
Metabolomics Reveals Nutritional Diversity among Six Coarse Cereals and Antioxidant Activity Analysis of Grain Sorghum and Sweet Sorghum
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Sample Preparation and Extraction
2.3. High-Performance Liquid Chromatography Conditions
2.4. ESI-Q TRAP-MS/MS
2.5. Identification of Metabolites
2.6. Determination of Flavonoids and Anthocyanin Contents
2.7. Determination of Antioxidant System Activity
2.8. Statistical Analysis
3. Results
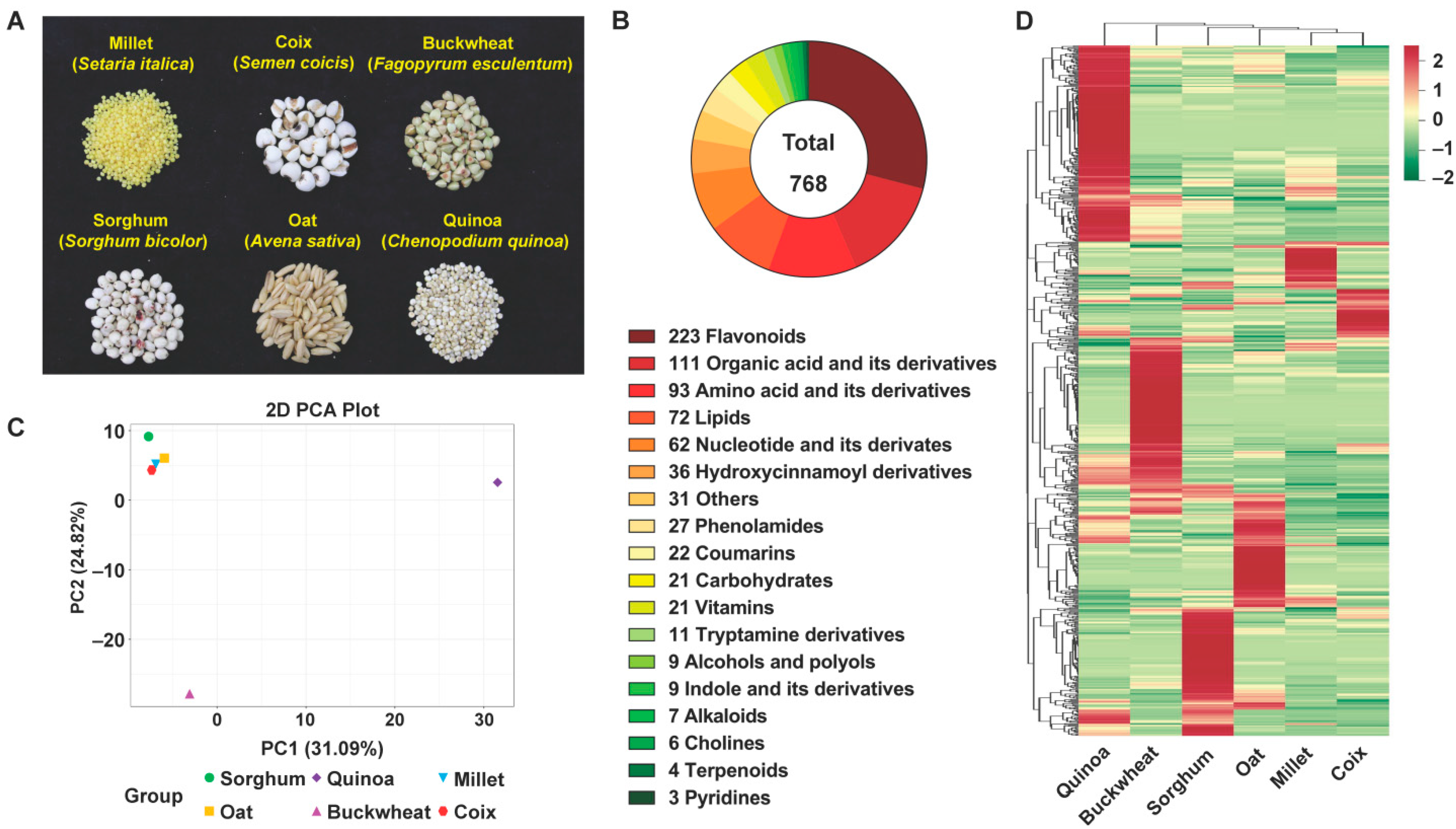
3.1. Whole Metabolome-Scale Comparative Analysis of Six Coarse Cereals
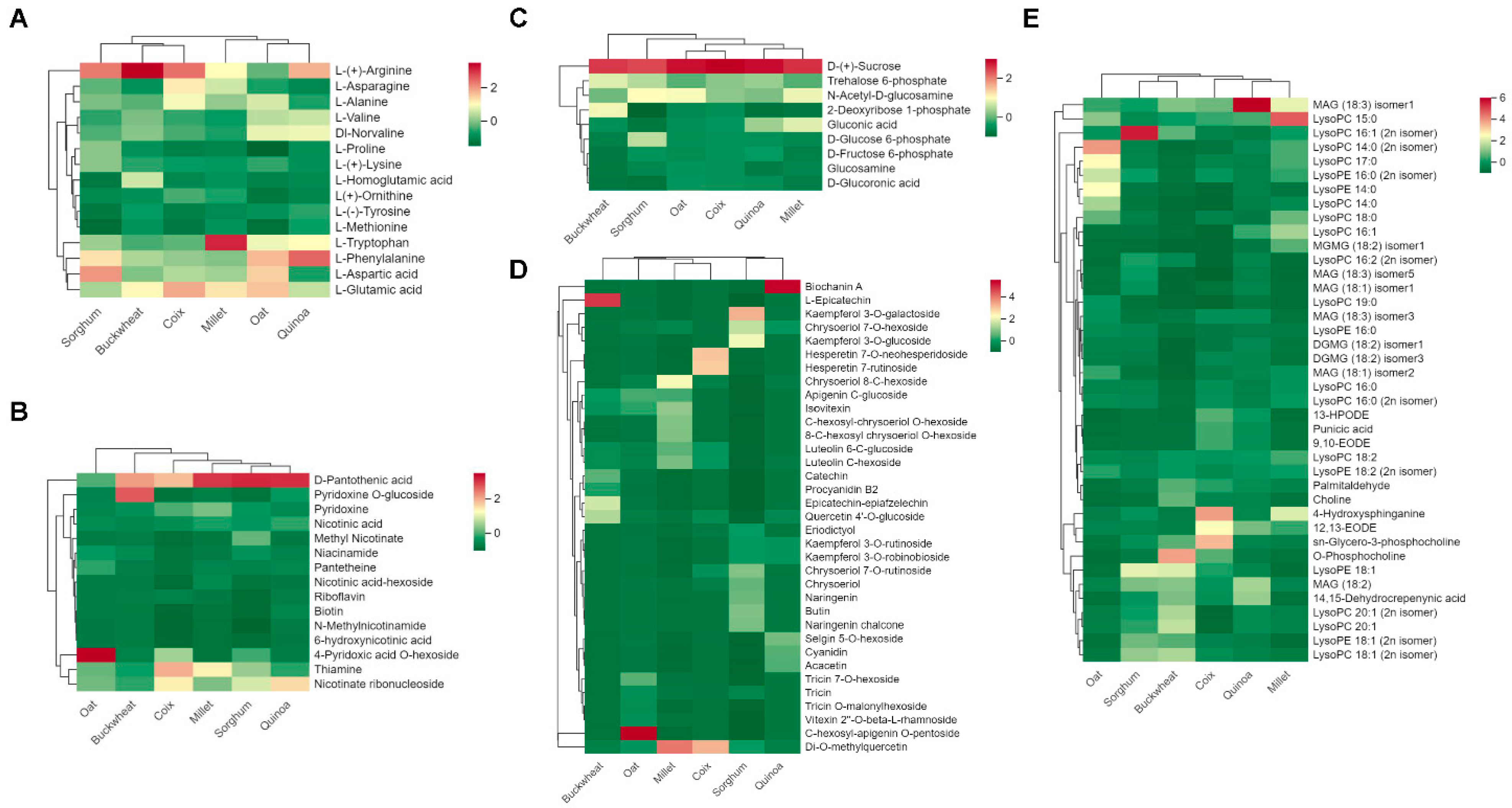
3.2. Comparative Analysis of Main Nutritional Metabolites in Six Coarse Cereals
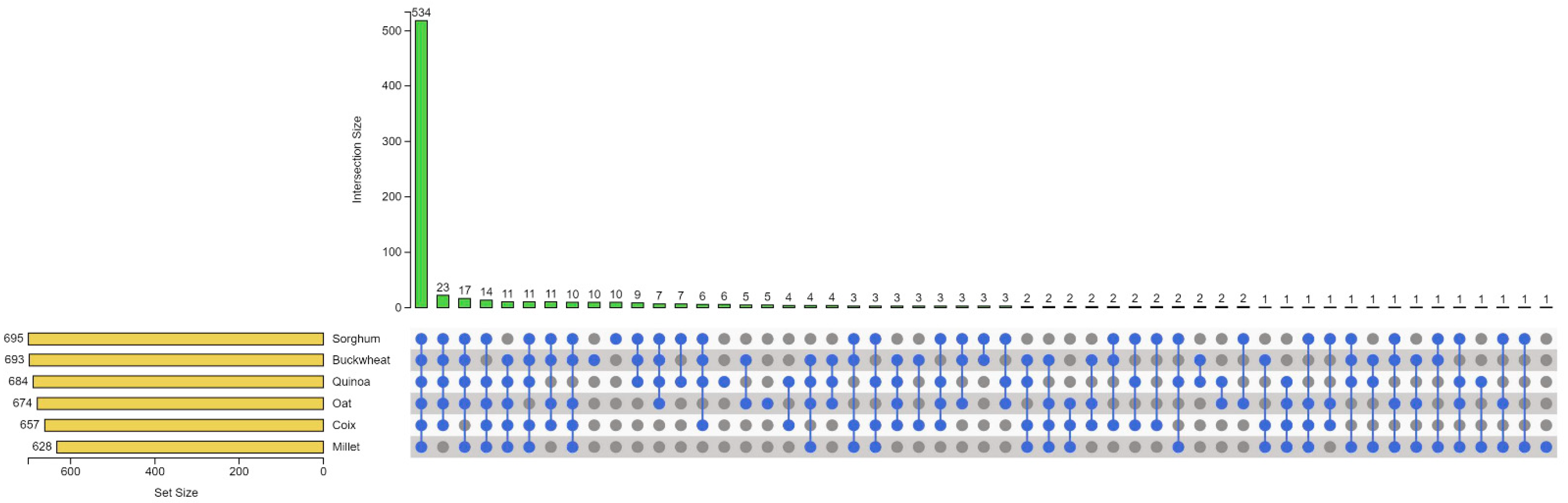
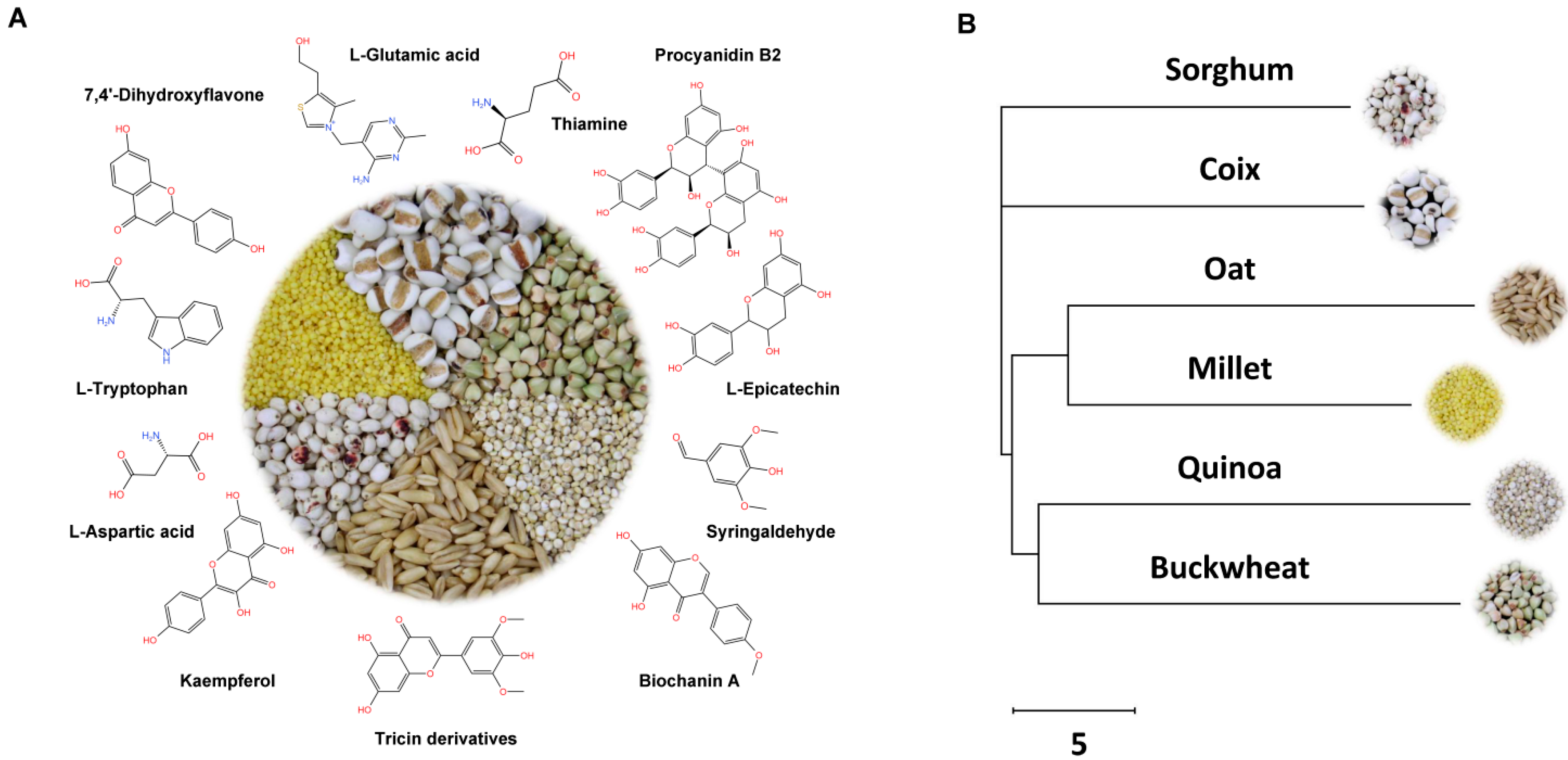
3.3. Analysis of Species-Specific Metabolites and Evolutionary Relationships of Six Coarse Cereals
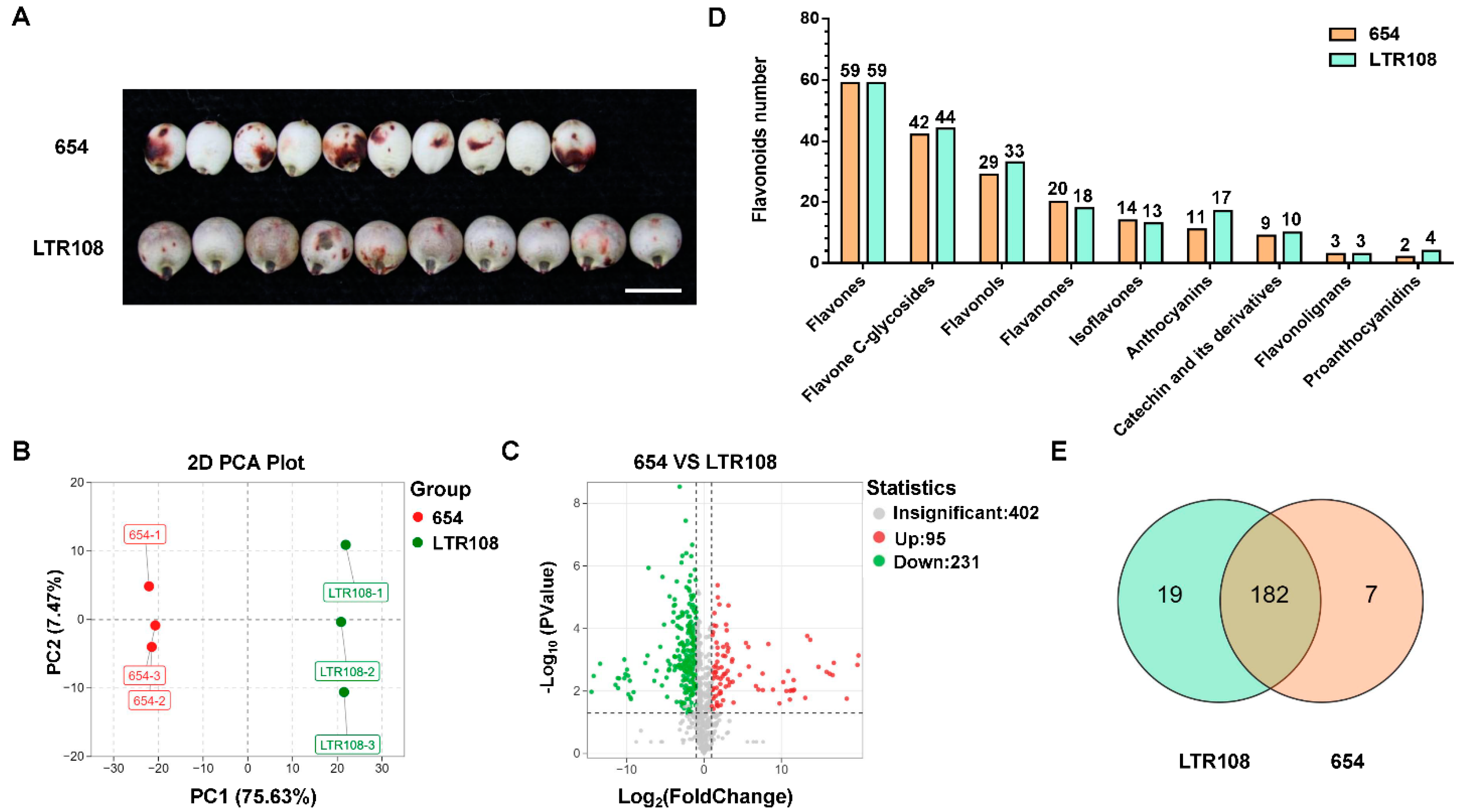
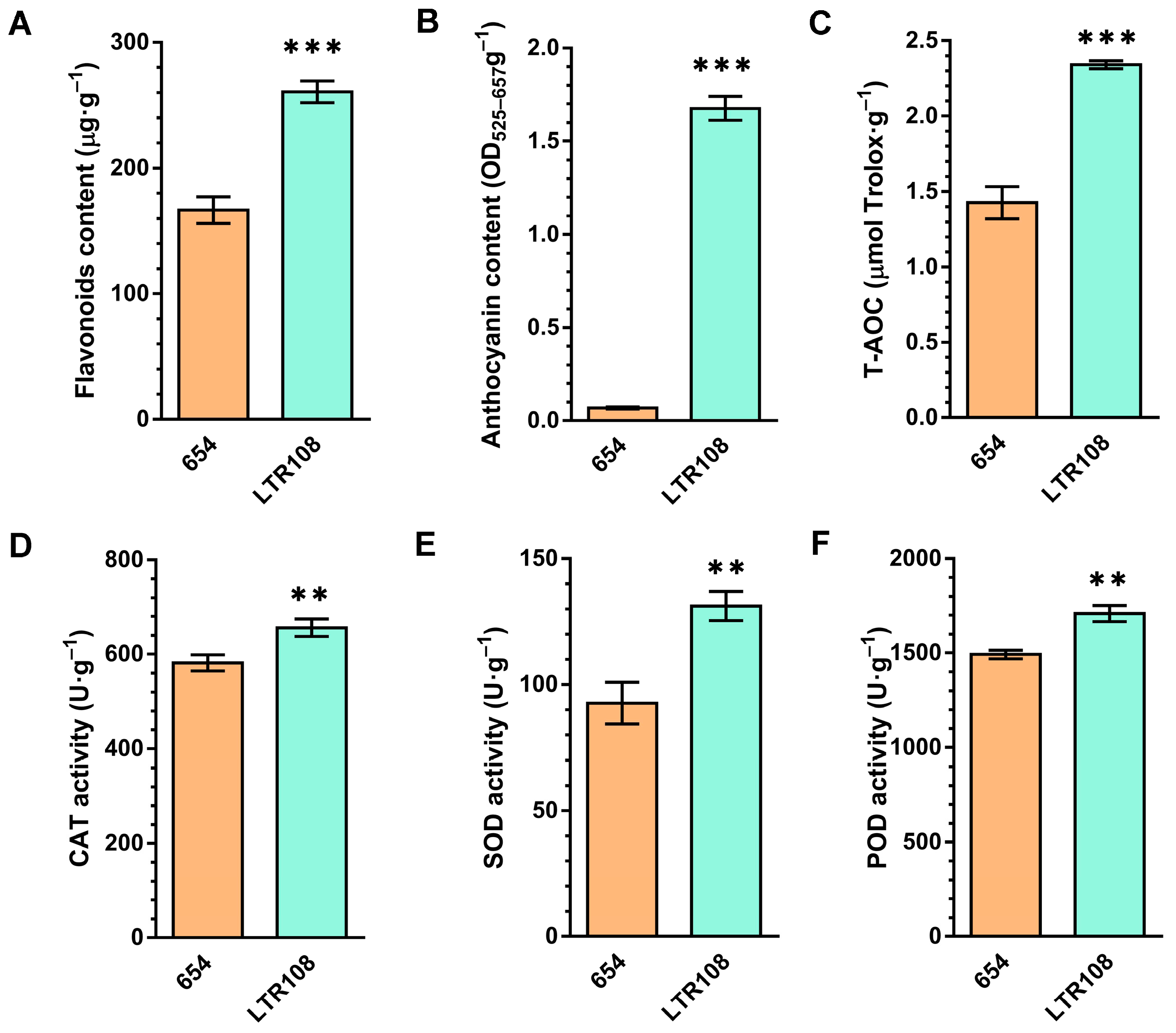
3.4. Analysis of Differential Metabolites and Antioxidant System Activities in Grain Sorghum and Sweet Sorghum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bassolino, L.; Petroni, K.; Polito, A.; Marinelli, A.; Azzini, E.; Ferrari, M.; Ficco, D.; Mazzucotelli, E.; Tondelli, A.; Fricano, A.; et al. Does plant breeding for antioxidant-rich foods have an impact on human health? Antioxidants 2022, 11, 794. [Google Scholar] [CrossRef] [PubMed]
- Rai, K.N.; Gowda, C.L.L.; Reddy, B.V.S.; Sehgal, S. Adaptation and potential uses of sorghum and pearl millet in alternative and health foods. Compr. Rev. Food Sci. Food Saf. 2009, 7, 340–352. [Google Scholar]
- Singh, A.K.; Singh, R.; Subramani, R.; Kumar, R.; Wankhede, D.P. Molecular approaches to understand nutritional potential of coarse cereals. Curr. Genom. 2016, 17, 177–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.; Luo, H.; Xu, J.; Cruickshank, A.; Zhao, X.; Teng, F.; Hathorn, A.; Wu, X.; Liu, Y.; Shatte, T.; et al. Extensive variation within the pan-genome of cultivated and wild sorghum. Nat. Plants 2021, 7, 766–773. [Google Scholar] [CrossRef]
- Punia, H.; Tokas, J.; Malik, A.; Sangwan, S.; Rani, A.; Yashveer, S.; Alansi, S.; Hashim, M.J.; El-Sheikh, M.A. Genome-wide transcriptome profiling, characterization, and functional identification of NAC transcription factors in sorghum under salt stress. Antioxidants 2021, 10, 1605. [Google Scholar] [CrossRef]
- Ritter, K.B.; McIntyre, C.L.; Godwin, I.D.; Jordan, D.R.; Chapman, S.C. An assessment of the genetic relationship between sweet and grain sorghums, within Sorghum bicolor ssp. bicolor (L.) Moench, using AFLP markers. Euphytica 2007, 157, 161–176. [Google Scholar]
- Kaur, K.D.; Jha, A.; Sabikhi, L.; Singh, A.K. Significance of coarse cereals in health and nutrition: A review. J. Food Sci. Technol. 2014, 51, 1429–1441. [Google Scholar] [CrossRef] [Green Version]
- Lie, L.; Brown, L.; Forrester, T.E.; Plange-Rhule, J.; Bovet, P.; Lambert, E.V.; Layden, B.T.; Luke, A.; Dugas, L.R. The association of dietary fiber intake with cardiometabolic risk in four countries across the epidemiologic transition. Nutrients 2018, 10, 628. [Google Scholar] [CrossRef] [Green Version]
- Ren, G.; Fan, X.; Teng, C.; Li, Y.; Everaert, N.; Blecker, C. The beneficial effect of coarse cereals on chronic diseases through regulating gut microbiota. Foods 2021, 10, 2891. [Google Scholar] [CrossRef]
- Hassan, Z.M.; Sebola, N.A.; Mabelebele, M. The nutritional use of millet grain for food and feed: A review. Agric. Food Secur. 2021, 10, 16. [Google Scholar] [CrossRef]
- Fan, Z. Coix: Chemical composition and health effects. Trends Food Sci. Technol. 2017, 61, 160–175. [Google Scholar]
- Sang, S.; Chu, Y. Whole grain oats, more than just a fiber: Role of unique phytochemicals. Mol. Nutr. Food Res. 2017, 61, 1600715. [Google Scholar] [CrossRef] [PubMed]
- Noreen, S.; Rizwan, B.; Khan, M.; Farooq, S. Health benefits of buckwheat (Fagopyrum esculentum), potential remedy for diseases, rare to cancer: A mini review. Infect. Disord. Drug Targets 2021, 21, e170721189478. [Google Scholar] [CrossRef] [PubMed]
- Graf, B.L.; Rojas-Silva, P.; Rojo, L.E.; Delatorre-Herrera, J.; Baldeón, M.E.; Raskin, I. Innovations in health value and functional food development of quinoa (Chenopodium quinoa Wild.). Compr. Rev. Food Sci. Food Saf. 2015, 14, 431–445. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Zhang, P.; Warner, R.D.; Fang, Z. Sorghum grain: From genotype, nutrition, and phenolic profile to its health benefits and food applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 2025–2046. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Zhang, Y.; Hu, Y.; Zhao, G.; Tang, Y.; Zou, L. Concise review: Coarse cereals exert multiple beneficial effects on human health. Food Chem. 2020, 325, 126761. [Google Scholar] [CrossRef]
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crozier, A.; Del Rio, D.; Clifford, M.N. Bioavailability of dietary flavonoids and phenolic compounds. Mol. Aspects Med. 2010, 31, 446–467. [Google Scholar] [CrossRef] [PubMed]
- Selma, M.V.; Espín, J.C.; Tomás-Barberán, F.A. Interaction between phenolics and gut microbiota: Role in human health. J. Agric. Food Chem. 2009, 57, 6485–6501. [Google Scholar] [CrossRef]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of intestinal microbiota in the bioavailability and physiological functions of dietary polyphenols. Molecules 2019, 24, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, D.; Tran, Y.; Shekhawat, G.S.; Gopinath, B. Dietary flavonoid intake and chronic sensory conditions: A scoping review. Antioxidants 2022, 11, 1214. [Google Scholar] [CrossRef] [PubMed]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K.; et al. MassBank: A public repository for sharing mass spectral data for life sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.; O’Maille, G.; Want, E.; Qin, C.; Trauger, S.; Brandon, T.; Custodio, D.; Abagyan, R.; Siuzdak, G. METLIN: A metabolite mass spectral database. Ther. Drug Monit. 2005, 27, 747–751. [Google Scholar] [CrossRef]
- Siddhu, N.; Saxena, J. Quantification of total phenolic and total flavonoid content of extracts of tagetes erecta flowers. Asian J. Pharm. Clin. Res. 2017, 10, 328–330. [Google Scholar]
- He, Y.; Zhang, X.; Li, L.; Sun, Z.; Li, J.; Chen, X.; Hong, G. SPX4 interacts with both PHR1 and PAP1 to regulate critical steps in phosphorus-status-dependent anthocyanin biosynthesis. New Phytol. 2021, 230, 205–217. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Kelly, B.; Pearce, E.L. Amino assets: How amino acids support immunity. Cell Metab. 2020, 32, 154–175. [Google Scholar] [CrossRef]
- Leese, H.J.; McKeegan, P.J.; Sturmey, R.G. Amino acids and the early mammalian embryo: Origin, fate, function and life-long legacy. Int. J. Environ. Res. Public Health 2021, 18, 9874. [Google Scholar] [CrossRef]
- Tardy, A.L.; Pouteau, E.; Marquez, D.; Yilmaz, C.; Scholey, A. Vitamins and minerals for energy, fatigue and cognition: A narrative review of the biochemical and clinical evidence. Nutrients 2020, 12, 228. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Ma, X.; Liu, X. Research progress on the functions of vitamins in body. J. Chin. Pharm. Sci. 2016, 25, 329–341. [Google Scholar]
- Butler, L.M.; Perone, Y.; Dehairs, J.; Lupien, L.E.; de Laat, V.; Talebi, A.; Loda, M.; Kinlaw, W.B.; Swinnen, J.V. Lipids and cancer: Emerging roles in pathogenesis, diagnosis and therapeutic intervention. Adv. Drug Deliv. Rev. 2020, 159, 245–293. [Google Scholar] [CrossRef]
- Cynober, L. Metabolism of dietary glutamate in adults. Ann. Nutr. Metab. 2018, 73, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Al-Nasser, M.N.; Mellor, I.R.; Carter, W.G. Is L-glutamate toxic to neurons and thereby contributes to neuronal loss and neurodegeneration? A Systematic Review. Brain Sci. 2022, 12, 577. [Google Scholar] [CrossRef] [PubMed]
- Marrs, C.; Lonsdale, D. Hiding in plain sight: Modern thiamine deficiency. Cells 2021, 10, 2595. [Google Scholar] [CrossRef] [PubMed]
- Alsharairi, N.A. Insights into the mechanisms of action of proanthocyanidins and anthocyanins in the treatment of nicotine-induced non-small cell lung cancer. Int. J. Mol. Sci. 2022, 23, 7905. [Google Scholar] [CrossRef]
- Alam, M.; Ali, S.; Ashraf, G.M.; Bilgrami, A.L.; Yadav, D.K.; Hassan, M.I. Epigallocatechin 3-gallate: From green tea to cancer therapeutics. Food Chem. 2022, 379, 132135. [Google Scholar] [CrossRef]
- Wu, J.; Fu, Y.S.; Lin, K.; Huang, X.; Chen, Y.J.; Lai, D.; Kang, N.; Huang, L.; Weng, C.F. A narrative review: The pharmaceutical evolution of phenolic syringaldehyde. Biomed. Pharmacother. 2022, 153, 113339. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Qiu, P.; Zhang, X.; Zhang, Y.; Mi, L.; Peng, C.; Pan, X.; Peng, F. Biochanin A from chinese medicine: An isoflavone with diverse pharmacological properties. Am. J. Chin. Med. 2021, 49, 1623–1643. [Google Scholar] [CrossRef]
- Jiang, B.; Song, J.; Jin, Y. A flavonoid monomer tricin in Gramineous plants: Metabolism, bio/chemosynthesis, biological properties, and toxicology. Food Chem. 2020, 320, 126617. [Google Scholar] [CrossRef]
- Amjad, E.; Sokouti, B.; Asnaashari, S. A systematic review of anti-cancer roles and mechanisms of kaempferol as a natural compound. Cancer Cell Int. 2022, 22, 260. [Google Scholar] [CrossRef]
- Park, H.; Jin, U.H.; Martin, G.; Chapkin, R.S.; Davidson, L.A.; Lee, K.; Jayaraman, A.; Safe, S. Structure-activity relationships among mono- and dihydroxy flavones as aryl hydrocarbon receptor (AhR) agonists or antagonists in Caco2 cells. Chem. Biol. Interact. 2022, 365, 110067. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Mu, C.L.; Farzi, A.; Zhu, W.Y. Tryptophan metabolism: A link between the gut microbiota and brain. Adv. Nutr. 2020, 11, 709–723. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Li, K.; Shi, Y.; Li, Y.; Dong, L.; Liu, L.; Li, M.; Ren, H.; Liu, X.; Fang, C.; et al. Cross-species comparison of metabolomics to decipher the metabolic diversity in ten fruits. Metabolites 2021, 11, 164. [Google Scholar] [CrossRef] [PubMed]
- Ain, N.U.; Haider, F.U.; Fatima, M.; Habiba; Zhou, Y.; Ming, R. Genetic determinants of biomass in C4 crops: Molecular and agronomic approaches to increase biomass for biofuels. Front. Plant Sci. 2022, 13, 839588. [Google Scholar]
- Zou, G.; Zhai, G.; Feng, Q.; Yan, S.; Wang, A.; Zhao, Q.; Shao, J.; Zhang, Z.; Zou, J.; Han, B.; et al. Identification of QTLs for eight agronomically important traits using an ultra-high-density map based on SNPs generated from high-throughput sequencing in sorghum under contrasting photoperiods. J. Exp. Bot. 2012, 63, 5451–5462. [Google Scholar] [CrossRef] [Green Version]
- Zou, G.; Zhai, G.; Yan, S.; Li, S.; Zhou, L.; Ding, Y.; Liu, H.; Zhang, Z.; Zou, J.; Zhang, L.; et al. Sorghum qTGW1a encodes a G-protein subunit and acts as a negative regulator of grain size. J. Exp. Bot. 2020, 71, 5389–5401. [Google Scholar] [CrossRef]
- Zhang, L.; Cui, D.; Ma, X.; Han, B.; Han, L. Comparative analysis of rice reveals insights into the mechanism of colored rice via widely targeted metabolomics. Food Chem. 2022, 399, 133926. [Google Scholar] [CrossRef]
- Khakimov, B.; Jespersen, B.M.; Engelsen, S.B. Comprehensive and comparative metabolomic profiling of wheat, barley, oat and rye using gas chromatography-mass spectrometry and advanced chemometrics. Foods 2014, 3, 569–585. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Li, X.; Zhang, Y.; Yang, Y.; Sun, R.; Li, Y.; Gao, J.; Han, Y. Differential flavonoids and carotenoids profiles in grains of six poaceae crops. Foods 2022, 11, 2068. [Google Scholar] [CrossRef]
- Yan, S.; Li, S.; Zhai, G.; Lu, P.; Deng, H.; Zhu, S.; Huang, R.; Shao, J.; Tao, Y.; Zou, G. Molecular cloning and expression analysis of duplicated polyphenol oxidase genes reveal their functional differentiations in sorghum. Plant Sci. 2017, 263, 23–30. [Google Scholar] [CrossRef]
- Shen, Y.; Jin, L.; Xiao, P.; Lu, Y.; Bao, J. Total phenolics, flavonoids, antioxidant capacity in rice grain and their relations to grain color, size and weight. J. Cereal Sci. 2009, 49, 106–111. [Google Scholar] [CrossRef]
- Dong, N.; Sun, Y.; Guo, T.; Shi, C.; Zhang, Y.; Kan, Y.; Xiang, Y.; Zhang, H.; Yang, Y.; Li, Y.; et al. UDP-glucosyltransferase regulates grain size and abiotic stress tolerance associated with metabolic flux redirection in rice. Nat. Commun. 2020, 11, 2629. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | ID | Q1 (Da) | RT (min) | Compounds | Class |
|---|---|---|---|---|---|
| Sorghum | pme0196 | 285 | 5.73 | Kaempferol | Flavonol |
| pme3288 | 329 | 6.63 | 3,7-Di-O-methylquercetin | Flavonol | |
| pme3300 | 301 | 4.51 | Tricetin | Flavone | |
| pme1496 | 267.1 | 6.33 | 4′-O-methyldaidzein | Isoflavone | |
| pme3502 | 429.1 | 4.59 | Formononetin 7-O-glucoside | Isoflavone | |
| pmb0631 | 581.1 | 3.17 | C-hexosyl-luteolin C-pentoside | Flavone C-glycosides | |
| pmb4777 | 337.1 | 4.4 | 4-Hydroxy-7-methoxycoumarin-beta-rhamnoside | Coumarins | |
| pmb2601 | 307.1 | 307.1 | 7-hydroxycoumarin-beta-rhamnoside | Coumarins | |
| pme0062 | 515.3 | 5.88 | Cucurbitacin D | Terpenoids | |
| pmb2833 | 529 | 2.13 | 3-O-Feruloyl quinic acid glucoside | Quinate derivatives | |
| Quinoa | pmb2835 | 181.1 | 3.95 | Syringaldehyde | Hydroxycinnamoyl derivatives |
| pme3468 | 739.2 | 3.35 | Kaempferol-3-O-robinoside-7-O-rhamnoside | Flavonol | |
| pme3399 | 445.1 | 5.16 | Sissotrin | Isoflavone | |
| pme1773 | 595 | 2.7 | Cyanidin 3-O-rutinoside | Anthocyanins | |
| pmb0542 | 535.1 | 2.99 | Cyanidin 3-O-malonylhexoside | Anthocyanins | |
| pmb3061 | 499.1 | 2.1 | 5-O-p-coumaroyl quinic acid O-hexoside | Quinate derivatives | |
| Millet | pme3507 | 253.1 | 4.75 | 7,4′-Dihydroxyflavone | Flavone |
| Oat | pme0434 | 653.1 | 4.16 | Tricin di-O-hexoside | Flavone |
| pmb3053 | 523.1 | 6 | Tricin O-eudesmic acid | Flavone | |
| pmb3031 | 403.1 | 5.23 | Tricin O-glycerol | Flavone | |
| pmb2979 | 549.2 | 3.91 | Hesperetin O-malonylhexoside | Flavone | |
| pmb3052 | 525.1 | 5.84 | Tricin 4′-O-β-guaiacylglycerol | Flavonolignan | |
| Buckwheat | pme0434 | 577 | 3.03 | Procyanidin B2 | Proanthocyanidins |
| pmb2976 | 755.1 | 3.74 | Chrysoeriol C-pentosyl-O-hexosyl-O-hexoside | Flavone C-glycosides | |
| pmb2977 | 739.2 | 3.87 | Chrysoeriol 8-C-pentosyl-O-rutinoside | Flavone C-glycosides | |
| pmb3047 | 657.1 | 4.50 | Tricin 4′-O-(syringyl alcohol) ether 5-O-hexoside | Flavonolignan | |
| pmb3066 | 481.1 | 2.62 | 5-O-p-coumaroyl shikimic acid O-hexoside | Quinate derivatives | |
| pmb3064 | 499.2 | 2.50 | 3-O-p-coumaroyl quinic acid O-hexoside | Quinate derivatives | |
| pmb3058 | 369 | 2.16 | Quinic acid O-glucuronic acid | Quinate derivatives | |
| pme1162 | 169 | 1.74 | Gallic acid | Benzoic acid derivatives | |
| pmb2947 | 865.1 | 3.44 | Catechin-catechin-catechin | Catechin derivatives | |
| pme3131 | 221.1 | 5.39 | 6,7-Dimethoxy-4-methylcoumarin | Coumarins |
| Species | ID | Q1 (Da) | RT (min) | Compounds | Class |
|---|---|---|---|---|---|
| 654 | pma0825 | 503 | 5.24 | Chrysin O-malonylhexoside | Flavone |
| pmb0744 | 435.3 | 5.48 | Tricin O-phenylformic acid | Flavone | |
| pma0787 | 551.1 | 4.63 | Quercetin-3-(6′-malonyl)-Glucoside | Flavonol | |
| pme2984 | 593 | 5.09 | Isosakuranetin-7-neohesperidoside | Flavanone | |
| pme3464 | 285.1 | 6.98 | Isosakuranetin | Flavanone | |
| pmb0631 | 445 | 3.54 | Glycitin | Isoflavone | |
| pmb2586 | 593.1 | 3.72 | Gallocatechin-catechin | Catechin derivatives | |
| LTR108 | pme1773 | 595 | 2.7 | Cyanidin 3-O-rutinoside | Anthocyanins |
| pmb0542 | 535.1 | 2.99 | Cyanidin 3-O-malonylhexoside | Anthocyanins | |
| pmb0557 | 621.1 | 3.26 | Cyanidin O-malonyl-malonylhexoside | Anthocyanins | |
| pme0094 | 447.3 | 2.61 | Cyanidin 3-O-glucoside | Anthocyanins | |
| pmb2959 | 489.1 | 2.94 | Cyanidin O-acetylhexoside | Anthocyanins | |
| pmb0558 | 637.1 | 3.27 | Delphinidin O-malonyl-malonylhexoside | Anthocyanins | |
| pmb0837 | 577.1 | 2.92 | Procyanidin A3 | Proanthocyanidins | |
| pme0434 | 577 | 3.03 | Procyanidin B2 | Proanthocyanidins | |
| pmb3044 | 653.1 | 4.16 | Tricin di-O-hexoside | Flavone | |
| pmb2999 | 461.1 | 3.87 | Chrysoeriol 5-O-hexoside | Flavone | |
| pmb3013 | 519.1 | 4.32 | Isorhamnetin O-acetyl-hexoside | Flavonol | |
| pme1588 | 315.1 | 5.85 | Isorhamnetin | Flavonol | |
| pme1478 | 317 | 4.7 | Myricetin | Flavonol | |
| pmb3026 | 505.1 | 3.8 | Quercetin O-acetylhexoside | Flavonol | |
| pmb0615 | 789.1 | 2.58 | Hesperetin C-hexosyl-O-hexosyl-O-hexoside | Flavone C-glycosides | |
| pmb0691 | 757.1 | 3.09 | Luteolin C-hexosyl-O-rhamnoside O-hexoside | Flavone C-glycosides | |
| pme0450 | 289 | 3.32 | L-Epicatechin | Catechin derivatives | |
| pmb2947 | 865.1 | 3.44 | Catechin-catechin-catechin | Catechin derivatives |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Zhai, G.; Li, X.; Tao, H.; Li, L.; He, Y.; Zhang, X.; Wang, F.; Hong, G.; Zhu, Y. Metabolomics Reveals Nutritional Diversity among Six Coarse Cereals and Antioxidant Activity Analysis of Grain Sorghum and Sweet Sorghum. Antioxidants 2022, 11, 1984. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11101984
Zhao Y, Zhai G, Li X, Tao H, Li L, He Y, Zhang X, Wang F, Hong G, Zhu Y. Metabolomics Reveals Nutritional Diversity among Six Coarse Cereals and Antioxidant Activity Analysis of Grain Sorghum and Sweet Sorghum. Antioxidants. 2022; 11(10):1984. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11101984
Chicago/Turabian StyleZhao, Yao, Guowei Zhai, Xuetong Li, Han Tao, Linying Li, Yuqing He, Xueying Zhang, Fulin Wang, Gaojie Hong, and Ying Zhu. 2022. "Metabolomics Reveals Nutritional Diversity among Six Coarse Cereals and Antioxidant Activity Analysis of Grain Sorghum and Sweet Sorghum" Antioxidants 11, no. 10: 1984. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11101984