayaba, R.D.A.; Raimundo, C.C.M.; dos Reis Martinez, C.B. DNA damage and oxidative stress induced by imidacloprid exposure in different tissues of the Neotropical fish Prochilodus lineatus. Chemosphere 2018, 195, 125–134. [Google Scholar] [CrossRef]
- Özdemir, S.; Altun, S.; Arslan, H. Imidacloprid exposure cause the histopathological changes, activation of TNF-α, iNOS, 8-OHdG biomarkers, and alteration of caspase 3, iNOS, CYP1A, MT1 gene expression levels in common carp (Cyprinus carpio L.). Toxicol. Rep. 2018, 5, 125–133. [Google Scholar]
- Hussain, A.; Audira, G.; Malhotra, N.; Uapipatanakul, B.; Chen, J.-R.; Lai, Y.-H.; Huang, J.-C.; Chen, K.H.-C.; Lai, H.-T.; Hsiao, C.-D. Multiple Screening of Pesticides Toxicity in Zebrafish and Daphnia Based on Locomotor Activity Alterations. Biomolecules 2020, 10, 1224. [Google Scholar] [CrossRef]
- Al-Badran, A.A.; Fujiwara, M.; Mora, M.A. Effects of insecticides, fipronil and imidacloprid, on the growth, survival, and behavior of brown shrimp Farfantepenaeus aztecus. PLoS ONE 2019, 14, e0223641. [Google Scholar] [CrossRef] [Green Version]
- Overmyer, J.; Mason, B.; Armbrust, K. Acute toxicity of imidacloprid and fipronil to a nontarget aquatic insect, Simulium vittatum Zetterstedt cytospecies IS-7. Bull. Environ. Contam. Toxicol. 2005, 74, 872–879. [Google Scholar] [CrossRef]
- Crosby, E.B.; Bailey, J.M.; Oliveri, A.N.; Levin, E.D. Neurobehavioral impairments caused by developmental imidacloprid exposure in zebrafish. Neurotoxicol. Teratol. 2015, 49, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Widdows, J. Effect of temperature and food on the heart beat, ventilation rate and oxygen uptake of Mytilus edulis. Mar. Biol. 1973, 20, 269–276. [Google Scholar] [CrossRef]
- Santoso, F.; Krylov, V.V.; Castillo, A.L.; Saputra, F.; Chen, H.-M.; Lai, H.-T.; Hsiao, C.-D. Cardiovascular Performance Measurement in Water Fleas by Utilizing High-Speed Videography and ImageJ Software and Its Application for Pesticide Toxicity Assessment. Animals 2020, 10, 1587. [Google Scholar] [CrossRef]
- Bownik, A.; Pawłocik, M.; Sokołowska, N. Effects of neonicotinoid insecticide acetamiprid on swimming velocity, heart rate and thoracic limb movement of Daphnia magna. Pol. J. Nat. Sci. 2017, 32, 481–493. [Google Scholar]
- Tišler, T.; Jemec, A.; Mozetič, B.; Trebše, P. Hazard identification of imidacloprid to aquatic environment. Chemosphere 2009, 76, 907–914. [Google Scholar] [CrossRef]
- Guo, L.; Dai, Z.; Guo, J.; Yang, W.; Ge, F.; Dai, Y. Oligotrophic bacterium Hymenobacter latericoloratus CGMCC 16346 degrades the neonicotinoid imidacloprid in surface water. Amb Express 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.-Y.; Luo, Q.-H.; Hou, C.-S.; Wang, Q.; Dai, P.-L.; Gao, J.; Liu, Y.-J.; Diao, Q.-Y. Sublethal effects of imidacloprid on targeting muscle and ribosomal protein related genes in the honey bee Apis mellifera L. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef]
- Nyman, A.-M.; Hintermeister, A.; Schirmer, K.; Ashauer, R. The insecticide imidacloprid causes mortality of the freshwater amphipod Gammarus pulex by interfering with feeding behavior. PLoS ONE 2013, 8, e62472. [Google Scholar] [CrossRef] [Green Version]
- Lind, R.J.; Clough, M.S.; Reynolds, S.E.; Earley, F.G. [3H] Imidacloprid Labels High-and Low-Affinity Nicotinic Acetylcholine Receptor-like Binding Sites in the AphidMyzus persicae (Hemiptera: Aphididae). Pestic. Biochem. Physiol. 1998, 62, 3–14. [Google Scholar] [CrossRef]
- Liu, W.; Zheng, W.; Ma, Y.; Liu, K.K. Sorption and degradation of imidacloprid in soil and water. J. Environ. Sci. Health Part B 2006, 41, 623–634. [Google Scholar] [CrossRef]
- Butcherine, P.; Kelaher, B.P.; Taylor, M.D.; Barkla, B.J.; Benkendorff, K. Impact of imidacloprid on the nutritional quality of adult black tiger shrimp (Penaeus monodon). Ecotoxicol. Environ. Saf. 2020, 198, 110682. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siregar, P.; Suryanto, M.E.; Chen, K.H.-C.; Huang, J.-C.; Chen, H.-M.; Kurnia, K.A.; Santoso, F.; Hussain, A.; Ngoc Hieu, B.T.; Saputra, F.; et al. Exploiting the Freshwater Shrimp Neocaridina denticulata as Aquatic Invertebrate Model to Evaluate Nontargeted Pesticide Induced Toxicity by Investigating Physiologic and Biochemical Parameters. Antioxidants 2021, 10, 391. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10030391
Siregar P, Suryanto ME, Chen KH-C, Huang J-C, Chen H-M, Kurnia KA, Santoso F, Hussain A, Ngoc Hieu BT, Saputra F, et al. Exploiting the Freshwater Shrimp Neocaridina denticulata as Aquatic Invertebrate Model to Evaluate Nontargeted Pesticide Induced Toxicity by Investigating Physiologic and Biochemical Parameters. Antioxidants. 2021; 10(3):391. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10030391
Chicago/Turabian StyleSiregar, Petrus, Michael Edbert Suryanto, Kelvin H.-C. Chen, Jong-Chin Huang, Hong-Ming Chen, Kevin Adi Kurnia, Fiorency Santoso, Akhlaq Hussain, Bui Thi Ngoc Hieu, Ferry Saputra, and et al. 2021. "Exploiting the Freshwater Shrimp Neocaridina denticulata as Aquatic Invertebrate Model to Evaluate Nontargeted Pesticide Induced Toxicity by Investigating Physiologic and Biochemical Parameters" Antioxidants 10, no. 3: 391. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10030391