
Opposite Effect of Thyroid Hormones on Oxidative Stress and on Mitochondrial Respiration in COVID-19 Patients
 , ,
, ,  , , , ,
, , , ,  ,
,  and
and
Abstract
:1. Introduction
2. Methods
2.1. Study Group
2.2. Thyroid Hormone Function Tests
2.3. Lethality
2.4. Measurement of the Reactive Oxygen Species (ROS) and of Biological Antioxidant Potential (BAP) in Serum of COVID-19 Patients with NTIS
2.5. Isolation of PBMC from COVID-19 Patients with NTIS
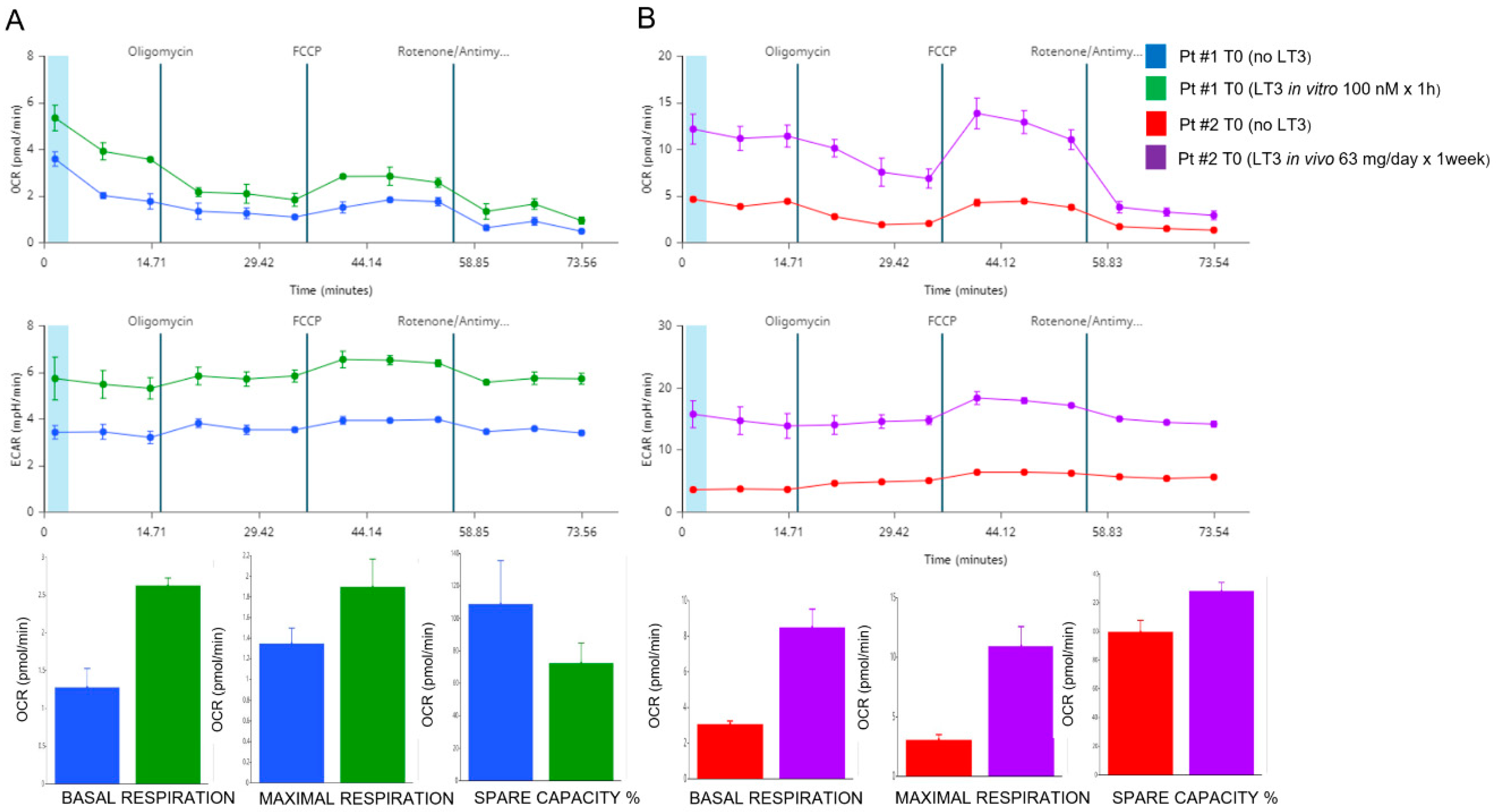
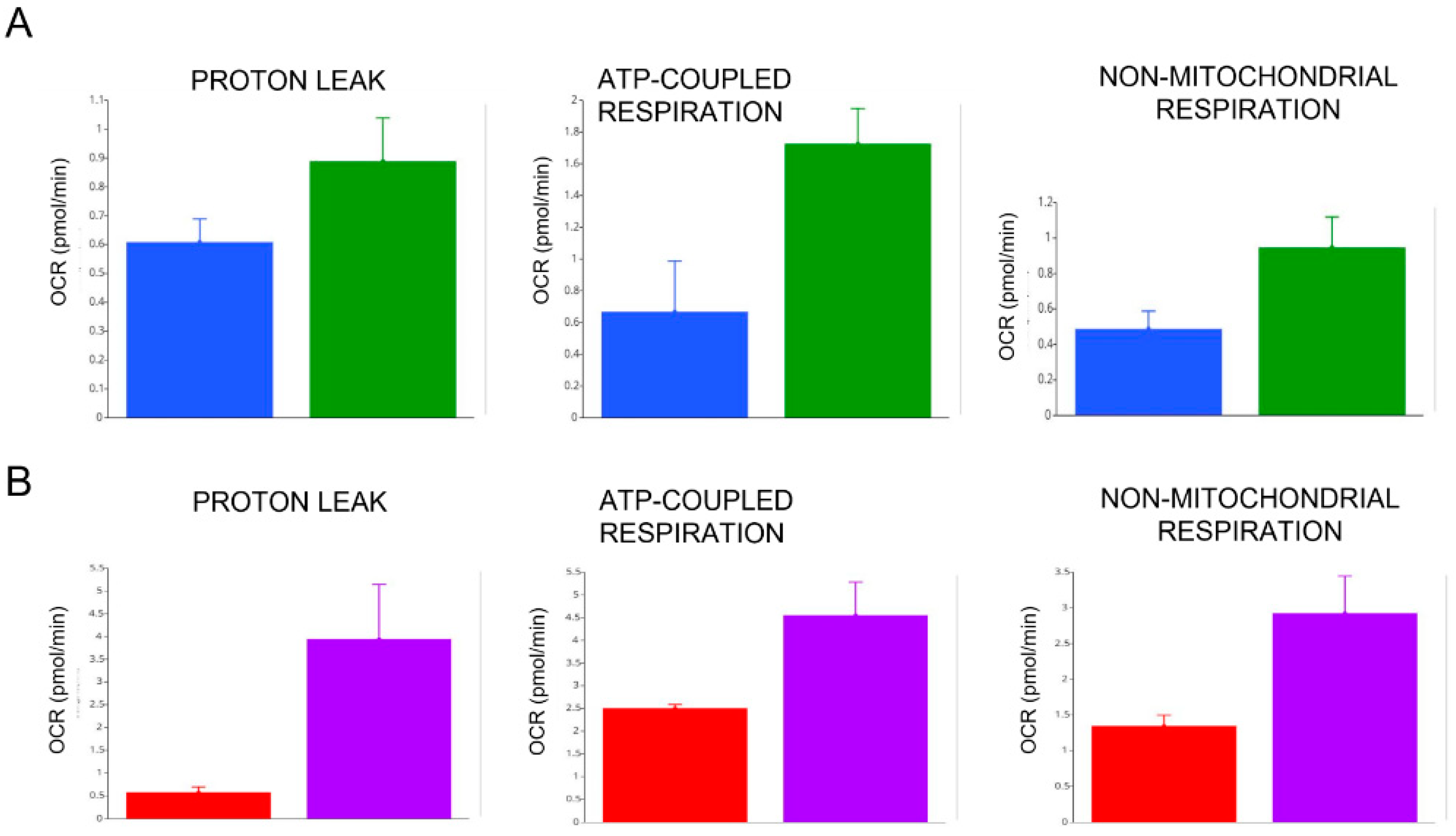
2.6. Assessment of Mitochondrial Respiration
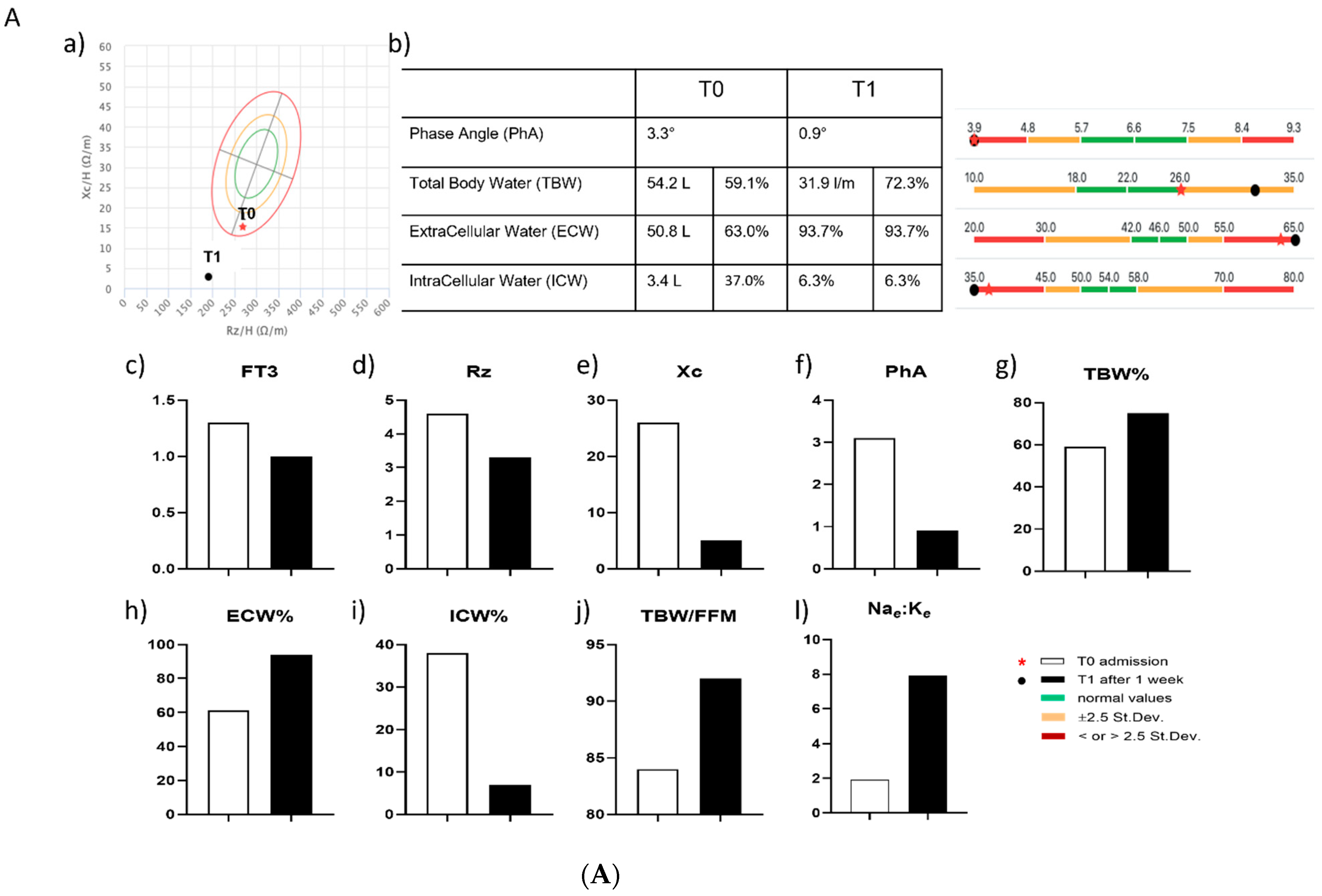
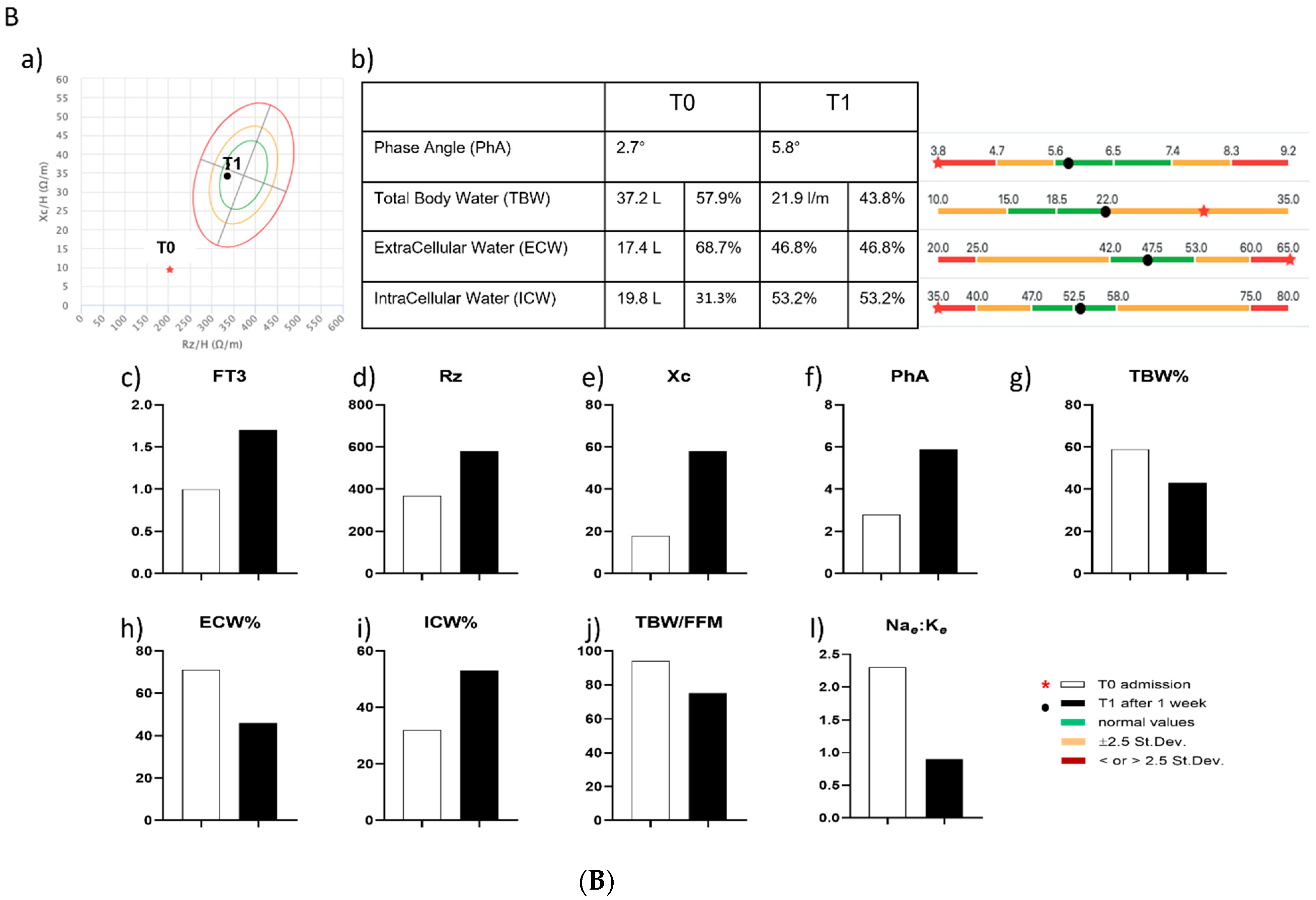
2.7. Assessment of BIA and of Mitochondrial Respiration in Two COVID-19 Patients with NTIS
2.8. Statistical Analysis
2.9. Ethics
3. Results
3.1. Lethality
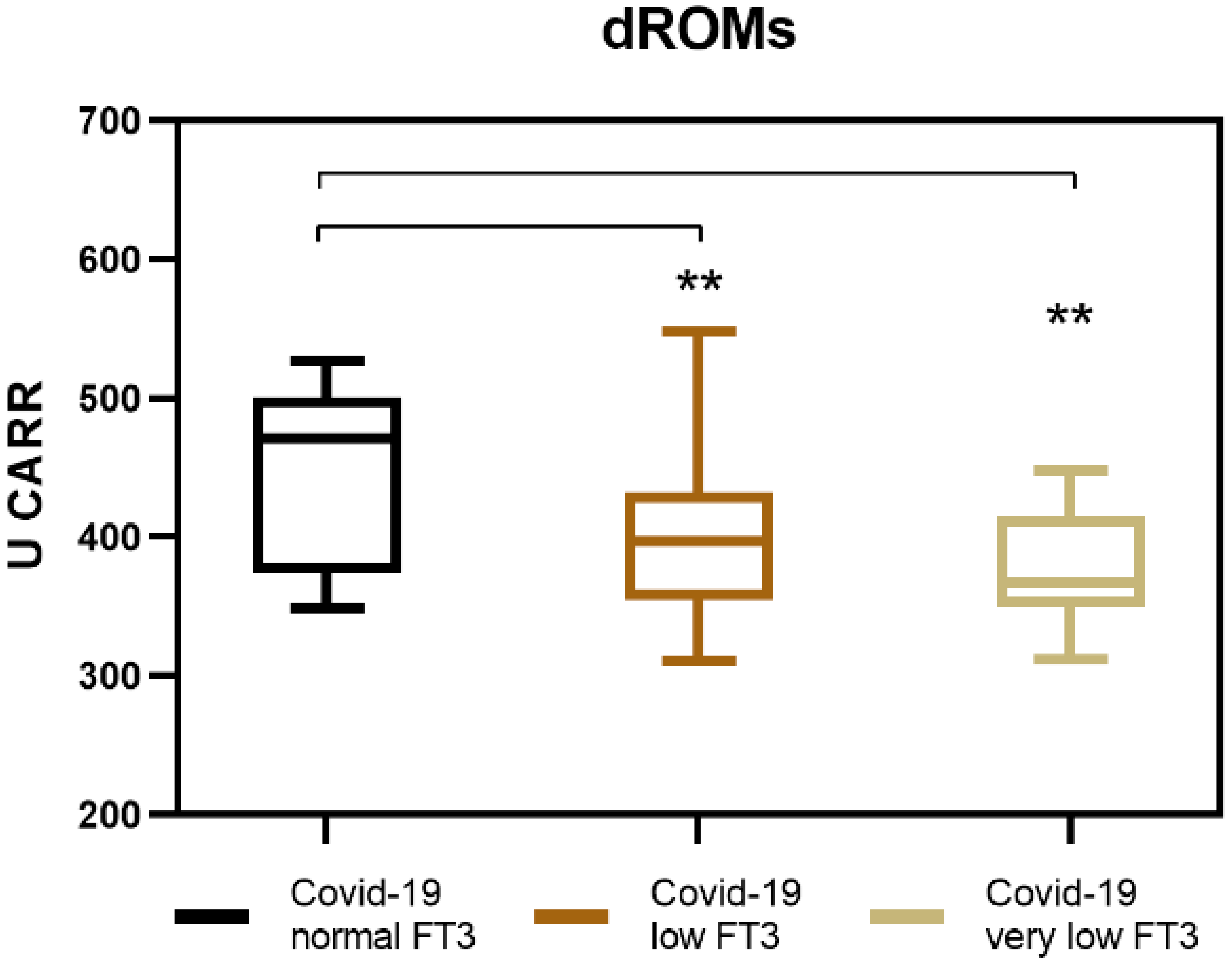
3.2. Generation of Reactive Oxygen Species (ROS) in COVID-19 Patients with NTIS
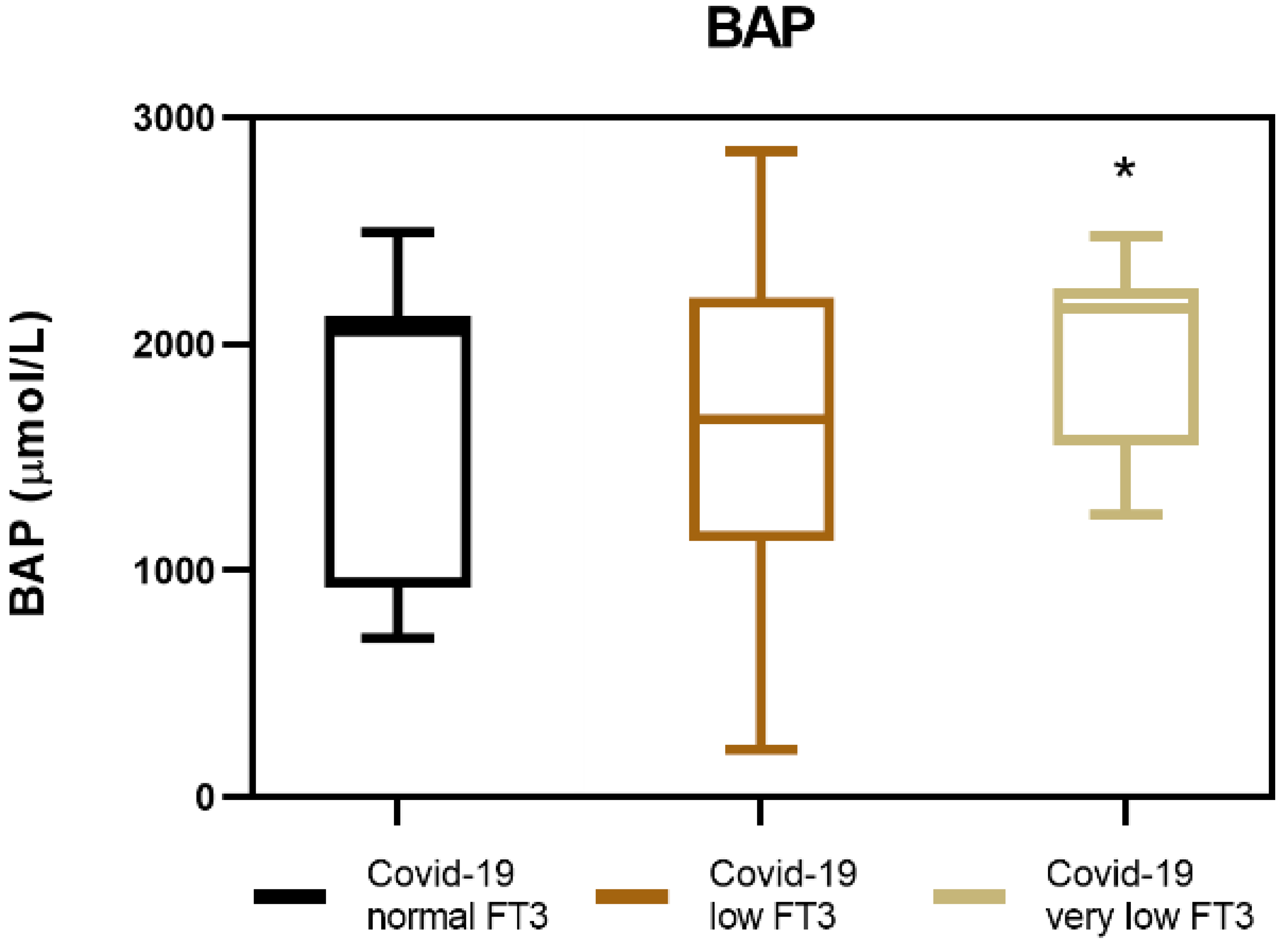
3.3. Biological Antioxidant Potential (BAP) in COVID-19 Patients with NTIS
3.4. Oxidative Stress Index (OSi) in COVID-19 Patients with NTIS
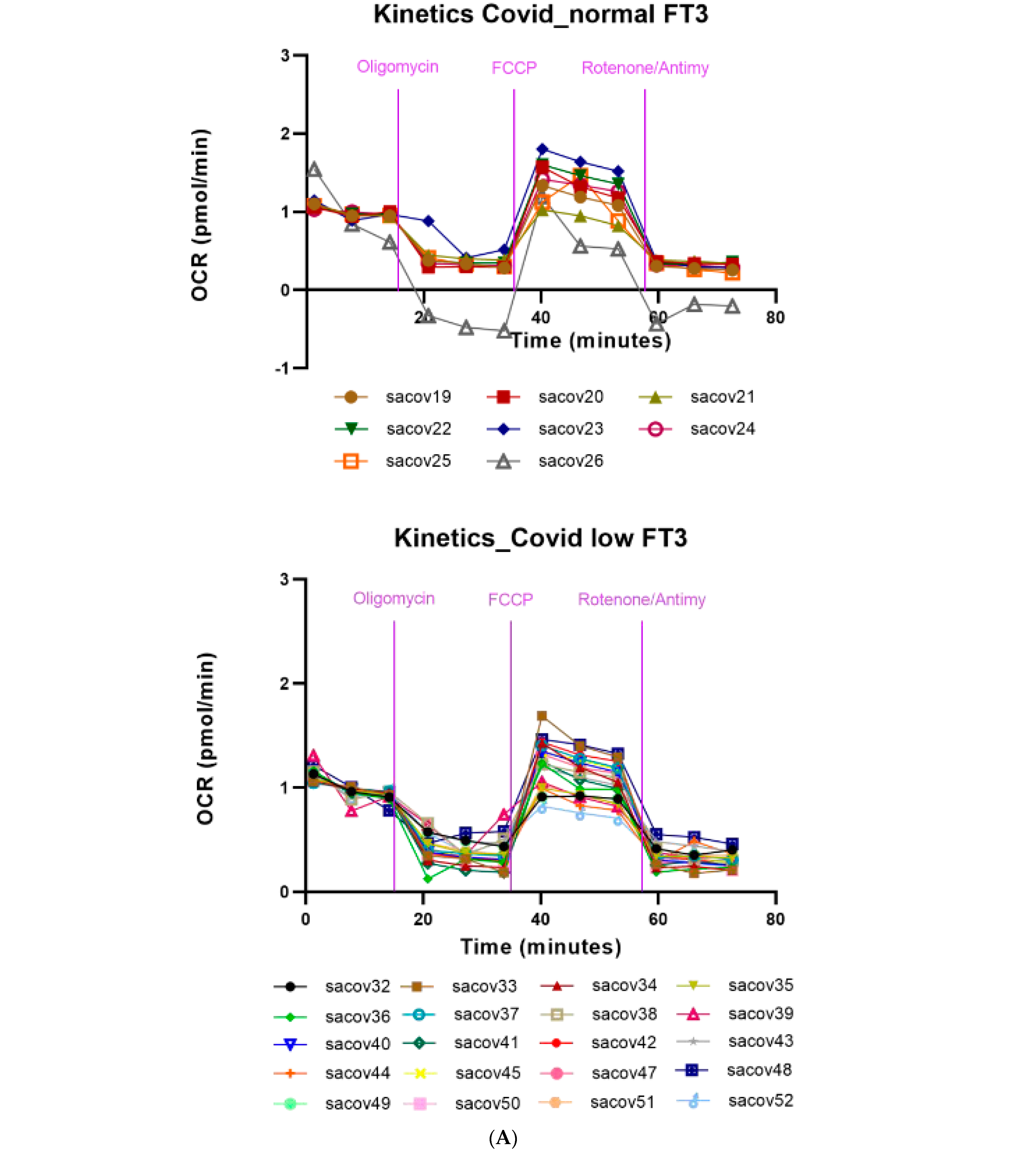
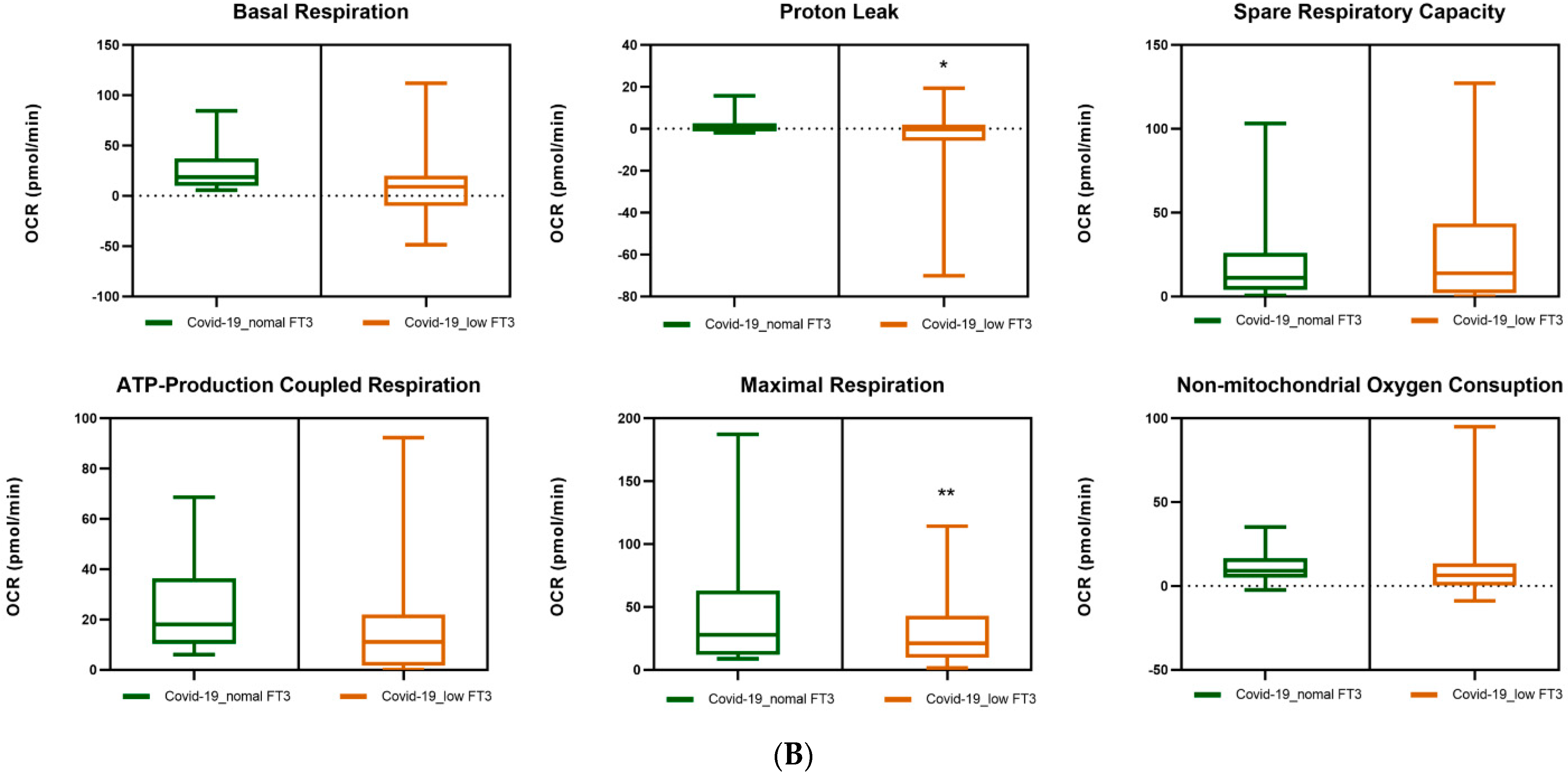
3.5. Mitochondrial Respiration in the PBMCs of COVID-19 Patients. Effects of FT3 Serum Levels
3.6. Effects of In Vitro and In Vivo LT3 Treatment in Two COVID-19 Patients with NTIS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Preiser, J.-C.; Ichai, C.; Orban, J.-C.; Groeneveld, A.B.J. Metabolic response to the stress of critical illness. BJA Br. J. Anaesth. 2014, 113, 945–954. [Google Scholar] [CrossRef] [Green Version]
- Chopra, I.J. Clinical review 86: Euthyroid sick syndrome: Is it a misnomer? J. Clin. Endocrinol. Metab. 1997, 82, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Bermudez, F.; Surks, M.I.; Oppenheimer, J.H. High incidence of decreased serum triiodothyronine concentration in patients with nonthyroidal disease. J. Clin. Endocrinol. Metab. 1975, 41, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.M.; Larsen, P.R.; Crantzf, R.; Dzau, V.J.; Rossing, T.H.; Haddow, J.E. Prevalence of abnormal thyroid function test results in patients with acute medical illnesses. Am. J. Med. 1982, 72, 9–16. [Google Scholar] [CrossRef]
- Plikat, K.; Langgartner, J.; Buettner, R.; Bollheimer, L.C.; Woenckhaus, U.; Schölmerich, J.; Wrede, C.E. Frequency and outcome of patients with nonthyroidal illness syndrome in a medical intensive care unit. Metab. Clin. Exp. 2007, 56, 239–244. [Google Scholar] [CrossRef]
- Langouche, L.; Jacobs, A.; Van den Berghe, G. Nonthyroidal Illness Syndrome Across the Ages. J. Endocr. Soc. 2019, 3, 2313–2325. [Google Scholar] [CrossRef] [Green Version]
- ICD-11, version 02/2022. Available online: https://icd.who.int/browse11/l-m/en (accessed on 31 July 2022).
- Chinga-Alayo, E.; Villena, J.; Evans, A.T.; Zimic, M. Thyroid hormone levels improve the prediction of mortality among patients admitted to the intensive care unit. Intensive Care Med. 2005, 31, 1356–1361. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, P.; Díez, J.J. Thyroid dysfunction and kidney disease. Eur. J. Endocrinol. 2009, 160, 503–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bello, G.; Pennisi, M.A.; Montini, L.; Silva, S.; Maviglia, R.; Cavallaro, F.; Bianchi, A.; De Marinis, L.; Antonelli, M. Nonthyroid illness syndrome and prolonged mechanical ventilation in patients admitted to the ICU. Chest 2009, 135, 1448–1454. [Google Scholar] [CrossRef] [Green Version]
- Sharshar, T.; Bastuji-Garin, S.; Polito, A.; De Jonghe, B.; Stevens, R.D.; Maxime, V.; Rodriguez, P.; Cerf, C.; Outin, H.; Touraine, P.; et al. Hormonal status in protracted critical illness and in-hospital mortality. Crit. Care 2011, 15, R47. [Google Scholar] [CrossRef]
- Peeters, R.P. Non thyroidal illness: To treat or not to treat? Annales d’Endocrinologie 2007, 68, 224–228. [Google Scholar] [CrossRef]
- Sciacchitano, S.; Capalbo, C.; Napoli, C.; Anibaldi, P.; Salvati, V.; De Vitis, C.; Mancini, R.; Coluzzi, F.; Rocco, M. Nonthyroidal Illness Syndrome: To Treat or Not to Treat? Have We Answered the Question? A Review of Metanalyses. Front. Endocrinol. 2022, 13, 850328. [Google Scholar] [CrossRef]
- Economidou, F.; Douka, E.; Tzanela, M.; Orfanos, S.; Kotanidou, A. Thyroid Function in Critical Illness. In Diet and Nutrition in Critical Care; Rajendram, R., Preedy, V.R., Patel, V.B., Eds.; Springer: New York, NY, USA, 2015. [Google Scholar] [CrossRef]
- Jonklaas, J.; Bianco, A.C.; Bauer, A.J.; Burman, K.D.; Cappola, A.R.; Celi, F.S.; Cooper, D.S.; Kim, B.W.; Peeters, R.P.; Rosenthal, M.S.; et al. Guidelines for the Treatment of Hypothyroidism: Prepared by the american thyroid association task force on thyroid hormone replacement. Thyroid 2014, 24, 1670–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sciacchitano, S.; De Vitis, C.; D’Ascanio, M.; Giovagnoli, S.; De Dominicis, C.; Laghi, A.; Anibaldi, P.; Petrucca, A.; Salerno, G.; Santino, I.; et al. Gene signature and immune cell profiling by high-dimensional, single-cell analysis in COVID-19 patients, presenting Low T3 syndrome and coexistent hematological malignancies. J. Transl. Med. 2021, 19, 139. [Google Scholar] [CrossRef] [PubMed]
- Sciacchitano, S.; Capalbo, C.; Napoli, C.; Negro, A.; De Biase, L.; Marcolongo, A.; Anibaldi, P.; Salvati, V.; Petrella, L.; Merlo, L.; et al. Nonthyroidal illness syndrome (NTIS) in severe COVID-19 patients: Role of T3 on the Na/K pump gene expression and on hydroelectrolytic equilibrium. J. Transl. Med. 2021, 19, 491. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Guo, W.; Guo, Y.; Shi, M.; Dong, G.; Wang, G.; Ge, Q.; Zhu, J.; Zhou, X. Thyroid hormone concentrations in severely or critically ill patients with COVID-19. J. Endocrinol. Investig. 2020, 44, 1031–1040. [Google Scholar] [CrossRef] [PubMed]
- Chacko, B.K.; Kramer, P.A.; Ravi, S.; Benavides, G.A.; Mitchell, T.; Dranka, B.P.; Ferrick, D.; Singal, A.K.; Ballinger, S.W.; Bailey, S.M.; et al. The Bioenergetic Health Index: A new concept in mitochondrial translational research. Clin. Sci. 2014, 127, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Burtscher, J.; Cappellano, G.; Omori, A.; Koshiba, T.; Millet, G.P. Mitochondria: In the Cross Fire of SARS-CoV-2 and Immunity. iScience 2020, 23, 101631. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.K.; Chaubey, G.; Chen, J.Y.; and Suravajhala, P. Decoding SARS-CoV-2 hijacking of host mitochondria in COVID-19 pathogenesis. Am. J. Physiol. Cell Physiol. 2020, 319, C258–C267. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. An introduction to the roles of purinergic signalling in neurodegeneration, neuroprotection and neuroregeneration. Neuropharmacology 2016, 104, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Zolkipli-Cunningham, Z.; Naviaux, J.C.; Nakayama, T.; Hirsch, C.M.; Monk, J.M.; Li, K.; Wang, L.; Le, T.P.; Meinardi, S.; Blake, D.R.; et al. Metabolic and behavioral features of acute hyperpurinergia and the maternal immune activation mouse model of autism spectrum disorder. PLoS ONE 2021, 16, e0248771. [Google Scholar] [CrossRef] [PubMed]
- Pei, L.; Wallace, D.C. Mitochondrial etiology of neuropsychiatric disorders. Biol. Psychiatry 2018, 83, 722–730. [Google Scholar] [CrossRef] [PubMed]
- Mullur, R.; Liu, Y.Y.; Brent, G.A. Thyroid hormone regulation of metabolism. Physiol Rev. 2014, 94, 355–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capelli, V.; Diéguez, C.; Mittag, J.; López, M. Thyroid wars: The rise of central actions. Trends Endocrinol. Metab. 2021, 32, 659–671. [Google Scholar] [CrossRef]
- Burger, A.G.; Berger, M.; Wimpfheimer, K.; Danforth, E. Interrelationships between energy metabolism and thyroid hormone metabolism during starvation in the rat. Acta Endocrinol 1980, 93, 322–331. [Google Scholar] [CrossRef]
- Yau, W.W.; Yen, P.M. Thermogenesis in Adipose Tissue Activated by Thyroid Hormone. Int. J. Mol. Sci. 2020, 21, 3020. [Google Scholar] [CrossRef]
- van der Spek, A.H.; Fliers, E.; Boelen, A. Thyroid hormone metabolism in innate immune cells. J. Endocrinol. 2017, 232, R67–R81. [Google Scholar] [CrossRef] [Green Version]
- Mancini, A.; Di Segni, C.; Raimondo, S.; Olivieri, G.; Silvestrini, A.; Meucci, E.; Curro, D. Thyroid hormones, oxidative stress, and inflammation. Mediat. Inflamm 2016, 2016, 6757154. [Google Scholar] [CrossRef] [Green Version]
- Morimoto, M.; Satomura, S.; Hashimoto, T.; Ito, E.; Kyotani, S. Oxidative Stress Measurement and Prediction of Epileptic Seizure in Children and Adults with Severe Motor and Intellectual Disabilities. J. Clin. Med. Res. 2016, 8, 437–444. [Google Scholar] [CrossRef]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Melnikova, T.I.; Porozov, Y.P.; Terentiev, A.A. Oxidative stress and advanced lipoxidation and glycation end products (ALEs and AGEs) in aging and age-related diseases. Oxid. Med. Cell Longev. 2019, 14, 3085756. [Google Scholar] [CrossRef] [Green Version]
- Loffredo, L.; Violi, F. COVID-19 and cardiovascular injury: A role for oxidative stress and antioxidant treatment? Int. J. Cardiol. 2020, 312, 136. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov Identifier: NCT04466657. Available online: ClinicalTrials.gov (accessed on 31 July 2022).
- Bektas, A.; Schurman, S.H.; Franceschi, C.; Ferrucci, L. A public health perspective of aging: Do hyper-inflammatory syndromes such as COVID-19, SARS, ARDS, cytokine storm syndrome, and post-ICU syndrome accelerate short- and long-term inflammaging? Immun. Ageing 2020, 17, 23. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yu, Y.; Xu, J.; Shu, H.; Xia, J.; Liu, H.; Wu, Y.; Zhang, L.; Yu, Z.; Fang, M.; et al. Clinical course and outcomes of critically ill patients with SARS-CoV-2 pneumonia in Wuhan, China: A single-centered, retrospective, observational study. Lancet Respir. Med. 2020, 8, 475–481. [Google Scholar] [CrossRef] [Green Version]
- López-Lluch, G.; Hernández-Camacho, J.D.; Fernández-Ayala, D.J.M.; Navas, P. Mitochondrial dysfunction in metabolism and ageing: Shared mechanisms and outcomes? Biogerontology 2018, 19, 461–480. [Google Scholar] [CrossRef]
- Moreno Fernández-Ayala, D.J.; Navas, P.; López-Lluch, G. Age-related mitochondrial dysfunction as a key factor in COVID-19 disease. Exp. Gerontol. 2020, 142, 111147. [Google Scholar] [CrossRef]
- Kwok, S.; Adam, S.; Ho, J.H.; Iqbal, Z.; Turkington, P.; Razvi, S.; Le Roux, C.W.; Soran, H.; Syed, A.A. Obesity: A critical risk factor in the COVID-19 pandemic. Clin. Obes. 2020, 10, e12403. [Google Scholar] [CrossRef]
- Bansal, R.; Gubbi, S.; Muniyappa, R. Metabolic syndrome and COVID 19: Endocrine-immune-vascular interactions shapes clinical course. Endocrinology 2020, 161, bqaa112. [Google Scholar] [CrossRef]
- Prasun, P. Mitochondrial dysfunction in metabolic syndrome. Biochim. Biophys. Acta Mol. Dis. 2020, 1866, 165838. [Google Scholar] [CrossRef]
- Maddaloni, E.; Buzzetti, R. COVID-19 and diabetes mellitus: Unveiling the interaction of two pandemics. Diabetes Metab. Res. Rev. 2020, 36, e33213321. [Google Scholar] [CrossRef]
- McMaster, W.G.; Kirabo, A.; Madhur, M.S.; Harrison, D.G. Inflammation, immunity, and hypertensive end-organ damage. Circ. Res. 2015, 116, 1022–1033. [Google Scholar] [CrossRef] [Green Version]
- Nishiga, M.; Wang, D.W.; Han, Y.; Lewis, D.B.; Wu, J.C. COVID-19 and cardiovascular disease: From basic mechanisms to clinical perspectives. Nat. Rev. Cardiol. 2020, 17, 543–558. [Google Scholar] [CrossRef]
- Zaki, N.; Alashwal, H.; Ibrahim, S. Association of hypertension, diabetes, stroke, cancer, kidney disease, and high-cholesterol with COVID-19 disease severity and fatality: A systematic review. Diabetes Metab. Syndr. 2020, 14, 1133–1142. [Google Scholar] [CrossRef]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Ajaz, S.; McPhail, M.J.; Singh, K.K.; Mujib, S.; Trovato, F.M.; Napoli, S.; Agarwal, K. Mitochondrial metabolic manipulation by SARS-CoV-2 in peripheral blood mononuclear cells of patients with COVID-19. Am. J. Physiol. Cell Physiol. 2021, 320, C57–C65. [Google Scholar] [CrossRef] [PubMed]
- De Biasi, S.; Meschiari, M.; Gibellini, L.; Bellinazzi, C.; Borella, R.; Fidanza, L.; Gozzi, L.; Iannone, A.; Lo Tartaro, D.; Mattioli, M.; et al. Marked T cell activation, senescence, exhaustion and skewing towards TH17 in patients with COVID-19 pneumonia. Nat. Commun. 2020, 11, 3434. [Google Scholar] [CrossRef]
- Gibellini, L.; De Biasi, S.; Paolini, A.; Borella, R.; Boraldi, F.; Mattioli, M.; Lo Tartaro, D.; Fidanza, L.; Caro-Maldonado, A.; Meschiari, M.; et al. Altered bioenergetics and mitochondrial dysfunction of monocytes in patients with COVID-19 pneumonia. EMBO Mol. Med. 2020, 12, e13001. [Google Scholar] [CrossRef] [PubMed]
- Medini, H.; Zirman, A.; Mishmar, D. Immune system cells from COVID-19 patients display compromised mitochondrial-nuclear expression co-regulation and rewiring toward glycolysis. iScience 2021, 24, 103471. [Google Scholar] [CrossRef]
- Harper, M.E.; Seifert, E.L. Thyroid hormone effects on mitochondrial energetics. Thyroid 2008, 18, 145–156. [Google Scholar] [CrossRef]
- Sterling, K.; Campbell, G.A.; Taliadouros, G.S.; Nunez, E.A. Mitochondrial binding of triiodothyronine (T3). Demonstration by electron- microscopic radioautography of dispersed liver cells. Cell Tissue Res. 1984, 236, 321–325. [Google Scholar]
- Resch, U.; Helsel, G.; Tatzber, F.; Sinzinger, H. Antioxidant status in thyroid dysfunction. Clin. Chem. Lab. Med. 2002, 40, 1132–1134. [Google Scholar] [CrossRef] [PubMed]
- Gaggini, M.; Traghella, I.; Vassalle, C. The Thyroid-Oxidative Stress Axis in Heart Failure. In Thyroid Heart; Iervasi, G., Pingitore, A., Gerdes, A., Razvi, S., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 171–186. [Google Scholar]
- Liu, X.Y.; Chen, W.; Wei, B.; Shan, Y.F.; Wang, C. IFN-induced TPR protein IFIT3 potentiates antiviral signaling by bridging MAVS and TBK1. J. Immunol. 2011, 187, 2559–2568. [Google Scholar] [CrossRef] [Green Version]
- Schroder, K.; Tschopp, J. The inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latz, E. The inflammasomes: Mechanisms of activation and function. Curr. Opin. Immunol. 2010, 22, 28–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, R.; Yazdi, A.S.; Menu, P.; Tschopp, J. A role for mitochondria in NLRP3 inflammasome activation. Nature 2011, 469, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhang, D.; Hu, D.; Zhou, X.; Zhou, Y. The role of mitochondria in NLRP3 inflammasome activation. Mol. Immunol. 2018, 103, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Marlein, C.R.; Piddock, R.E.; Mistry, J.J.; Zaitseva, L.; Hellmich, C.; Horton, R.H.; Zhou, Z.; Auger, M.J.; Bowles, K.M.; Rushworth, S.A. CD38-Driven Mitochondrial Trafficking Promotes Bioenergetic Plasticity in Multiple Myeloma. Cancer Res. 2019, 79, 2285–2297. [Google Scholar] [CrossRef] [Green Version]
- Hayakawa, K.; Esposito, E.; Wang, X.; Terasaki, Y.; Liu, Y.; Xing, C.; Ji, X.; Lo, E.H. Transfer of mitochondria from astrocytes to neurons after stroke. Nature 2016, 535, 551–555, Erratum in Nature 2016, 539, 123. [Google Scholar] [CrossRef] [Green Version]
- Camacho-Pereira, J.; Tarragó, M.G.; Chini, C.C.S.; Nin, V.; Escande, C.; Warner, G.M.; Puranik, A.S.; Schoon, R.A.; Reid, J.M.; Galina, A.; et al. CD38 Dictates Age-Related NAD Decline and Mitochondrial Dysfunction through an SIRT3-Dependent Mechanism. Cell Metab. 2016, 23, 1127–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, H.C.; Gerbeth, C.; Thiru, P.; Vögtle, N.F.; Knoll, M.; Shahsafaei, A.; Samocha, K.E.; Huang, C.X.; Harden, M.M.; Song, R.; et al. A respiratory chain-controlled signal transduction cascade in the mitochondrial intermembrane space mediates hydrogen peroxide signaling. Proc. Natl. Acad. Sci. USA 2015, 112, E5679–E5688, Erratum in Proc. Natl. Acad. Sci. USA 2016, 113, E2094. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.H.; Wang, Y.; Gupta, P.; Goldenberg, D.M. Extensive crosslinking of CD22 by epratuzumab triggers BCR signaling and caspase-dependent apoptosis in human lymphoma cells. MAbs 2015, 7, 199–211. [Google Scholar] [CrossRef]
- Divakaruni, A.S.; Paradyse, A.; Ferrick, D.A.; Murphy, A.N.; Jastroch, M. Analysis and interpretation of microplate-based oxygen consumption and pH data. Methods Enzym. 2014, 547, 309–354. [Google Scholar]
- Nicholas, D.; Proctor, E.A.; Raval, F.M.; Ip, B.C.; Habib, C.; Ritou, E.; Grammatopoulos, T.N.; Steenkamp, D.; Dooms, H.; Apovian, C.M.; et al. Advances in the quantification of mitochondrial function in primary human immune cells through extracellular flux analysis. PLoS ONE 2017, 12, e0170975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrus, A.T.; Lighezan, D.L.; Danila, M.D.; Duicu, O.M.; Sturza, A.; Muntean, D.M.; Ionita, I. Assessment of platelet respiration as emerging biomarker of disease. Physiol. Res. 2019, 68, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Reyes, A.Z.; Hu, K.A.; Teperman, J.; Wampler Muskardin, T.L.; Tardif, J.C.; Shah, B.; Pillinger, M.H. Anti-inflammatory therapy for COVID-19 infection: The case for colchicine. Ann. Rheum. Dis. 2021, 80, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Streng, L.W.J.M.; de Wijs, C.J.; Raat, N.J.H.; Specht, P.A.C.; Sneiders, D.; van der Kaaij, M.; Endeman, H.; Mik, E.G.; Harms, F.A. In Vivo and Ex Vivo Mitochondrial Function in COVID-19 Patients on the Intensive Care Unit. Biomedicines 2022, 10, 1746. [Google Scholar] [CrossRef]
- Bohn, M.K.; Hall, A.; Sepiashvili, L.; Jung, B.; Steele, S.; Adeli, K. Pathophysiology of COVID-19: Mechanisms Underlying Disease Severity and Progression. Physioloigy 2020, 35, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Yau, W.W.; Singh, B.K.; Lesmana, R.; Zhou, J.; Sinha, R.A.; Wong, K.A.; Wu, Y.; Bay, B.H.; Sugii, S.; Sun, L.; et al. Thyroid hormone (T3) stimulates brown adipose tissue activation via mitochondrial biogenesis and MTOR-mediated mitophagy. Autophagy 2019, 15, 131–150. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Tzouvelekis, A.; Wang, R.; Herazo-Maya, J.D.; Ibarra, G.H.; Srivastava, A.; de Castro, J.P.W.; DeIuliis, G.; Ahangari, F.; Woolard, T.; et al. Thyroid hormone inhibits lung fibrosis in mice by improving epithelial mitochondrial function. Nat. Med. 2018, 24, 39–49. [Google Scholar] [CrossRef]
- Lombardi, A.; Moreno, M.; de Lange, P.; Iossa, S.; Busiello, R.A.; Goglia, F. Regulation of skeletal muscle mitochondrial activity by thyroid hormones: Focus on the “old” triiodothyronine and the “emerging” 3,5-diiodothyronine. Front. Physiol. 2015, 6, 237. [Google Scholar] [CrossRef] [Green Version]
- Brand, M.D. Uncoupling to survive? The role of mitochondrial inefficiency in ageing. Exp. Gerontol. 2000, 35, 811–820. [Google Scholar] [CrossRef]
- Porter, R.K.; Brand, M.D. Causes of the differences in respiration rate of hepatocytes from mammals of different body mass. Am. J. Physiol. 1995, 269, R1213–R1224. [Google Scholar] [CrossRef] [PubMed]
- Hafner, R.P.; Nobes, C.D.; McGown, A.D.; Brand, M.D. Altered relationship between protonmotive force and respiration rate in non-phosphorylating liver mitochondria isolated from rats of different thyroid hormone status. Eur. J. Biochem. 1988, 178, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Mourouzis, I.; Apostolaki, V.; Trikas, A.; Kokkinos, L.; Alexandrou, N.; Avdikou, M.; Giannoulopoulou, M.; Vassi, A.; Tseti, I.; Pantos, C. The Potential of Thyroid Hormone Therapy in Severe COVID-19: Rationale and Preliminary Evidence. Int. J. Environ. Res. Public Health 2022, 19, 8063. [Google Scholar] [CrossRef]
- Ibrahim, H.; Perl, A.; Smith, D.; Lewis, T.; Kon, Z.; Goldenberg, R.; Yarta, K.; Staniloae, C.; Williams, M. Therapeutic blockade of inflammation in severe COVID-19 infection with intravenous N-acetylcysteine. Clin. Immunol. 2020, 219, 108544. [Google Scholar] [CrossRef]
- Ghosh, A.; Joseph, B.; Anil, S. Nitric Oxide in the Management of Respiratory Consequences in COVID-19: A Scoping Review of a Different Treatment Approach. Cureus 2022, 14, e23852. [Google Scholar] [CrossRef]
- McAllister, R.M.; Albarracin, I.; Price, E.M.; Smith, T.K.; Turk, J.R.; Wyatt, K.D. Thyroid status and nitric oxide in rat arterial vessels. J. Endocrinol. 2005, 185, 111–119. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PATIENTS’S CHARACTERISTICS | ALL (n = 54) | LOW FT3 (n = 35) | VERY LOW FT3 (n = 9) | NORMAL FT3 (n = 10) |
|---|---|---|---|---|
| Age | 68 (29–93) | 69 (29–93) | 67 (29–93) | 69 (29–93) |
| Sex | ||||
| M | 37 (69%) | 22 (63%) | 7 (78%) | 8 (80%) |
| F | 17 (31%) | 13 (37%) | 2 (22%) | 2 (20%) |
| Comorbidity | 34 (63%) | 25 (71%) | 5 (56%) | 4 (40%) |
| Diabets | 5 (9%) | 3 (9%) | 1 (11%) | 1 (10%) |
| Hypertension | 21 (39%) | 15 (43%) | 3 (33%) | 3 (30%) |
| Heart disease | 15 (28%) | 12 (34%) | 3 (33%) | 0 (0%) |
| Obesity | 4 (7%) | 1 (3%) | 2 (22%) | 1 (10%) |
| PTS | GENDER | AGE (yrs) | HEIGHT (cm) | WEIGHT (Kg) | BMI (Kg/m2) | SOFA Score | TIME | FT3 (pg/mL) | FT4 (ng/dL) | TSH (µlU/mL) | THERAPY |
|---|---|---|---|---|---|---|---|---|---|---|---|
| #1 | M | 58 | 170 | 75 | 26 | 5 | T0 | 1.3 | 1 | 0.16 | Dexamethasone (4 mg OD) |
| T1 | 1 | 0.9 | 0.07 | ||||||||
| #2 | F | 53 | 170 | 85 | 29.4 | 2 | T0 | 1 | 0.9 | 0.39 | Dexamethasone (6 mg OD) + Liothyroinine (21.3 µg TID) |
| T1 | 1.8 | 0.7 | 0.03 |
| VERY LOW FT3 (<1.0 pg/mL) | LOW FT3 (>1.0 ≤ 1.7 pg/mL) | NORMAL FT3 (>1.8 pg/mL) | p VALUES (VERY LOW VS. NORMAL) | |
|---|---|---|---|---|
| dROMs (U-CARR) medium values (±SD) | 376.5 (±44.6) | 395.0 (±55.8) | 449.5 (±62.1) | p < 0.005 |
| BAP (µmol/L) medium values (±SD) | 2068.4 (±370.2) | 1680.8 (±682.6) | 1605.4 (±690.4) | p < 0.05 |
| OSi Index (DROMs/BAP) medium values (±SD) | 0.18 (±0.06) | 0.23 (±0.03) | 0.27 (±0.1) | p < 0.005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Vitis, C.; Capalbo, C.; Torsello, A.; Napoli, C.; Salvati, V.; Loffredo, C.; Blandino, G.; Piaggio, G.; Auciello, F.R.; Pelliccia, F.; et al. Opposite Effect of Thyroid Hormones on Oxidative Stress and on Mitochondrial Respiration in COVID-19 Patients. Antioxidants 2022, 11, 1998. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11101998
De Vitis C, Capalbo C, Torsello A, Napoli C, Salvati V, Loffredo C, Blandino G, Piaggio G, Auciello FR, Pelliccia F, et al. Opposite Effect of Thyroid Hormones on Oxidative Stress and on Mitochondrial Respiration in COVID-19 Patients. Antioxidants. 2022; 11(10):1998. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11101998
Chicago/Turabian StyleDe Vitis, Claudia, Carlo Capalbo, Alessandra Torsello, Christian Napoli, Valentina Salvati, Chiara Loffredo, Giovanni Blandino, Giulia Piaggio, Francesca Romana Auciello, Flaminia Pelliccia, and et al. 2022. "Opposite Effect of Thyroid Hormones on Oxidative Stress and on Mitochondrial Respiration in COVID-19 Patients" Antioxidants 11, no. 10: 1998. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11101998