
Impact of Seminal Plasma Antioxidants on Donkey Sperm Cryotolerance

 ,
,  ,
,  , ,
, ,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Ejaculates
2.2. Isolation of Seminal Plasma (SP)
2.3. Measurement of Antioxidants Activity Levels and OSI in SP
2.4. Sperm Cryopreservation
2.5. Assessment of Sperm Motility
2.6. Flow Cytometry Analyses
2.6.1. Evaluation of Sperm Plasma Membrane Integrity
2.6.2. Evaluation of Membrane Lipid Disorder
2.6.3. Evaluation of Mitochondrial Membrane Potential
2.6.4. Evaluation of Intracellular ROS Levels
2.6.5. Evaluation of Intracellular Calcium Levels
2.7. Experimental Design
2.8. Statistical Analysis
2.8.1. Hierarchical Clusters and Comparison between GFE and PFE
2.8.2. Correlation Matrix
3. Results
3.1. Classification of Donkey Ejaculates into GFE and PFE Groups According to Their Post-Thaw Sperm Quality and Functionality Parameters
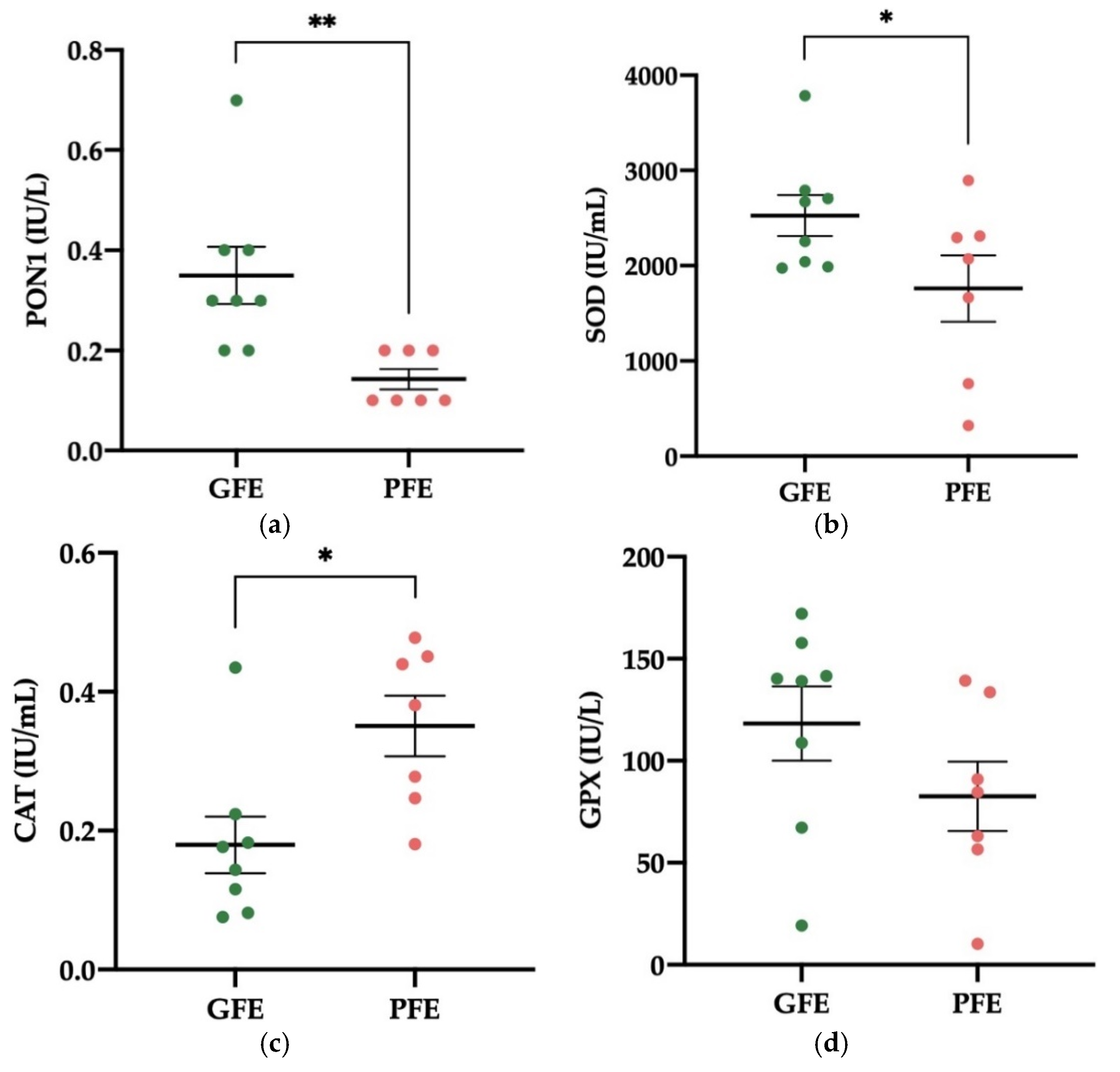
3.2. Activity Levels of Enzymatic Antioxidants in Donkey SP
3.3. Activity Levels of Non-Enzymatic Antioxidants in Donkey SP
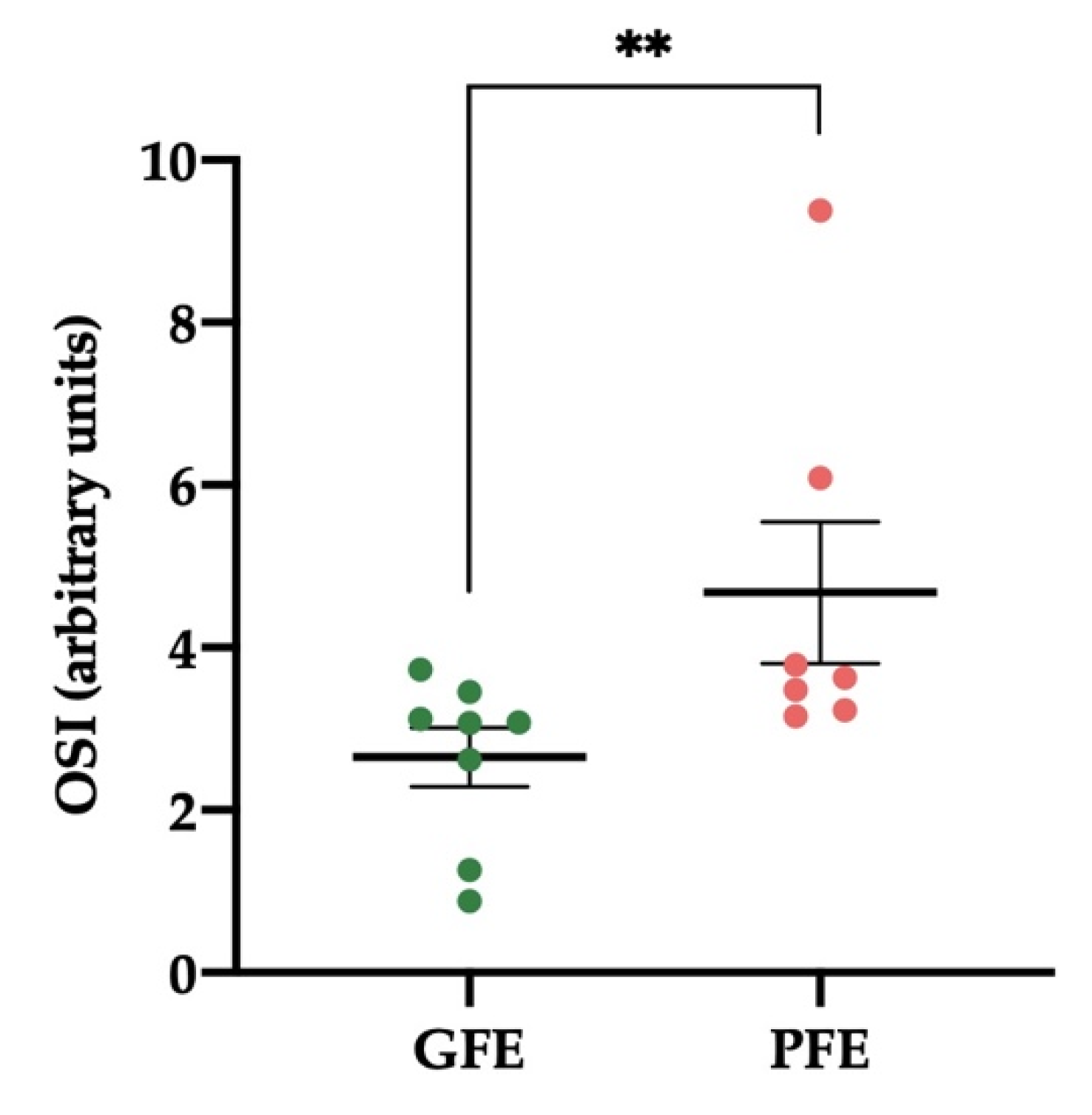
3.4. Levels of OSI in Donkey SP
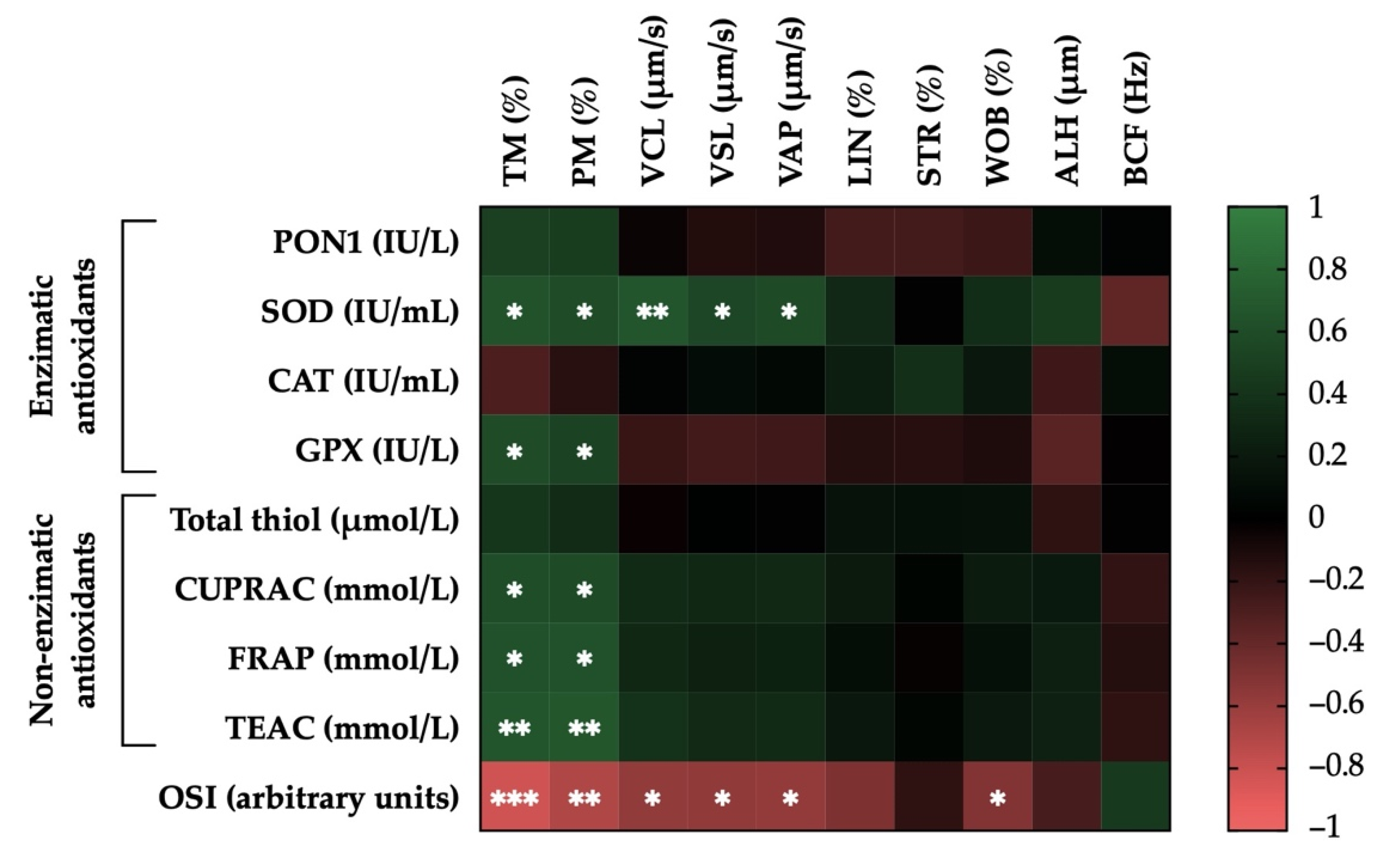
3.5. Correlations of the Activity Levels of Enzymatic and Non-Enzymatic Antioxidants and OSI in Donkey SP with Post-Thaw Sperm Motility Parameters
3.6. Correlations of the Activity Levels of Enzymatic and Non-Enzymatic Antioxidants and OSI in Donkey SP with Post-Thaw Sperm Functionality Parameters
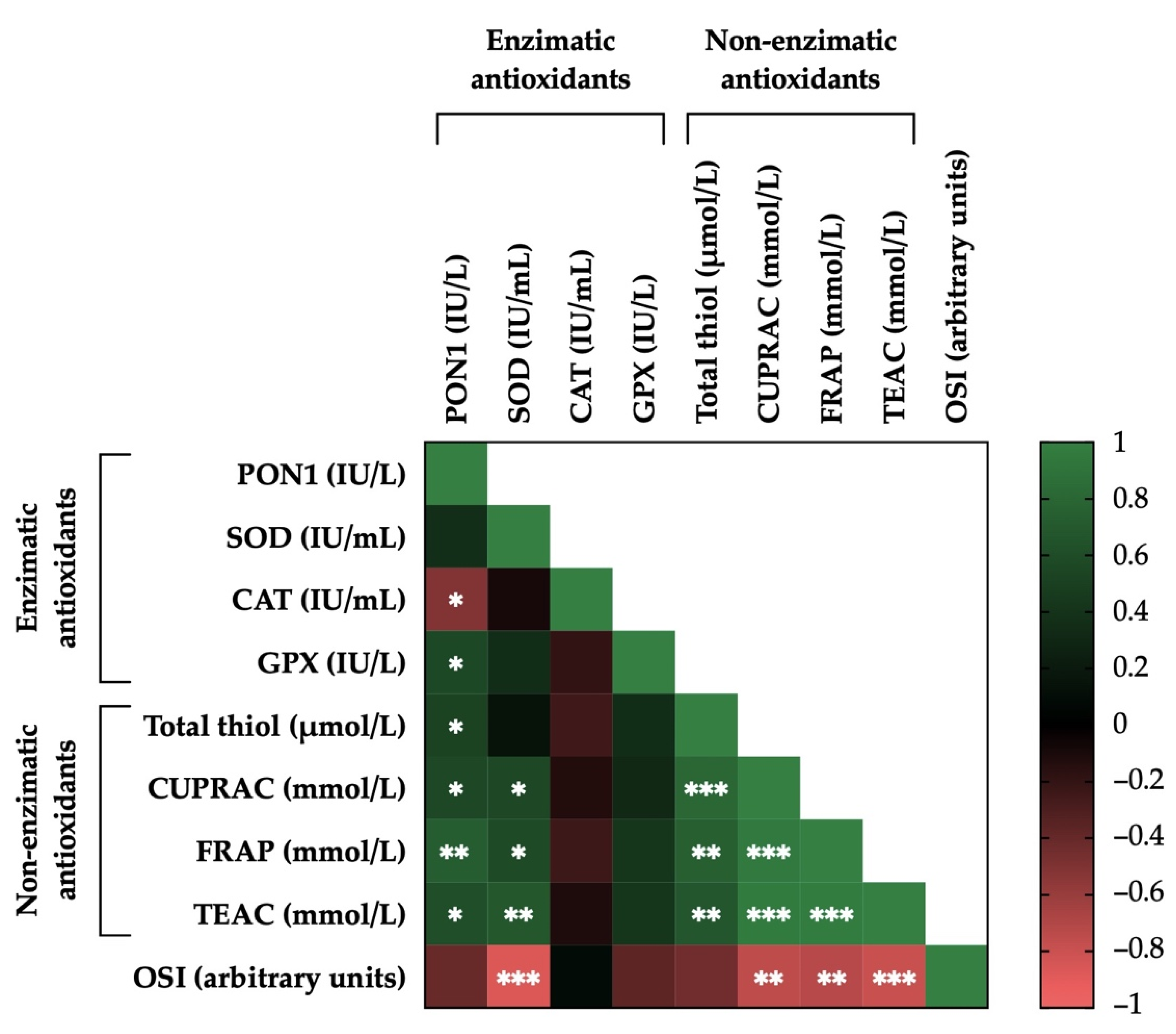
3.7. Correlations between the Activity Levels of Enzymatic and Non-Enzymatic Antioxidants and OSI in Donkey SP
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Camillo, F.; Rota, A.; Biagini, L.; Tesi, M.; Fanelli, D.; Panzani, D. The current situation and trend of donkey industry in Europe. J. Equine Vet. Sci. 2018, 65, 44–49. [Google Scholar] [CrossRef]
- Canisso, I.F.; Panzani, D.; Miró, J.; Ellerbrock, R.E. Key aspects of donkey and mule reproduction. Vet. Clin. N. Am. Equine Pract. 2019, 35, 607–642. [Google Scholar] [CrossRef] [PubMed]
- Di Palma, T.; Cecchini, S.; MacChia, G.; Pasolini, M.P.; Cocchia, N.; Boni, R. Kinematic, bioenergetic and oxidative evaluations of donkey sperm preserved at +4 °C. Zygote 2020, 28, 300–307. [Google Scholar] [CrossRef]
- Martini, M.; Altomonte, I.; Licitra, R.; Salari, F. Nutritional and nutraceutical quality of donkey milk. J. Equine Vet. Sci. 2018, 65, 33–37. [Google Scholar] [CrossRef]
- Miró, J.; Marín, H.; Catalán, J.; Papas, M.; Gacem, S.; Yeste, M. Seminal plasma, sperm concentration, and sperm-pmn interaction in the donkey: An in vitro model to study endometrial inflammation at post-insemination. Int. J. Mol. Sci. 2020, 21, 3478. [Google Scholar] [CrossRef]
- Papas, M.; Catalan, J.; Barranco, I.; Arroyo, L.; Bassols, A.; Yeste, M.; Miró, J. Total and specific activities of superoxide dismutase (SOD) in seminal plasma are related with the cryotolerance of jackass spermatozoa. Cryobiology 2020, 92, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, C.; González, N.; Luño, V.; Martínez, F.; Gil, L. Alternatives in Donkey semen cryopreservation: Mare vs. Jenny Colostrum. Reprod. Domest. Anim. 2019, 54, 94–97. [Google Scholar] [CrossRef]
- Vidament, M.; Vincent, P.; Martin, F.X.; Magistrini, M.; Blesbois, E. Differences in ability of jennies and mares to conceive with cooled and frozen semen containing glycerol or not. Anim. Reprod. Sci. 2009, 112, 22–35. [Google Scholar] [CrossRef]
- Canisso, I.F.; Carvalho, G.R.; Morel, M.D.; Ker, P.G.; Rodrigues, A.L.; Silva, E.C.; Da Silva, M.A.C. Seminal parameters and field fertility of cryopreserved donkey jack semen after insemination of horse mares. Equine Vet. J. 2011, 43, 179–183. [Google Scholar] [CrossRef]
- Oliveira, J.V.; Papa, F.O.; Melo-Oña, C.M.; Monteiro, G.A.; Puoli-Filho, J.N.P.; Alvarenga, M.A. New procedures to freeze donkey semen and its influence on mares and jennies fertility. J. Equine Vet. Sci. 2012, 32, 503–504. [Google Scholar] [CrossRef]
- Vilés, K.; Rabanal, R.; Rodríguez-Prado, M.; Miró, J. Effect of ketoprofen treatment on the uterine inflammatory response after AI of jennies with frozen semen. Theriogenology 2013, 79, 1019–1026. [Google Scholar] [CrossRef] [PubMed]
- Miró, J.; Papas, M. Post–artificial insemination endometrial inflammation and its control in donkeys. J. Equine Vet. Sci. 2018, 65, 38–43. [Google Scholar] [CrossRef]
- Mateo-Otero, Y.; Zambrano, F.; Catalán, J.; Sánchez, R.; Yeste, M.; Miro, J.; Fernandez-Fuertes, B. Seminal plasma, and not sperm, induces time and concentration-dependent neutrophil extracellular trap release in donkeys. Equine Vet. J. 2021, 54, 415–426. [Google Scholar] [CrossRef]
- Yeste, M.; Estrada, E.; Rocha, L.G.; Marín, H.; Rodríguez-Gil, J.E.; Miró, J. Cryotolerance of stallion spermatozoa is related to ROS production and mitochondrial membrane potential rather than to the integrity of sperm nucleus. Andrology 2015, 3, 395–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, W.V. Basic aspects of frozen storage of semen. Anim. Reprod. Sci. 2000, 62, 3–22. [Google Scholar] [CrossRef]
- Watson, P.F. The causes of reduced fertility with cryopreserved semen. Anim. Reprod. Sci. 2000, 60, 481–492. [Google Scholar] [CrossRef]
- Papas, M.; Catalán, J.; Fernandez-Fuertes, B.; Arroyo, L.; Bassols, A.; Miró, J.; Yeste, M. Specific activity of superoxide dismutase in stallion seminal plasma is related to sperm cryotolerance. Antioxidants 2019, 8, 539. [Google Scholar] [CrossRef] [Green Version]
- Yeste, M. Sperm cryopreservation update: Cryodamage, markers, and factors affecting the sperm freezability in pigs. Theriogenology 2016, 85, 47–64. [Google Scholar] [CrossRef]
- Sieme, H.; Oldenhof, H.; Wolkers, W. Sperm membrane behaviour during cooling and cryopreservation. Reprod. Domest. Anim. 2015, 50, 20–26. [Google Scholar] [CrossRef]
- Amidi, F.; Pazhohan, A.; Shabani Nashtaei, M.; Khodarahmian, M.; Nekoonam, S. The role of antioxidants in sperm freezing: A review. Cell Tissue Bank. 2016, 17, 745–756. [Google Scholar] [CrossRef]
- Aitken, R.J.; De Iuliis, G.N.; Drevet, J.R. Role of Oxidative Stress in the Etiology of Male Infertility and the Potential Therapeutic Value of Antioxidants. In Oxidants, Antioxidants, and Impact of the Oxidative Status in Male Reproduction; Elsevier: London, UK, 2018; pp. 91–100. [Google Scholar] [CrossRef]
- Papas, M.; Arroyo, L.; Bassols, A.; Catalán, J.; Bonilla-Correal, S.; Gacem, S.; Yeste, M.; Miró, J. Activities of antioxidant seminal plasma enzymes (SOD, CAT, GPX and GSR) are higher in jackasses than in stallions and are correlated with sperm motility in jackasses. Theriogenology 2019, 140, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Muiño-Blanco, T.; Pérez-Pé, R.; Cebrián-Pérez, J. Seminal plasma proteins and sperm resistance to stress. Reprod. Domest. Anim. 2008, 43, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.A.; Wolf, C.A.; Viu, M.A.O.; Gambarini, M.L. Addition of glutathione to an extender for frozen equine semen. J. Equine Vet. Sci. 2013, 33, 1148–1152. [Google Scholar] [CrossRef] [Green Version]
- Gadea, J.; García-Vazquez, F.; Matás, C.; Gardón, J.C.; Cánovas, S.; Gumbao, D. Cooling and freezing of boar spermatozoa: Supplementation of the freezing media with reduced glutathione preserves sperm function. J. Androl. 2005, 26, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.V.; Soares, A.T.; Batista, A.M.; Almeida, F.C.; Nunes, J.F.; Peixoto, C.A.; Guerra, M.M.P. In vitro and in vivo evaluation of ram sperm frozen in tris egg-yolk and wupplemented with superoxide dismutase and reduced glutathione. Reprod. Domest. Anim. 2011, 46, 874–881. [Google Scholar] [CrossRef]
- Gangwar, C.; Saxena, A.; Patel, A.; Singh, S.P.; Yadav, S.; Kumar, R.; Singh, V. Effect of reduced glutathione supplementation on cryopreservation induced sperm cryoinjuries in Murrah bull semen. Anim. Reprod. Sci. 2018, 192, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Ogata, K.; Sasaki, A.; Kato, Y.; Takeda, A.; Wakabayashi, M.; Sarentonglaga, B.; Yamaguchi, M.; Hara, A.; Fukumori, R.; Nagao, Y. Glutathione supplementation to semen extender improves the quality of frozen-thawed canine spermatozoa for transcervical insemination. J. Reprod. Dev. 2015, 61, 116–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowsett, K.F.; Knott, L.M. The influence of age and breed on stallion semen. Theriogenology 1996, 46, 397–412. [Google Scholar] [CrossRef]
- Sieme, H.; Harrison, R.A.P.; Petrunkina, A.M. Cryobiological determinants of frozen semen quality, with special reference to stallion. Anim. Reprod. Sci. 2008, 107, 276–292. [Google Scholar] [CrossRef]
- Loomis, P.R.; Graham, J.K. Commercial semen freezing: Individual male variation in cryosurvival and the response of stallion sperm to customized freezing protocols. Anim. Reprod. Sci. 2008, 105, 119–128. [Google Scholar] [CrossRef]
- Mari, G.; Castagnetti, C.; Rizzato, G.; Mislei, B.; Iacono, E.; Merlo, B. Density gradient centrifugation of sperm from a subfertile stallion and effect of seminal plasma addition on fertility. Anim. Reprod. Sci. 2011, 126, 96–100. [Google Scholar] [CrossRef]
- Cabrera, P.; Sánchez, R.; Risopatrón, J. Selección espermática en semen congelado/descongelado de equino: Evaluación de las membranas plasmática, acrosomal y potencial de membrana mitocondrial. Int. J. Morphol. 2014, 32, 725–731. [Google Scholar] [CrossRef] [Green Version]
- Neuhauser, S.; Gösele, P.; Handler, J. Postthaw addition of autologous seminal plasma improves sperm motion characteristics in fair and poor freezer stallions. J. Equine Vet. Sci. 2019, 72, 117–123. [Google Scholar] [CrossRef]
- Aurich, J.E.; Kühne, A.; Hoppe, H.; Aurich, C. Seminal plasma affects membrane integrity and motility of equine spermatozoa after cryopreservation. Theriogenology 1996, 46, 791–797. [Google Scholar] [CrossRef]
- Brinsko, S.P.; Crockett, E.C.; Squires, E.L. Effect of centrifugation and partial removal of seminal plasma on equine spermatozoal motility after cooling and storage. Theriogenology 2000, 54, 129–136. [Google Scholar] [CrossRef]
- Bromfield, J.J. A role for seminal plasma in modulating pregnancy outcomes in domestic species. Reproduction 2016, 152, R223–R232. [Google Scholar] [CrossRef] [Green Version]
- Mogielnicka-Brzozowska, M.; Kordan, W. Characteristics of selected seminal plasma proteins and their application in the improvement of the reproductive processes in mammals. Pol. J. Vet. Sci. 2011, 14, 489–499. [Google Scholar] [CrossRef]
- Li, J.; Barranco, I.; Tvarijonaviciute, A.; Molina, M.F.; Martinez, E.A.; Rodriguez-Martinez, H.; Parrilla, I.; Roca, J. Seminal plasma antioxidants are directly involved in boar sperm cryotolerance. Theriogenology 2018, 107, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Kenney, R.M.; Bergman, R.V.; Cooper, W.L.; Morse, G.W. Minimal contamination techniques for breeding mares: Technique and preliminary findings. Proc. Am. Assoc. Equine Pract. 1975, 21, 327–336. [Google Scholar]
- Bamba, K. Evaluation of acrosomal integrity of boar spermatozoa by bright field microscopy using an eosin-nigrosin stain. Theriogenology 1988, 29, 1245–1251. [Google Scholar] [CrossRef]
- Jocelyn, P.C. Spectrophotometric assay of thiols. Methods Enzymol. 1987, 143, 44–67. [Google Scholar] [CrossRef]
- Da Costa, C.M.; Dos Santos, R.C.C.; Lima, E.S. A simple automated procedure for thiol measurement in human serum samples. J. Bras. Patol. Med. Lab. 2006, 42, 345–350. [Google Scholar] [CrossRef]
- Erel, O. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin. Biochem. 2004, 37, 277–285. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “Antioxidant Power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Campos, C.; Guzmán, R.; López-Fernández, E.; Casado, Á. Evaluation of the copper(II) reduction assay using bathocuproinedisulfonic acid disodium salt for the total antioxidant capacity assessment: The CUPRAC–BCS assay. Anal. Biochem. 2009, 392, 37–44. [Google Scholar] [CrossRef]
- Barranco, I.; Roca, J.; Tvarijonaviciute, A.; Rubér, M.; Vicente-Carrillo, A.; Atikuzzaman, M.; Ceron, J.J.; Martinez, E.A.; Rodriguez-Martinez, H. Measurement of activity and concentration of paraoxonase 1 (PON-1) in seminal plasma and identification of PON-2 in the sperm of boar ejaculates. Mol. Reprod. Dev. 2015, 82, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem. 2005, 38, 1103–1111. [Google Scholar] [CrossRef]
- Wu, R.; Feng, J.; Yang, Y.; Dai, C.; Lu, A.; Li, J.; Liao, Y.; Xiang, M.; Huang, Q.; Wang, D.; et al. Significance of serum total oxidant/antioxidant status in patients with colorectal cancer. PLoS ONE 2017, 12, e0170003. [Google Scholar] [CrossRef]
- Flores, E.; Taberner, E.; Rivera, M.M.; Peña, A.; Rigau, T.; Miró, J.; Rodríguez-Gil, J.E. Effects of freezing/thawing on motile sperm subpopulations of boar and donkey ejaculates. Theriogenology 2008, 70, 936–945. [Google Scholar] [CrossRef]
- Lee, J.A.; Spidlen, J.; Boyce, K.; Cai, J.; Crosbie, N.; Dalphin, M.; Furlong, J.; Gasparetto, M.; Goldberg, M.; Goralczyk, E.M.; et al. MIFlowCyt: The minimum information about a flow cytometry experiment. Cytom. Part A 2008, 73A, 926–930. [Google Scholar] [CrossRef] [Green Version]
- Petrunkina, A.M.; Waberski, D.; Bollwein, H.; Sieme, H. Identifying non-sperm particles during flow cytometric physiological assessment: A simple approach. Theriogenology 2010, 73, 995–1000. [Google Scholar] [CrossRef]
- Garner, D.L.; Johnson, L.A. Viability assessment of mammalian sperm using SYBR-14 and propidium iodide. Biol. Reprod. 1995, 53, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.; Ashworth, P.; Miller, N. Bicarbonate/CO2, an effector of capacitation, induces a rapid and reversible change in the lipid architecture of boar sperm plasma membranes. Mol. Reprod. Dev. 1996, 45, 378–391. [Google Scholar] [CrossRef]
- Catalán, J.; Papas, M.; Trujillo-Rojas, L.; Blanco-Prieto, O.; Bonilla-Correal, S.; Rodríguez-Gil, J.E.; Miró, J.; Yeste, M. Red LED light acts on the mitochondrial electron chain of donkey sperm and its effects depend on the time of exposure to light. Front. Cell Dev. Biol. 2020, 8, 588621. [Google Scholar] [CrossRef]
- Guthrie, H.D.; Welch, G.R. Determination of intracellular reactive oxygen species and high mitochondrial membrane potential in Percoll-treated viable boar sperm using fluorescence-activated flow cytometry1. J. Anim. Sci. 2006, 84, 2089–2100. [Google Scholar] [CrossRef] [Green Version]
- Yeste, M.; Fernández-Novell, J.M.; Ramió-Lluch, L.; Estrada, E.; Rocha, L.G.; Cebrián-Pérez, J.A.; Muiño-Blanco, T.; Concha, I.I.; Ramírez, A.; Rodríguez-Gil, J.E. Intracellular calcium movements of boar spermatozoa during ‘in vitro’ capacitation and subsequent acrosome exocytosis follow a multiple-storage place, extracellular calcium-dependent model. Andrology 2015, 3, 729–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadirvel, G.; Kumar, S.; Kumaresan, A.; Kathiravan, P. Capacitation status of fresh and frozen-thawed buffalo spermatozoa in relation to cholesterol level, membrane fluidity and intracellular calcium. Anim. Reprod. Sci. 2009, 116, 244–253. [Google Scholar] [CrossRef]
- Morató, R.; Prieto-Martínez, N.; Muiño, R.; Hidalgo, C.O.; Rodríguez-Gil, J.E.; Bonet, S.; Yeste, M. Aquaporin 11 is related to cryotolerance and fertilising ability of frozen–thawed bull spermatozoa. Reprod. Fertil. Dev. 2018, 30, 1099–1108. [Google Scholar] [CrossRef]
- Szczykutowicz, J.; Kałuza, A.; Kaźmierowska-Niemczuk, M.; Ferens-Sieczkowska, M. The potential role of seminal plasma in the fertilization outcomes. BioMed Res. Int. 2019, 2019, 5397804. [Google Scholar] [CrossRef]
- Carretero, M.I.; Fumuso, F.G.; Giuliano, M.S. Effect of seminal plasma in the conservation of semen in zootechnical species. Spermova 2016, 6, 93–98. [Google Scholar] [CrossRef]
- Troedsson, M.H.T.; Desvousges, A.; Alghamdi, A.S.; Dahms, B.; Dow, C.A.; Hayna, J.; Valesco, R.; Collahan, P.T.; Macpherson, M.L.; Pozor, M.; et al. Components in seminal plasma regulating sperm transport and elimination. Anim. Reprod. Sci. 2005, 89, 171–186. [Google Scholar] [CrossRef]
- Jonakova, V.; Jonak, J.; Ticha, M. Proteomics of Male Seminal Plasma. In Reproductive Genomics in Domestic Animals; Wiley-Blackwell: Oxford, UK, 2010; pp. 339–366. [Google Scholar]
- Barranco, I.; Padilla, L.; Tvarijonaviciute, A.; Parrilla, I.; Martínez, E.A.; Rodriguez-Martinez, H.; Yeste, M.; Roca, J. Levels of activity of superoxide dismutase in seminal plasma do not predict fertility of pig AI-semen doses. Theriogenology 2019, 140, 18–24. [Google Scholar] [CrossRef]
- Yánez-Ortiz, I.; Catalán, J.; Mateo-Otero, Y.; Dordas-Perpinyà, M.; Gacem, S.; Yeste, N.; Bassols, A.; Yeste, M.; Miró, J. Extracellular reactive oxygen species (ROS) production in fresh donkey sperm exposed to reductive stress, oxidative stress and NETosis. Antioxidants 2021, 10, 1367. [Google Scholar] [CrossRef] [PubMed]
- Baumber, J.; Ball, B.A. Determination of glutathione peroxidase and superoxide dismutase-like activities in equine spermatozoa, seminal plasma, and reproductive tissues. Am. J. Vet. Res. 2005, 66, 1415–1419. [Google Scholar] [CrossRef] [PubMed]
- Waheed, M.M.; Gouda, E.M.; Khalifa, T.A.A. Impact of seminal plasma superoxide dismutase and glutathione peroxidase on cryopreserved buffalo spermatozoa. Anim. Reprod. Sci. 2013, 142, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Buffone, M.G.; Calamera, J.C.; Brugo-Olmedo, S.; Vincentiis, S.; Calamera, M.M.; Storey, B.T.; Doncel, G.F.; Alvarez, J.G. Superoxide dismutase content in sperm correlates with motility recovery after thawing of cryopreserved human spermatozoa. Fertil. Steril. 2012, 97, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Kowalowka, M.; Wysocki, P.; Fraser, L.; Strzezek, J. Extracellular superoxide dismutase of boar seminal plasma. Reprod. Domest. Anim. 2008, 43, 490–496. [Google Scholar] [CrossRef]
- Silvestre, M.A.; Yániz, J.L.; Peña, F.J.; Santolaria, P.; Castelló-Ruiz, M. Role of antioxidants in cooled liquid storage of mammal spermatozoa. Antioxidants 2021, 10, 1096. [Google Scholar] [CrossRef]
- Kehrer, J.P. The Haber–Weiss reaction and mechanisms of toxicity. Toxicology 2000, 149, 43–50. [Google Scholar] [CrossRef]
- Koppers, A.J.; De Iuliis, G.N.; Finnie, J.M.; McLaughlin, E.A.; Aitken, R.J. Significance of mitochondrial reactive oxygen species in the generation of oxidative stress in spermatozoa. J. Clin. Endocrinol. Metab. 2008, 93, 3199–3207. [Google Scholar] [CrossRef] [Green Version]
- Ghaleno, L.R.; Valojerdi, M.R.; Janzamin, E.; Chehrazi, M.; Sharbatoghli, M.; Yazdi, R.S. Evaluation of conventional semen parameters, intracellular reactive oxygen species, DNA fragmentation and dysfunction of mitochondrial membrane potential after semen preparation techniques: A flow cytometric study. Arch. Gynecol. Obstet. 2014, 289, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.M.; Janes, M.S.; Pehar, M.; Monette, J.S.; Ross, M.F.; Hagen, T.M.; Murphy, M.P.; Beckman, J.S. Selective fluorescent imaging of superoxide in vivo using ethidium-based probes. Proc. Natl. Acad. Sci. USA 2006, 103, 15038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, A.; Boni, R.; Buia, M.C.; Monfrecola, V.; Esposito, M.C.; Tosti, E. Ocean acidification impact on ascidian Ciona robusta spermatozoa: New evidence for stress resilience. Sci. Total Environ. 2019, 697, 134100. [Google Scholar] [CrossRef] [PubMed]
- De Iuliis, G.N.; Wingate, J.K.; Koppers, A.J.; McLaughlin, E.A.; Aitken, R.J. Definitive evidence for the nonmitochondrial production of superoxide anion by human spermatozoa. J. Clin. Endocrinol. Metab. 2006, 91, 1968–1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aitken, R.J.; Iuliis, G.N. De Role of Oxidative Stress in the Etiology of Sperm DNA Damage. In Sperm Chromatin; Springer: New York, NY, USA, 2011; pp. 277–293. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2019, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Kasimanickam, R.; Pelzer, K.D.; Kasimanickam, V.; Swecker, W.S.; Thatcher, C.D. Association of classical semen parameters, sperm DNA fragmentation index, lipid peroxidation and antioxidant enzymatic activity of semen in ram-lambs. Theriogenology 2006, 65, 1407–1421. [Google Scholar] [CrossRef]
- Rubio-Riquelme, N.; Huerta-Retamal, N.; Gómez-Torres, M.J.; Martínez-Espinosa, R.M. Catalase as a molecular target for male infertility diagnosis and monitoring: An overview. Antioxidants 2020, 9, 78. [Google Scholar] [CrossRef] [Green Version]
- Aitken, R.J.; Buckingham, D.; Harkiss, D. Use of a xanthine oxidase free radical generating system to investigate the cytotoxic effects of reactive oxygen species on human spermatozoa. Reproduction 1993, 97, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Aitken, R.J.; Drevet, J.R. The importance of oxidative stress in determining the functionality of mammalian spermatozoa: A two-edged sword. Antioxidants 2020, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Baiardi, G.; Ruiz, R.D.; Cuneo, M.F.; Ponce, A.A.; Lacuara, J.L.; Vincent, L. Differential effects of pharmacologically generated reactive oxygen species upon functional activity of epididymal mouse spermatozoa. Can. J. Physiol. Pharmacol. 1997, 75, 173–178. [Google Scholar] [CrossRef]
- Baumber, J.; Ball, B.A.; Gravance, C.G.; Medina, V.; Davies-Morel, M.C.G. The effect of reactive oxygen species on equine sperm motility, viability, acrosomal integrity, mitochondrial membrane potential, and membrane lipid peroxidation. J. Androl. 2000, 21, 895–902. [Google Scholar] [PubMed]
- Maxwell, W.; Stojanov, T. Liquid storage of ram semen in the absence or presence of some antioxidants. Reprod. Fertil. Dev. 1996, 8, 1013–1020. [Google Scholar] [CrossRef] [PubMed]
- Peruma, P.; Chamuah, J.K.; Rajkhowa, C. Effect of catalase on the liquid storage of mithun (Bos frontalis) semen. Asian Pac. J. Reprod. 2013, 2, 209–214. [Google Scholar] [CrossRef]
- Michael, A.J.; Alexopoulos, C.; Pontiki, E.A.; Hadjipavlou-Litina, D.J.; Saratsis, P.; Ververidis, H.N.; Boscos, C.M. Effect of antioxidant supplementation in semen extenders on semen quality and reactive oxygen species of chilled canine spermatozoa. Anim. Reprod. Sci. 2009, 112, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Barranco, I.; Tvarijonaviciute, A.; Perez-Patiño, C.; Alkmin, D.V.; Ceron, J.J.; Martinez, E.A.; Rodriguez-Martinez, H.; Roca, J. The activity of paraoxonase type 1 (PON-1) in boar seminal plasma and its relationship with sperm quality, functionality, and in vivo fertility. Andrology 2015, 3, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Verit, F.F.; Verit, A.; Ciftci, H.; Erel, O.; Celik, H. Paraoxonase-1 activity in subfertile men and relationship to sperm parameters. J. Androl. 2009, 30, 183–189. [Google Scholar] [CrossRef]
- Camps, J.; Marsillach, J.; Joven, J. The paraoxonases: Role in human diseases and methodological difficulties in measurement. Crit. Rev. Clin. Lab. Sci. 2009, 46, 83–106. [Google Scholar] [CrossRef]
- Mackness, M.I.; Arrol, S.; Abbott, C.; Durrington, P.N. Protection of low-density lipoprotein against oxidative modification by high-density lipoprotein associated paraoxonase. Atherosclerosis 1993, 104, 129–135. [Google Scholar] [CrossRef]
- Aviram, M.; Rosenblat, M.; Billecke, S.; Erogul, J.; Sorenson, R.; Bisgaier, C.L.; Newton, R.S.; La Du, B. Human serum paraoxonase (PON 1) is inactivated by oxidized low density lipoprotein and preserved by antioxidants. Free Radic. Biol. Med. 1999, 26, 892–904. [Google Scholar] [CrossRef]
- Lazaros, L.A.; Xita, N.V.; Hatzi, E.G.; Kaponis, A.I.; Stefos, T.J.; Plachouras, N.I.; Makrydimas, G.V.; Sofikitis, N.V.; Zikopoulos, K.A.; Georgiou, I.A. Association of paraoxonase gene polymorphisms with sperm parameters. J. Androl. 2011, 32, 394–401. [Google Scholar] [CrossRef]
- Cecchini, G.S.; Giangaspero, B.; Di Palma, T.; Macchia, G.; Carluccio, A.; Boni, R. Oxidative profile and protease regulator potential to predict sperm functionality in donkey (Equus asinus). Sci. Rep. 2021, 11, 20551. [Google Scholar] [CrossRef]
- Rubio, C.P.; Martínez-Subiela, S.; Hernández-Ruiz, J.; Tvarijonaviciute, A.; Cerón, J.J.; Allenspach, K. Serum biomarkers of oxidative stress in dogs with idiopathic inflammatory bowel disease. Vet. J. 2017, 221, 56–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niki, E. Assessment of antioxidant capacity of natural products. Curr. Pharm. Biotechnol. 2010, 11, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Bathgate, R. Antioxidant mechanisms and their benefit on post-thaw boar sperm quality. Reprod. Domest. Anim. 2011, 46, 23–25. [Google Scholar] [CrossRef]
- López-Rodriguez, G.; Suárez-Dieguez, T. Albumin and transferrin are antioxidants that prevent lipoperoxidation in vitro. Rev. Latinoam. Quím. 2010, 38, 159–167. [Google Scholar]
- Roche, M.; Rondeau, P.; Singh, N.R.; Tarnus, E.; Bourdon, E. The antioxidant properties of serum albumin. FEBS Lett. 2008, 582, 1783–1787. [Google Scholar] [CrossRef] [PubMed]
- Bansal, A.K.; Bilaspuri, G.S. Impacts of oxidative stress and antioxidants on semen functions. Vet. Med. Int. 2011, 2011, 686137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abuelo, A.; Hernández, J.; Benedito, J.L.; Castillo, C. Oxidative stress index (OSi) as a new tool to assess redox status in dairy cattle during the transition period. Animal 2013, 7, 1374–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barranco, I.; Rubio, C.P.; Tvarijonaviciute, A.; Rodriguez-Martinez, H.; Roca, J. Measurement of oxidative stress index in seminal plasma can predict in vivo fertility of liquid-stored porcine artificial insemination semen doses. Antioxidants 2021, 10, 1203. [Google Scholar] [CrossRef]
- Contri, A.; Gloria, A.; Robbe, D.; Sfirro, M.P.; Carluccio, A. Effect of sperm concentration on characteristics of frozen-thawed semen in donkeys. Anim. Reprod. Sci. 2012, 136, 74–80. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Catalán, J.; Yánez-Ortiz, I.; Tvarijonaviciute, A.; González-Arostegui, L.G.; Rubio, C.P.; Yeste, M.; Miró, J.; Barranco, I. Impact of Seminal Plasma Antioxidants on Donkey Sperm Cryotolerance. Antioxidants 2022, 11, 417. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11020417
Catalán J, Yánez-Ortiz I, Tvarijonaviciute A, González-Arostegui LG, Rubio CP, Yeste M, Miró J, Barranco I. Impact of Seminal Plasma Antioxidants on Donkey Sperm Cryotolerance. Antioxidants. 2022; 11(2):417. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11020417
Chicago/Turabian StyleCatalán, Jaime, Iván Yánez-Ortiz, Asta Tvarijonaviciute, Luis Guillermo González-Arostegui, Camila P. Rubio, Marc Yeste, Jordi Miró, and Isabel Barranco. 2022. "Impact of Seminal Plasma Antioxidants on Donkey Sperm Cryotolerance" Antioxidants 11, no. 2: 417. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11020417