
Venetoclax-Resistant MV4-11 Leukemic Cells Activate PI3K/AKT Pathway for Metabolic Reprogramming and Redox Adaptation for Survival

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Venetoclax (ABT199)-Resistant Model
2.3. Cell Viability Analysis
2.4. Apoptosis
2.5. Global Untargeted Metabolomics Profiling Using LC-MS
2.5.1. Metabolite Extraction
2.5.2. HPLC Workflow
2.5.3. Mass Spectrometry
2.5.4. Data Analysis
2.5.5. Data Processing
2.5.6. Metabolomics Pathway Analysis
2.6. Gene Expression Assay
2.7. ROS Assay
2.8. Determination of GSH Levels
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
3.1. ABT199-R Cells Are Highly Resistant to Venetoclax Treatment
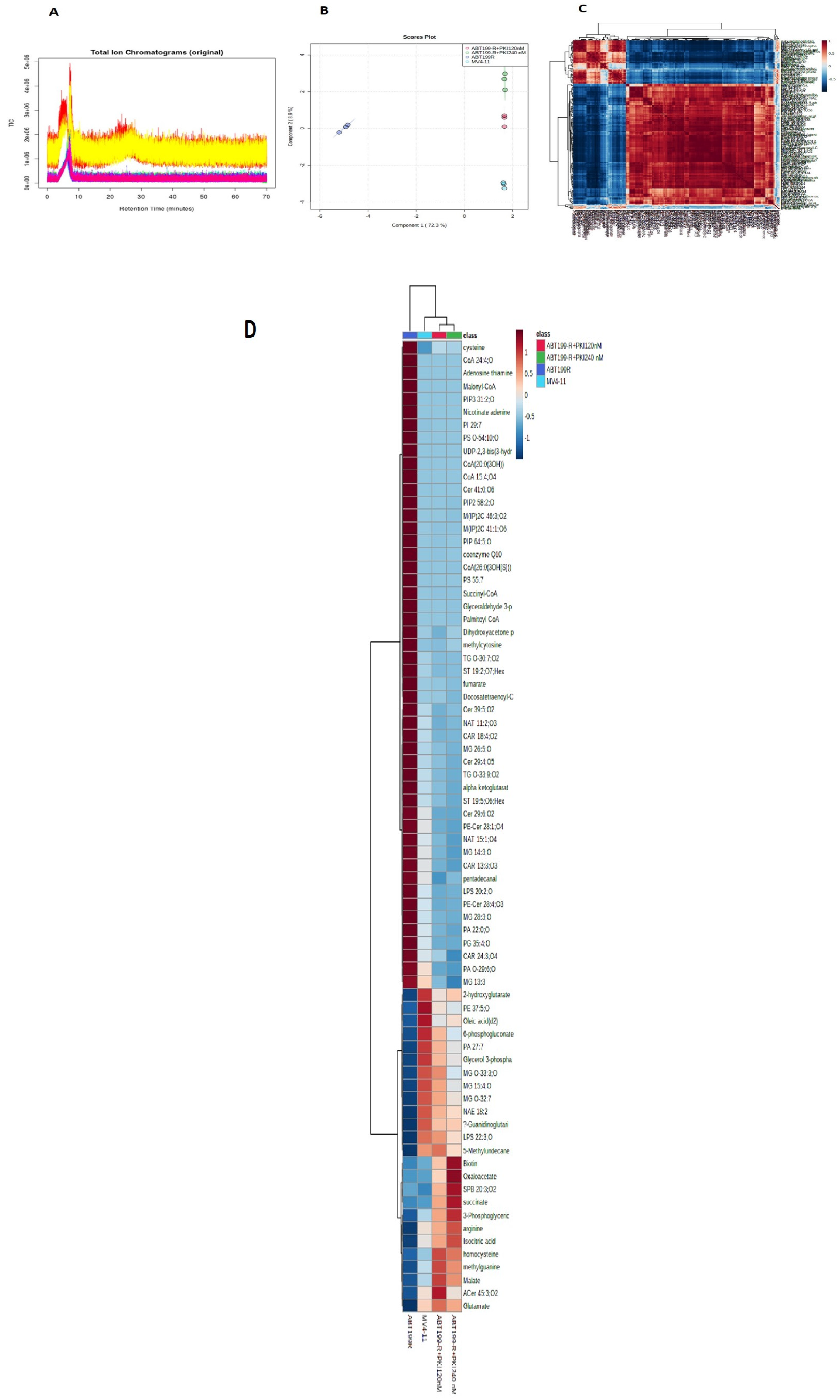
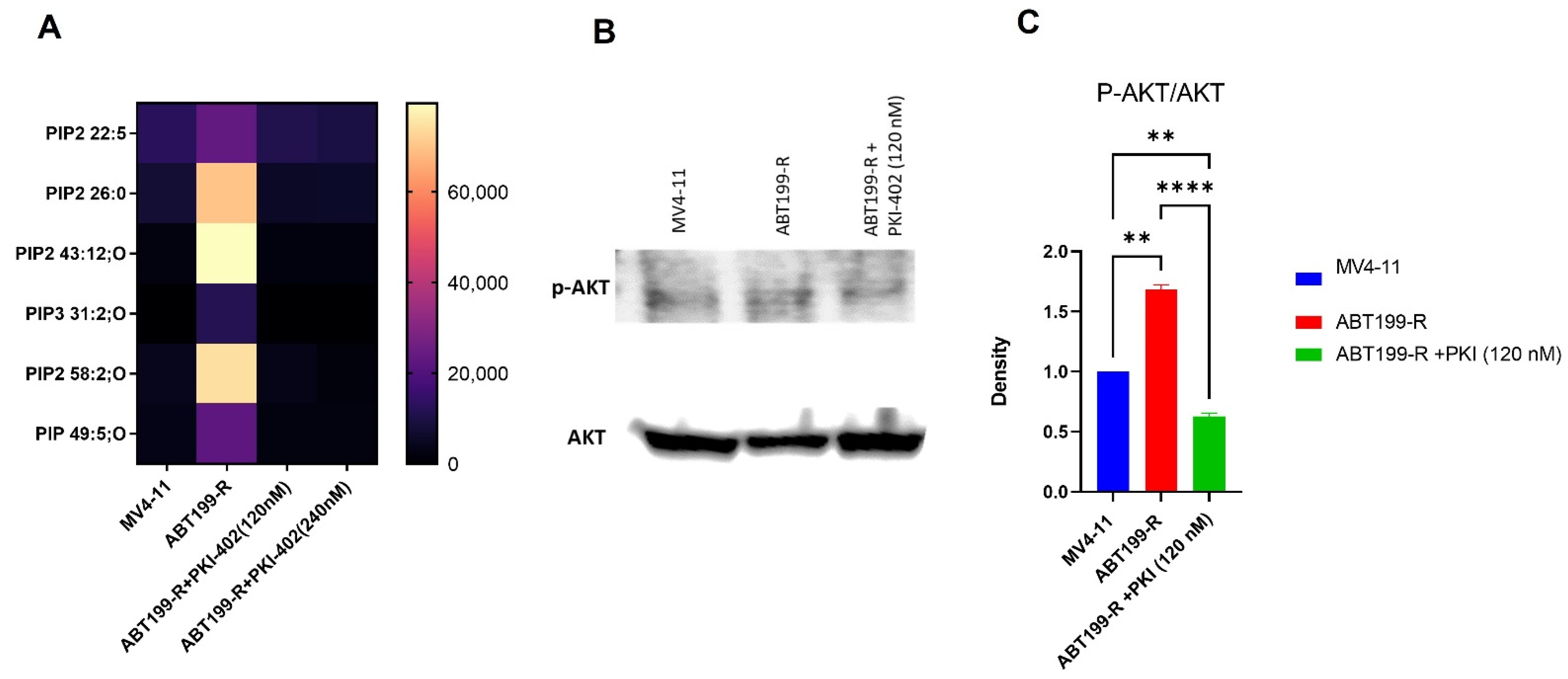
3.2. Metabolomics Showed Accumulation of Metabolites Related to PI3K/AKT Pathway in ABT199-R Cells
3.3. Targeting PI3K/AKT Pathway Shifts the Metabolic Profile of Venetoclax-Resistant Cells Closer to MV4-11
3.4. ABT-199R Cells Maintain Heightened p-AKT Signaling
3.5. ABT199-R Cells Possess High Glycolytic Phenotypes
3.6. ABT199-R Cells Exhibit Decrease in TCA Cycle and OXPHOS Metabolites
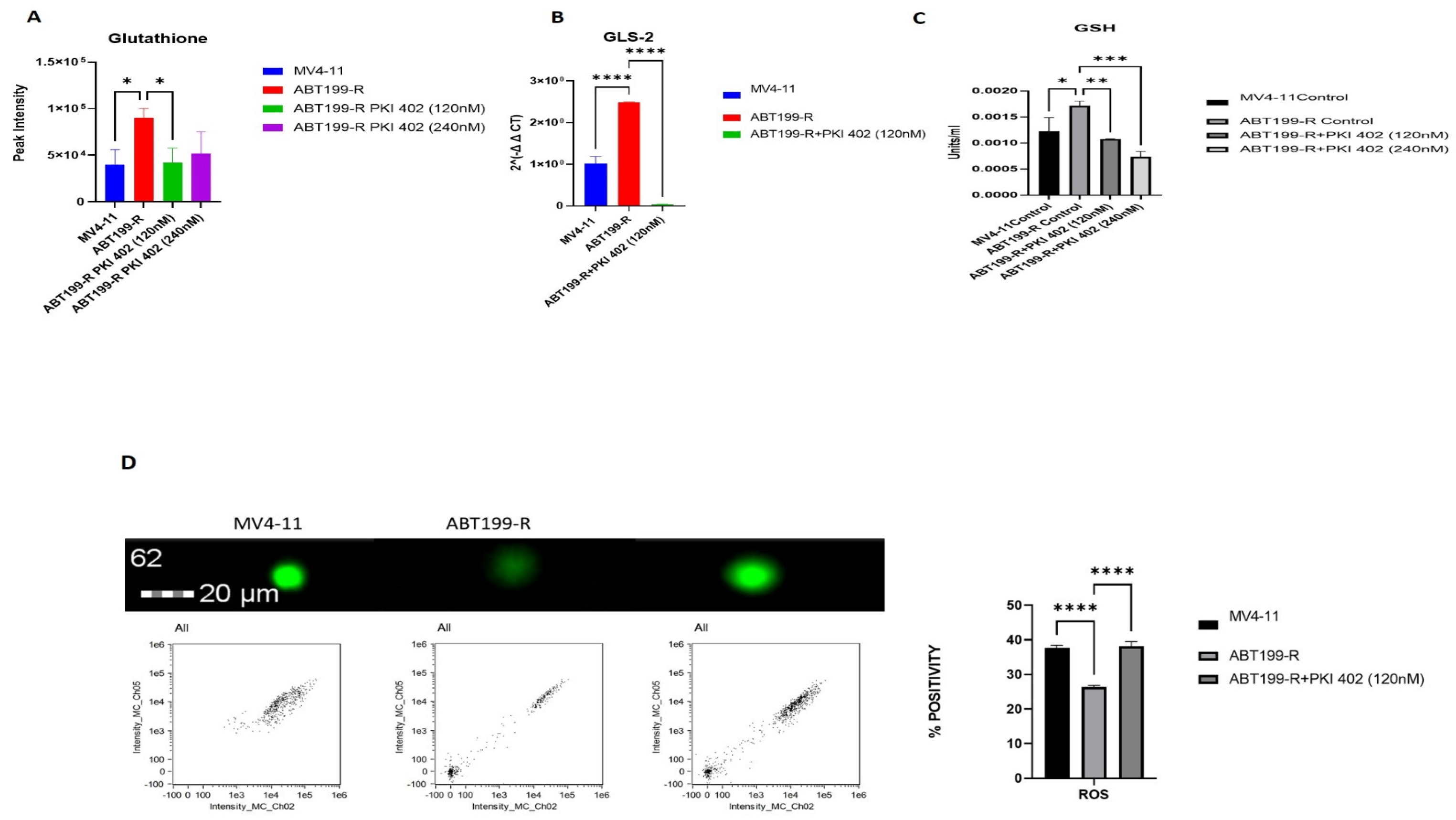
3.7. ABT199-R Cells Possess High Redox Potential
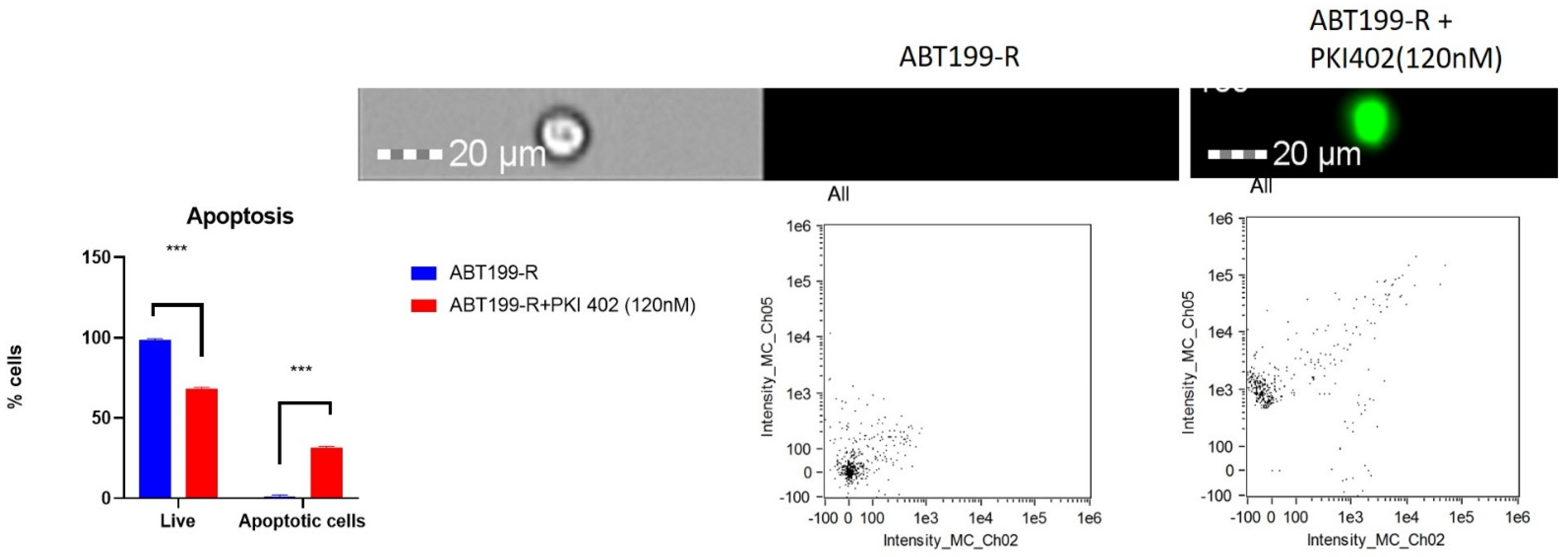
3.8. PI3K/AKT Pathway Inhibition Promotes Apoptosis in ABT199-R Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mehta, S.V.; Shukla, S.N.; Vora, H.H. Overexpression of Bcl2 Protein Predicts Chemoresistance in Acute Myeloid Leukemia: Its Correlation with FLT3. Neoplasma 2013, 60, 666–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra, V.A.; DiNardo, C.; Konopleva, M. Venetoclax-Based Therapies for Acute Myeloid Leukemia. Best Pract. Res. Clin. Haematol. 2019, 32, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, L.; Camidge, D.R.; de Oliveira, M.R.; Bonomi, P.; Gandara, D.; Khaira, D.; Hann, C.L.; McKeegan, E.M.; Litvinovich, E.; Hemken, P.M.; et al. Phase I Study of Navitoclax (ABT-263), a Novel Bcl-2 Family Inhibitor, in Patients with Small-Cell Lung Cancer and Other Solid Tumors. J. Clin. Oncol. 2011, 29, 909–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenberg, C.J.; Cory, S. Brief Report LYMPHOID NEOPLASIA ABT-199, a New Bcl-2-Specific BH3 Mimetic, Has in Vivo Efficacy against Aggressive Myc-Driven Mouse Lymphomas without Provoking Thrombocytopenia. Blood J. Am. Soc. Hematol. 2013, 121, 2285–2288. [Google Scholar] [CrossRef]
- Souers, A.J.; Leverson, J.D.; Boghaert, E.R.; Ackler, S.L.; Catron, N.D.; Chen, J.; Dayton, B.D.; Ding, H.; Enschede, S.H.; Fairbrother, W.J.; et al. ABT-199, a Potent and Selective BCL-2 Inhibitor, Achieves Antitumor Activity While Sparing Platelets. Nat. Med. 2013, 19, 202–208. [Google Scholar] [CrossRef]
- Schieber, M.; Ma, S. The Expanding Role of Venetoclax in Chronic Lymphocytic Leukemia and Small Lymphocytic Lymphoma. Blood Lymphat. Cancer Targets Ther. 2019, 9, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Shimony, S.; Stone, R.M.; Stahl, M. Venetoclax Combination Therapy in Acute Myeloid Leukemia and Myelodysplastic Syndromes. Curr. Opin. Hematol. 2021, 29, 63–73. [Google Scholar] [CrossRef]
- Thijssen, R.; Diepstraten, S.T.; Moujalled, D.; Chew, E.; Flensburg, C.; Shi, M.X.; Dengler, M.A.; Litalien, V.; Macraild, S.; Chen, M.; et al. Intact TP-53 Function Is Essential for Sustaining Durable Responses to BH3-Mimetic Drugs in Leukemias. Blood J. Am. Soc. Hematol. 2021, 137, 2721–2735. [Google Scholar] [CrossRef]
- Dinardo, C.D.; Tiong, I.S.; Quaglieri, A.; Macraild, S.; Loghavi, S.; Brown, F.C.; Thijssen, R.; Pomilio, G.; Ivey, A.; Salmon, J.M.; et al. Molecular Patterns of Response and Treatment Failure after Frontline Venetoclax Combinations in Older Patients with AML. Blood 2020, 135, 791–803. [Google Scholar] [CrossRef]
- Chyla, B.; Daver, N.; Doyle, K.; McKeegan, E.; Huang, X.; Ruvolo, V.; Wang, Z.; Chen, K.; Souers, A.; Leverson, J.; et al. Genetic Biomarkers of Sensitivity and Resistance to Venetoclax Monotherapy in Patients with Relapsed Acute Myeloid Leukemia. Am. J. Hematol. 2018, 93, E202–E205. [Google Scholar] [CrossRef] [Green Version]
- Nepstad, I.; Hatfield, K.J.; Grønningsaeter, I.S.; Reikvam, H. Molecular Sciences The PI3K-Akt-mTOR Signaling Pathway in Human Acute Myeloid Leukemia (AML) Cells. Int. J. Mol. Sci. 2020, 21, 2907. [Google Scholar] [CrossRef] [Green Version]
- Piddock, R.E.; Bowles, K.M.; Rushworth, S.A. Cancers The Role of PI3K Isoforms in Regulating Bone Marrow Microenvironment Signaling Focusing on Acute Myeloid Leukemia and Multiple Myeloma. Cancers 2017, 9, 29. [Google Scholar] [CrossRef] [Green Version]
- Roca-Portoles, A.; Rodriguez-Blanco, G.; Sumpton, D.; Cloix, C.; Mullin, M.; Mackay, G.M.; O’Neill, K.; Lemgruber, L.; Luo, X.; Tait, S.W.G. Venetoclax Causes Metabolic Reprogramming Independent of BCL-2 Inhibition. Cell Death Dis. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Arciuch, V.G.A.; Russo, M.A.; Kang, K.S.; di Cristofano, A. Molecular, and Cellular Pathobiology Inhibition of AMPK and Krebs Cycle Gene Expression Drives Metabolic Remodeling of Pten-Deficient Preneoplastic Thyroid Cells. Cancer Res. 2013, 73, 5459–5472. [Google Scholar] [CrossRef] [Green Version]
- Gioran, A.; Nicotera, P.; Bano, D. Impaired Mitochondrial Respiration Promotes Dendritic Branching via the AMPK Signaling Pathway. Cell Death Dis. 2014, 5, e1175. [Google Scholar] [CrossRef] [Green Version]
- Stevens, B.M.; Jones, C.L.; Pollyea, D.A.; Culp-Hill, R.; D’Alessandro, A.; Winters, A.; Krug, A.; Abbott, D.; Goosman, M.; Pei, S.; et al. Fatty Acid Metabolism Underlies Venetoclax Resistance in Acute Myeloid Leukemia Stem Cells. Nat. Cancer 2020, 1, 1176–1187. [Google Scholar] [CrossRef]
- Jones, C.L.; Stevens, B.M.; Pollyea, D.A.; Culp-Hill, R.; Reisz, J.A.; Nemkov, T.; Gehrke, S.; Gamboni, F.; Krug, A.; Winters, A.; et al. Nicotinamide Metabolism Mediates Resistance to Venetoclax in Relapsed Acute Myeloid Leukemia Stem Cells. Cell Stem Cell 2020, 27, 748–764.e4. [Google Scholar] [CrossRef]
- Alghamdi, A.A.; Mohammed, M.R.S.; Zamzami, M.A.; Al-Malki, A.L.; Qari, M.H.; Khan, M.I.; Choudhry, H. Untargeted Metabolomics Identifies Key Metabolic Pathways Altered by Thymoquinone in Leukemic Cancer Cells. Nutrients 2020, 12, 1792. [Google Scholar] [CrossRef]
- Alzahrani, A.M.; Shait Mohammed, M.R.; Alghamdi, R.A.; Ahmad, A.; Zamzami, M.A.; Choudhry, H.; Khan, M.I. Urolithin a and b Alter Cellular Metabolism and Induce Metabolites Associated with Apoptosis in Leukemic Cells. Int. J. Mol. Sci. 2021, 22, 5465. [Google Scholar] [CrossRef]
- Shait Mohammed, M.R.; Alghamdi, R.A.; Alzahrani, A.M.; Zamzami, M.A.; Choudhry, H.; Khan, M.I. Compound C, a Broad Kinase Inhibitor Alters Metabolic Fingerprinting of Extra Cellular Matrix Detached Cancer Cells. Front. Oncol. 2021, 11, 612778. [Google Scholar] [CrossRef]
- Wilson, I.D.; Plumb, R.; Granger, J.; Major, H.; Williams, R.; Lenz, E.M. HPLC-MS-Based Methods for the Study of Metabonomics. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2005, 817, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Tautenhahn, R.; Patti, G.J.; Rinehart, D.; Siuzdak, G. XCMS Online: A Web-Based Platform to Process Untargeted Metabolomic Data. Anal. Chem. 2012, 84, 5035–5039. [Google Scholar] [CrossRef] [Green Version]
- Soltani, A.; Torki, S.; Ghahfarokhi, M.S.; Jami, M.S.; Ghatrehsamani, M. Targeting the Phosphoinositide 3-Kinase/AKT Pathways by Small Molecules and Natural Compounds as a Therapeutic Approach for Breast Cancer Cells. Mol. Biol. Rep. 2019, 46, 4809–4816. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, T.; Du, J.; Li, Y.; Wang, X.; Zhou, Y.; Yu, X.; Fan, W.; Zhu, Q.; Tong, X.; et al. The Critical Role of PTEN/PI3K/AKT Signaling Pathway in Shikonin-Induced Apoptosis and Proliferation Inhibition of Chronic Myeloid Leukemia. Cell. Physiol. Biochem. 2018, 47, 981–993. [Google Scholar] [CrossRef] [PubMed]
- Ediriweera, M.K.; Tennekoon, K.H.; Samarakoon, S.R. Role of the PI3K/AKT/MTOR Signaling Pathway in Ovarian Cancer: Biological and Therapeutic Significance. Semin. Cancer Biol. 2019, 59, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.M.; Zhang, T.; Liu, Y.B.; Deng, S.H.; Han, R.; Liu, T.; Li, J.; Xu, Y. The PAX6-ZEB2 Axis Promotes Metastasis and Cisplatin Resistance in Non-Small Cell Lung Cancer through PI3K/AKT Signaling. Cell Death Dis. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Rahmani, F.; Ziaeemehr, A.; Shahidsales, S.; Gharib, M.; Khazaei, M.; Ferns, G.A.; Ryzhikov, M.; Avan, A.; Hassanian, S.M. Role of Regulatory MiRNAs of the PI3K/AKT/MTOR Signaling in the Pathogenesis of Hepatocellular Carcinoma. J. Cell. Physiol. 2020, 235, 4146–4152. [Google Scholar] [CrossRef]
- Rittler, D.; Baranyi, M.; Molnár, E.; Garay, T.; Jalsovszky, I.; Varga, I.K.; Hegedűs, L.; Aigner, C.; Tóvári, J.; Tímár, J.; et al. The Antitumor Effect of Lipophilic Bisphosphonate BPH1222 in Melanoma Models: The Role of the PI3K/Akt Pathway and the Small G Protein Rheb. Int. J. Mol. Sci. 2019, 20, 4917. [Google Scholar] [CrossRef] [Green Version]
- Sedlic, F.; Sepac, A.; Pravdic, D.; Camara, A.K.S.; Bienengraeber, M.; Brzezinska, A.K.; Wakatsuki, T.; Bosnjak, Z.J. Mitochondrial Depolarization Underlies Delay in Permeability Transition by Preconditioning with Isoflurane: Roles of ROS and Ca2+. Am. J. Physiol. Cell Physiol. 2010, 299, 506–515. [Google Scholar] [CrossRef] [Green Version]
- Sedlic, F.; Seiwerth, F.; Sepac, A.; Sikiric, S.; Cindric, M.; Milavic, M.; Vuletic, L.B.; Jakopovic, M.; Seiwerth, S. Mitochondrial ROS Induce Partial Dedifferentiation of Human Mesothelioma via Upregulation of NANOG. Antioxidants 2020, 9, 606. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Genes | Primers |
|---|---|---|
| 6515 | GLUT-3 | F: 5′-TGCCTTTGGCACTCTCAACCAG-3′ R: 5′-GCCATAGCTCTTCAGACCCAAG-3′ |
| 3099 | HK-2 | F: 5′-GAGTTTGACCTGGATGTGGTTGC-3′ R: 5′-CCTCCATGTAGCAGGCATTGCT-3′ |
| 5211 | PFKL | F: 5′-AAGAAGTAGGCTGGCACGACGT-3′ R: 5′-GCGGATGTTCTCCACAATGGAC-3′ |
| 2026 | ENO-2 | F: 5′-TCATGGTGAGTCATCGCTCAGGAG-3′ R: 5′-ATGTCCGGCAAAGCGAGCTTCATC-3′ |
| 3939 | LDHA | F: 5′-GGATCTCCAACATGGCAGCCTT-3′ R: 5′-AGACGGCTTTCTCCCTCTTGCT-3′ |
| 5163 | PDK-1 | F: 5′-CATGTCACGCTGGGTAATGAGG-3′ R: 5′-CTCAACACGAGGTCTTGGTGCA-3′ |
| 4540 | MT-ND5 | F: 5′-TCACTTCAACCTCCCTCACC-3′ R: 5′-CAGGGAGGTAGCGATGAGAG-3′ |
| 4519 | MT-CYB | F: 5′-ATCACTCGAGACGTAAATTATGGCT-3′ R: 5′-TGAACTAGGTCTGTCCCAATGTATG-3′ |
| 4514 | MT-CO3 | F: 5′-AGTAAGCCTCTACCTGCACG-3′ R: 5′-GAGGAGCGTTATGGAGTGGA-3′ |
| 6175 | RPLP0 | F: 5′-TGGTCATCCAGCAGGTGTTCGA-3′ R: 5′-ACAGACACTGGCAACATTGCGG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alkhatabi, H.A.; Zohny, S.F.; Shait Mohammed, M.R.; Choudhry, H.; Rehan, M.; Ahmad, A.; Ahmed, F.; Khan, M.I. Venetoclax-Resistant MV4-11 Leukemic Cells Activate PI3K/AKT Pathway for Metabolic Reprogramming and Redox Adaptation for Survival. Antioxidants 2022, 11, 461. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11030461
Alkhatabi HA, Zohny SF, Shait Mohammed MR, Choudhry H, Rehan M, Ahmad A, Ahmed F, Khan MI. Venetoclax-Resistant MV4-11 Leukemic Cells Activate PI3K/AKT Pathway for Metabolic Reprogramming and Redox Adaptation for Survival. Antioxidants. 2022; 11(3):461. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11030461
Chicago/Turabian StyleAlkhatabi, Hind A., Samir F. Zohny, Mohammed Razeeth Shait Mohammed, Hani Choudhry, Mohd Rehan, Aamir Ahmad, Farid Ahmed, and Mohammad Imran Khan. 2022. "Venetoclax-Resistant MV4-11 Leukemic Cells Activate PI3K/AKT Pathway for Metabolic Reprogramming and Redox Adaptation for Survival" Antioxidants 11, no. 3: 461. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11030461