
Inhibition of Tumor Growth and Modulation of Antioxidant Activity of Rhodoxanthin Isolated from Taxus baccata Aril against B16F10 Murine Malignant Melanoma
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Identification and Collection of Plant Materials
2.3. Plant Extract Preparation
2.4. Cell Line and Culture
2.5. Experimental Animals
2.6. Tumor Model and In Vivo Treatment Regimen
2.7. Hematological and Biochemical Analysis
2.8. ELISA Technique for Determining Carcinogenesis Indicators and DNA Oxidation Level
2.9. Histopathology Analysis
2.10. Statistical Analysis
3. Results and Discussions
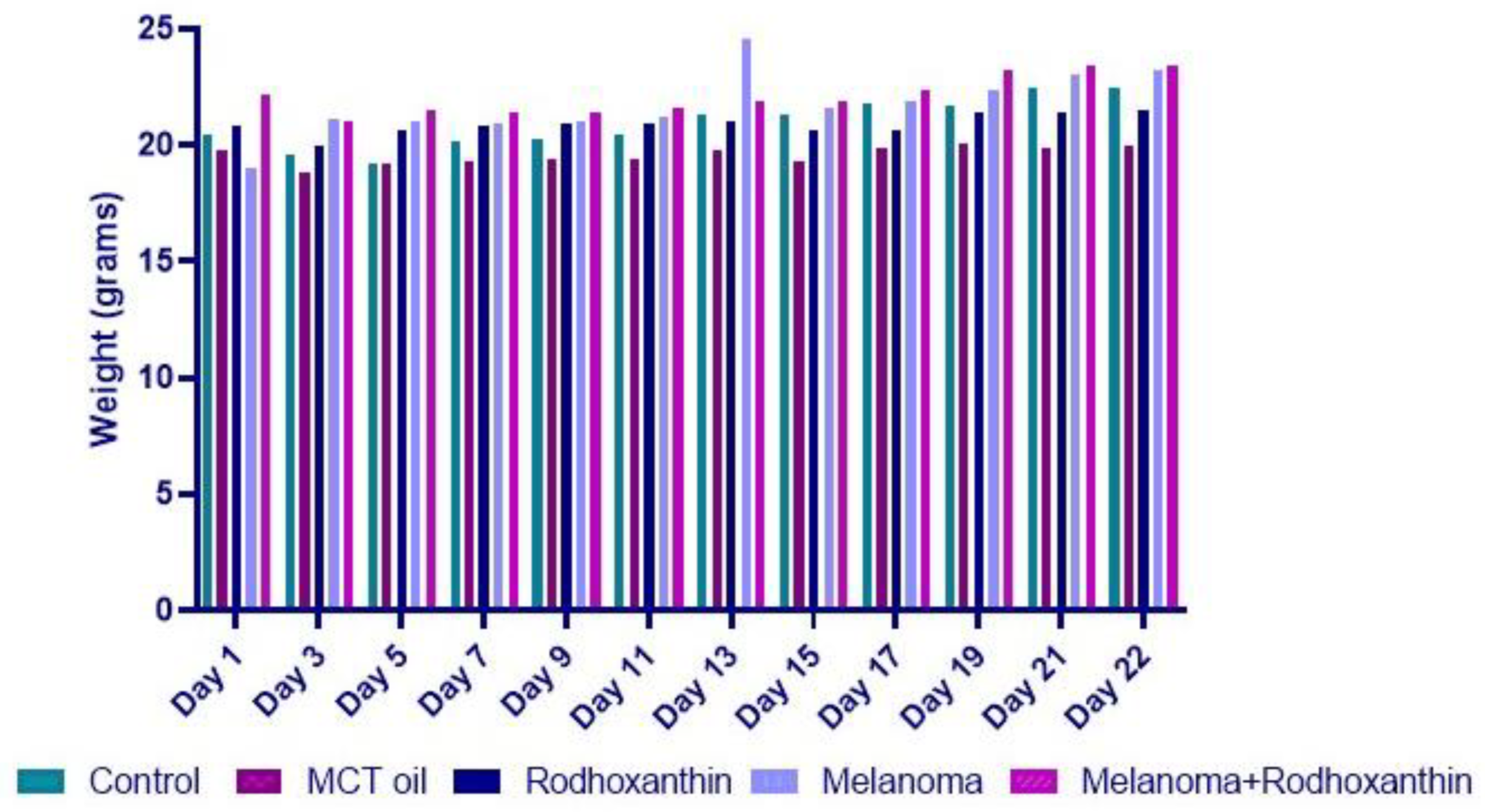
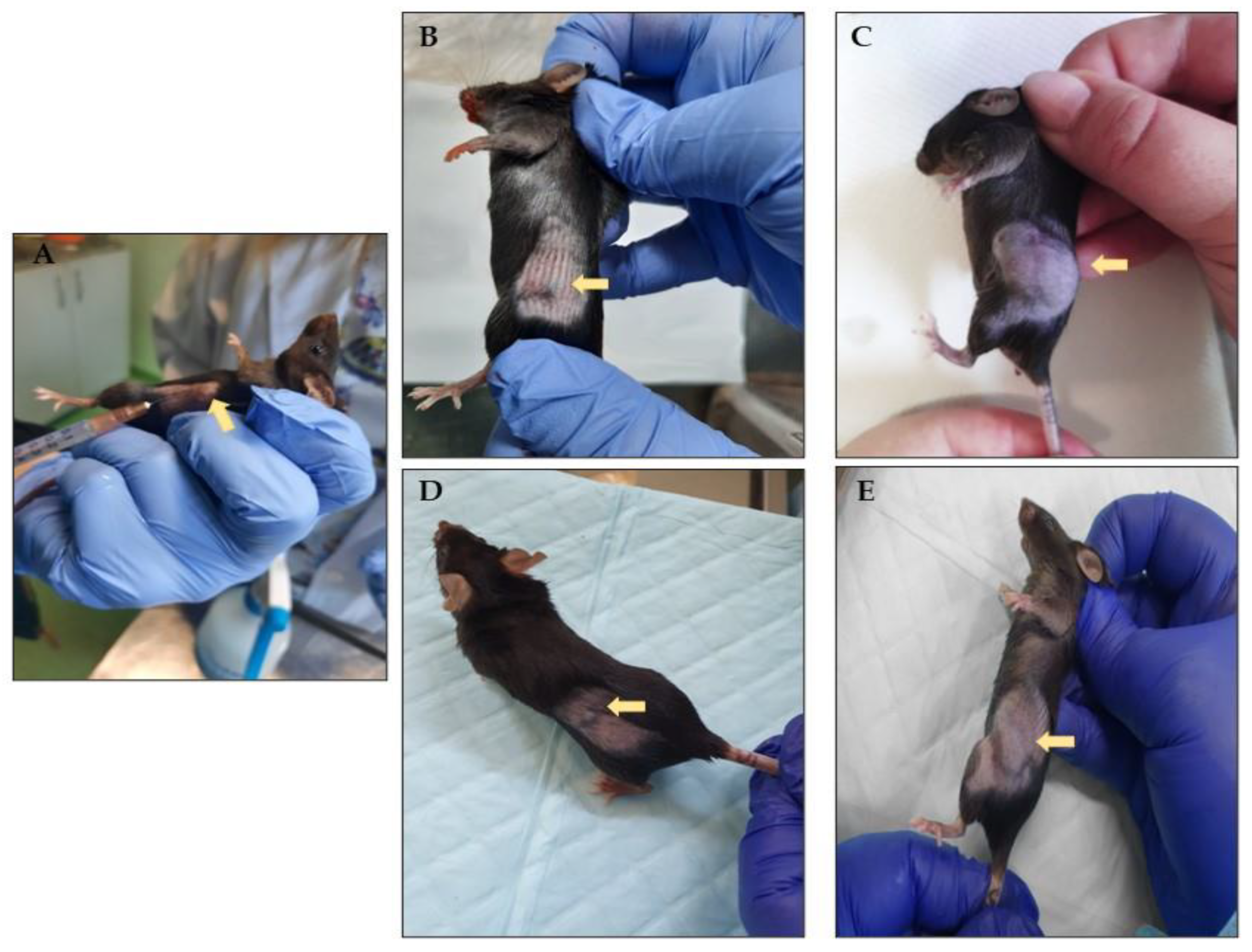
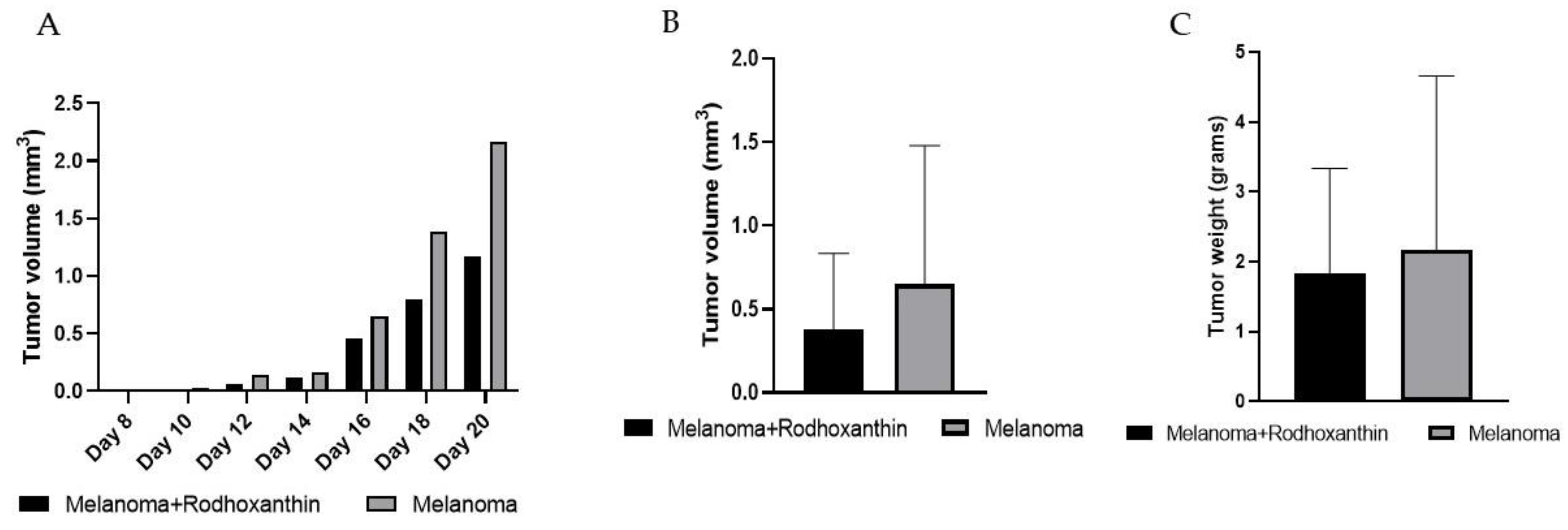
3.1. Rhodoxanthin’s Effect on B16F10 Murine Melanoma Growth in C57BL/6J Mice
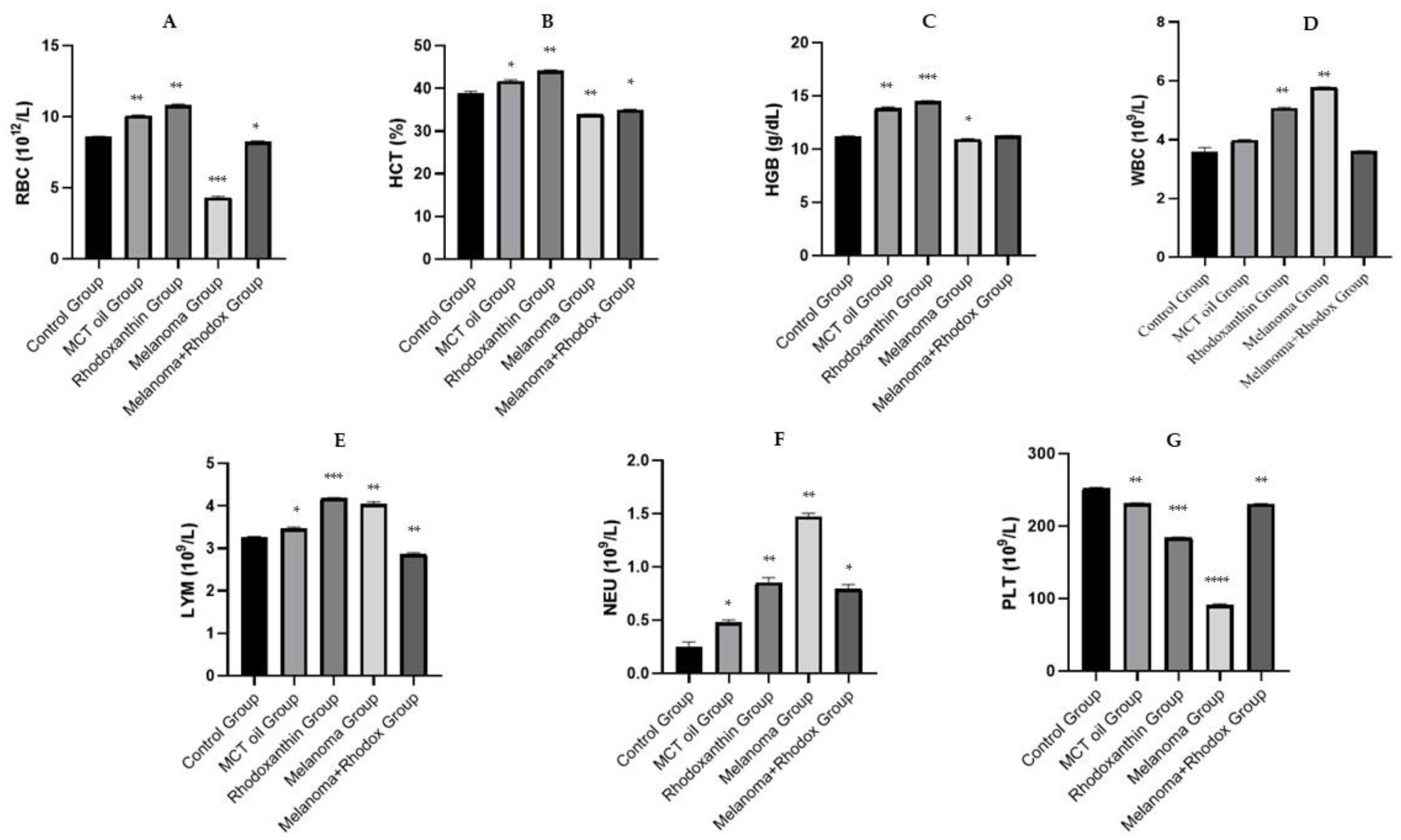
3.2. Hematological Analysis
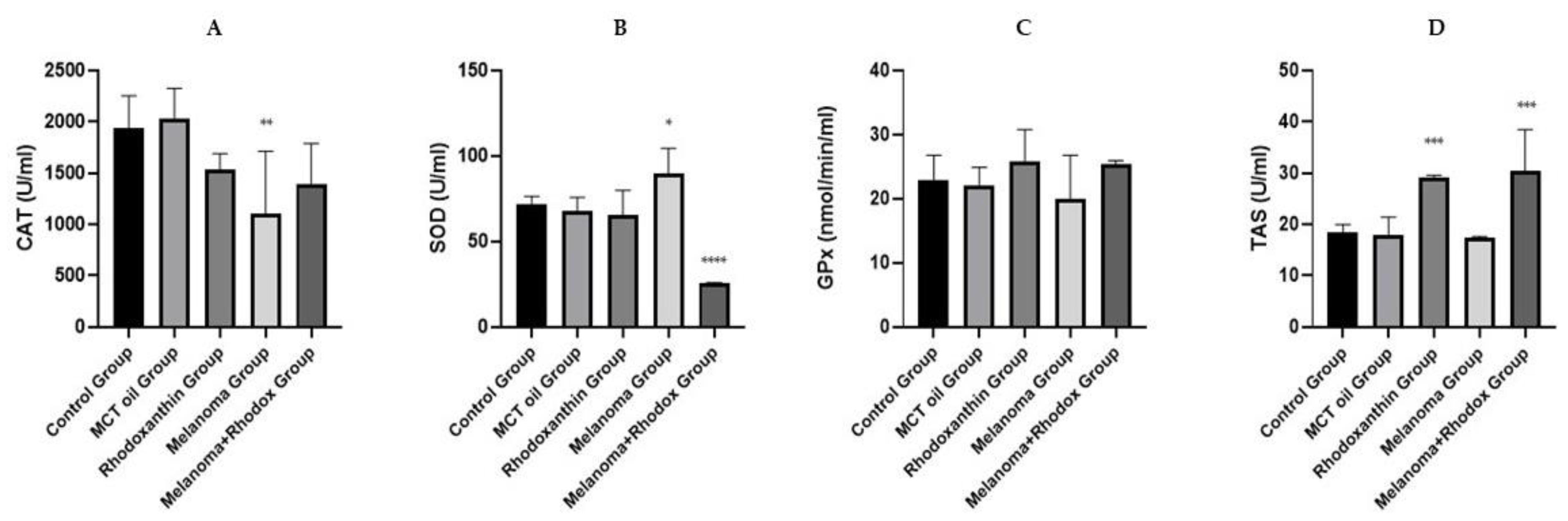
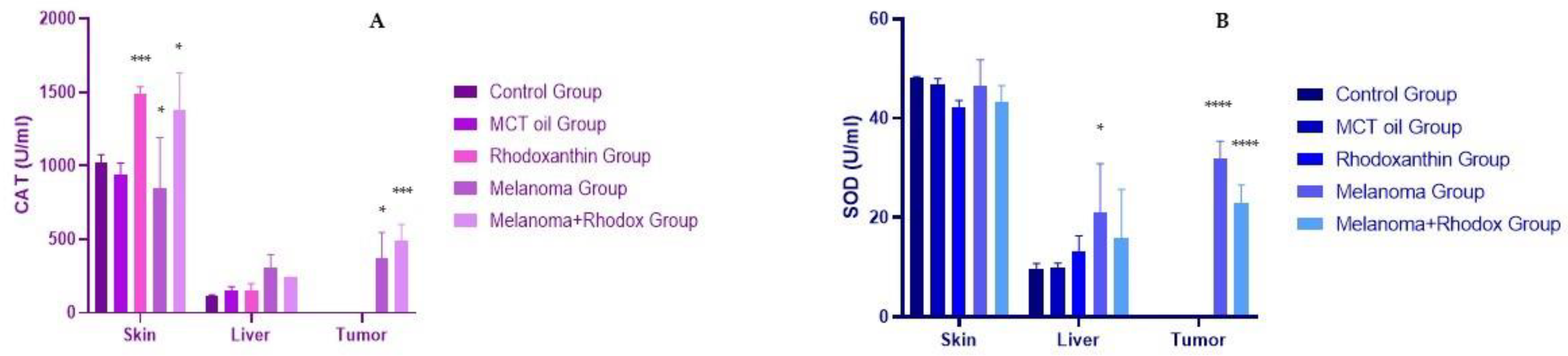
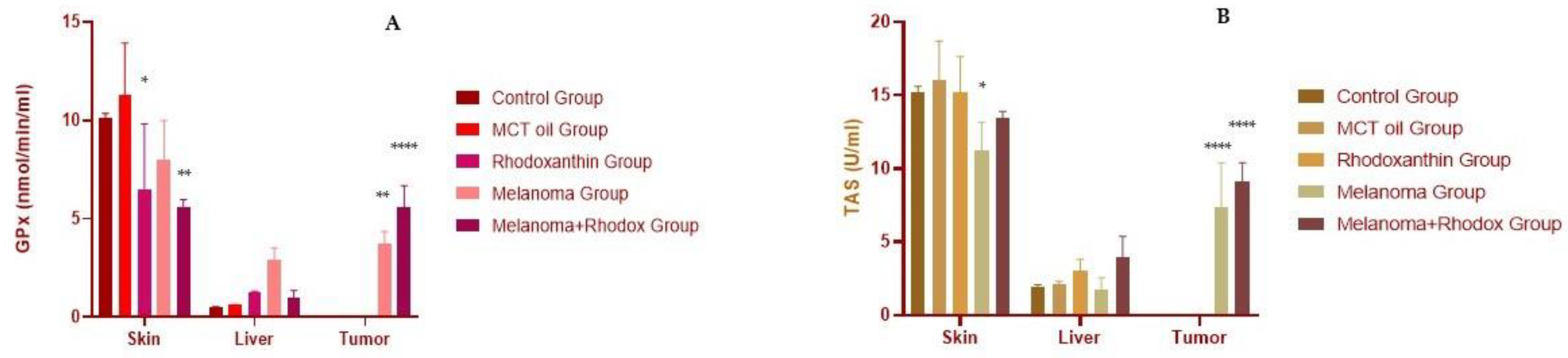
3.3. The Determination of Antioxidant Enzyme Activity
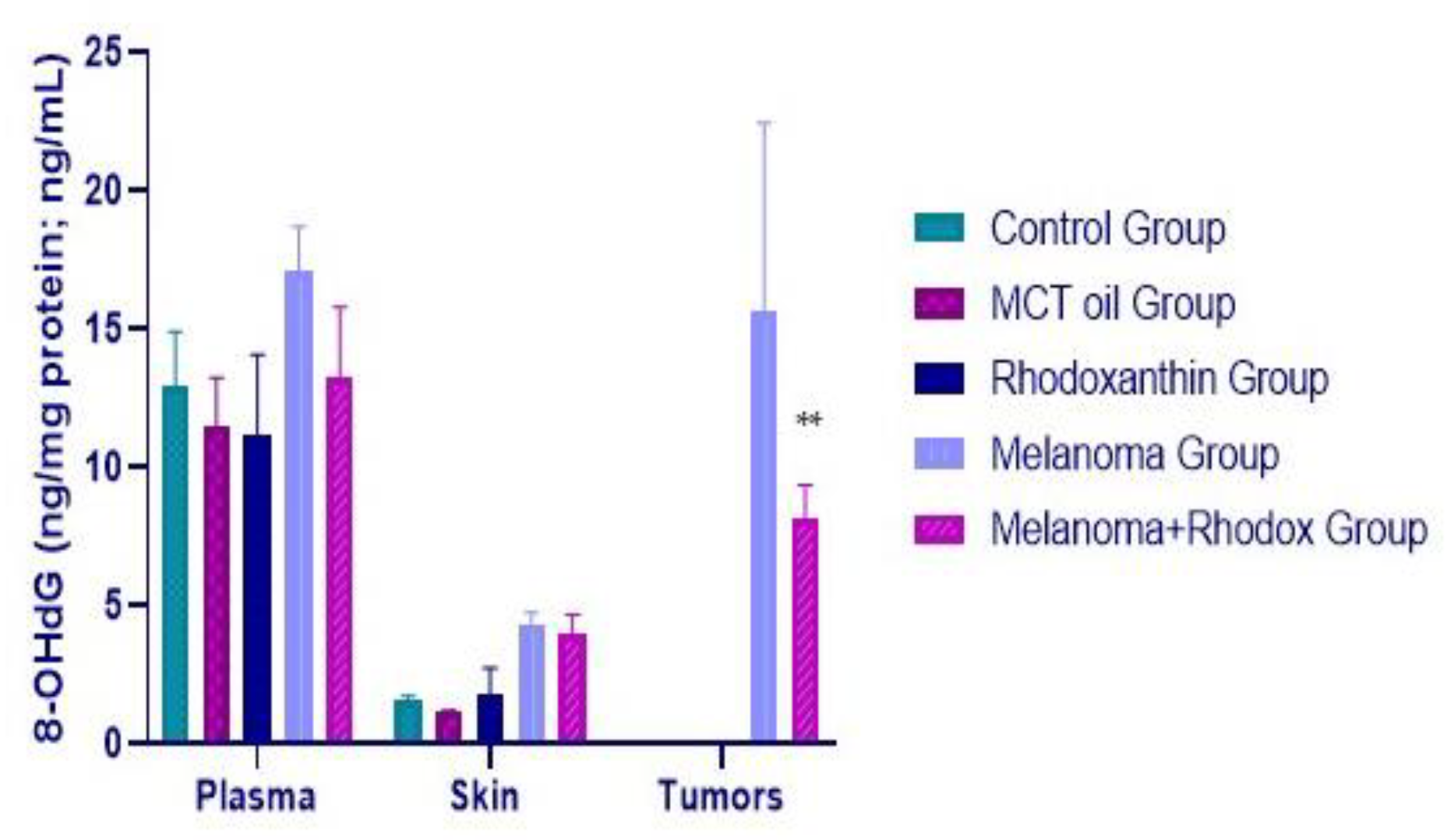
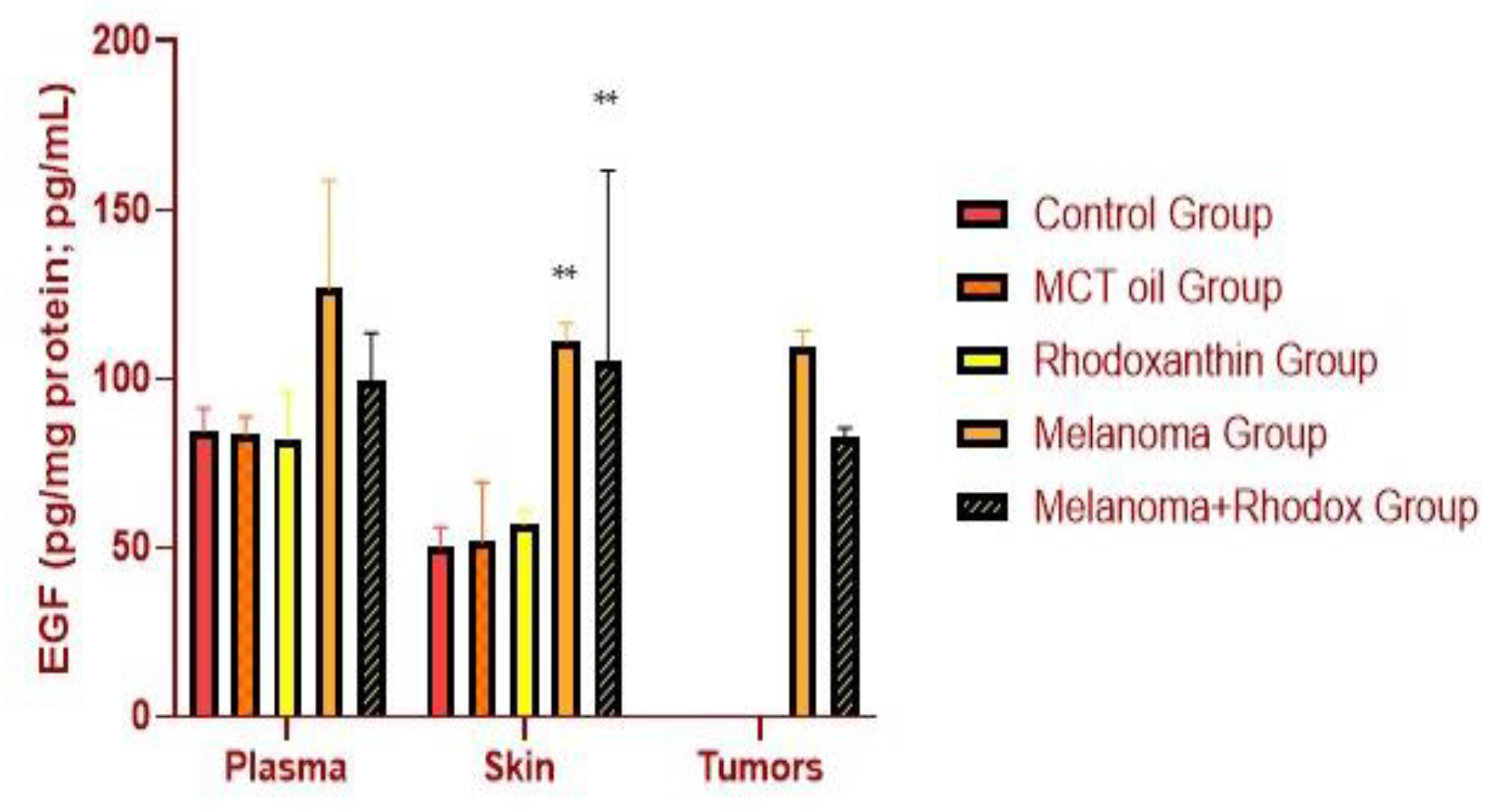
3.4. Determination of 8OHdG and EGF Activity
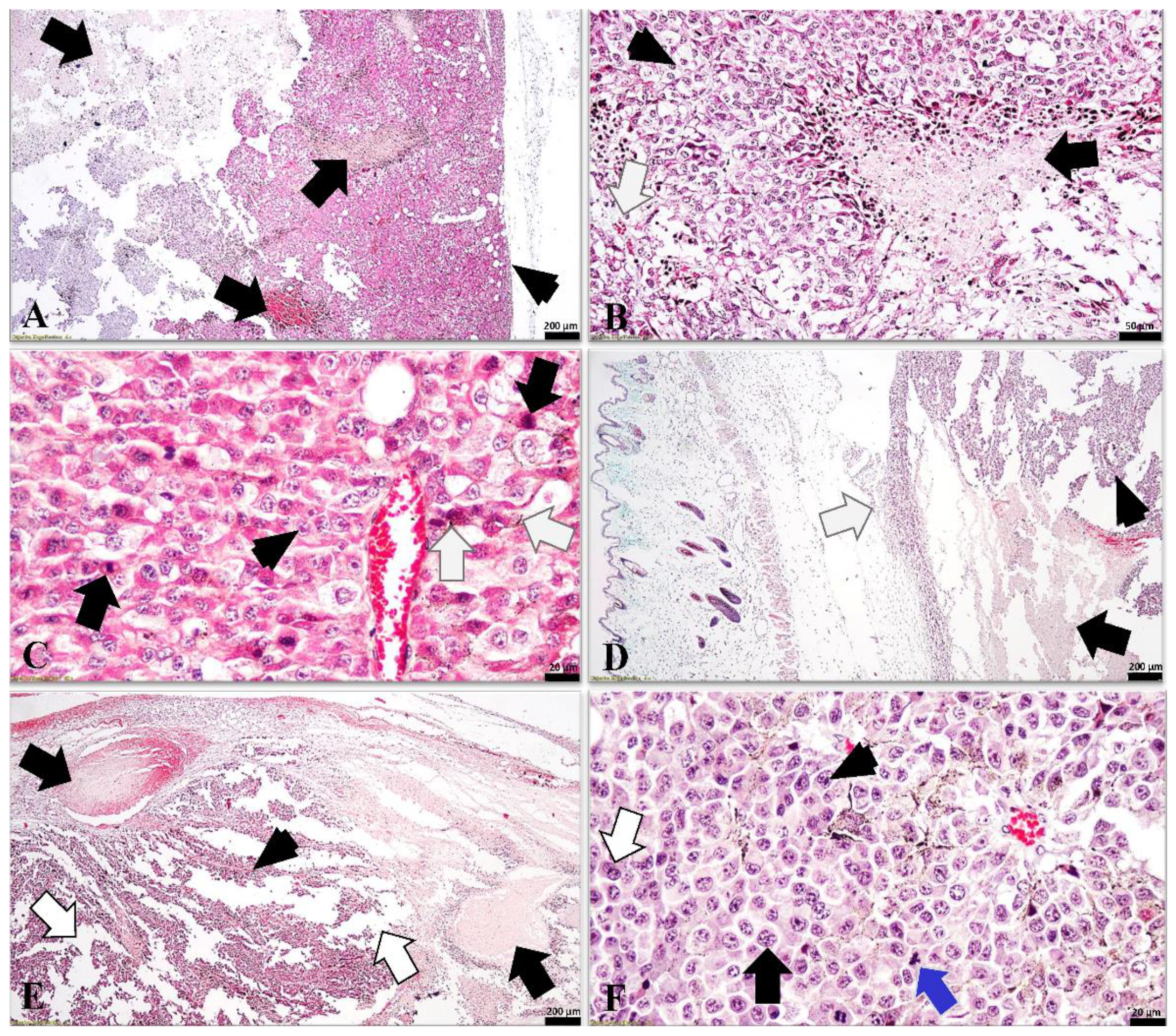
3.5. Histological Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gastaldello, G.H.; Cazeloto, A.C.V.; Ferreira, J.C.; Rodrigues, D.M.; Bastos, J.K.; Campo, V.L.; Zoccal, K.F.; Tefé-Silva, C. Green Propolis Compounds (Baccharin and p-Coumaric Acid) Show Beneficial Effects in Mice for Melanoma Induced by B16f10. Medicines 2021, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Iranzo, C.C.; Rubia-Ortí, J.E.; Castillo, S.S.; Firmino-Canhoto, J. Lesões cutâneas malignas e pré-malignas: Conhecimentos, hábitos e campanhas de prevenção solar. Acta Paul. Enferm. 2015, 28, 2–6. [Google Scholar] [CrossRef] [Green Version]
- Dumitraș, D.-A.; Andrei, S. Recent Advances in the Antiproliferative and Proapoptotic Activity of Various Plant Extracts and Constituents against Murine Malignant Melanoma. Molecules 2022, 27, 2585. [Google Scholar] [CrossRef] [PubMed]
- Pinello, K.; Pires, I.; Castro, A.F.; Carvalho, P.T.; Santos, A.; de Matos, A.; Queiroga, F.; Canadas-Sousa, A.; Dias-Pereira, P.; Catarino, J.; et al. Cross Species Analysis and Comparison of Tumors in Dogs and Cats, by Age, Sex, Topography and Main Morphologies. Data from Vet-OncoNet. Vet. Sci. 2022, 9, 167. [Google Scholar] [CrossRef] [PubMed]
- Goldschmidt, M.H.; Hendrick, M.J. Tumors of the skin and soft tissues. In Tumors in Domestic Animals, 4th ed.; Meuten, D.J., Ed.; Blackwell: Ames, IA, USA, 2002; pp. 45–117. [Google Scholar]
- Gross, T.L.; Ihrke, P.E.; Walder, E.J.; Affolter, V.K. Skin Diseases of the Dog and Cat: Clinical and Histopathologic Diagnosis, 2nd ed.; Blackwell Science: Oxford, UK, 2005; pp. 589–596. [Google Scholar]
- Miller, W.H., Jr.; Griffin, C.E.; Campbell, K.L. Muller&Kirk’s Small Animal Dermatology, 7th ed.; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Withrow, S.J.; Vail, D.M. Small Animal Clinical Oncology, 4th ed.; Saunders Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Jahanban-Esfahlan, A.; Modaeinama, S.; Abasi, M.; Abbasi, M.M.; Jahanban-Esfahlan, R. Anti proliferative properties of Melissa ofcinalis in diferent human cancer cells. Asian Pac. J. Cancer Prev. 2015, 16, 5703–5707. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.-C.; Chang, Y.-H.; Chang, C.-C.; Cheng, Y.-M.; Ou, Y.-C.; Chien, C.-C.C.; Hsu, Y.-C. Induction of Apoptosis in Endometrial Cancer (Ishikawa) Cells by Pogostemon cablin Aqueous Extract (PCAE). Int. J. Mol. Sci. 2015, 16, 12424–12435. [Google Scholar] [CrossRef] [Green Version]
- Yan, B.; Liu, L.; Zhao, Y.; Xiu, L.-J.; Sun, D.-Z.; Liu, X.; Lu, Y.; Shi, J.; Zhang, Y.-C.; Li, Y.-J.; et al. Xiaotan Sanjie decoction attenuates tumor angiogenesis by manipulating Notch-1-regulated proliferation of gastric cancer stem-like cells. World J. Gastroenterol. 2014, 20, 13105–13118. [Google Scholar] [CrossRef]
- Yi, J.-M.; Park, J.-S.; Lee, J.; Hong, J.T.; Bang, O.-S.; Kim, N.S. Anti-angiogenic potential of an ethanol extract of Annona atemoya seeds in vitro and in vivo. BMC Complement. Altern. Med. 2014, 14, 353. [Google Scholar]
- Benham, S.E.; Houston Durrant, T.; Caudullo, G.; de Rigo, D. Taxus baccata in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the EU: Luxembourg, 2016; p. e015921. [Google Scholar]
- Natasha, G.; Chan, M.; Gue, Y.X.; Gorog, D.A. Fatal heart block from intentional yew tree (Taxus baccata) ingestion: A case report. Eur. Heart J.- Case Rep. 2020, 4, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Zutter, A.; Hauri, K.; Evers, K.S.; Uhde, S.; Fassl, J.; Reuthebuch, O.T.; Berset, A.; Kuhne, M.; Donner, B.C. “Chaotic arrhythmia” during successful resuscitation after ingestion of yew (Taxus baccata) needles. Pediatr. Emerg. Care 2017, 35, e223–e225. [Google Scholar] [CrossRef]
- Willaert, W.; Claessens, P.; Vankelecom, B.; Vanderheyden, M. Intoxication with Taxus baccata: Cardiac arrhythmias following yew leaves ingestion. Pacing Clin. Electrophysiol. 2002, 25 Pt 1, 511–512. [Google Scholar] [CrossRef]
- Hudon, J.; Derbyshire, D.; Leckie, S.; Flinn, T. Diet-induced plumage erythrism in Baltimore Orioles as a result of the spread of introduced shrubs. Wilson J. Ornithol. 2013, 125, 88–96. [Google Scholar] [CrossRef]
- Tabaszewska, M.; Rutkowska, J.; Skoczylas, Ł.; Słupski, J.; Antoniewska, A.; Smolen, S.; Łukasiewicz, M.; Baranowski, D.; Duda, I.; Pietsch, J. Red Arils of Taxus baccata L.—A New Source of Valuable Fatty Acids and Nutrients. Molecules 2021, 26, 723. [Google Scholar] [CrossRef]
- Schex, R.; Lieb, V.M.; Schäfer, C.; Schweiggert, R.; Steingass, C.B. Carotenoid profiles of red-and yellow-colored arils of cultivars of Taxus baccata L. and Taxus× media Rehder. Phytochemistry 2021, 186, 112741. [Google Scholar] [CrossRef]
- Dumitraş, D.-A.; Bunea, A.; Vodnar, D.C.; Hanganu, D.; Pall, E.; Cenariu, M.; Gal, A.F.; Andrei, S. Phytochemical Characterization of Taxus baccata L. Aril with Emphasis on Evaluation of the Antiproliferative and Pro-Apoptotic Activity of Rhodoxanthin. Antioxidants 2022, 11, 1039. [Google Scholar] [CrossRef]
- Britton, G. UV/vis spectroscopy. In Carotenoids; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhauser: Basel, Switzerland; Boston, MA, USA; Berlin, Germany, 1995; Volume 1B: Spectroscopy, pp. 13–62. [Google Scholar]
- Roland, S.; Schweiggert, F.; Wüstenberg, B.; Bonrath, W.; Schäfer, C.; Schweiggert, R. Kinetic and Thermodynamic Study of the Thermally Induced (E/Z)-Isomerization of the retro-Carotenoid Rhodoxanthin. J. Agric. Food Chem. 2020, 68, 5259–5269. [Google Scholar] [CrossRef]
- Andrei, S.; Pintea, A.; Bunea, A. Studies regarding retro carotenoids content from Taxus baccata. In Proceedings of the XL Croatian Symposium on Agriculture with International Participation, Opatija, Croatia, 15–18 February 2005; pp. 653–655. [Google Scholar]
- ISO 10993-6; Biological Evaluation of Medical Devices—Part2: Animal Welfare Requirements. International Organization for Standardization: Geneva, Switzerland, 2006.
- Conway, T.F., Jr.; Sabel, M.S.; Sugano, M.; Frelinger, J.G.; Egilmez, N.K.; Chen, F.; Bankert, R.B. Growth of human tumor xenografts in SCID mice quantified using an immunoassay fot tumor marker protein in serum. J. Immunol. Methods 2000, 233, 57–65. [Google Scholar] [CrossRef]
- AMVA Panel of Euthanasia. Amva Guidelines for the Euthanasia of Animals: 2020 Edition; American Veterinary Medical Association: Schaumburg, IL, USA, 2020. [Google Scholar]
- Rajasekar, S.; Park, D.J.; Park, C.; Park, S.; Park, Y.H.; Kim, S.T.; Choi, Y.H.; Choi, Y.W. In vitro and in vivo anticancer effects of Lithospermum erythrorhizon extract on B16F10 murine melanoma. J. Ethnopharmacol. 2012, 144, 335–345. [Google Scholar] [CrossRef]
- Weichselbaum, T.E. An accurate and rapid method for the determination of proteins in small amounts of blood serum and plasma. Am. J. Clin. Pathol. 1946, 10, 40–49. [Google Scholar] [CrossRef]
- Zeng, F.; Harris, R.C. Epidermal growth factor, from gene organization to bedside. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2014; Volume 28. [Google Scholar]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2′-deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health Part C 2009, 27, 120–139. [Google Scholar] [CrossRef] [Green Version]
- Qing, X.; Shi, D.; Lv, X.; Wang, B.; Chen, S.; Shao, Z. Prognostic significance of 8-hydroxy-2′-deoxyguanosine in solid tumors: A meta-analysis. BMC Cancer 2019, 19, 997. [Google Scholar] [CrossRef] [PubMed]
- Gabe, M. Techniques Histologiques; Masson et Cie Editeurs: Paris, France, 1968; Chapter 9: Les Colorations Topographiques; pp. 203–256. [Google Scholar]
- Available online: https://www.nih.gov/news-events/nih-research-matters/mechanism-muscle-loss-cancer (accessed on 4 October 2022).
- Havanur, S.; Batish, I.; Cheruku, S.P.; Gourishetti, K.; JagadeeshBabu, P.E.; Kumar, N. Poly (N, N-diethyl acrylamide)/functionalized graphene quantum dots hydrogels loaded with doxorubicin as a nano-drug carrier for metastatic lung cancer in mice. Mater. Sci. Eng. C 2019, 105, 110094. [Google Scholar] [CrossRef] [PubMed]
- Boussabbeh, M.; Ben Salem, I.; Rjiba-Touati, K.; Bouyahya, C.; Neffati, F.; Najjar, M.F.; Abid-Essefi, S. The potential effect of patulin on mice bearing melanoma cells: An anti-tumour or carcinogenic effect? Tumor Biol. 2016, 37, 6285–6295. [Google Scholar] [CrossRef] [PubMed]
- Kamran, N.; Li, Y.; Sierra, M.; Alghamri, M.S.; Kadiyala, P.; Appelman, H.D.; Edwards, M.; Lowenstein, P.R.; Castro, M.G. Melanoma induced immunosuppression is mediated by hematopoietic dysregulation. Oncoimmunology 2018, 7, e1408750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alupei, M.C.; Licarete, E.; Patras, L.; Banciu, M. Liposomal simvastatin inhibits tumor growth via targeting tumor-associated macrophages-mediated oxidative stress. Cancer Lett. 2015, 356 Pt B, 946–952. [Google Scholar] [CrossRef]
- Sharma, S.; Koneri, R.; Sharma, G.K.; Chandrul, K.K. Assessment of Chemoprotective Potential of Curcumin against DMBA-Croton Oil Induced Skin Cancer in Mice. Eur. J. Med. Plants 2020, 31, 57–71. [Google Scholar] [CrossRef]
- Oberley, T.D.; Oberley, L.W. Antioxidant enzyme levels in cancer. Histology and Histopathology 1997, 12, 525–535. [Google Scholar]
- Nishikawa, M.; Hyoudou, K.; Kobayashi, Y.; Umeyama, Y.; Takakura, Y.; Hashida, M. Inhibition of metastatic tumor growth by targeted delivery of antioxidant enzymes. J. Control. Release 2005, 109, 101–107. [Google Scholar] [CrossRef]
- Andrei, S.; Bunea, A.; Pintea, A. Stresul Oxidativ şi Antioxidanţi Naturali; Editura Academic Press: Cluj-Napoca, Romania, 2014. [Google Scholar]
- Plavec, T.; Nemec-Svete, A.; Butinar, J.; Tozon, N.; Prezelj, M.; Kandel, B.; Kessler, M. Antioxidant status in canine cancer patients. Acta Vet. 2008, 58, 275–286. [Google Scholar]
- Vazquez, F.; Lim, J.H.; Chim, H.; Bhalla, K.; Girnun, G.; Pierce, K.; Clish, C.B.; Granter, S.R.; Widlund, H.R.; Spiegelman, B.M.; et al. PGC1α expression defines a subset of human melanoma tumors with increased mitochondrial capacity and resistance to oxidative stress. Cancer Cell 2013, 23, 287–301. [Google Scholar] [CrossRef] [Green Version]
- Cannavò, S.P.; Tonacci, A.; Bertino, L.; Casciaro, M.; Borgia, F.; Gangemi, S. The role of oxidative stress in the biology of melanoma: A systematic review. Pathol.-Res. Pract. 2019, 215, 21–28. [Google Scholar] [CrossRef]
- Sander, C.S.; Hamm, F.; Elsner, P.; Thiele, J.J. Oxidative stress in malignant melanoma and non-melanoma skin cancer. Br. J. Dermatol. 2003, 148, 913–922. [Google Scholar] [CrossRef]
- Nogués, M.R.; Giralt, M.; Cervelló, I.; del Castillo, D.; Espeso, O.; Argany, N.; Aliaga, A.; Mallol, J. Parameters related to oxygen free radicals in human skin: A study comparing healthy epidermis and skin cancer tissue. J. Invest. Dermatol. 2002, 119, 645–652. [Google Scholar] [CrossRef]
- Woźniak, A.; Drewa, G.; Woźniak, B.; Schachtschabel, D.O. Activity of antioxidant enzymes and concentration of lipid peroxidation products in selected tissues of mice of different ages, both healthy and melanoma-bearing. Z. Gerontol. Geriatr. 2004, 37, 184–189. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Control Group | MCT Oil Group | Rhodoxanthin Group | Melanoma Group | Melanoma + Rodhox Group |
|---|---|---|---|---|---|
| WBC (109/L) | 3.603 ± 0.14 | 3.977 ± 0.03 | 5.056 ± 0.04 | 5.777 ± 0.12 | 3.626 ± 0.05 |
| LYM (109/L) | 3.253 ± 0.03 | 3.466 ± 0.21 | 4.166 ± 0.03 | 4.036 ± 0.07 | 2.850 ± 0.05 |
| NEU (109/L) | 0.253 ± 0.04 | 0.476 ± 0.02 | 0.853 ± 0.04 | 1.473 ± 0.03 | 0.793 ± 0.04 |
| RBC (1012/L) | 8.593 ± 0.03 | 10.096 ± 0.02 | 10.826 ± 0.06 | 4.332 ± 0.05 | 8.250 ± 0.05 |
| HGB (g/dL) | 11.210 ± 0.04 | 13.876 ± 0.08 | 14.486 ± 0.09 | 10.943 ± 0.05 | 11.260 ± 0.03 |
| HCT (%) | 38.926 ± 0.41 | 41.713 ± 0.28 | 44.210 | 33.916 ± 0.13 | 35.010 ± 0.11 |
| PLT (109/L) | 252.12 ± 1.42 | 231.943 ± 0.41 | 184.5 ± 0.51 | 91.333 ± 1.52 | 230.643 ± 0.66 |
| Tissue/Parameters | Total Proteins (mg/mL) |
|---|---|
| Skin | |
| Control group | 1.39 ± 0.28 |
| MCT oil group | 1.53 ± 0.39 |
| Rhodoxanthin group | 1.08 ± 0.03 |
| Melanoma group | 2.05 ± 0.89 |
| Melanoma + Rhodoxanthin group | 1.102 ± 0.13 |
| Liver | |
| Control group | 14.30 ± 0.28 |
| MCT oil group | 10.56 ± 1.89 |
| Rhodoxanthin group | 8.71 ± 1.201 |
| Melanoma group | 5.94 ± 1.22 |
| Melanoma + Rhodoxanthin group | 5.50 ± 2.15 |
| Tumor | |
| Melanoma group | 3.18 ± 0.62 |
| Melanoma + Rhodoxanthin group | 3.19 ± 0.44 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dumitraş, D.-A.; Dreanca, A.I.; Pall, E.; Gal, A.F.; Rus, V.; Morohoschi, A.G.; Cotul, M.; Nan, M.I.; Andrei, S. Inhibition of Tumor Growth and Modulation of Antioxidant Activity of Rhodoxanthin Isolated from Taxus baccata Aril against B16F10 Murine Malignant Melanoma. Antioxidants 2022, 11, 2264. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11112264
Dumitraş D-A, Dreanca AI, Pall E, Gal AF, Rus V, Morohoschi AG, Cotul M, Nan MI, Andrei S. Inhibition of Tumor Growth and Modulation of Antioxidant Activity of Rhodoxanthin Isolated from Taxus baccata Aril against B16F10 Murine Malignant Melanoma. Antioxidants. 2022; 11(11):2264. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11112264
Chicago/Turabian StyleDumitraş, Daria-Antonia, Alexandra Iulia Dreanca, Emoke Pall, Adrian Florin Gal, Vasile Rus, Andreea Georgiana Morohoschi, Mihaela Cotul, Monica Irina Nan, and Sanda Andrei. 2022. "Inhibition of Tumor Growth and Modulation of Antioxidant Activity of Rhodoxanthin Isolated from Taxus baccata Aril against B16F10 Murine Malignant Melanoma" Antioxidants 11, no. 11: 2264. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11112264