
Flunixin Meglumine Reduces Milk Isoprostane Concentrations in Holstein Dairy Cattle Suffering from Acute Coliform Mastitis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Selection and Study Design
2.2. Blood and Milk Sample Collection
2.3. Blood and Milk Biomarker Analysis
2.4. Reagents
2.5. Solid-Phase Extraction (SPE) of Blood and Milk
2.6. LC/MS/MS Analysis
2.7. Statistical Analysis
3. Results
3.1. Survival Rate
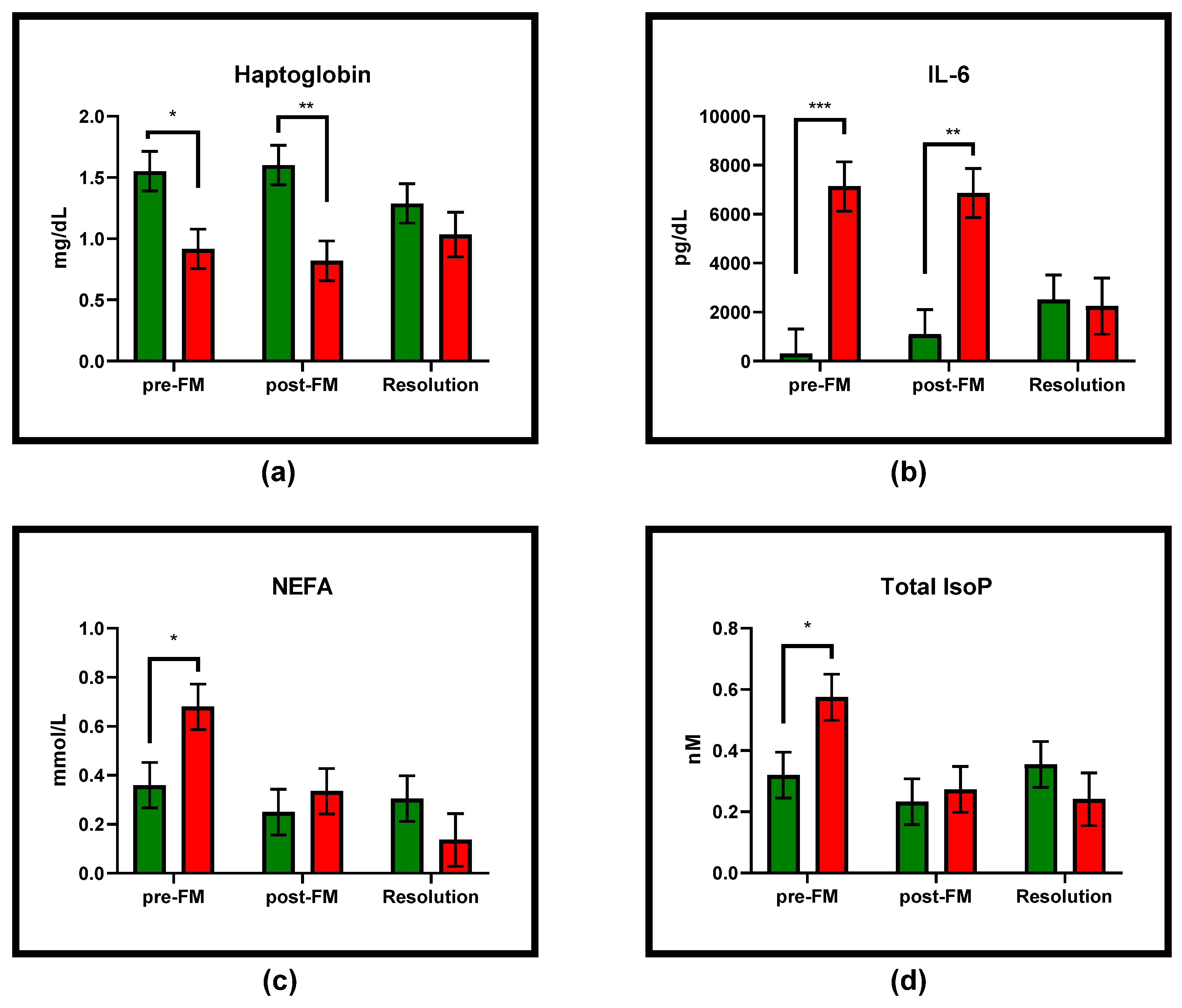
3.2. Blood
3.3. Milk
4. Discussion
4.1. Traditional Inflammatory Markers
4.2. Oxidant Status and Isoprostanes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rollin, E.; Dhuyvetter, K.C.; Overton, M.W. The cost of clinical mastitis in the first 30 days of lactation: An economic modeling tool. Prev. Vet. Med. 2015, 122, 257–264. [Google Scholar] [CrossRef] [Green Version]
- National Animal Health Monitoring Service (NAHMS). Dairy 2014. Part. 1: Reference of Dairy Health and Management in the United States; USDA APHIS Veterinary Services: Fort Collins, CO, USA, 2014.
- Erskine, R.J.; Eberhart, R.J.; Hutchinson, L.J.; Spencer, S.B.; A Campbell, M. Incidence and types of clinical mastitis in dairy herds with high and low somatic cell counts. J. Am. Veter. Med. Assoc. 1988, 192, 761–765. [Google Scholar]
- Nakajima, N.; Mikami, O.; Yoshioka, M.; Motoi, Y. Elevated levels of tumor necrosis factor-a (TNFA-a) and interleukin-6 (IL-6) activities in the sera and milk of cows with naturally occurring coliform mastitis. Res. Vet. Sci. 1997, 62, 297–298. [Google Scholar] [CrossRef]
- Ohtsuka, H.; Kudo, K.; Mori, K.; Nagai, F.; Hatsugaya, A.; Tajima, M.; Tamura, K.; Hoshi, F.; Koiwa, M.; Kawamura, S.-I. Acute Phase Response in Naturally Occurring Coliform Mastitis. J. Veter. Med. Sci. 2001, 63, 675–678. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, L.; Hulland, C.; Ruegg, P. Characterization of clinical mastitis occurring in cows on 50 large dairy herds in Wis-consin. J. Dairy Sci. 2013, 96, 7538–7549. [Google Scholar] [CrossRef] [PubMed]
- Sordillo, L.; Pighetti, G.; Davis, M. Enhanced production of bovine tumor necrosis factor-α during the periparturient period. Veter. Immunol. Immunopathol. 1995, 49, 263–270. [Google Scholar] [CrossRef]
- Hoeben, D.; Burvenich, C.; Trevisi, E.; Bertoni, G.; Hamann, J.; Bruckmaier, R.M.; Blum, J.W. Role of endotoxin and TNF-α in the pathogenesis of experimentally induced coliform mastitis in periparturient cows. J. Dairy Res. 2000, 67, 503–514. [Google Scholar] [CrossRef]
- De Schepper, S.; De Ketelaere, A.; Bannerman, D.D.; Paape, M.J.; Peelman, L.; Burvenich, C. The toll-like receptor-4 (TLR-4) pathway and its possible role in the pathogenesis of Escherichia coli mastitis in dairy cattle. Vet. Res. 2008, 39, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Aitken, S.L.; Corl, C.M.; Sordillo, L.M. Pro-inflammatory and pro-apoptotic responses of TNF-α stimulated bovine mammary endothelial cells. Veter. Immunol. Immunopathol. 2011, 140, 282–290. [Google Scholar] [CrossRef]
- Aitken, S.L.; Corl, C.M.; Sordillo, L.M. Immunopathology of Mastitis: Insights into Disease Recognition and Resolution. J. Mammary Gland. Biol. Neoplasia 2011, 16, 291–304. [Google Scholar] [CrossRef]
- Ryman, V.E.; Packiriswamy, N.; Sordillo, L.M. Role of endothelial cells in bovine mammary gland health and disease. Anim. Health Res. Rev. 2015, 16, 135–149. [Google Scholar] [CrossRef]
- Sordillo, L.M. Mammary Gland Immunobiology and Resistance to Mastitis. Veter. Clin. N. Am. Food Anim. Pr. 2018, 34, 507–523. [Google Scholar] [CrossRef] [PubMed]
- Mavangira, V.; Brown, J.; Gandy, J.C.; Sordillo, L.M. 20-hydroxyeicosatetraenoic acid alters endothelial cell barrier integrity independent of oxidative stress and cell death. Prostaglandins Other Lipid Mediat. 2020, 149, 106425. [Google Scholar] [CrossRef]
- Bodiga, S.; Gruenloh, S.K.; Gao, Y.; Manthati, V.L.; Dubasi, N.; Falck, J.R.; Medhora, M.; Jacobs, E.R. 20-HETE-induced nitric oxide production in pulmonary artery endothelial cells is mediated by NADPH oxidase, H2O2, and PI3-kinase/Akt. Am. J. Physiol. Cell. Mol. Physiol. 2010, 298, L564–L574. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in Inflammation, Immunity, and Disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Porcherie, A.; Cunha, P.; Trotereau, A.; Roussel, P.; Gilbert, F.B.; Rainard, P.; Germon, P. Repertoire of Escherichia coli agonists sensed by innate immunity receptors of the bovine udder and mammary epithelial cells. Veter. Res. 2012, 43, 14. [Google Scholar] [CrossRef] [Green Version]
- Kumar, H.; Kawai, T.; Akira, S. Pathogen Recognition by the Innate Immune System. Int. Rev. Immunol. 2011, 30, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Goldammer, T.; Zerbe, H.; Molenaar, A.; Schuberth, H.-J.; Brunner, R.M.; Kata, S.R.; Seyfert, H.-M. Mastitis increases mammary mRNA abundance of b-defensin 5, toll-like-receptor 2 (TLR2), and TLR4 but not TLR9 in cattle. J. Clin. Diag. Lab. Immunol. 2004, 11, 174–185. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Zhou, Y.; Shen, P. NF-kappaB and its regulation on the immune system. Cell. Mol. Immunol. 2004, 1, 343–350. [Google Scholar]
- Lloyd, E.E.; Gaubatz, J.W.; Burns, A.R.; Pownall, H.J. Sustained elevations in NEFA induce cycloocygenase-2 activity and potentiate THP-1 macrophage foam cell formation. Atherosclerosis 2007, 192, 49–55. [Google Scholar] [CrossRef]
- Hinson, R.M.; Williams, J.A.; Shacter, E. Elevated interleukin 6 is induced by prostaglandin E2 in a murine model of in-flammation: Possible role of cyclooxygenase-2. Proc. Natl. Acad. Sci. USA 1996, 93, 4885–4890. [Google Scholar] [CrossRef] [Green Version]
- Contreras, G.A.; Sordillo, L.M. Lipid mobilization and inflammatory responses during the transition period of dairy cows. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 281–289. [Google Scholar] [CrossRef]
- Sordillo, L.M.; Mavangira, V. The nexus between nutrient metabolism, oxidative stress and inflammation in transition cows. Anim. Prod. Sci. 2014, 54, 1204–1214. [Google Scholar] [CrossRef]
- Mavangira, V.; Mangual, M.; Gandy, J.; Sordillo, L. 15-F 2t -Isoprostane Concentrations and Oxidant Status in Lactating Dairy Cattle with Acute Coliform Mastitis. J. Veter. Intern. Med. 2016, 30, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, M.J.; Mavangira, V.; Gandy, J.C.; Sordillo, L.M. Production of 15-F2t-isoprostane as an assessment of oxidative stress in dairy cows at different stages of lactation. J. Dairy Sci. 2018, 101, 9287–9295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.S.; White, A.; Perry, R.J.; Camporez, J.P.; Hidalgo, J.; Shulman, G.I.; Davis, R.J. Regulation of adipose tissue inflammation by interleukin 6. Proc. Natl. Acad. Sci. USA 2020, 117, 2751–2760. [Google Scholar] [CrossRef] [Green Version]
- Hussey, S.E.; Lum, H.; Alvarez, A.; Cipriani, Y.; Garduño-Garcia, J.; Anaya, L.; Dube, J.; Musi, N. A sustained increase in plasma NEFA upregulates the Toll-like receptor netowrk in human muscle. Diabetologia 2013, 57, 583–591. [Google Scholar]
- Contreras, G.A.; Raphael, W.; Mattmiller, S.A.; Gandy, J.; Sordillo, L.M. Nonesterified fatty acids modify inflammatory re-sponse and eicosanoid biosynthesis in bovine endothelial cells. J. Dairy Sci. 2012, 95, 5011–5023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, X.; Zhang, H.; Zhao, Z.; Peng, Z.; Wang, Z.; Liu, G.; Li, X. Non-esterified fatty acids over-activate the TLR2/4-NK-Kb signaling pathway to increase inflammatory cytokine synthesis in neutrophils from ketotic cows. Cell. Physiol. Biochem. 2018, 48, 827–837. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sordillo, L.M.; Aitken, S.L. Impact of oxidative stress on the health and immune function of dairy cattle. Veter. Immunol. Immunopathol. 2009, 128, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Lykkesfeldt, J.; Svendsen, O. Oxidants and antioxidants in disease: Oxidative stress in farm animals. Veter. J. 2007, 173, 502–511. [Google Scholar] [CrossRef]
- Hernanz, R.; Briones, A.M.; Salaices, M.; Alonso, M.J. New roles for old pathways? A circuitous relationship between reactive oxygen species and cyclo-oxygenase in hypertension. Clin. Sci. 2013, 126, 111–121. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [Green Version]
- Milne, G.L.; Dai, Q.; Roberts, L.J., II. The isoprostanes—25 years later. Biochim. Biophys. Acta 2015, 1851, 433–445. [Google Scholar] [CrossRef] [Green Version]
- Milne, G.L.; Yin, H.; Brooks, J.D.; Sanchez, S.; Roberts II, L.J.; Morrow, J.D. Quantification of F2-isoprostanes in biological fluids and tissues as a measure of oxidant stress. Methods Enzymol. 2007, 433, 113–126. [Google Scholar]
- Celi, P. Biomarkers of oxidative stress in ruminant medicine. Immunopharmacol. Immunotoxicol. 2010, 33, 233–240. [Google Scholar] [CrossRef]
- US FDA (United States Food and Drug Administration). Implantation or Injectable Dosage Form New Animal Drugs; Flunixin Meglumine; US Federal Register: White Oak, ML, USA, 1998; 63, pp. 38749–38750.
- Myers, M.J.; Scott, M.L.; Deaver, C.M.; Farrell, D.E.; Yancy, H.F. Biomarkers of inflammation in cattle determining the ef-fectiveness of anti-inflammatory drugs. J. Vet. Pharmacol. Therap. 2009, 33, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yeiser, E.; Leslie, K.; McGilliard, M.; Petersson-Wolfe, C. The effects of experimentally induced Escherichia coli mastitis and flunixin meglumine administration on activity measures, feed intake, and milk parameters. J. Dairy Sci. 2012, 95, 4939–4949. [Google Scholar] [CrossRef] [PubMed]
- Stock, M.L.; Coetzee, J.F. Clinical Pharmacology of Analgesic Drugs in Cattle. Food Anim. Pract. 2015, 31, 113–138. [Google Scholar] [CrossRef] [PubMed]
- Donalisio, C.; Barbero, R.; Cuniberti, B.; Ver Celli, C.; Casalone, M.; Re, G. Effects of flunixin meglumine and ketoprofen on mediator production in ex vivo and in vitro models of inflammation in healthy dairy cows. J. Vet. Parmacol. Therap. 2012, 36, 130–139. [Google Scholar] [CrossRef]
- Sintes, G.; Bruckmaier, R.; Wellnitz, O. Nonsteroidal anti-inflammatory drugs affect the mammary epithelial barrier during inflammation. J. Dairy Sci. 2020, 103, 10742–10753. [Google Scholar] [CrossRef]
- Putman, A.; Brown, J.; Gandy, J.; Wisnieski, L.; Sordillo, L. Changes in biomarkers of nutrient metabolism, inflammation, and oxidative stress in dairy cows during the transition into the early dry period. J. Dairy Sci. 2018, 101, 9350–9359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Mavangira, V.; Gandy, J.C.; Zhang, C.; Ryman, V.E.; Jones, A.D.; Sordillo, L.M. Polyunsaturated fatty acids influence differential biosynthesis of oxylipids and other lipid mediators during bovine coliform mastitis. J. Dairy Sci. 2015, 98, 6202–6215. [Google Scholar] [CrossRef] [PubMed]
- Pezeshki, A.; Stordeur, P.; Wallemacq, H.; Schynts, F.; Stevens, M.; Boutet, P.; Peelman, L.J.; De Spiegeleer, B.; Duchateau, L.; Bureau, F.; et al. Variation of inflammatory dynamics and mediators in primiparous cows after intramammary challenge with Escherichia coli. Veter. Res. 2011, 42, 15. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, A.C.C.; Davoodi, S.; Kaur, M.; Veira, D.; Melo, L.E.H.; Cerri, R.L.A. Effect of repeated intravenous lipopolyssa-charide infusions on systemic inflammatory response and edomentrium gene expression in Holstein heifers. J. Dairy Sci. 2019, 102, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sordillo, L.M.; Peel, J.E. Effect of interferon-y on the production of tumor necrosis factor during acute Escherichia coli mastitis. J. Dairy Sci. 1992, 75, 2119–2125. [Google Scholar] [CrossRef]
- Garaude, J.; Acín-Pérez, R.; Martínez-Cano, S.; Enamorado, M.; Ugolini, M.; Nistal-Villán, E.; Hervás-Stubbs, E.N.-V.S.; Pelegrín, P.; E Sander, M.U.L.; Enríquez, J.A.; et al. Mitochondrial respiratory-chain adaptations in macrophages contribute to antibacterial host defense. Nat. Immunol. 2016, 17, 1037–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zentella de Piña, M.; Vázquez-Meza, H.; Agundis, C.; Pereyra, M.A.; Pardo, J.P.; Villalobos-Molina, R.; Piña, E. Inhibition of cAMP-dependent protein kinase A: A novel cyclo-oxygenase-independent effect of non-steroidal anti-inflammatory drugs in adipocytes. Auton. Autacoid Pharmacol. 2007, 27, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.L.; Malkowski, M.G. Interactions of fatty acids, nonsteroidal anti-inflammatory drugs, and coxibs with the catalytic and allosteric subunits of cyclooxygenases-1 and -2. J. Biol. Chem. 2019, 294, 1697–1705. [Google Scholar] [CrossRef] [Green Version]
- Mavangira, V.; Sordillo, L.M. Role of lipid mediators in the regulation of oxidative stress and inflammatory responses in dairy cattle. Res. Veter. Sci. 2018, 116, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Van’t Erve, T.J.; Lih, F.B.; Kadiiska, M.B.; Deterding, L.J.; Eling, T.E.; Mason, R.P. Reinterpreting the best biomarker of oxi-dative stress: The 8-iso-PGF2α/PGF2α ratio distinguishes chemical from enzymatic lipid peroxidation. Free Radic. Biol. Med. 2015, 83, 245–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, H.; Yuan, C.; Dong, L.; Sidhu, R.S.; Hong, Y.H.; Kuklev, D.V.; Smith, W.L. Human cyclooxygenase-1 activity and its responses to COX inhibitors are allosterically regulated by nonsubstrate fatty acids. J. Lipid Res. 2012, 53, 1336–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, L.; Sharma, N.P.; Jurban, B.J.; Smith, W.L. Pre-existent asymmetry in the human cyclooxygenase-2 sequence ho-modimer. J. Biol. Chem. 2013, 288, 28641–28655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, L.; Yuan, C.; Orlando, B.J.; Malkowski, M.G.; Smith, W.L. Fatty acid binding to the allosteric subunit of cyclooxygen-ase-2 relieves a tonic inhibition of the catalytic subunit. J. Biol. Chem. 2016, 291, 25641–25655. [Google Scholar] [CrossRef] [Green Version]
- Milne, G.L.; Yin, H.; Hardy, K.D.; Davies, S.S.; Roberts, L.J., 2nd. Isoprostane generation and function. Chem. Rev. 2011, 111, 5973–5996. [Google Scholar] [CrossRef] [Green Version]
- Caldeira, M.; Bruckmaier, R.; Wellnitz, O. Effects of local or systemic administration of meloxicam on mammary gland inflammatory responses to lipopolysaccharide-induced mastitis in dairy cows. J. Dairy Sci. 2021, 104, 1039–1052. [Google Scholar] [CrossRef] [PubMed]
- Barragan, A.; Hovingh, E.; Bas, S.; Lakritz, J.; Byler, L.; Ludwikowski, A.; Takitch, S.; Zug, J.; Hann, S. Effects of postpartum acetylsalicylic acid on metabolic status, health, and production in lactating dairy cattle. J. Dairy Sci. 2020, 103, 8443–8452. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| (a) All animals | Concentrations | ||||||||
| Pre-FM Treatment (N = 8) | Post-FM Treatment (N = 8) | Resolution (N = 7) | |||||||
| Biomarker | Mean | SEM | Mean | SEM | Mean | SEM | |||
| ROS (RFU/µL) | 2.358 ab | ± | 0.227 | 1.859 a | ± | 0.227 | 2.914 b | ± | 0.239 |
| AOP (TE/µL) | 4.415 a | ± | 0.170 | 5.043 b | ± | 0.170 | 4.270 a | ± | 0.181 |
| OSi (ROS/AOP) | 0.538 a | ± | 0.056 | 0.373 b | ± | 0.056 | 0.716 c | ± | 0.059 |
| (b) Survival only | Concentrations | ||||||||
| Pre-FM Treatment (N = 4) | Post-FM Treatment (N = 4) | Resolution (N = 4) | |||||||
| Biomarker | Mean | SEM | Mean | SEM | Mean | SEM | |||
| ROS (RFU/µL) | 2.528 a | ± | 0.310 | 1.880 a | ± | 0.310 | 2.535 a | ± | 0.310 |
| AOP (TE/µL) | 4.532 ab | ± | 0.224 | 5.335 a | ± | 0.224 | 4.505 b | ± | 0.224 |
| OSi (ROS/AOP) | 0.562 a | ± | 0.073 | 0.348 b | ± | 0.073 | 0.602 a | ± | 0.073 |
| (a) All animals | Concentrations | ||||||||
| Pre-FM Treatment (N = 8) | Post-FM Treatment (N = 8) | Resolution (N = 7) | |||||||
| Biomarker | Mean | SEM | Mean | SEM | Mean | SEM | |||
| ROS (RFU/µL) | 15,221.000 a | ± | 2985.490 | 7695.380 b | ± | 2985.490 | 11,000.00 ab | ± | 3066.370 |
| AOP (TE/µL) | 6.363 a | ± | 0.900 | 8.563 b | ± | 0.900 | 6.918 a | ± | 0.922 |
| OSi (ROS/AOP) | 3750.410 a | ± | 1031.890 | 1467.460 b | ± | 1031.890 | 3031.870 ab | ± | 1059.940 |
| (b) Survival only | Concentrations | ||||||||
| Pre-FM Treatment (N = 4) | Post-FM Treatment (N = 4) | Resolution (N = 4) | |||||||
| Biomarker | Mean | SEM | Mean | SEM | Mean | SEM | |||
| ROS (RFU/µL) | 11,757.000 a | ± | 4339.200 | 7317.380 a | ± | 4339.200 | 9031.750 a | ± | 4339.200 |
| AOP (TE/µL) | 7.750 a | ± | 0.578 | 11.550 b | ± | 0.578 | 9.325 a | ± | 0.578 |
| OSi (ROS/AOP) | 1556.820 a | ± | 1056.530 | 646.830 a | ± | 1056.530 | 984.500 a | ± | 1056.530 |
| (c) Death only | Concentrations | ||||||||
| Pre-FM Treatment (N = 4) | Post-FM Treatment (N = 4) | Resolution (N = 3) | |||||||
| Biomarker | Mean | SEM | Mean | SEM | Mean | SEM | |||
| ROS (RFU/µL) | 18,685.000 a | ± | 4339.200 | 8073.380 b | ± | 4339.200 | 13,548.000 ab | ± | 4566.120 |
| AOP (TE/µL) | 4.975 a | ± | 0.578 | 5.575 a | ± | 0.578 | 4.537 a | ± | 0.636 |
| OSi (ROS/AOP) | 5944.010 a | ± | 1056.530 | 2288.080 b | ± | 1056.530 | 5542.340 a | ± | 1111.05 |
| (a) All animals | Concentrations | ||||||||
| Pre-FM Treatment (N = 8) | Post-FM Treatment (N = 8) | Resolution (N = 7) | |||||||
| Biomarker (nM) | Mean | SEM | Mean | SEM | Mean | SEM | |||
| 5-iso-iPF2α-VI | 9.300 a | ± | 1.3163 | 3.550 b | ± | 1.3163 | 3.993 b | ± | 1.3867 |
| 8,12-iso-iPF2α-VI | 22.388 a | ± | 3.1743 | 7.225 b | ± | 3.1743 | 7.458 b | ± | 3.3755 |
| 8-iso-15®-PGF2α | 2.100 a | ± | 0.364 | 1.600 a | ± | 0.364 | 1.119 a | ± | 0.3878 |
| 8-iso-15-keto-PGE2 | 3.500 a | ± | 0.6616 | 1.538 a | ± | 0.6616 | 2.206 a | ± | 0.7058 |
| Total IsoP | 37.288 a | ± | 4.769 | 13.913 b | ± | 4.769 | 14.733 b | ± | 5.0478 |
| (b) Survival only | Concentrations | ||||||||
| Pre-FM Treatment (N = 4) | Post-FM Treatment (N = 4) | Resolution (N = 4) | |||||||
| Biomarker (nM) | Mean | SEM | Mean | SEM | Mean | SEM | |||
| 5-iso-iPF2α-VI | 10.775 a | ± | 1.9483 | 3.400 b | ± | 1.9483 | 4.525 ab | ± | 1.9483 |
| 8,12-iso-iPF2α-VI | 26.350 a | ± | 4.6684 | 7.250 b | ± | 4.6684 | 8.125 b | ± | 4.6684 |
| 8-iso-15®-PGF2α | 2.050 a | ± | 0.5489 | 1.625 a | ± | 0.5489 | 1.400 a | ± | 0.5489 |
| 8-iso-15-keto-PGE2 | 3.300 a | ± | 0.996 | 1.575 a | ± | 0.996 | 1.725 a | ± | 0.996 |
| Total IsoP | 42.475 a | ± | 7.0826 | 13.850 b | ± | 7.0826 | 15.775 b | ± | 7.0826 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walker, C.C.F.; Brester, J.L.; Sordillo, L.M. Flunixin Meglumine Reduces Milk Isoprostane Concentrations in Holstein Dairy Cattle Suffering from Acute Coliform Mastitis. Antioxidants 2021, 10, 834. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10060834
Walker CCF, Brester JL, Sordillo LM. Flunixin Meglumine Reduces Milk Isoprostane Concentrations in Holstein Dairy Cattle Suffering from Acute Coliform Mastitis. Antioxidants. 2021; 10(6):834. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10060834
Chicago/Turabian StyleWalker, Carsten C. F., Jill L. Brester, and Lorraine M. Sordillo. 2021. "Flunixin Meglumine Reduces Milk Isoprostane Concentrations in Holstein Dairy Cattle Suffering from Acute Coliform Mastitis" Antioxidants 10, no. 6: 834. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10060834