
Vasoactive Properties of a Cocoa Shell Extract: Mechanism of Action and Effect on Endothelial Dysfunction in Aged Rats


 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. CSE Extraction and Characterization
2.2. Experimental Animals
2.3. Assessment of Vascular Function
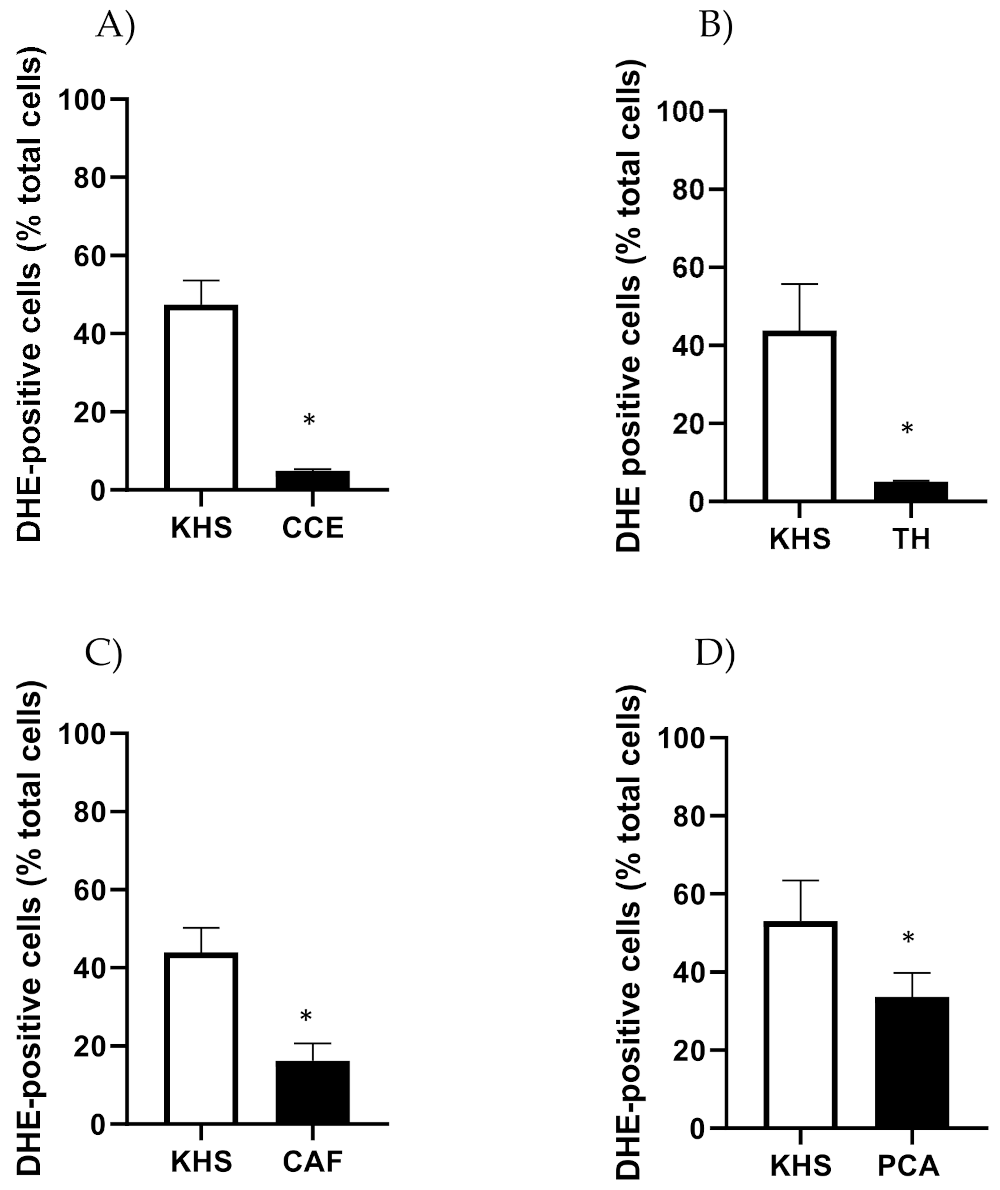
2.4. Determination of Basal Superoxide Anion Levels
2.5. Statistical Analysis
3. Results
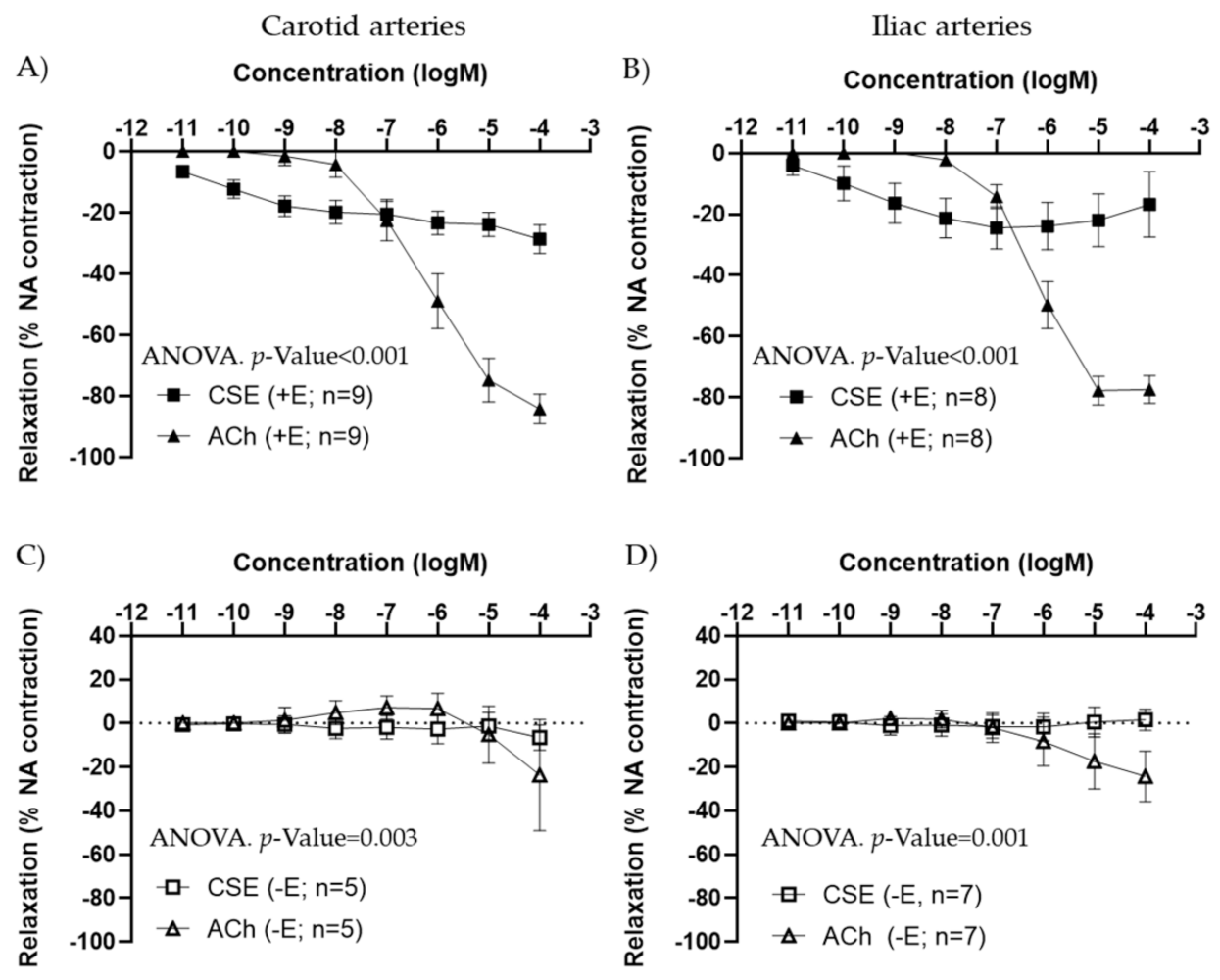
3.1. Vasoactive Effects of CSE
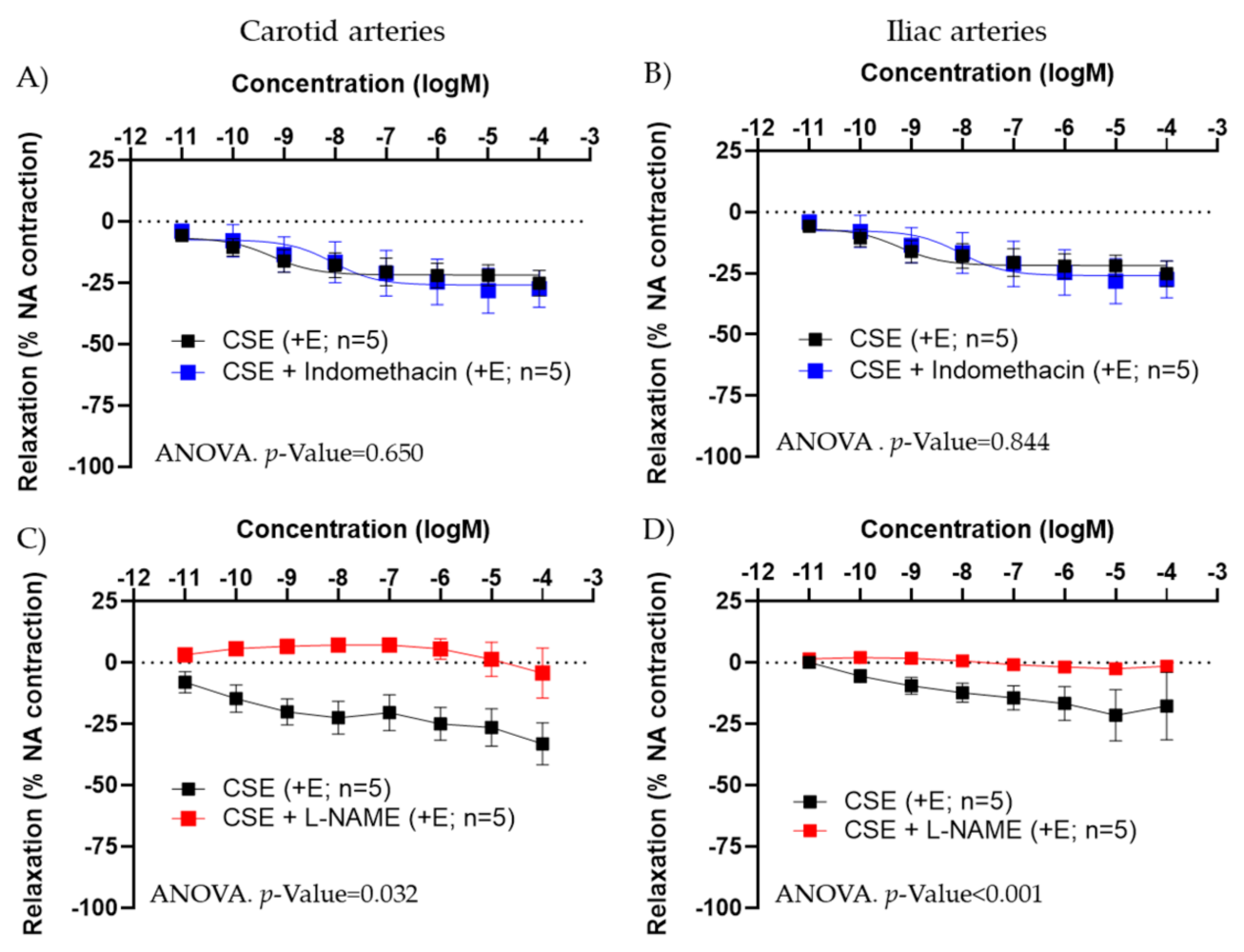
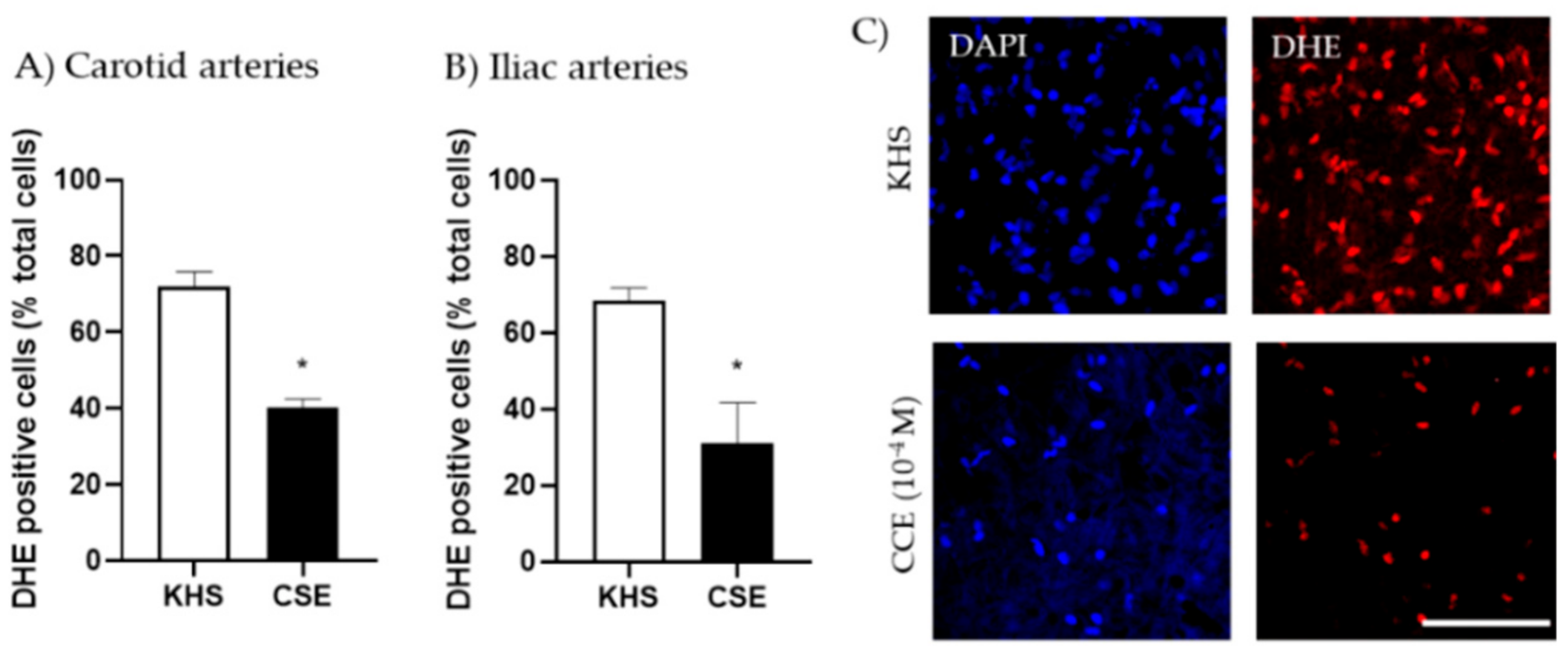
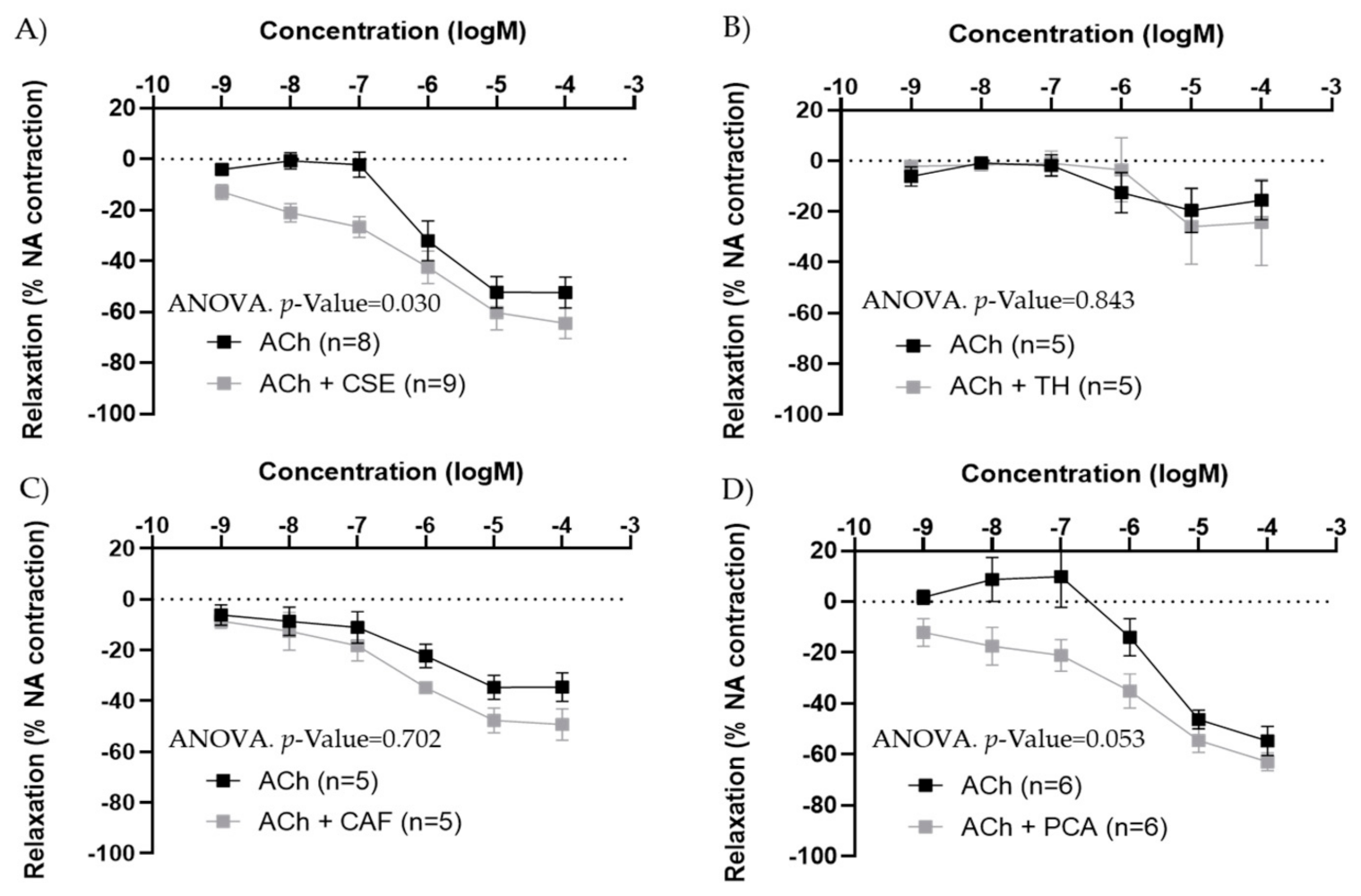
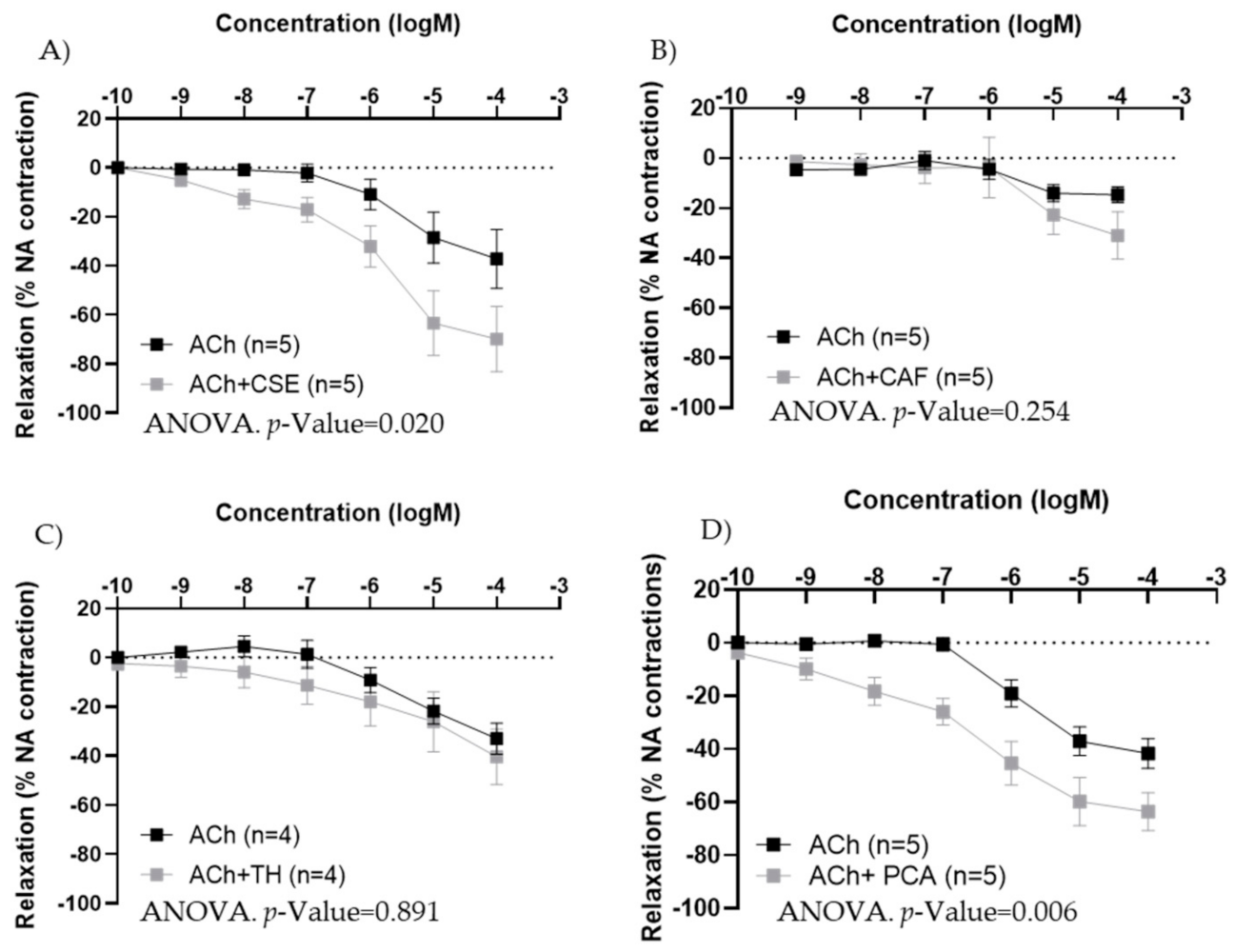
3.2. Mechanism of Action of CSE
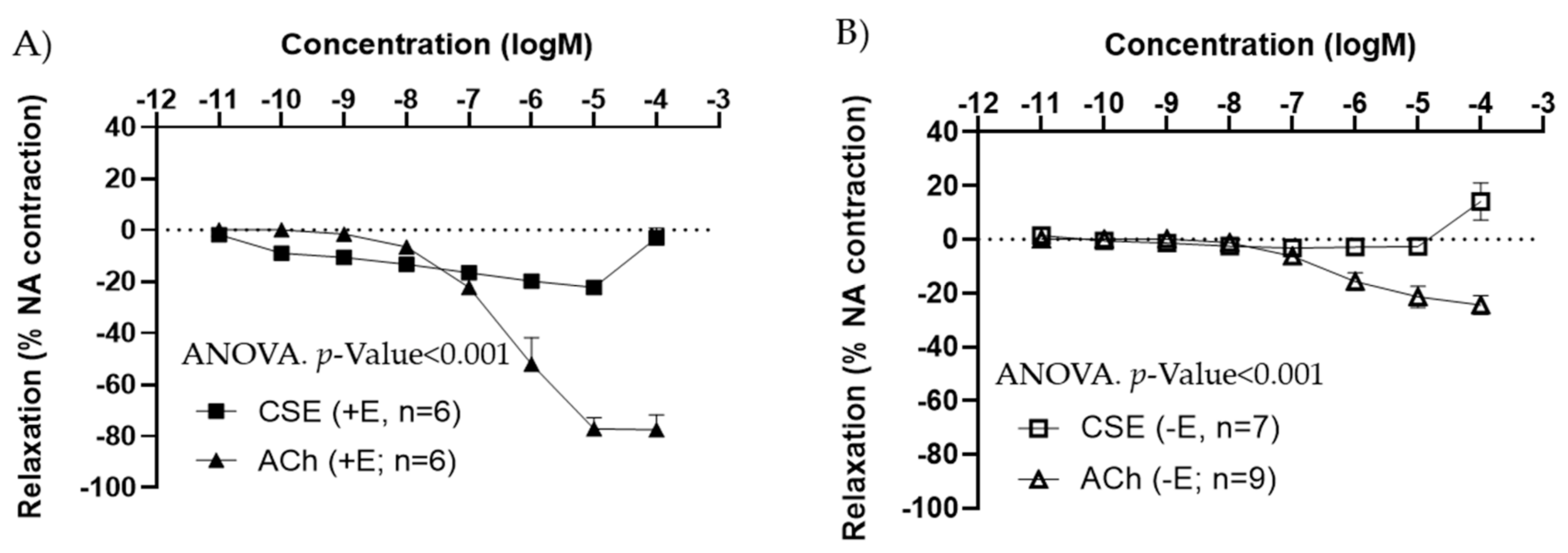
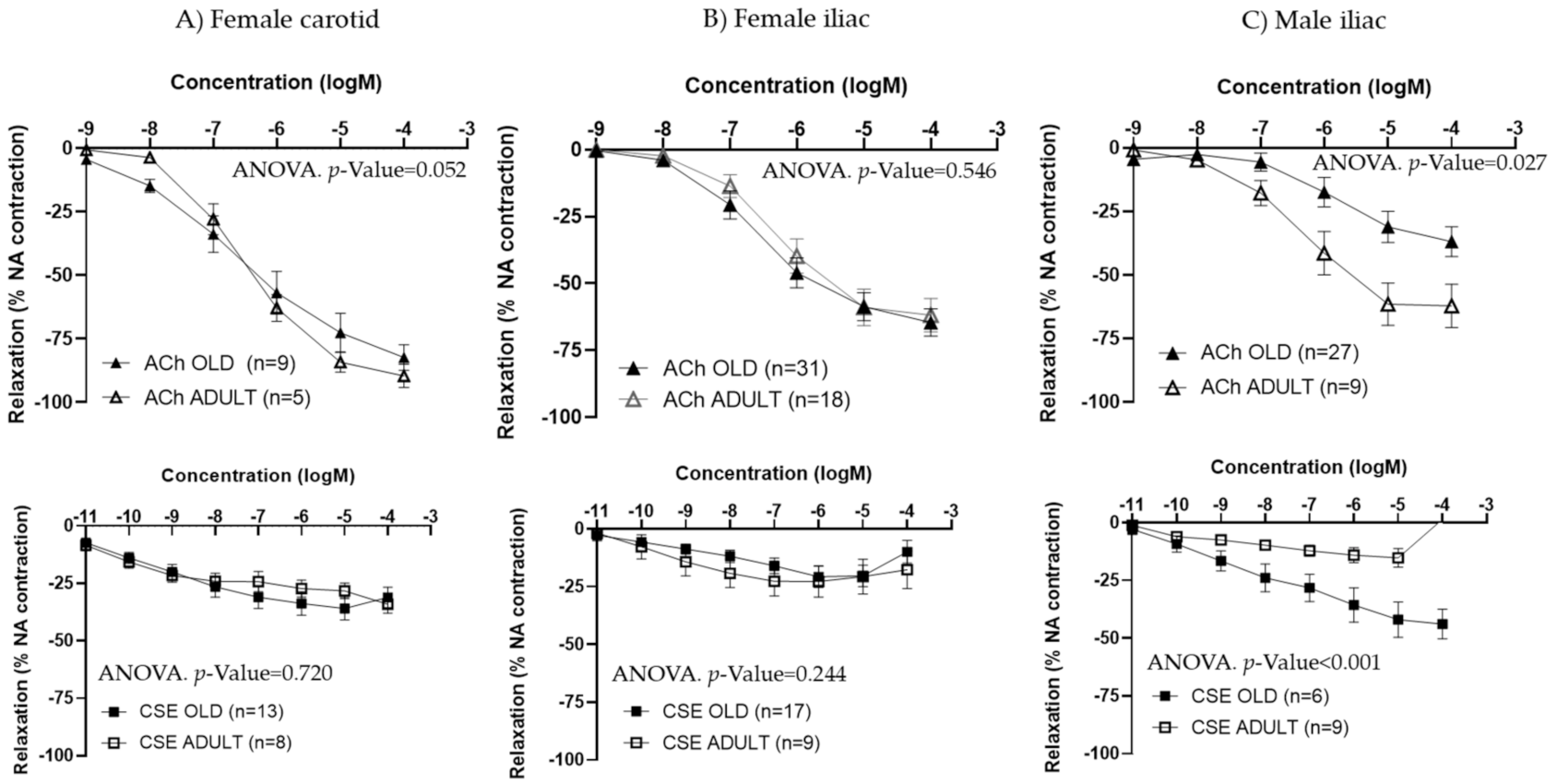
3.3. Effect of CSE on Aged Rats
3.4. Effect of the Major Bioactive Components of CSE on Aged Male and Female Rats
4. Discussion
5. Conclusions
- An aqueous extract derived from the cocoa shell has vasodilatory properties in different vessels, age ranges, and both sexes.
- CSE vasodilatory actions are, at least in part, related to its antioxidant properties, protecting NO from superoxide anion degradation.
- CSE has a higher potency compared to classical endothelium-dependent vasodilators, relaxing arteries at very low concentrations.
- CSE vasodilatory actions are more prominent in arteries with endothelial dysfunction, also improving vasodilatation induced by other agents such as acetylcholine.
- Our results indicate the possible use of this extract as a dietary supplement to improve endothelial dysfunction targeting aging and CVD.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P.; et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990–2019: Update from the GBD 2019 Study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef] [PubMed]
- Konukoglu, D.; Uzun, H. Endothelial Dysfunction and Hypertension. Adv. Exp. Med. Biol. 2017, 956, 511–540. [Google Scholar] [PubMed]
- McCarron, J.G.; Lee, M.D.; Wilson, C. The Endothelium Solves Problems that Endothelial Cells do Not Know Exist. Trends Pharmacol. Sci. 2017, 38, 322–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.J.; Wu, Z.Y.; Nie, X.W.; Bian, J.S. Role of Endothelial Dysfunction in Cardiovascular Diseases: The Link between Inflammation and Hydrogen Sulfide. Front. Pharmacol. 2020, 10, 1568. [Google Scholar] [CrossRef] [Green Version]
- Widmer, R.J.; Lerman, A. Endothelial Dysfunction and Cardiovascular Disease. Glob. Cardiol. Sci. Pract. 2014, 2014, 291–308. [Google Scholar] [CrossRef]
- Rodgers, J.L.; Jones, J.; Bolleddu, S.I.; Vanthenapalli, S.; Rodgers, L.E.; Shah, K.; Karia, K.; Panguluri, S.K. Cardiovascular Risks Associated with Gender and Aging. J. Cardiovasc. Dev. Dis. 2019, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chun, O.K.; Song, W.O. Plasma and Dietary Antioxidant Status as Cardiovascular Disease Risk Factors: A Review of Human Studies. Nutrients 2013, 5, 2969–3004. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.A.; Alam, F.; Solayman, M.; Khalil, M.I.; Kamal, M.A.; Gan, S.H. Dietary Phytochemicals: Natural Swords Combating Inflammation and Oxidation-Mediated Degenerative Diseases. Oxidative Med. Cell. Longev. 2016, 2016, 5137431. [Google Scholar] [CrossRef] [Green Version]
- Nasri, H.; Baradaran, A.; Shirzad, H.; Rafieian-Kopaei, M. New Concepts in Nutraceuticals as Alternative for Pharmaceuticals. Int. J. Prev. Med. 2014, 5, 1487–1499. [Google Scholar]
- Santini, A.; Cammarata, S.M.; Capone, G.; Ianaro, A.; Tenore, G.C.; Pani, L.; Novellino, E. Nutraceuticals: Opening the Debate for a Regulatory Framework. Br. J. Clin. Pharmacol. 2018, 84, 659–672. [Google Scholar] [CrossRef] [Green Version]
- Martín, M.A.; Ramos, S. Cocoa Polyphenols in Oxidative Stress: Potential Health Implications. J. Funct. Foods 2016, 27, 570–588. [Google Scholar] [CrossRef] [Green Version]
- Katz, D.L.; Doughty, K.; Ali, A. Cocoa and Chocolate in Human Health and Disease. Antioxid. Redox Signal. 2011, 15, 2779–2811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Zhang, I.; Li, A.; Manson, J.E.; Sesso, H.D.; Wang, L.; Liu, S. Cocoa Flavanol Intake and Biomarkers for Cardiometabolic Health: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. J. Nutr. 2016, 146, 2325–2333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.; Khymenets, O.; Urpi-Sarda, M.; Tulipani, S.; Garcia-Aloy, M.; Monagas, M.; Mora-Cubillos, X.; Llorach, R.; Andres-Lacueva, C. Cocoa Polyphenols and Inflammatory Markers of Cardiovascular Disease. Nutrients 2014, 6, 844–880. [Google Scholar] [CrossRef] [Green Version]
- Beg, M.S.; Ahmad, S.; Jan, K.; Bashir, K. Status, Supply Chain and Processing of Cocoa—A Review. Trends Food Sci. Technol. 2017, 66, 108–116. [Google Scholar] [CrossRef]
- Adamafio, N.A. Theobromine Toxicity and Remediation of Cocoa by-Products: An Overview. J. Biol. Sci. (Faisalabad Pak.) 2013, 13, 570–576. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, H.; Czajkowska, K.; Cichowska, J.; Lenart, A. What’s New in Biopotential of Fruit and Vegetable by-Products Applied in the Food Processing Industry. Trends Food Sci. Technol. 2017, 67, 150–159. [Google Scholar] [CrossRef]
- Felice, F.; Fabiano, A.; De Leo, M.; Piras, A.M.; Beconcini, D.; Cesare, M.M.; Braca, A.; Zambito, Y.; Di Stefano, R. Antioxidant Effect of Cocoa by-Product and Cherry Polyphenol Extracts: A Comparative Study. Antioxidants 2020, 9, 132. [Google Scholar] [CrossRef] [Green Version]
- Rebollo-Hernanz, M.; Zhang, Q.; Aguilera, Y.; Martin-Cabrejas, M.A.; de Mejia, E.G. Relationship of the Phytochemicals from Coffee and Cocoa by-Products with their Potential to Modulate Biomarkers of Metabolic Syndrome in Vitro. Antioxidants 2019, 8, 279. [Google Scholar] [CrossRef] [Green Version]
- Jokic, S.; Gagic, T.; Knez, Z.; Subaric, D.; Skerget, M. Separation of Active Compounds from Food by-Product (Cocoa Shell) using Subcritical Water Extraction. Molecules 2018, 23, 1408. [Google Scholar] [CrossRef] [Green Version]
- Rebollo-Hernanz, M.; Cañas, S.; Taladrid, D.; Segovia, Á.; Bartolomé, B.; Aguilera, Y.; Martín-Cabrejas, M.A. Extraction of Phenolic Compounds from Cocoa Shell: Modeling using Response Surface Methodology and Artificial Neural Networks. Sep. Purif. Technol. 2021, 270, 118779. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Zhang, Q.; Aguilera, Y.; Martin-Cabrejas, M.A.; de Mejia, E.G. Cocoa Shell Aqueous Phenolic Extract Preserves Mitochondrial Function and Insulin Sensitivity by Attenuating Inflammation between Macrophages and Adipocytes in Vitro. Mol. Nutr. Food Res. 2019, 63, e1801413. [Google Scholar] [CrossRef]
- Somoza, B.; Abderrahim, F.; Gonzalez, J.M.; Conde, M.V.; Arribas, S.M.; Starcher, B.; Regadera, J.; Fernandez-Alfonso, M.S.; Diaz-Gil, J.J.; Gonzalez, M.C. Short-Term Treatment of Spontaneously Hypertensive Rats with Liver Growth Factor Reduces Carotid Artery Fibrosis, Improves Vascular Function, and Lowers Blood Pressure. Cardiovasc. Res. 2006, 69, 764–771. [Google Scholar] [CrossRef]
- Gonzalez, J.M.; Somoza, B.; Conde, M.V.; Fernandez-Alfonso, M.S.; Gonzalez, M.C.; Arribas, S.M. Hypertension Increases Middle Cerebral Artery Resting Tone in Spontaneously Hypertensive Rats: Role of Tonic Vasoactive Factor Availability. Clin. Sci. 2008, 114, 651–659. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Rojo-Poveda, O.; Barbosa-Pereira, L.; Zeppa, G.; Stevigny, C. Cocoa Bean Shell-A by-Product with Nutritional Properties and Biofunctional Potential. Nutrients 2020, 12, 1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Vanhoutte, P.M.; Leung, S.W. Vascular Nitric Oxide: Beyond eNOS. J. Pharmacol. Sci. 2015, 129, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Kellogg, D.L.; Zhao, J.L.; Coey, U.; Green, J.V. Acetylcholine-Induced Vasodilation is Mediated by Nitric Oxide and Prostaglandins in Human Skin. J. Appl. Physiol. 2005, 98, 629–632. [Google Scholar] [CrossRef] [Green Version]
- Medow, M.S.; Glover, J.L.; Stewart, J.M. Nitric Oxide and Prostaglandin Inhibition during Acetylcholine-Mediated Cutaneous Vasodilation in Humans. Microcirculation 2008, 15, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Ponte, A.; Marin, J.; Arribas, S.; Gonzalez, R.; Barrus, M.T.; Salaices, M.; Sanchez-Ferrer, C.F. Endothelial Modulation of Ouabain-Induced Contraction and Sodium Pump Activity in Aortas of Normotensive Wistar-Kyoto and Spontaneously Hypertensive Rats. J. Vasc. Res. 1996, 33, 164–174. [Google Scholar] [CrossRef]
- Kelm, M. Nitric Oxide Metabolism and Breakdown. Biochim. Biophys. Acta 1999, 1411, 273–289. [Google Scholar] [CrossRef] [Green Version]
- Vieira-Rocha, M.S.; Rodríguez-Rodríguez, P.; Ferreira-Duarte, M.; Faira, M.; Sousa, J.B.; Morato, M.; Arribas, S.M.; Diniz, C. Fetal Undernutrition Modifies Vascular RAS Balance Enhancing Oxidative Damage and Contributing to Remodeling. Int. J. Mol. Sci. 2022, 23, 1233. [Google Scholar] [CrossRef] [PubMed]
- Siques, P.; de Pablo, Á.L.L.; Brito, J.; Arribas, S.M.; Flores, K.; Arriaza, K.; Naveas, N.; Gonzalez, M.C.; Hoorntje, A.; Leon-Velarde, F.; et al. Nitric Oxide and Superoxide Anion Balance in Rats Exposed to Chronic and Long Term Intermittent Hypoxia. Biomed. Res. Int. 2014, 2014, 610474. [Google Scholar] [CrossRef] [PubMed]
- Cañas, S.; Rebollo-Hernanz, M.; Aguilera, Y.; Benítez, V.; Braojos, C.; Arribas, S.; Martín-Cabrejas, M. Bioaccessibility of Phenolic Compounds from Cocoa Shell Subjected to in Vitro Digestion and its Antioxidant Activity in Intestinal and Hepatic Cells. Med. Sci. Forum 2020, 2, 8612. [Google Scholar] [CrossRef]
- Semaming, Y.; Pannengpetch, P.; Chattipakorn, S.C.; Chattipakorn, N. Pharmacological Properties of Protocatechuic Acid and its Potential Roles as Complementary Medicine. Evid. Based Complement. Altern. Med. 2015, 2015, 593902. [Google Scholar] [CrossRef] [Green Version]
- Echeverri, D.; Montes, F.R.; Cabrera, M.; Galan, A.; Prieto, A. Caffeine’s Vascular Mechanisms of Action. Int. J. Vasc. Med. 2010, 2010, 834060. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Pinilla, E.; Oñatibia-Astibia, A.; Franco, R. The Relevance of Theobromine for the Beneficial Effects of Cocoa Consumption. Front. Pharmacol. 2015, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Edwards, M.; Czank, C.; Woodward, G.M.; Cassidy, A.; Kay, C.D. Phenolic Metabolites of Anthocyanins Modulate Mechanisms of Endothelial Function. J. Agric. Food Chem. 2015, 63, 2423–2431. [Google Scholar] [CrossRef] [Green Version]
- El-Sonbaty, Y.A.; Suddek, G.M.; Megahed, N.; Gameil, N.M. Protocatechuic Acid Exhibits Hepatoprotective, Vasculoprotective, Antioxidant and Insulin-Like Effects in Dexamethasone-Induced Insulin-Resistant Rats. Biochimie 2019, 167, 119–134. [Google Scholar] [CrossRef]
- Luo, J.; Mills, K.; le Cessie, S.; Noordam, R.; van Heemst, D. Ageing, Age-Related Diseases and Oxidative Stress: What to do Next? Ageing Res. Rev. 2020, 57, 100982. [Google Scholar] [CrossRef]
- Gomez-Zamudio, J.H.; Garcia-Macedo, R.; Lazaro-Suarez, M.; Ibarra-Barajas, M.; Kumate, J.; Cruz, M. Vascular Endothelial Function is Improved by Oral Glycine Treatment in Aged Rats. Can. J. Physiol. Pharmacol. 2015, 93, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Stanhewicz, A.E.; Wenner, M.M.; Stachenfeld, N.S. Sex Differences in Endothelial Function Important to Vascular Health and overall Cardiovascular Disease Risk Across the Lifespan. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H1569–H1588. [Google Scholar] [CrossRef] [PubMed]
- Touyz, R.M.; Rios, F.J.; Alves-Lopes, R.; Neves, K.B.; Camargo, L.L.; Montezano, A.C. Oxidative Stress: A Unifying Paradigm in Hypertension. Can. J. Cardiol. 2020, 36, 659–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebollo-Hernanz, M.; Cañas, S.; Aguilera, Y.; Benitez, V.; Gila-Díaz, A.; Rodriguez-Rodriguez, P.; Cobeta, I.M.; de Pablo, A.L.L.; Gonzalez, M.C.; Arribas, S.M.; et al. Validation of Cocoa Shell as a Novel Antioxidant Dietary Fiber Food Ingredient: Nutritional Value, Functional Properties, and Safety. Curr. Dev. Nutr. 2020, 4, 773. [Google Scholar] [CrossRef]
- Wan, M.L.Y.; Co, V.A.; El-Nezami, H. Dietary Polyphenol Impact on Gut Health and Microbiota. Crit. Rev. Food Sci. Nutr. 2021, 61, 690–711. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Female Artery | EC50 ACh | EC50 CSE | p-Value | Max ACh (%) | Max CSE (%) | p-Value |
|---|---|---|---|---|---|---|
| Carotid (n = 9) | 7.46 × 10−7 [5.24 × 10−7; 3.13 × 10−6] | 7.26 × 10−10 [1.98 × 10−10; 1.07 × 10−7] | <0.001 a | −69.71 ± 4.38 | −23.90 ± 3.92 | <0.001 b |
| Iliac (n = 8) | 5.51 × 10−7 [1.96 × 10−7; 1.48 × 10−6] | 4.16 × 10−9 [4.16 × 10−10; 2.44 × 10−6] | 0.045 a | −74.46 ± 4.79 | −21.92 ± 8.67 | <0.001 b |
| Male Artery | EC50 ACh | EC50 CSE | p-Value | Max. Ach (%) | Max. CSE (%) | p-Value |
|---|---|---|---|---|---|---|
| Iliac (n = 6) | 3.04 × 10−7 [1.75 × 10−7; 1.22 × 10−6] | 1.20 × 10−9 [3.67 × 10−11; 9.45 × 10−9] | 0.002 a | −77.22 ± 4.40 | −22.41 ± 2.50 | <0.001 b |
| Artery | Age | EC50 ACh | p-Value | Max ACh (%) | p-Value |
|---|---|---|---|---|---|
| Carotid | Old (n = 9) | 2.08 × 10−7 [1.52 × 10−7; 3.75 × 10−6] | 0.606 | −79.34 [−89.55; −58.29] | 0.307 |
| Adult (n = 5) | 3.31 × 10−7 [1.74 × 10−7; 7.64 × 10−7] | −87.21 [−90.44; −76.51] | |||
| Iliac | Old (n = 31) | 4.92 × 10−7 [8.50 × 10−8; 6.82 × 10−7] | 0.396 | −70.62 [−79.65; −33.93] | 0.886 |
| Adult (n = 18) | 4.93 × 10−7 [2.04 × 10−7; 2.37 × 10−6] | −68.09 [−83.70; −31.95] | |||
| Artery | Age | EC50 CSE | p-Value | Max CSE (%) | p-Value |
| Carotid | Old (n = 13) | 9.37 × 10−10 [2.87 × 10−10; 7.19 × 10−9] | 0.939 | −38.41 [−47.93; −20.23] | 0.307 |
| Adult (n = 8) | 5.96 × 10−10 [1.23 × 10−10; 1.23 × 10−7] | −25.57 [−32.70; −20.41] | |||
| Iliac | Old (n = 15) | 2.70 × 10−9 [3.47 × 10−10; 3.14 × 10−8] | 0.794 | −12.69 [−69.91; −6.04] | 0.879 |
| Adult (n = 9) | 2.15 × 10−9 [2.32 × 10−10; 1.63 × 10−6] | −16.84 [−41.45; −0.13] |
| Artery | Age | EC50 ACh | p-Value | Max ACh (%) | p-Value |
|---|---|---|---|---|---|
| Iliac | Old (n = 27) | 7.56 × 10−7 [1.16 × 10−7; 5.07 × 10−6] | 0.295 | −40.10 [−51.34; −18.00] | 0.016 |
| Adult (n = 9) | 4.47 × 10−7 [1.88 × 10−7; 9.13 × 10−7] | −71.45 [−84.04; −30.63] | |||
| Artery | Age | EC50 CSE | p-Value | Max CSE (%) | p-Value |
| Iliac | Old (n = 6) | 1.20 × 10−8 [2.65 × 10−9; 5.04 × 10−7] | 0.228 | −36.83 [−64.12; −25.27] | 0.005 |
| Adult (n = 8) | 2.40 × 10−9 [6.02 × 10−11; 2.39 × 10−8] | −21.05 [−24.67; −3.77] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Rodríguez, P.; Ragusky, K.; Phuthong, S.; Ruvira, S.; Ramiro-Cortijo, D.; Cañas, S.; Rebollo-Hernanz, M.; Morales, M.D.; López de Pablo, Á.L.; Martín-Cabrejas, M.A.; et al. Vasoactive Properties of a Cocoa Shell Extract: Mechanism of Action and Effect on Endothelial Dysfunction in Aged Rats. Antioxidants 2022, 11, 429. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11020429
Rodríguez-Rodríguez P, Ragusky K, Phuthong S, Ruvira S, Ramiro-Cortijo D, Cañas S, Rebollo-Hernanz M, Morales MD, López de Pablo ÁL, Martín-Cabrejas MA, et al. Vasoactive Properties of a Cocoa Shell Extract: Mechanism of Action and Effect on Endothelial Dysfunction in Aged Rats. Antioxidants. 2022; 11(2):429. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11020429
Chicago/Turabian StyleRodríguez-Rodríguez, Pilar, Kendal Ragusky, Sophida Phuthong, Santiago Ruvira, David Ramiro-Cortijo, Silvia Cañas, Miguel Rebollo-Hernanz, María Dolores Morales, Ángel L. López de Pablo, María A. Martín-Cabrejas, and et al. 2022. "Vasoactive Properties of a Cocoa Shell Extract: Mechanism of Action and Effect on Endothelial Dysfunction in Aged Rats" Antioxidants 11, no. 2: 429. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11020429