
Moringa oleifera Protects SH-SY5YCells from DEHP-Induced Endoplasmic Reticulum Stress and Apoptosis

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Moringa Preparation, Extraction, and Analysis by Gas Chromatography—Tandem Mass Spectrometry (GC-MS/MS) and Liquid Chromatography—Tandem Mass Spectrometry (LC-MS/MS)
2.3. Cell Culture and Treatment
2.4. Cell Toxicity Assay (MTT Assay)
2.5. Reactive Oxygen Species Determination
2.6. Measurement of Mitochondrial Complex Activities
2.7. RNA isolation and qRT-PCR
2.8. Protein Extraction and Western Blotting
2.9. Mitochondrial Membrane Potential (ΔΨm) Assay
2.10. Cell Death Induced by DEHP
2.11. Caspase-3 Activity Assay
2.12. Statistical Analysis
3. Results
3.1. Analysis of MOe by LC/MS and GC/MS
3.2. Effect of MOe on DEHP-Induced Cytotoxicity
3.3. Effect of MOe on DEHP-Induced ROS Generation
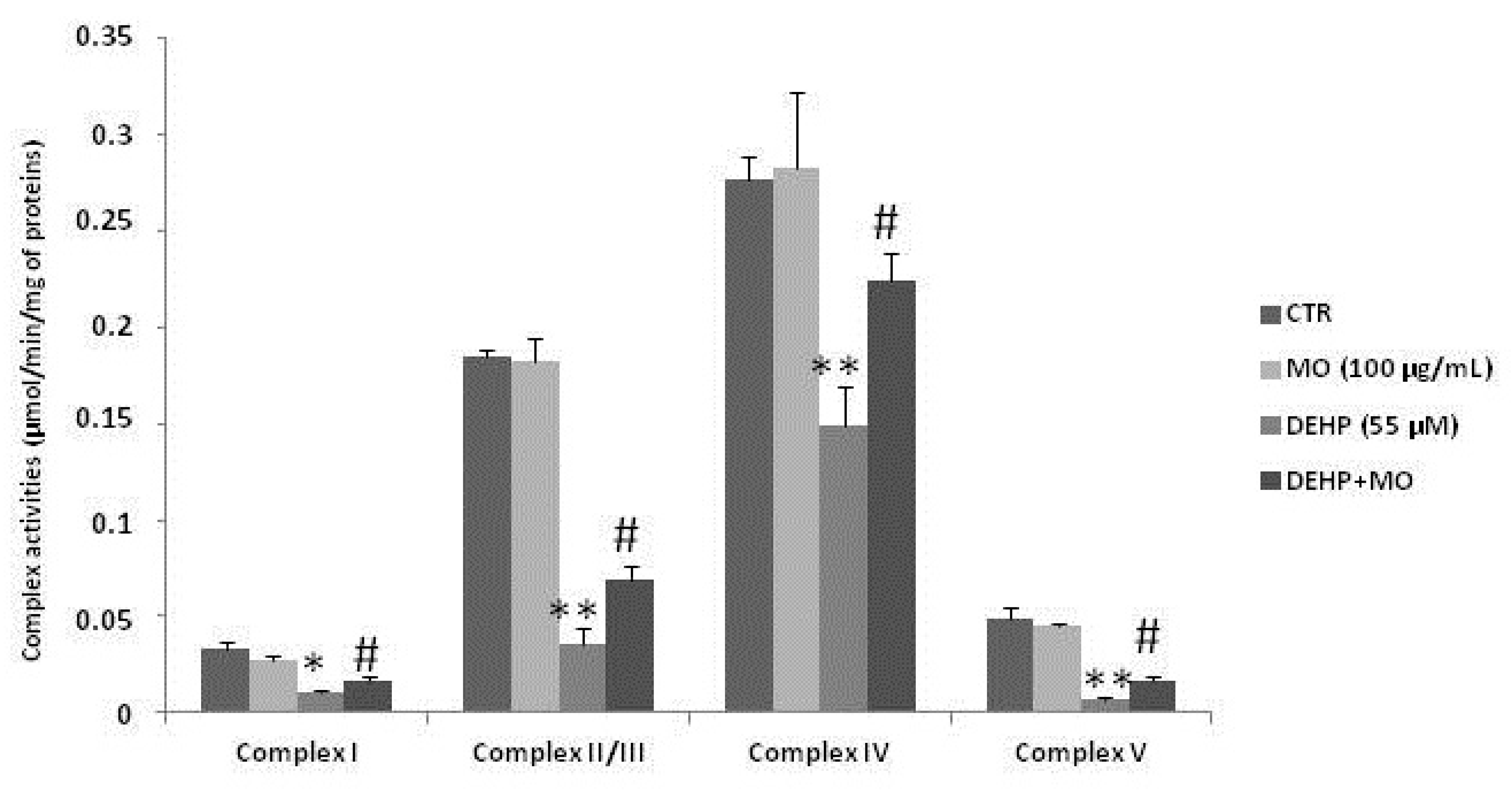
3.4. Effect of MOe on DEHP-Induced Mitochondrial Dysfunctions
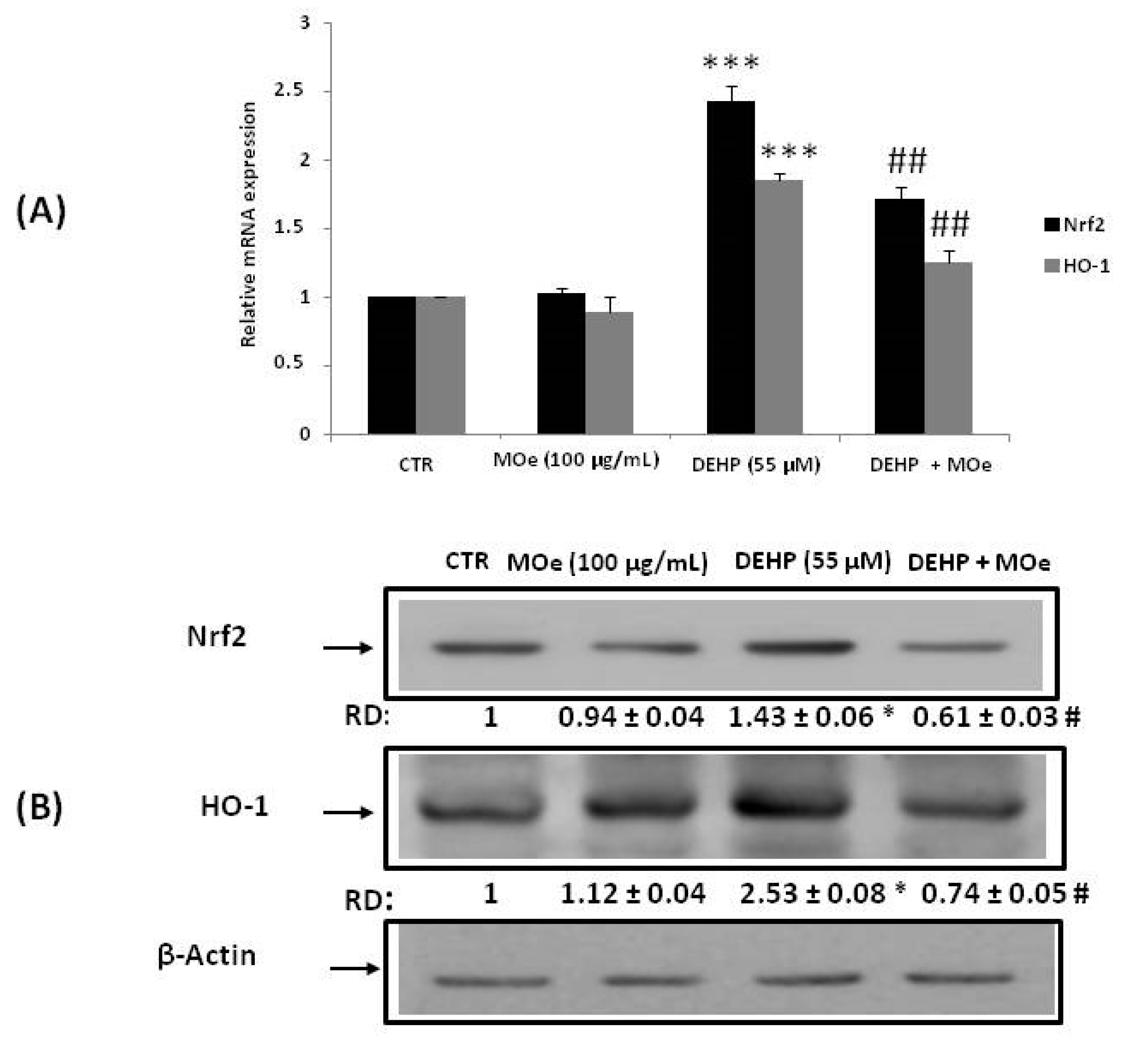
3.5. Effect of MOe on NRF2 and HO-1 Expression
3.6. Effect of MOe on DEHP-Induced Endoplasmic Reticulum (ER) Stress
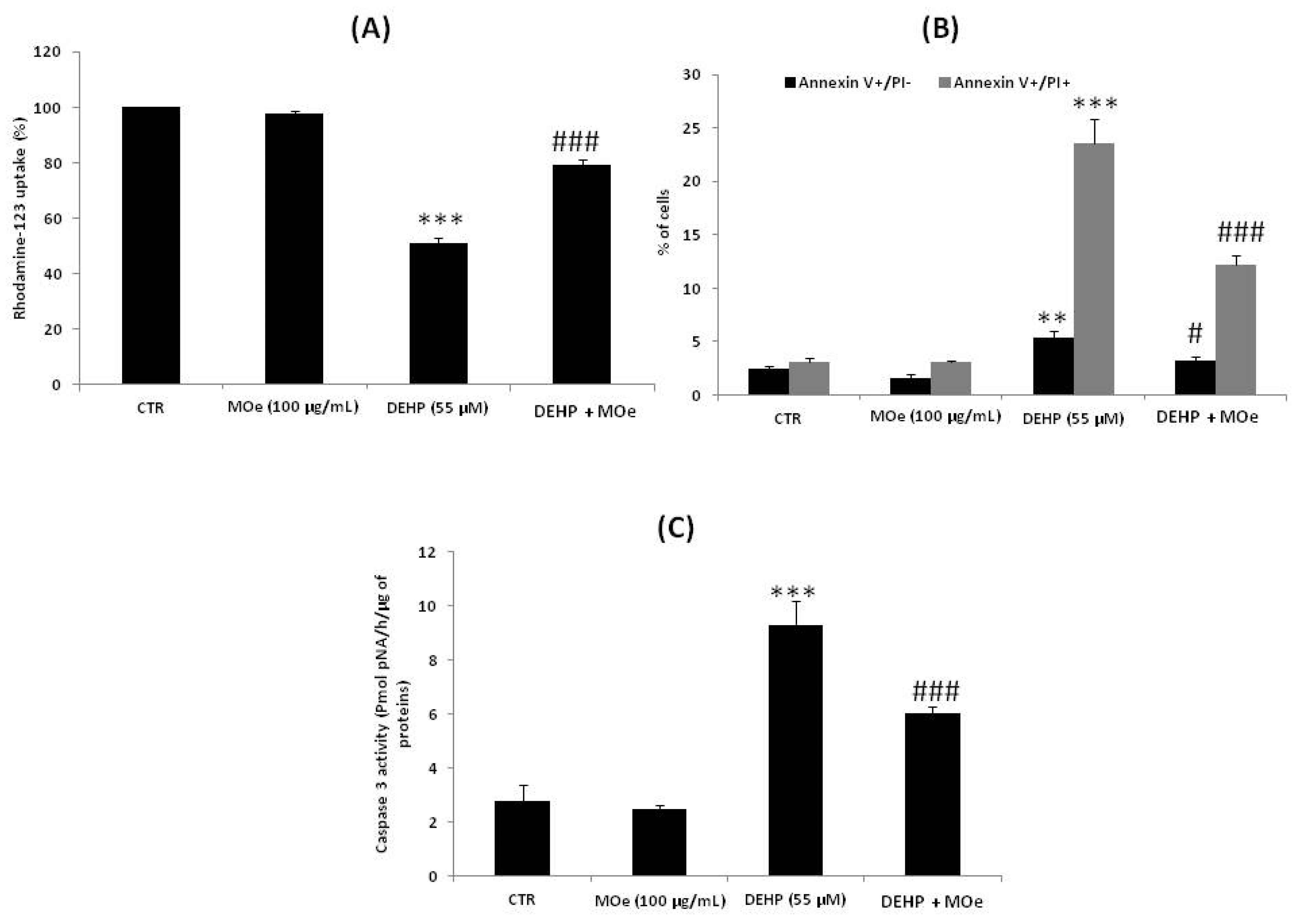
3.7. Effect of MOe on DEHP-Induced Cell Death
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MO | Moringa oleifera |
| SH-SY5Y | Human neuroblastoma cells |
| DEHP | Di-(2-ethylhexyl) phthalate |
| ROS | Reactive oxygen species |
| ER | Endoplasmic reticulum |
| Nrf2 | Nuclear factor erythroid 2-related factor |
| Keap1 | Kelch-like ECH-associated protein 1 |
| ARE | Antioxidant response element |
| NQO1 | NAD(P)H:quinone oxidoreductase 1 |
| HO-1 | Heme oxygenase 1 |
| GST | Glutathione S-transferase |
| HSP | Heat shock proteins |
| PC12 | Pheochromocytoma 12 cells |
| PBA | Chaperone 4-phenylbutyric acid |
| ΔΨm | Mitochondrial transmembrane potential |
| VSMCs | Vascular smooth muscle cells |
| Complex I | NADH:ubiquinone oxidoreductase |
| Complex II-III | Succinate-cytochrome c oxidoreductase |
| Complex IV | Cytochrome c oxidase |
| Complex V | ATP synthase |
| CTR | Control |
| UPR | Unfolded protein response |
| DMEM | Dulbecco’s Modified Eagles Medium |
| FBS | Fetal Bovine Serum |
| PBS | Phosphate Buffer Saline |
| DCFH-DA | 2,7-dichlorodihydrofluorescein diacetate |
| MTT | 3-4,5-Dimethylthiazol-2-yl,2,5-Diphenyltetrazolium Bromide |
| DCFH | Non-fluorescent dichlorodihydrofluorescein |
| AnnV/PI | Annexin V/propidium iodide |
| AMC | Aminomethylcoumarin |
References
- Rowdhwal, S.S.S.; Chen, J. Toxic Effects of Di-2-ethylhexyl Phthalate: An Overview. Biomed. Res. Int. 2018, 1, 1750368. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, B.; Lin, T.; Wu, S.; Wei, G. Protective effects of vitamin E against reproductive toxicity induced by di(2 ethylhexyl) phthalate via PPAR-dependent mechanisms. Toxicol. Mech. Methods 2017, 27, 551–559. [Google Scholar] [CrossRef]
- Chen, H.P.; Pan, M.H.; Chou, Y.Y.; Sung, C.; Lee, K.H.; Leung, C.M.; Hsu, P.C. Effects of di(2-ethylhexyl)phthalate exposure on 1,2-dimethyhydrazine-induced colon tumor promotion in rats. Food Chem. Toxicol. 2017, 103, 157–167. [Google Scholar] [CrossRef]
- Pradhan, A.; Olsson, P.E.; Jass, J. Di(2-ethylhexyl) phthalate and diethyl phthalate disrupt lipid metabolism, reduce fecundity and shortens lifespan of Caenorhabditis elegans. Chemosphere 2018, 190, 375–382. [Google Scholar] [CrossRef]
- You, M.; Dong, J.; Fu, Y.; Cong, Z.; Fu, H.; Wei, L.; Wang, Y.; Wang, Y.; Chen, J. Exposure to Di-(2-ethylhexyl) Phthalate during Perinatal Period Gender-Specifically Impairs the Dendritic Growth of Pyramidal Neurons in Rat Offspring. Front. Neurosci. 2018, 12, 444. [Google Scholar] [CrossRef] [Green Version]
- Amara, I.; Timoumi, R.; Annabi, E.; Di Rosa, G.; Scuto, M.; Najjar, M.F.; Calabrese, V.; Abid-Essefi, S. Di (2-ethylhexyl) phthalate targets the thioredoxin system and the oxidative branch of the pentose phosphate pathway in liver of Balb/c mice. Environ. Toxicol. 2020, 35, 78–86. [Google Scholar] [CrossRef]
- Luo, Y.; Li, X.N.; Zhao, Y.; Du, Z.H.; Li, J.L. DEHP triggers cerebral mitochondrial dysfunction and oxidative stress in quail (Coturnix japonica) via modulating mitochondrial dynamics and biogenesis and activating Nrf2-mediated defense response. Chemosphere 2019, 224, 626–633. [Google Scholar] [CrossRef]
- Aung, K.H.; Win-Shwe, T.T.; Kanaya, M.; Takano, H.; Tsukahara, S. Involvement of hemeoxygenase-1 in di(2-ethylhexyl) phthalate (DEHP)-induced apoptosis of Neuro-2a cells. J. Toxicol. Sci. 2014, 39, 217–229. [Google Scholar] [CrossRef] [Green Version]
- Tanida, T.; Warita, K.; Ishihara, K.; Fukui, S.; Mitsuhashi, T.; Sugawara, T.; Tabuchi, Y.; Nanmori, T.; Qi, W.M.; Inamoto, T.; et al. Fetal and neonatal exposure to three typical environmental chemicals with different mechanisms of action: Mixed exposure to phenol, phthalate, and dioxin cancels the effects of sole exposure on mouse midbrain dopaminergic nuclei. Toxicol. Lett. 2009, 189, 40–47. [Google Scholar] [CrossRef]
- Smith, C.A.; Macdonald, A.; Holahan, M.R. Acute postnatal exposure to di(2-ethylhexyl) phthalate adversely impacts hippocampal development in the male rat. Neuroscience 2011, 193, 100–108. [Google Scholar] [CrossRef]
- Masu, Y.; Morita, M.; Oka, S.; Ishido, M. Motor hyperactivity caused by a deficit in dopaminergic neurons and the effects of endocrine disruptors: A study inspired by the physiological roles of PACAP in the brain. Regul. Pept. 2004, 123, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Testa, C.; Nuti, F.; Hayek, J.; De Felice, C.; Chelli, M.; Rovero, P.; Latini, G.; Papini, A.M. Di-(2-ethylhexyl) phthalate and autism spectrum disorders. ASN Neuro 2012, 4, 223–229. [Google Scholar] [CrossRef]
- El-Shemy, H.A.; Aboul-Enein, A.M.; Aboul-Enein, K.M.; Fujita, K. Willow Leaves’ Extracts Contain Anti-Tumor Agents Effective against Three Cell Types. PLoS ONE 2007, 2, e178. [Google Scholar] [CrossRef] [Green Version]
- Leone, A.; Fiorillo, G.; Criscuoli, F.; Ravasenghi, S.; Santagostini, L.; Fico, G.; Spadafranca, A.; Battezzati, A.; Schiraldi, A.; Pozzi, F.; et al. Nutritional characterization and phenolic profiling of Moringa oleifera leale grown in chad, sahrawi refugee camps, and haiti. Int. J. Mol. Sci. 2015, 16, 18923–18937. [Google Scholar] [CrossRef] [Green Version]
- Rockwood, J.L.; Anderson, B.G.; Casamatta, D.A. Potential uses of Moringa oleifera and an examination of antibiotic efficacy conferred by M. oleifera seed and leaf extracts using crude extraction techniques available to under- served indigenous populations. Int. J. Phytothearpy Res. 2013, 3, 61–71. [Google Scholar]
- Mbikay, M. Therapeutic Potential of Moringa oleifera Leaves in Chronic Hyperglycemia and Dyslipidemia: A Review. Front. Pharmacol. 2012, 3, 24. [Google Scholar] [CrossRef] [Green Version]
- Edeogu, C.O.; Kalu, M.E.; Famurewa, A.C.; Asogwa, N.T.; Onyeji, G.N.; Ikpemo, K.O. Nephroprotective Effect of Moringa Oleifera Seed Oil on Gentamicin-Induced Nephrotoxicity in Rats: Biochemical Evaluation of Antioxidant, Anti-inflammatory, and Antiapoptotic Pathways. J. Am. Coll. Nutr. 2019, 39, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Mousa, A.A.; El-Gansh, H.A.I.; Eldaim, M.A.A.; Mohamed, M.A.E.; Morsi, A.H.; El Sabagh, H.S. Protective effect of Moringa oleifera leaves ethanolic extract against thioacetamide-induced hepatotoxicity in rats via modulation of cellular antioxidant, apoptotic and inflammatory markers. Environ. Sci. Pollut. Res. Int. 2019, 26, 32488–32504. [Google Scholar] [CrossRef]
- Elblehi, S.S.; El Euony, O.I.; El-Nahas, A.F. Partial ameliorative effect of Moringa leaf ethanolic extract on the reproductive toxicity and the expression of steroidogenic genes induced by subchronic cadmium in male rats. Environ. Sci. Pollut. Res. 2019, 26, 23306–23318. [Google Scholar] [CrossRef]
- Kandeil, M.A.; Mohammed, E.T.; Hashem, K.S.; Aleya, L.; Abdel-Daim, M.M. Moringa seed extract alleviates titanium oxide nanoparticles (TiO2-NPs)-induced cerebral oxidative damage, and increases cerebral mitochondrial viability. Environ. Sci. Pollut. Res. Int. 2019. [Google Scholar] [CrossRef] [Green Version]
- Cirmi, S.; Ferlazzo, N.; Gugliandolo, A.; Musumeci, L.; Mazzon, E.; Bramanti, A.; Navarra, M. Moringin from Moringa Oleifera Seeds Inhibits Growth, Arrests Cell-Cycle, and Induces Apoptosis of SH-SY5Y Human Neuroblastoma Cells through the Modulation of NF-κB and Apoptotic Related Factors. Int. J. Mol. Sci. 2019, 20, 1930. [Google Scholar] [CrossRef] [Green Version]
- Barhoi, D.; Upadhaya, P.; Barbhuiya, S.N.; Giri, A.; Giri, S. Aqueous Extract of Moringa oleifera Exhibit Potential Anticancer Activity and can be used as a Possible Cancer Therapeutic Agent: A Study Involving In Vitro and In Vivo Approach. J. Am. Coll. Nutr. 2020, 40, 70–85. [Google Scholar] [CrossRef]
- Tiloke, C.; Phulukdaree, A.; Chuturgoon, A.A. The antiproliferative effect of Moringa oleifera crude aqueosus leaf extract on cancerous human alveolar epithelial cells. BMC Complementary Altern. Med. 2013, 13, 226. [Google Scholar] [CrossRef] [Green Version]
- Albasher, G.; Al Kahtani, S.; Alwahibi, M.S.; Almeer, R. Effect of Moringa oleifera Lam. Methanolic extract on lead induced oxidative stress-mediated hepatic demage and inflammation in rats. Environ. Sci. Pollut. Res. Int. 2020, 27, 19877–19887. [Google Scholar] [CrossRef] [PubMed]
- Abdou, K.H.; Moselhy, W.A.; Mohamed, H.M.; El-Nahass, E.S.; Khalifa, A.G. Moringa oleifera Leaves Extract Protects Titanium Dioxide Nanoparticles-Induced Nephrotoxicity via Nrf2/HO-1 Signaling and Amelioration of Oxidative Stress. Biol. Trace Elem. Res. 2019, 187, 181–191. [Google Scholar] [CrossRef]
- Esteras, N.; Dinkova-Kostova, A.T.; Abramov, A.Y. Nrf2 activation in the treatment of neurodegenerative diseases: A focus on its role in mitochondrial bioenergetics and function. Biol. Chem. 2016, 397, 383–400. [Google Scholar] [CrossRef] [PubMed]
- Ryoo, I.G.; Kwak, M.K. Regulatory crosstalk between the oxidative stress-related transcription factor Nfe2l2/Nrf2 and mitochondria. Toxicol. Appl. Pharmacol. 2018, 359, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Li, H.X.; Guo, J.Y.; Huang, Y.Q.; Wang, H.; Talukder, M.; Li, J.L. Di (2-ethyl hexyl)phthalate (DEHP)-induced spleen toxicity in quail (Coturnix japonica) via disturbing Nrf2-mediated defense response. Environ. Pollut. 2019, 251, 984–989. [Google Scholar] [CrossRef]
- Yeager, R.L.; Reisman, S.A.; Aleksunes, L.M.; Klaassen, C.D. Introducing the “TCDD-inducible AhR-Nrf2 gene battery”. Toxicol. Sci. 2009, 111, 238–246. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.Y.; Wu, C.R.; Chang, S.W.; Wang, Y.J.; Wu, J.J.; Tsai, C.W. Induction of the pi class of glutathione S-transferase by carnosic acid in rat Clone 9 cells via the p38/Nrf2 pathway. Food Funct. 2015, 6, 1936–1943. [Google Scholar] [CrossRef]
- Bartolini, D.; Galli, F. The functional interactome of GSTP: A regulatory biomolecular network at the interface with the Nrf2 adaption response to oxidative stress. J. Chromatogr. B 2016, 1019, 29–44. [Google Scholar] [CrossRef] [PubMed]
- TrovatoSalinaro, A.; Cornelius, C.; Koverech, G.; Koverech, A.; Scuto, M.; Lodato, F.; Fronte, V.; Muccilli, V.; Reibaldi, M.; Longo, A.; et al. Cellular stress response, redox status, and vitagenes in glaucoma: A systemic oxidant disorder linked to Alzheimer’s disease. Front. Pharmacol. 2014, 5, 129. [Google Scholar]
- Calabrese, V.; Cornelius, C.; Dinkova-Kostova, A.T.; Calabrese, E.J.; Mattson, M.P. Cellular stress responses, the hormesis paradigm, and vitagenes: Novel targets for therapeutic intervention in neurodegenerative disorders. Antioxid. Redox Signal. 2010, 13, 1763–1811. [Google Scholar] [CrossRef]
- Scuto, M.C.; Mancuso, C.; Tomasello, B.; Laura Ontario, M.; Cavallaro, A.; Frasca, F.; Maiolino, L.; Trovato Salinaro, A.; Calabrese, E.J.; Calabrese, V. Curcumin, Hormesis and the Nervous System. Nutrients 2019, 11, 2417. [Google Scholar] [CrossRef] [Green Version]
- Leri, M.; Scuto, M.; Ontario, M.L.; Calabrese, V.; Calabrese, E.J.; Bucciantini, M.; Stefani, M. Healthy Effects of Plant Polyphenols: Molecular Mechanisms. Int. J. Mol. Sci. 2020, 21, 1250. [Google Scholar] [CrossRef] [Green Version]
- Scuto, M.; Di Mauro, P.; Ontario, M.L.; Amato, C.; Modafferi, S.; Ciavardelli, D.; Trovato Salinaro, A.; Maiolino, L.; Calabrese, V. Nutritional Mushroom Treatment in Meniere’s Disease with Coriolus versicolor: A Rationale for Therapeutic Intervention in Neuroinflammation and Antineurodegeneration. Int. J. Mol. Sci. 2019, 21, 284. [Google Scholar] [CrossRef] [Green Version]
- Trovato Salinaro, A.; Pennisi, M.; Di Paola, R.; Scuto, M.; Crupi, R.; Cambria, M.T.; Ontario, M.L.; Tomasello, M.; Uva, M.; Maiolino, L.; et al. Neuroinflammation and neurohormesis in the pathogenesis of Alzheimer’s disease and Alzheimer-linked pathologies: Modulation by nutritional mushrooms. Immun. Ageing 2018, 15, 8. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, V.; Butterfield, D.A.; Scapagnini, G.; Stella, A.M.; Maines, M.D. Redox regulation of heat shock protein expression by signaling involving nitric oxide and carbon monoxide: Relevance to brain aging, neurodegenerative disorders, and longevity. Antioxid. Redox Signal. 2006, 8, 444–477. [Google Scholar] [CrossRef] [PubMed]
- Trovato, A.; Siracusa, R.; Di Paola, R.; Scuto, M.; Ontario, M.L.; Bua, O.; Di Mauro, P.; Toscano, M.A.; Petralia, C.C.T.; Maiolino, L.; et al. Redox modulation of cellular stress response and lipoxin A4 expression by HericiumErinaceus in rat brain: Relevance to Alzheimer’s disease pathogenesis. Immun. Ageing 2016, 13, 23. [Google Scholar] [CrossRef] [Green Version]
- Di Rosa, G.; Brunetti, G.; Scuto, M.; TrovatoSalinaro, A.; Calabrese, E.J.; Crea, R.; Schmitz-Linneweber, C.; Calabrese, V.; Saul, N. Healthspan Enhancement by Olive Polyphenols in C. elegans Wild Type and Parkinson’s Models. Int. J. Mol. Sci. 2020, 21, 3893. [Google Scholar] [CrossRef] [PubMed]
- Scuto, M.; Trovato Salinaro, A.; Modafferi, S.; Polimeni, A.; Pfeffer, T.; Weigand, T.; Calabrese, V.; Schmitt, C.P.; Peters, V. Carnosine activates cellular stress response in podocytes and reduces glycative and lipo-peroxidative stress. Biomedicines 2020, 8, 177. [Google Scholar] [CrossRef]
- Trovato, A.; Siracusa, R.; Di Paola, R.; Scuto, M.; Fronte, V.; Koverech, G.; Luca, M.; Serra, A.; Toscano, M.A.; Petralia, A.; et al. Redox modulation of cellular stress response and lipoxin A4 expression by Coriolus versicolor in rat brain: Relevance to Alzheimer’s disease pathogenesis. Neurotoxicology 2016, 53, 350–358. [Google Scholar] [CrossRef]
- Siracusa, R.; Scuto, M.; Fusco, R.; Trovato, A.; Ontario, M.L.; Crea, R.; Di Paola, R.; Cuzzocrea, S.; Calabrese, V. Article Anti-inflammatory and Anti-oxidant Activity of Hidrox® in Rotenone-Induced Parkinson’s Disease in Mice. Antioxidants 2020, 9, 824. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, V.; Scuto, M.; Salinaro, A.T.; Dionisio, G.; Modafferi, S.; Ontario, M.L.; Greco, V.; Sciuto, S.; Schmitt, C.P.; Calabrese, E.J.; et al. Hydrogen Sulfide and Carnosine: Modulation of Oxidative Stress and Inflammation in Kidney and Brain Axis. Antioxidants 2020, 9, 1303. [Google Scholar] [CrossRef]
- Lo Dico, G.M.; Ulrici, A.; Pulvirenti, A.; Cammilleri, G.; Macaluso, A.; Vella, A.; Giaccone, V.; Lo Cascio, G.; Graci, S.; Scuto, M.; et al. Multivariate statistical analysis of the polyphenols content for the discrimination of honey produced in Sicily (Southern Italy). J. Food Compos. Anal. 2019. [Google Scholar] [CrossRef]
- Amara, I.; Scuto, M.; Zappalà, A.; Ontario, M.L.; Petralia, A.; Abid-Essefi, S.; Maiolino, L.; Signorile, A.; Trovato Salinaro, A.; Calabrese, V. Hericium Erinaceus Prevents DEHP-Induced Mitochondrial Dysfunction and Apoptosis in PC12 Cells. Int. J. Mol. Sci. 2020, 21, 2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Gutiérrez, N.; del Mar Aguilera-Luiz, M.; Romero-González, R.; Martínez Vidal, J.L.; Frenic, A.G. Fast analysis of polyphenols in royal jelly products using automated TurboFlow™-liquid chromatography-Orbitrap high resolution mass spectrometry. J. Chromatogr. B 2014, 973, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Wester, R.C.; Melendres, J.; Sedik, L.; Maibach, H.; Riviere, J.E. Percutaneous absorption of salicylic acid, theophylline, 2,4-dimethylamine, diethyl hexyl phthalic acid, and p-aminobenzoic acid in the isolated perfused porcine skin flap compared to man in vivo. Toxicol. Appl. Pharmacol. 1998, 151, 159–165. [Google Scholar] [CrossRef]
- Signorile, A.; Micelli, L.; De Rasmo, D.; Santeramo, A.; Papa, F.; Ficarella, R.; Gattoni, G.; Scacco, S.; Papa, S. Regulation of the biogenesis of OXPHOS complexes in cell transition from replicating to quiescent state Involvement of PKA and effect of hydroxytyrosol. Biochim. Biophys. Acta 2006, 1843, 675–684. [Google Scholar] [CrossRef] [Green Version]
- De Rasmo, D.; Micelli, L.; Santeramo, A.; Signorile, A.; Lattanzio, P.; Papa, S. cAMP regulates the functional activity, coupling efficiency and structural organization of mammalian FOF1 ATP synthase. Biochim. Biophys. Acta 2016, 1857, 350–358. [Google Scholar] [CrossRef]
- Morris, J.A.; Dorner, A.J.; Edwards, C.A.; Hendershot, L.M.; Kaufman, R.J. Immunoglobulin binding protein (BiP) function is required to protect cells from endoplasmic reticulum stress but is not required for the secretion of selective proteins. J. Biol. Chem. 1997, 272, 4327–4334. [Google Scholar] [CrossRef] [Green Version]
- Ozcan, U.; Yilmaz, E.; Ozcan, L.; Furuhashi, M.; Vaillancourt, E.; Smith, R.O.; Görgün, C.Z.; Hotamisligil, G.S. Chemical chaperones reduce ER stress and restore glucose homeostasis in a mouse model of type 2 diabetes. Science 2006, 313, 1137–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharaf el dein, O.; Gallerne, C.; Deniaud, A.; Brenner, C.; Lemaire, C. Role of the permeability transition pore complex in lethal inter organelle crosstalk. Front. Biosci. 2009, 14, 3465–3482. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, M.Z.; Shen, Y.; Lin, Y.; Wang, H.R.; Talukder, M.; Li, J.L. Lycopene prevents DEHP-Induced Leydig Cell Damage with the Nrf2 Antioxidant Signaling Pathway in Mice. J. Agric. Food. Chem. 2020, 68, 2031–2040. [Google Scholar] [CrossRef] [PubMed]
- Głombik, K.; Basta-Kaim, A.; Sikora-Polaczek, M.; Kubera, M.; Starowicz, G.; Styrna, J. Curcumin influences semen quality parameters and reverses the di(2 ethylhexyl)phthalate (DEHP)-induced testicular damage in mice. Pharmacol. Rep. 2014, 66, 782–787. [Google Scholar] [CrossRef] [PubMed]
- Helal, M.A. Celery oil modulates DEHP-induced reproductive toxicity in male rats. Reprod. Biol. 2014, 14, 182–189. [Google Scholar] [CrossRef]
- Abd-Ellah, M.F.; Aly, H.A.; Mokhlis, H.A.; Abdel-Aziz, A.H. Quercetin attenuates di-(2-ethylhexyl) phthalate-induced testicular toxicity in adult rats. Hum. Exp. Toxicol. 2016, 35, 232–243. [Google Scholar] [CrossRef]
- Li, J.; Zheng, L.; Wang, X.; Yao, K.; Shi, L.; Sun, X.; Yang, G.; Jiang, L.; Zhang, C.; Wang, Y.; et al. Taurine protects INS-1 cells from apoptosis induced by Di(2-ethylhexyl) phthalate via reducing oxidative stress and autophagy. Toxicol. Mech. Methods 2019, 4, 1–12. [Google Scholar] [CrossRef]
- Kang, K.B.; Lee, K.H.; Chae, S.W.; Zhang, R.; Jung, M.S.; Lee, Y.K.; Kim, S.Y.; Kim, H.S.; Joo, H.G.; Park, J.W.; et al. Eckol isolated from Ecklonia cava attenuates oxidative stress induced cell damage in lung fibroblast cells. FEBS Lett. 2005, 79, 6295–6304. [Google Scholar] [CrossRef] [Green Version]
- Mahaman, Y.A.R.; Huang, F.; Wu, M.; Wang, Y.; Wei, Z.; Bao, J.; Salissou, M.T.M.; Ke, D.; Wang, Q.; Liu, R.; et al. Moringa Oleifera Alleviates Homocysteine-Induced Alzheimer’s Disease-Like Pathology and Cognitive Impairments. J. Alzheimers Dis. 2018, 63, 1141–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaafaru, M.S.; Nordin, N.; Shaari, K.; Rosli, R.; AbdullRazis, A.F. Isothiocyanate from Moringa oleifera seeds mitigates hydrogen peroxide-induced cytotoxicity and preserved morphological features of human neuronal cells. PLoS ONE 2018, 3, e0196403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Bao, Y.; Shen, X.; Zengin, G.; Lyu, Y.; Xiao, J.; Weng, Z. Niazirin from Moringa oleifera Lam. attenuates high glucose-induced oxidative stress through PKCζ/Nox4 pathway. Phytomedicine 2019, 8, 153066. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yang, W.; Li, Z.; Luo, D.; Li, W.; Zhang, Y.; Wang, X.; Fang, M.; Chen, Q.; Jin, X. Moringa oleifera stem extract protect skin keratinocytes against oxidative stress injury by enhancement of antioxidant defense systems and activation of PPARα. Biomed. Pharmacother. 2018, 107, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Aju, B.Y.; Rajalakshmi, R.; Mini, S. Protective role of Moringa oleifera leaf extract on cardiac antioxidant status and lipid peroxidation in streptozocin induced diabetic rats. Heliyon 2019, 5, e02935. [Google Scholar] [CrossRef]
- Goudarzi, M.; Haghi Karamallah, M.; Malayeri, A.; Kalantar, M.; Mansouri, E.; Kalantar, H. Protective effect of alpha-lipoic acid on di-(2-ethylhexyl) phthalate-induced testicular toxicity in mice. Environ. Sci. Pollut. Res. Int. 2020, 27, 13670–13678. [Google Scholar] [CrossRef]
- Bahrami, N.; Goudarzi, M.; Hosseinzadeh, A.; Sabbagh, S.; Reiter, R.J.; Mehrzadi, S. Evaluating the protective effects of melatonin on di(2-ethylhexyl) phthalate-induced testicular injury in adult mice. Biomed. Pharmacother. 2018, 108, 515–523. [Google Scholar] [CrossRef]
- Bahrami, N.; Mehrzadi, S.; Goudarzi, M.; Mansouri, E.; Fatemi, I. Lycopene abrogates di-(2-ethylhexyl) phthalate induced testicular injury by modulating oxidative, endocrine and inflammatory changes in mice. Life Sci. 2018, 15, 265–271. [Google Scholar] [CrossRef]
- Lo, D.; Wang, Y.T.; Wu, M.C. Hepatoprotective effect of silymarin on di(2-ethylhexyl)phthalate (DEHP) induced injury in liver FL83B cells. Environ. Toxicol. Pharmacol. 2014, 38, 112–118. [Google Scholar] [CrossRef]
- Paradies, G.; Paradies, V.; Ruggiero, F.M.; Petrosillo, G. Mitochondrial bioenergetics and cardiolipin alterations in myocardial ischemia/ reperfusion injury. Implications for pharmacological cardioprotection. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H1341–H1352. [Google Scholar] [CrossRef]
- Marella, M.; Seo, B.B.; Yagi, A.M.; Yagi, T. Mechanism of Cell Death Caused by Complex I Defects in a Rat Dopaminergic Cell Line. J. Biol. Chem. 2007, 282, 24146–24156. [Google Scholar] [CrossRef] [Green Version]
- De Rasmo, D.; Signorile, A.; Larizza, M.; Pacelli, C.; Cocco, T.; Papa, S. Activation of the cAMP cascade in human fibroblast cultures rescues the activity of oxidatively damaged complex I. Free Radic. Biol. Med. 2012, 52, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Dröse, S.; Brandt, U.; Wittig, I. Mitochondrial respiratory chain complexes as sources and targets of thiol-based redox-regulation1. Biochim. Biophys. Acta 2014, 1844, 1344–1354. [Google Scholar] [CrossRef]
- Yu, H.-T.; Fu, X.-Y.; Liang, B.; Wang, S.; Liu, J.-K.; Wang, S.-R.; Feng, Z.-H. Oxidative damage of mitochondrial respiratory chain in different organs of a rat model of diet-induced obesity. Eur. J. Nutr. 2018. [Google Scholar] [CrossRef]
- Suzuki, T.; Yamamoto, M. Stress-sensing mechanisms and the physiological roles of the Keap1-Nrf2 system during cellular stress. J. Biol. Chem. 2017, 292, 16817–16824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basak, P.; Sadhukhan, P.; Sarkar, P.; Sil, P.C. Perspectives of the Nrf-2 signaling pathway in cancer progression and therapy. Toxicol. Rep. 2017, 4, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Hetz, C. The unfolded protein response: Controlling cell fate decisions under ER stress and beyond. Nat. Rev. Mol. Cell Biol. 2012, 13, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Glover-Cutter, K.M.; Lin, S.; Blackwell, T.K. Integration of the unfolded protein and oxidative stress responses through SKN-1/Nrf. PLoS Genet. 2013, 9, e1003701. [Google Scholar] [CrossRef] [Green Version]
- Gardner, B.M.; Pincus, D.; Gotthardt, K.; Gallagher, C.M.; Walter, P. Endoplasmic Reticulum Stress Sensing in the Unfolded Protein Response. Cold Spring Harb. Perspect. Biol. 2013, 5, a013169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sano, R.; Reed, J.C. ER stress-induced cell death mechanisms. Biochim. Biophys. Acta 2013, 1833, 3460–3470. [Google Scholar] [CrossRef] [Green Version]
- Im, J.S.; Na Lee, H.; Oh, J.W.; Yoon, Y.J.; Park, J.S.; Park, J.W.; Kim, J.H.; Kim, Y.S.; Cha, D.S.; Jeon, H. Moringa oleifera Prolongs Lifespan via DAF-16/FOXO Transcriptional Factor in Caenorhabditis elegans. Nat. Prod. Sci. 2016, 22, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Peropadre, A.; Fernández Freire, P.; Pérez Martín, J.M.; Herrero, Ó.; Hazen, M.J. Endoplasmic reticulum stress as a novel cellular response to di (2-ethylhexyl) phthalate exposure. Toxicol. Vitro 2015, 30, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Joung, H.; Kim, B.; Park, H.; Lee, K.; Kim, H.H.; Sim, H.C.; Do, H.J.; Hyun, C.K.; Do, M.S. Fermented Moringa oleifera Decreases Hepatic Adiposity and Ameliorates Glucose Intolerance in High-Fat Diet-Induced Obese Mice. J. Med. Food 2017, 20, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Zinszner, H.; Kuroda, M.; Wang, X.; Batchvarova, N.; Lightfoot, R.T.; Remotti, H.; Stevens, J.L.; Ron, D. CHOP is implicated in programmed cell death in response to impaired function of the endoplasmic reticulum. Genes Dev. 1998, 12, 982–995. [Google Scholar] [CrossRef]
- Santos, X.C.; Tanaka, L.Y.; Wosniak, J., Jr.; Laurindo, R.M. Mechanisms and Implications of Reactive Oxygen Species Generation During the Unfolded Protein Response: Roles of Endoplasmic Reticulum Oxidoreductases, Mitochondrial Electron Transport, and NADPH Oxidase. Antioxid. Redox Signal. 2009, 11, 2409–2427. [Google Scholar] [CrossRef] [PubMed]
- Lucero, M.; Suarez, A.E.; Chambers, J.W. Phosphoregulation on mitochondria: Integration of cell and organelle responses. CNS Neurosci. Ther. 2019, 25, 837–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Zhou, X.; Liu, P.; Fei, W.; Li, L.; Yun, H. Isoflurane reduces hypoxia/reoxygenation-induced apoptosis and mitochondrial permeability transition in rat primary cultured cardiocytes. BMC Anesthesiol. 2014, 14, 17. [Google Scholar] [CrossRef] [Green Version]
- Newcomb-Fernandez, J.K.; Zhao, X.; Pike, B.R. Concurrent assessment of calpain and caspase-3 activation after oxygen-glucose deprivation in primary septo-hippocampal cultures. J. Cereb. Blood Flow Metab. 2001, 21, 1281–1294. [Google Scholar] [CrossRef] [Green Version]
- Abdul Hisam, E.E.; Rofiee, M.S.; Khalid, A.M.; Jalaluddin, A.F.; Mohamad Yusof, M.I.; Idris, M.H.; Ramli, S.; James, R.J.; Jack Yoeng, W.; Lay Kek, T.; et al. Combined extract of Moringa oleifera and Centellaasiatica modulates oxidative stress and senescence in hydrogen peroxide-induced human dermal fibroblasts. Turk. J. Biol. 2018, 42, 33–44. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GC-MS/MS Analysis | ||
|---|---|---|
| Retention Time (minutes) | Name | Relative abundance (%) |
| 10.39 | Astaxanthin | 0.91 |
| 12.7 | Carotene, 1,1′,2,2′-tetrahydro-1,1′-dimethoxy | 0.18 |
| 13.46 | 1-Monolinoleoylglycerol | 1.01 |
| 17.3 | Decanoic acid | 0.19 |
| 17.78 | Rhodopin | 3.05 |
| 19.41 | Destruxin A | 0.19 |
| 20.46 | Hexadecanoic acid, ethyl ester | 0.18 |
| 20.79 | 5,8,11,14,17-Eicosapentaenoic acid, methyl ester, | 1.05 |
| 22.07 | 9-Octadecynoic acid, methyl ester | 2.01 |
| 22.18 | Methyl 8,11,14-heptadecatrienoate | 1.09 |
| 27.64 | 2-Nonadecanone 2,4-dinitrophenylhydrazine | 3.49 |
| 28.82 | Quercetin | 5.31 |
| 30.08 | Gallic acid | 41.8 |
| 32.47 | Oleic acid, 3-(octadecyloxy)propyl ester | 13.26 |
| 35.14 | Vitamin E | 6.91 |
| 36.25 | Kaempferol | 7.79 |
| 37.21 | β-Sitosterol | 6.63 |
| 39.12 | Oleyl oleate | 4.95 |
| LC-OrbitrapTM-MS Analysis | ||
| Retention Time (minutes) | Name (accurate masses) | Relative abundance (%) |
| 2.71 | Gallic acid (168.96) | 11.45 |
| 3.61 | Chlorogenic acid (353.09) | 11.21 |
| 5.02 | Vanillin (150.99) | 4.52 |
| 5.72 | Syringic acid (197.10) | 10.63 |
| 6.66 | Ferulic acid (193.03) | 14.75 |
| 6.84 | Rutin (609.11) | 3.32 |
| 7.32 | Quercetin (301.01) | 15.11 |
| 8.53 | Kaempferol (285.04) | 29.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amara, I.; Ontario, M.L.; Scuto, M.; Lo Dico, G.M.; Sciuto, S.; Greco, V.; Abid-Essefi, S.; Signorile, A.; Salinaro, A.T.; Calabrese, V. Moringa oleifera Protects SH-SY5YCells from DEHP-Induced Endoplasmic Reticulum Stress and Apoptosis. Antioxidants 2021, 10, 532. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10040532
Amara I, Ontario ML, Scuto M, Lo Dico GM, Sciuto S, Greco V, Abid-Essefi S, Signorile A, Salinaro AT, Calabrese V. Moringa oleifera Protects SH-SY5YCells from DEHP-Induced Endoplasmic Reticulum Stress and Apoptosis. Antioxidants. 2021; 10(4):532. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10040532
Chicago/Turabian StyleAmara, Ines, Maria Laura Ontario, Maria Scuto, Gianluigi Maria Lo Dico, Sebastiano Sciuto, Valentina Greco, Salwa Abid-Essefi, Anna Signorile, Angela Trovato Salinaro, and Vittorio Calabrese. 2021. "Moringa oleifera Protects SH-SY5YCells from DEHP-Induced Endoplasmic Reticulum Stress and Apoptosis" Antioxidants 10, no. 4: 532. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10040532