
Transcriptional and Catalytic Responsiveness of the Antarctic Fish Trematomus bernacchii Antioxidant System toward Multiple Stressors


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Chemical Analyses
2.3. Transcriptional Analyses
2.4. Biological Responses
2.5. Statistical Analyses
3. Results
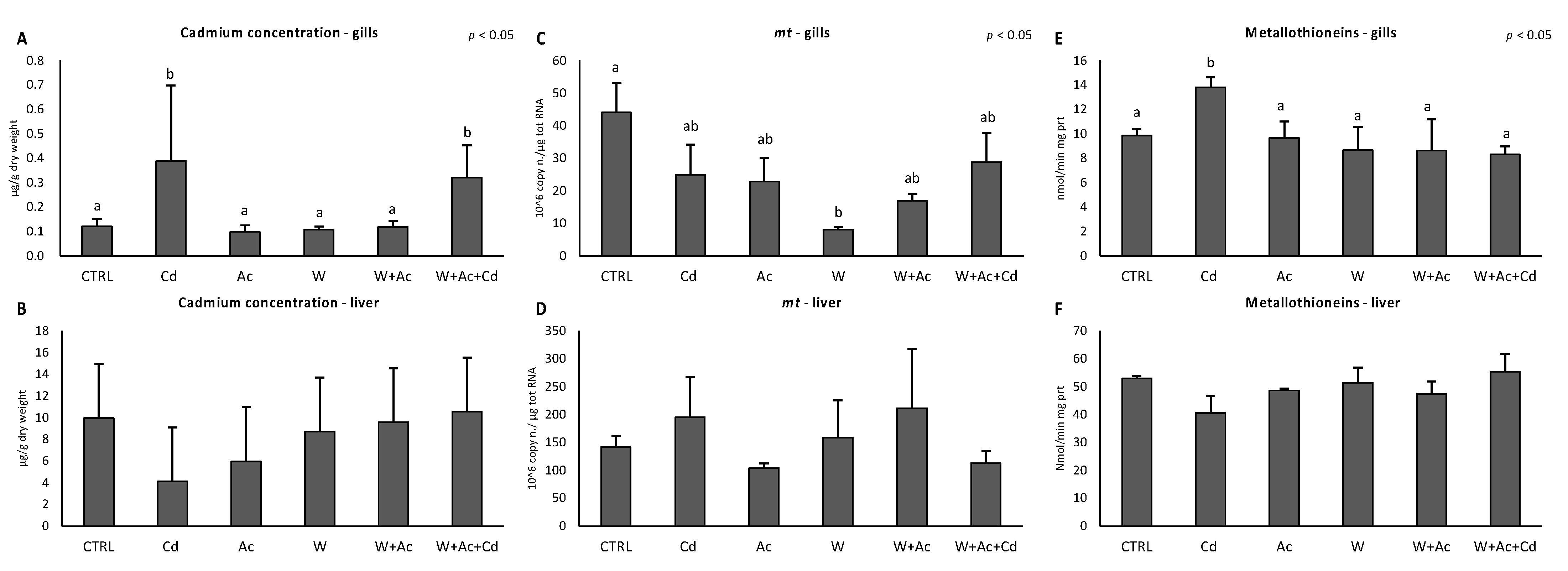
3.1. Cadmium Concentration, Metallothioneins mRNA and Protein Levels
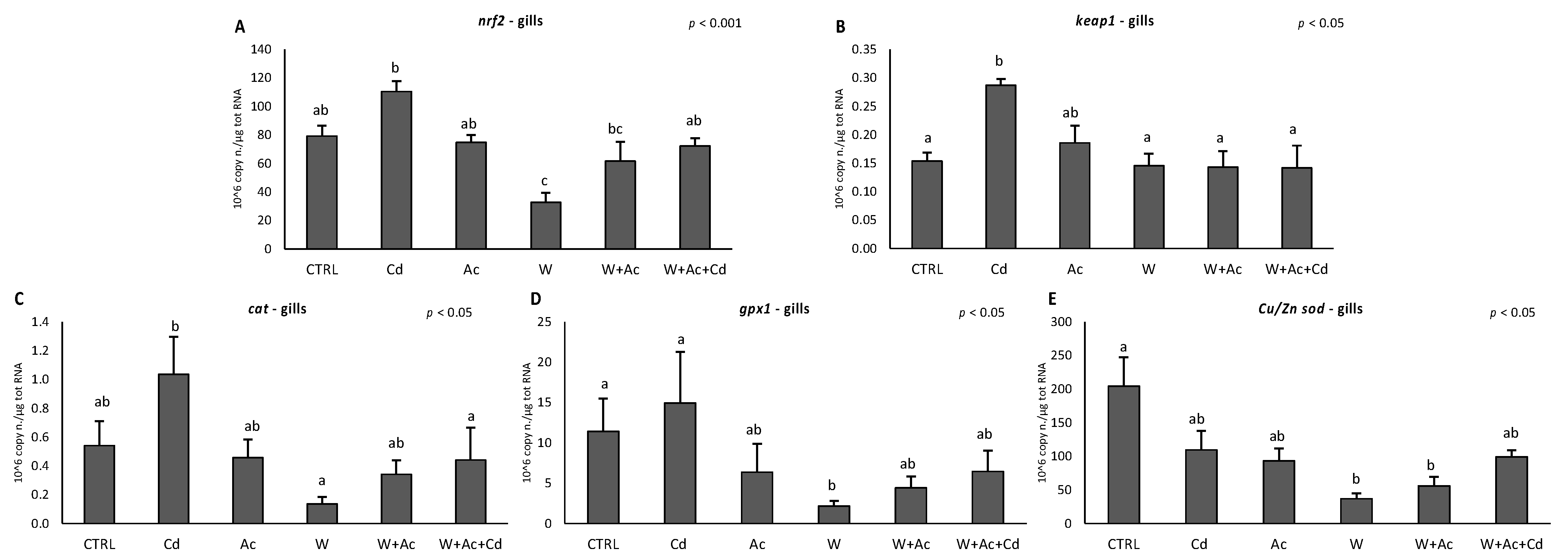
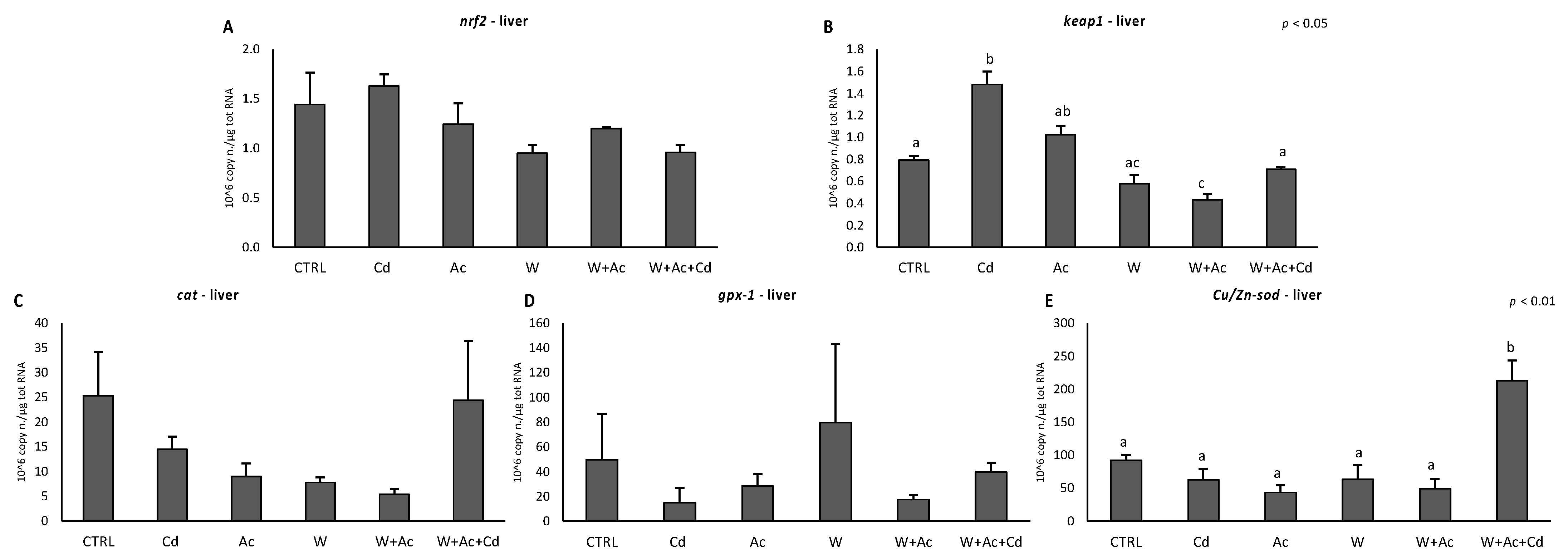
3.2. Antioxidants mRNA Levels
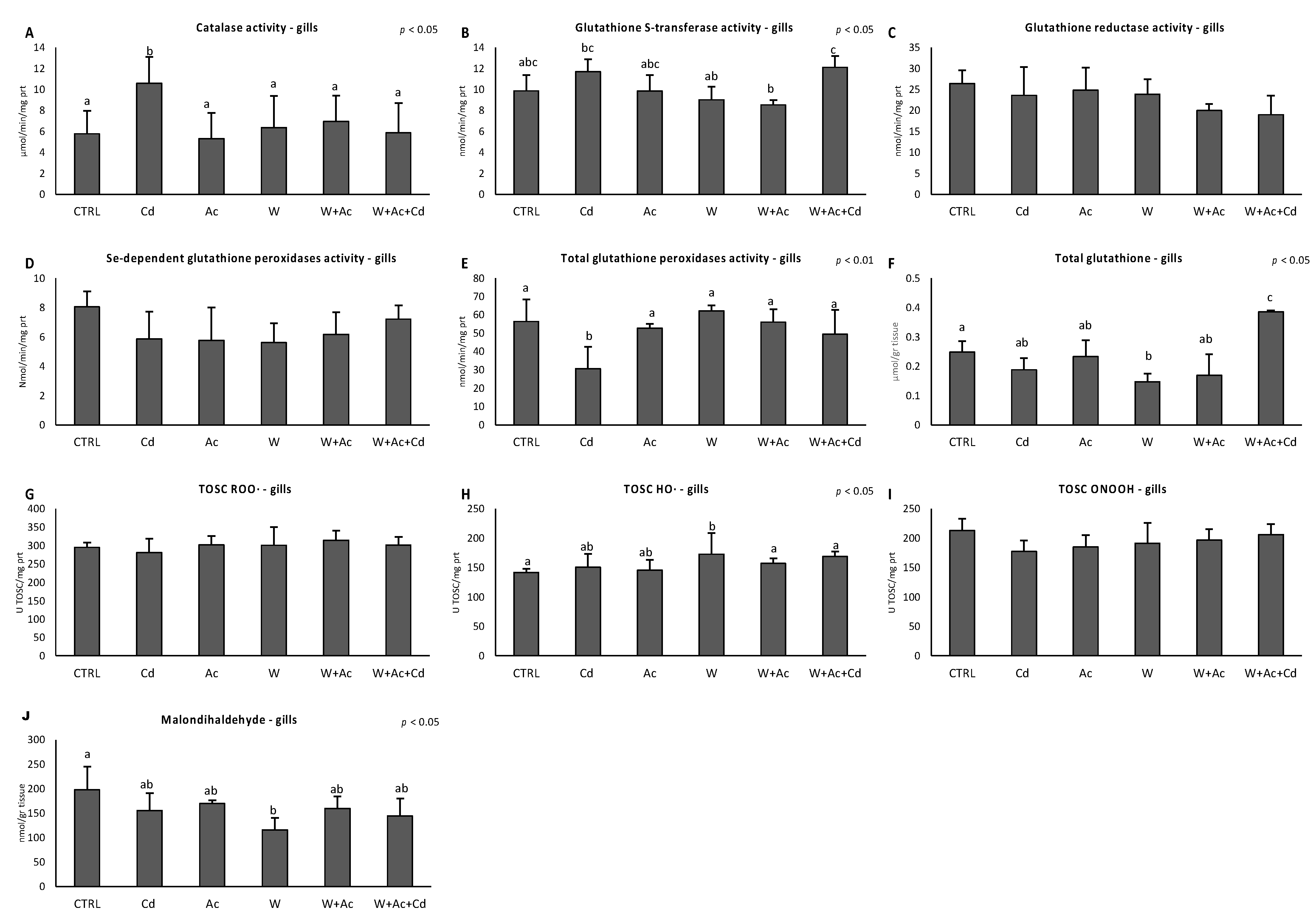
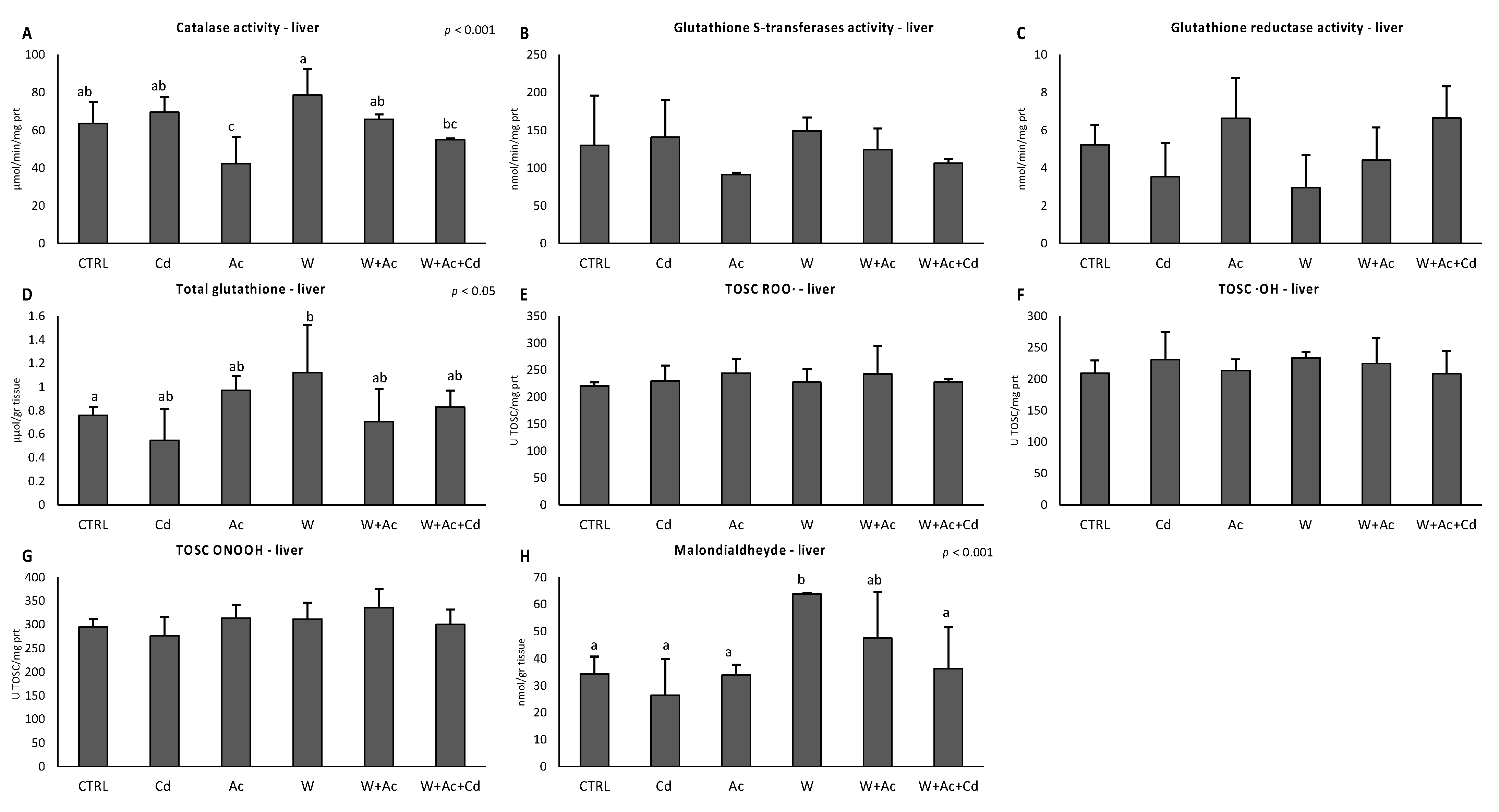
3.3. Antioxidant System Functional Analyses
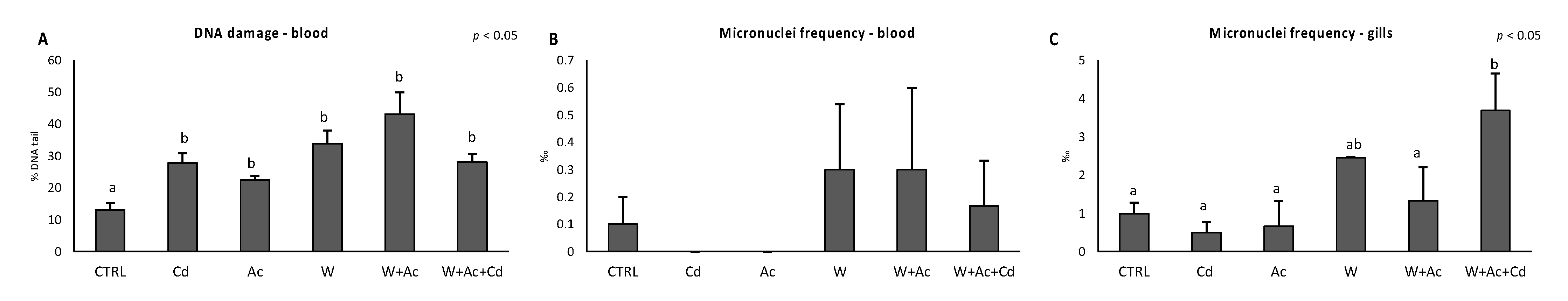
3.4. Genotoxic Damage
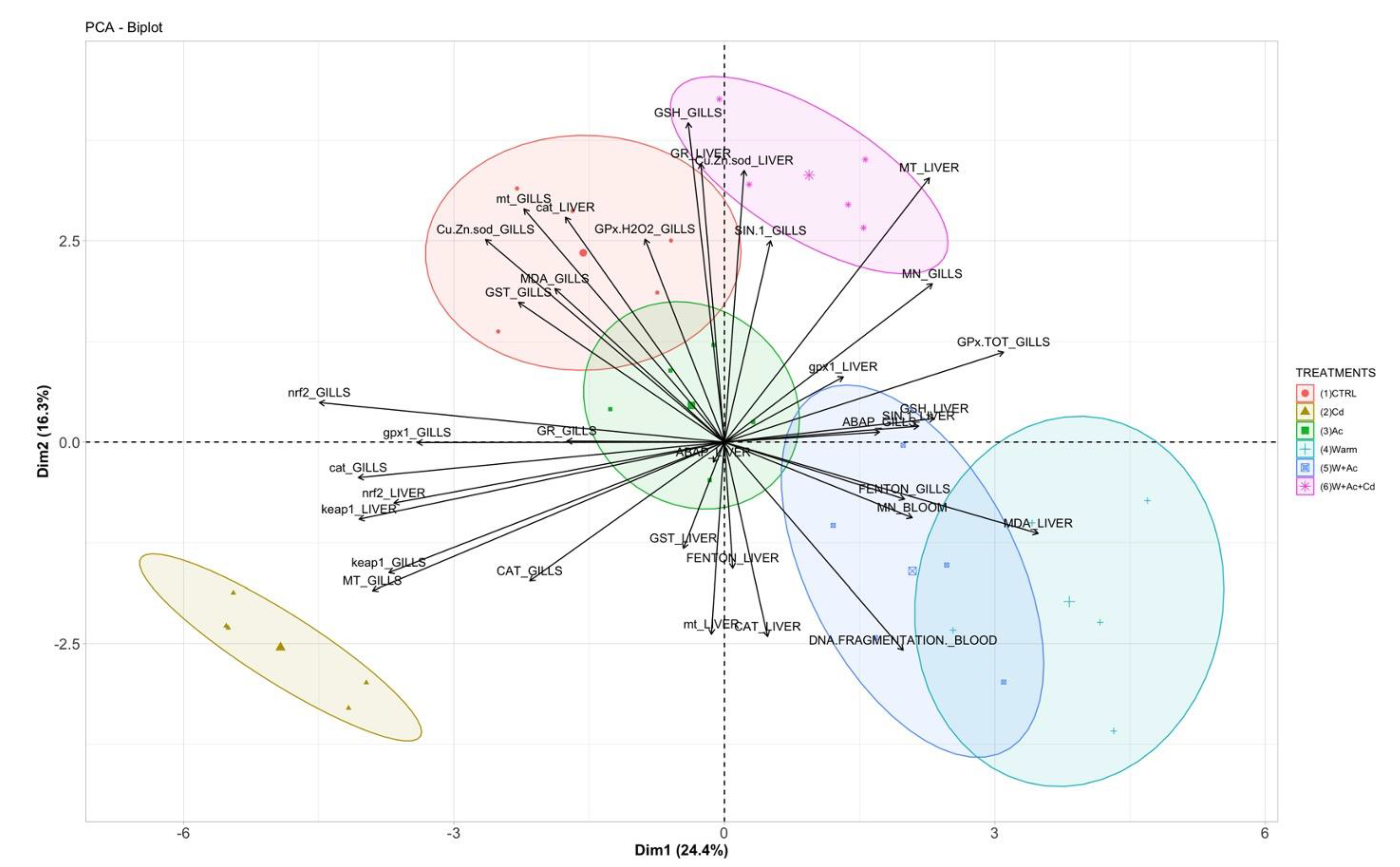
3.5. Principal Components Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robinson, S.A.; Klekociuk, A.R.; King, D.H.; Pizarro Rojas, M.; Zúñiga, G.E.; Bergstrom, D.M. The 2019/2020 summer of Antarctic heatwaves. Glob. Change Biol. 2020, 26, 3178–3180. [Google Scholar] [CrossRef]
- Meredith, M.; Sommerkorn, M.; Cassotta, S.; Derksen, C.; Ekaykin, A.; Hollowed, A.; Kofinas, G.; Mackintosh, A.; Melbourne-Thomas, J. Polar Regions. In IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; Pörtner, H.-O., Roberts, D.C., Masson-Delmotte, V., Zhai, P., Tignor, M., Poloczanska, E., Mintenbeck, K., Alegría, A., Nicolai, M., Okem, A., et al., Eds.; IPCC: Geneva, Switzerland, 2019. [Google Scholar]
- Bednaršek, N.; Tarling, G.A.; Bakker, D.C.E.; Fielding, S.; Jones, E.M.; Venables, H.J.; Ward, P.; Kuzirian, A.; Lézé, B.; Feely, R.A.; et al. Extensive dissolution of live pteropods in the Southern Ocean. Nat. Geosci. 2012, 5, 881–885. [Google Scholar] [CrossRef] [Green Version]
- Benedetti, M.; Lanzoni, I.; Nardi, A.; d’Errico, G.; Di Carlo, M.; Fattorini, D.; Nigro, M.; Regoli, F. Oxidative responsiveness to multiple stressors in the key Antarctic species, Adamussium colbecki: Interactions between temperature, acidification and cadmium exposure. Mar. Environ. Res. 2016, 121, 20–30. [Google Scholar] [CrossRef]
- Byrne, M.; Ho, M.A.; Koleits, L.; Price, C.; King, C.K.; Virtue, P.; Tilbrook, B.; Lamare, M. Vulnerability of the calcifying larval stage of the Antarctic sea urchin Sterechinus neumayeri to near-future ocean acidification and warming. Glob. Change Biol. 2013, 19, 2264–2275. [Google Scholar] [CrossRef]
- Dell’Acqua, O.; Ferrando, S.; Chiantore, M.; Asnaghi, V. The impact of ocean acidification on the gonads of three key Antarctic benthic macroinvertebrates. Aquatic. Toxicol. 2019, 210, 19–29. [Google Scholar] [CrossRef]
- Yu, P.C.; Sewell, M.A.; Matson, P.G.; Rivest, E.B.; Kapsenberg, L.; Hofmann, G.E. Growth Attenuation with Developmental Schedule Progression in Embryos and Early Larvae of Sterechinus neumayeri Raised under Elevated CO2. PLoS ONE 2013, 8, e52448. [Google Scholar] [CrossRef] [PubMed]
- Strobel, A.; Bennecke, S.; Leo, E.; Mintenbeck, K.; Pörtner, H.O.; Mark, F.C. Metabolic shifts in the Antarctic fish Notothenia rossii in response to rising temperature and PCO2. Front. Zool. 2012, 9, 28. [Google Scholar] [CrossRef] [Green Version]
- Strobel, A.; Graeve, M.; Poertner, H.O.; Mark, F.C. Mitochondrial acclimation capacities to ocean warming and acidification are limited in the Antarctic nototheniid fish, Notothenia rossii and Lepidonotothen squamifrons. PLoS ONE 2013, 8, e68865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enzor, L.A.; Place, S.P. Is warmer better? Decreased oxidative damage in notothenioid fish after long-term acclimation to multiple stressors. J. Exp. Biol. 2014, 217, 3301–3310. [Google Scholar] [CrossRef] [Green Version]
- Enzor, L.A.; Hunter, E.M.; Place, S.P. The effects of elevated temperature and ocean acidification on the metabolic pathways of notothenioid fish. Conserv. Physiol. 2017, 5, cox019. [Google Scholar] [CrossRef] [Green Version]
- Flynn, E.E.; Bjelde, B.E.; Miller, N.A.; Todgham, A.E. Ocean acidification exerts negative effects during warming conditions in a developing Antarctic fish. Conserv. Physiol. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Huth, T.J.; Place, S.P. Transcriptome wide analyses reveal a sustained cellular stress response in the gill tissue of Trematomus bernacchii after acclimation to multiple stressors. BMC Genom. 2016, 17, 127. [Google Scholar] [CrossRef] [Green Version]
- Davis, B.E.; Flynn, E.E.; Miller, N.A.; Nelson, F.A.; Fangue, N.A.; Todgham, A.E. Antarctic emerald rockcod have the capacity to compensate for warming when uncoupled from CO2-acidification. Glob. Change Biol. 2018, 24, e655–e670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braga, A.C.; Pereira, V.; Marçal, R.; Marques, A.; Guilherme, S.; Costa, P.R.; Pacheco, M. DNA damage and oxidative stress responses of mussels Mytilus galloprovincialis to paralytic shellfish toxins under warming and acidification conditions—Elucidation on the organ-specificity. Aquat. Toxicol. 2020, 228, 105619. [Google Scholar] [CrossRef] [PubMed]
- Coppola, F.; Almeida, Â.; Henriques, B.; Soares, A.M.V.M.; Figueira, E.; Pereira, E.; Freitas, R. Biochemical responses and accumulation patterns of Mytilus galloprovincialis exposed to thermal stress and Arsenic contamination. Ecotox. Environ. Safe. 2018, 147, 954–962. [Google Scholar] [CrossRef]
- Cao, R.; Zhang, T.; Li, X.; Zhao, Y.; Wang, Q.; Yang, D.; Qu, Y.; Liu, H.; Dong, Z.; Zhao, J. Seawater acidification increases copper toxicity: A multi-biomarker approach with a key marine invertebrate, the Pacific Oyster Crassostrea gigas. Aquat. Toxicol. 2019, 210, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, M.E.; Filippini, G.; Nardi, A. Season specific influence of projected ocean changes on the response to cadmium of stress-related genes in Mytilus galloprovincialis. Mar. Environ. Res. 2020, 162, 105091. [Google Scholar] [CrossRef]
- Mezzelani, M.; Nardi, A.; Bernardini, I.; Milan, M.; Peruzza, L.; d’Errico, G.; Fattorini, D.; Gorbi, S.; Patarnello, T.; Regoli, F. Environmental pharmaceuticals and climate change: The case study of carbamazepine in M. galloprovincialis under ocean acidification scenario. Environ. Int. 2021, 146, 106269. [Google Scholar] [CrossRef] [PubMed]
- Munari, M.; Matozzo, V.; Chemello, G.; Riedl, V.; Pastore, P.; Badocco, D.; Marin, M.G. Seawater acidification and emerging contaminants: A dangerous marriage for haemocytes of marine bivalves. Environ. Res. 2019, 175, 11–21. [Google Scholar] [CrossRef]
- Nardi, A.; Mincarelli, L.F.; Benedetti, M.; Fattorini, D.; d’Errico, G.; Regoli, F. Indirect effects of climate changes on cadmium bioavailability and biological effects in the Mediterranean mussel Mytilus galloprovincialis. Chemosphere 2017, 169, 493–502. [Google Scholar] [CrossRef]
- Nardi, A.; Benedetti, M.; Fattorini, D.; Regoli, F. Oxidative and interactive challenge of cadmium and ocean acidification on the smooth scallop Flexopecten glaber. Aquat. Toxicol. 2018, 196, 53–60. [Google Scholar] [CrossRef]
- Nardi, A.; Benedetti, M.; d’Errico, G.; Fattorini, D.; Regoli, F. Effects of ocean warming and acidification on accumulation and cellular responsiveness to cadmium in mussels Mytilus galloprovincialis: Importance of the seasonal status. Aquat. Toxicol. 2018, 204, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Gerringa, L.J.A.; Alderkamp, A.-C.; van Dijken, G.; Laan, P.; Middag, R.; Arrigo, K.R. Dissolved Trace Metals in the Ross Sea. Front. Mar. Sci. 2020, 7, 577098. [Google Scholar] [CrossRef]
- Scarponi, G.; Capodaglio, G.; Barbante, C.; Toscano, G.; Cecchini, M.; Gambaro, A.; Cescon, P. Concentration Changes in Cadmium and Lead in Antarctic Coastal Seawater (Ross Sea) during the Austral Summer and Their Relationship with the Evolution of Biological Activity. In Ross Sea Ecology; Faranda, F.M., Guglielmo, L., Ianora, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 585–594. [Google Scholar]
- Benedetti, M.; Nigro, M.; Regoli, F. Characterisation of antioxidant defences in three Antarctic notothenioid species from Terra Nova Bay (Ross Sea). Chem. Ecol. 2010, 26, 305–314. [Google Scholar] [CrossRef]
- Regoli, F.; Nigro, M.; Benedetti, M.; Gorbi, S.; Pretti, C.; Gervasi, P.G.; Fattorini, D. Interactions between metabolism of trace metals and xenobiotic agonists of the aryl hydrocarbon receptor in the antarctic fish Trematomus bernacchii: Environmental perspectives. Environ. Toxicol. Chem. 2005, 24, 1475–1482. [Google Scholar] [CrossRef] [PubMed]
- Cuypers, A.; Plusquin, M.; Remans, T.; Jozefczak, M.; Keunen, E.; Gielen, H.; Opdenakker, K.; Nair, A.R.; Munters, E.; Artois, T.J.; et al. Cadmium stress: An oxidative challenge. BioMetals 2010, 23, 927–940. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, M.; Martuccio, G.; Fattorini, D.; Canapa, A.; Barucca, M.; Nigro, M.; Regoli, F. Oxidative and modulatory effects of trace metals on metabolism of polycyclic aromatic hydrocarbons in the Antarctic fish Trematomus bernacchii. Aquat. Toxicol. 2007, 85, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, M.; Fattorini, D.; Martuccio, G.; Nigro, M.; Regoli, F. Interactions between trace metals (Cu, Hg, Ni, Pb) and 2,3,7,8- tetrachlorodibenzo-p-dioxin in the antarctic fish Trematomus bernacchii: Oxidative effects on biotransformation pathway. Environ. Toxicol. Chem. 2009, 28, 818–825. [Google Scholar] [CrossRef]
- Canapa, A.; Barucca, M.; Gorbi, S.; Benedetti, M.; Zucchi, S.; Biscotti, M.A.; Olmo, E.; Nigro, M.; Regoli, F. Vitellogenin gene expression in males of the Antarctic fish Trematomus bernacchii from Terra Nova Bay (Ross Sea): A role for environmental cadmium? Chemosphere 2007, 66, 1270–1277. [Google Scholar] [CrossRef]
- Giuliani, M.E.; Benedetti, M.; Nigro, M.; Regoli, F. Nrf2 and regulation of the antioxidant system in the Antarctic silverfish, Pleuragramma antarctica: Adaptation to environmental changes of pro-oxidant pressure. Mar. Environ. Res. 2017, 129, 1–13. [Google Scholar] [CrossRef]
- Benedetti, M.; Giuliani, M.E.; Regoli, F. Oxidative metabolism of chemical pollutants in marine organisms: Molecular and biochemical biomarkers in environmental toxicology. Ann. N. Y. Acad. Sci. 2015, 1340, 8–19. [Google Scholar] [CrossRef]
- Regoli, F.; Giuliani, M.E.; Benedetti, M.; Arukwe, A. Molecular and biochemical biomarkers in environmental monitoring: A comparison of biotransformation and antioxidant defense systems in multiple tissues. Aquat. Toxicol. 2011, 105 (Suppl. S3–S4), 56–66. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, M.E.; Benedetti, M.; Arukwe, A.; Regoli, F. Transcriptional and catalytic responses of antioxidant and biotransformation pathways in mussels, Mytilus galloprovincialis, exposed to chemical mixtures. Aquat. Toxicol. 2013, 134–135, 120–127. [Google Scholar] [CrossRef]
- Vieweg, I.; Benedetti, M.; Lanzoni, I.; Regoli, F.; Nahrgang, J. Antioxidant defenses in polar cod (Boreogadus saida) and responsiveness toward dietary crude oil exposure. Mar. Environ. Res. 2017, 130, 48–59. [Google Scholar] [CrossRef] [Green Version]
- Bargagli, R. Environmental contamination in Antarctic ecosystems. Sci. Total Environ. 2008, 400, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Moreira, A.; Figueira, E.; Mestre, N.C.; Schrama, D.; Soares, A.M.V.M.; Freitas, R.; Bebianno, M.J. Impacts of the combined exposure to seawater acidification and arsenic on the proteome of Crassostrea angulata and Crassostrea gigas. Aquat. Toxicol. 2018, 203, 117–129. [Google Scholar] [CrossRef]
- Abele, D.; Puntarulo, S. Formation of reactive species and induction of antioxidant defence systems in polar and temperate marine invertebrates and fish. Comp. Biochem. Phys. A 2004, 138, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Viarengo, A.; Burlando, B.; Ceratto, N.; Panfoli, I. Antioxidant role of metallothioneins: A comparative overview. Cell Mol. Biol. (Noisy-le-grand) 2000, 46, 407–417. [Google Scholar]
- Regoli, F.; Giuliani, M.E. Oxidative pathways of chemical toxicity and oxidative stress biomarkers in marine organisms. Mar. Environ. Res. 2014, 93, 106–117. [Google Scholar] [CrossRef]
- Abele, D.; Heise, K.; Pörtner, H.O.; Puntarulo, S. Temperature-dependence of mitochondrial function and production of reactive oxygen species in the intertidal mud clam Mya arenaria. J. Exp. Biol. 2002, 205, 1831–1841. [Google Scholar]
- Madeira, D.; Vinagre, C.; Diniz, M.S. Are fish in hot water? Effects of warming on oxidative stress metabolism in the commercial species Sparus aurata. Ecol. Indic. 2016, 63, 324–331. [Google Scholar] [CrossRef]
- Tomanek, L.; Zuzow, M.J.; Ivanina, A.V.; Beniash, E.; Sokolova, I.M. Proteomic response to elevated PCO2 level in eastern oysters, Crassostrea virginica: Evidence for oxidative stress. J. Exp. Biol. 2011, 214, 1836–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storch, D.; Pörtner, H.O. The protein synthesis machinery operates at the same expense in eurythermal and cold stenothermal pectinids. Physiol. Biochem. Zool. 2003, 76, 28–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viarengo, A.; Canesi, L.; Garcia Martinez, P.; Peters, L.D.; Livingstone, D.R. Pro-oxidant processes and antioxidant defence systems in the tissues of the Antarctic scallop (Adamussium colbecki) compared with the Mediterranean scallop (Pecten jacobaeus). Comp. Biochem. Phys. B 1995, 111, 119–126. [Google Scholar] [CrossRef]
- Regoli, F.; Nigro, M.; Bompadre, S.; Winston, G.W. Susceptibility to oxyradical toxicity in Antarctic, Arctic, and Mediterranean scallops. Mar. Environ. Res. 2000, 50, 547. [Google Scholar] [CrossRef]
- Camus, L.; Gulliksen, B.; Depledge, M.H.; Jones, M.B. Polar bivalves are characterized by high antioxidant defences. Polar Res. 2005, 24, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Gieseg, S.P.; Cuddihy, S.; Hill, J.V.; Davison, W. A comparison of plasma vitamin C and E levels in two Antarctic and two temperate water fish species. Comp. Biochem. Phys. B 2000, 125, 371–378. [Google Scholar] [CrossRef]
- Heise, K.; Estevez, M.S.; Puntarulo, S.; Galleano, M.; Nikinmaa, M.; Pörtner, H.O.; Abele, D. Effects of seasonal and latitudinal cold on oxidative stress parameters and activation of hypoxia inducible factor (HIF-1) in zoarcid fish. J. Comp. Physiol. B 2007, 177, 765–777. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Chen, M.G.; Ma, Q. Activation of Nrf2 in defense against cadmium-induced oxidative stress. Chem. Res. Toxicol. 2008, 21, 1375–1383. [Google Scholar] [CrossRef]
- Chen, J.; Shaikh, Z.A. Activation of Nrf2 by cadmium and its role in protection against cadmium-induced apoptosis in rat kidney cells. Toxicol. Appl. Pharm. 2009, 241, 81–89. [Google Scholar] [CrossRef]
- Choundury, C.; Mazumder, R.; Kumar, R.; Dhar, B.; Sengupta, M. Cadmium induced oxystress alters Nrf2-Keap1 signaling and triggers apoptosis in piscine head kidney macrophages. Aquat. Toxicol. 2021, 231. [Google Scholar] [CrossRef]
- Giuliani, M.E.; Regoli, F. Identification of the Nrf2-Keap1 pathway in the European eel Anguilla anguilla: Role for a transcriptional regulation of antioxidant genes in aquatic organisms. Aquat. Toxicol. 2014, 150, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Lee, O.-H.; Jain, A.K.; Papusha, V.; Jaiswal, A.K. An auto-regulatory loop between stress sensors INrf2 and Nrf2 controls their cellular abundance. J. Biol. Chem. 2007, 282, 36412–36420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carney Almroth, B.; Asker, N.; Wassmur, B.; Rosengren, M.; Jutfelt, F.; Gräns, A.; Sundell, K.; Axelsson, M.; Sturve, J. Warmer water temperature results in oxidative damage in an Antarctic fish, the bald notothen. J. Exp. Mar. Biol. Ecol. 2015, 468, 130–137. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Name | Primer Sequences | Amplicon a | Annealing b | Accession Number |
|---|---|---|---|---|---|
| mt | metallothioneins | Fwd: 5′ GTTTGACTTCCTTCATCCCTGTG 3′ Rev: 5′ TGCTGTGTTTGGTTCCTTGG 3′ | 105 | 65 | AJ011585, Z72485 |
| cat | catalase | Fwd: 5′ GCCTGATGGTTTCCGTCATA 3′ Rev: 5′ CCTGACATGTTCTTTATGCCTTG 3′ | 126 | 62 | LT962462 |
| gpx1 | glutathione peroxidase 1 | Fwd: 5′ TTGTGTTCCTGAGGGAGATG 3′ Rev: 5′ ACGTCGTTCCTGCATACTG 3′ | 103 | 58 | XM_034139369 |
| Cu/Zn-sod | Cu/Zn superoxide dismutase | Fwd: 5′ TTCTTCGAGCAGGAGAATG 3′ Rev: 5′ GATGCACCCGTTTGTATTG 3′ | 120 | 56 | AY736280 |
| nrf2 | nuclear factor erythroid 2-related factor 2 | Fwd: 5′ GAGTGAGAAGAGCGAGAACATC 3′ Rev: 5′ GGAGTATTCGGAGGGAGATAA 3′ | 127 | 57 | XM_034128735 |
| keap1 | Kelch-like ECH associating protein 1 | Fwd: 5′ AGCTACCTGGAGGCGTATAA 3′ Rev: 5′ CTTCCTCCAACAGCGTAGAAA 3′ | 122 | 62 | XM_034129957 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giuliani, M.E.; Nardi, A.; Di Carlo, M.; Benedetti, M.; Regoli, F. Transcriptional and Catalytic Responsiveness of the Antarctic Fish Trematomus bernacchii Antioxidant System toward Multiple Stressors. Antioxidants 2021, 10, 410. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10030410
Giuliani ME, Nardi A, Di Carlo M, Benedetti M, Regoli F. Transcriptional and Catalytic Responsiveness of the Antarctic Fish Trematomus bernacchii Antioxidant System toward Multiple Stressors. Antioxidants. 2021; 10(3):410. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10030410
Chicago/Turabian StyleGiuliani, Maria Elisa, Alessandro Nardi, Marta Di Carlo, Maura Benedetti, and Francesco Regoli. 2021. "Transcriptional and Catalytic Responsiveness of the Antarctic Fish Trematomus bernacchii Antioxidant System toward Multiple Stressors" Antioxidants 10, no. 3: 410. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10030410