
Antioxidant and Antiaging Properties of a Novel Synergistic Nutraceutical Complex: Readouts from an In Cellulo Study and an In Vivo Prospective, Randomized Trial




 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Cellulo Evaluation
2.1.1. Cell Cultures
2.1.2. Real-Time PCR Analysis
Transcriptional Levels of Proteasome Subunits
Relative Telomere Length
2.1.3. Immunoblot Analysis
Oxidized Protein Levels
Proteasome Subunits
2.1.4. DNA Methylation Levels
2.1.5. Enzymatic Activity Assays
Foxo1 Transcriptional Activity Assay
SIRT1 Activity Assay
Proteasome Activity Assay
2.2. In Vivo Evaluation
2.2.1. Clinical Trial Design
2.2.2. Blood Handling, PBMCs and Plasma Isolation
2.2.3. Immunoenzymatic Assays
Determination of 20S Proteasome Content
Oxidized Proteins in Human Plasma
2.3. Statistical Analysis
3. Results
3.1. Formulation of the Administered Nutritional Supplement
3.2. Antiaging and Antioxidant Properties of the Composition Assessed In Cellulo
3.2.1. The Composition Extends the Lifespan of Human Diploid Fibroblasts In Vitro
3.2.2. The Composition Increases the Proteasome Content and Activity and Diminishes the Levels of Oxidized Proteins
3.2.3. The Composition Attenuates Age-Related Telomere Attrition
3.2.4. The Composition Protects from Age-Related Changes in DNA Methylation
3.2.5. The Composition Enhances FoxO1 Transcriptional Activity
3.2.6. The Composition Activates SIRT1
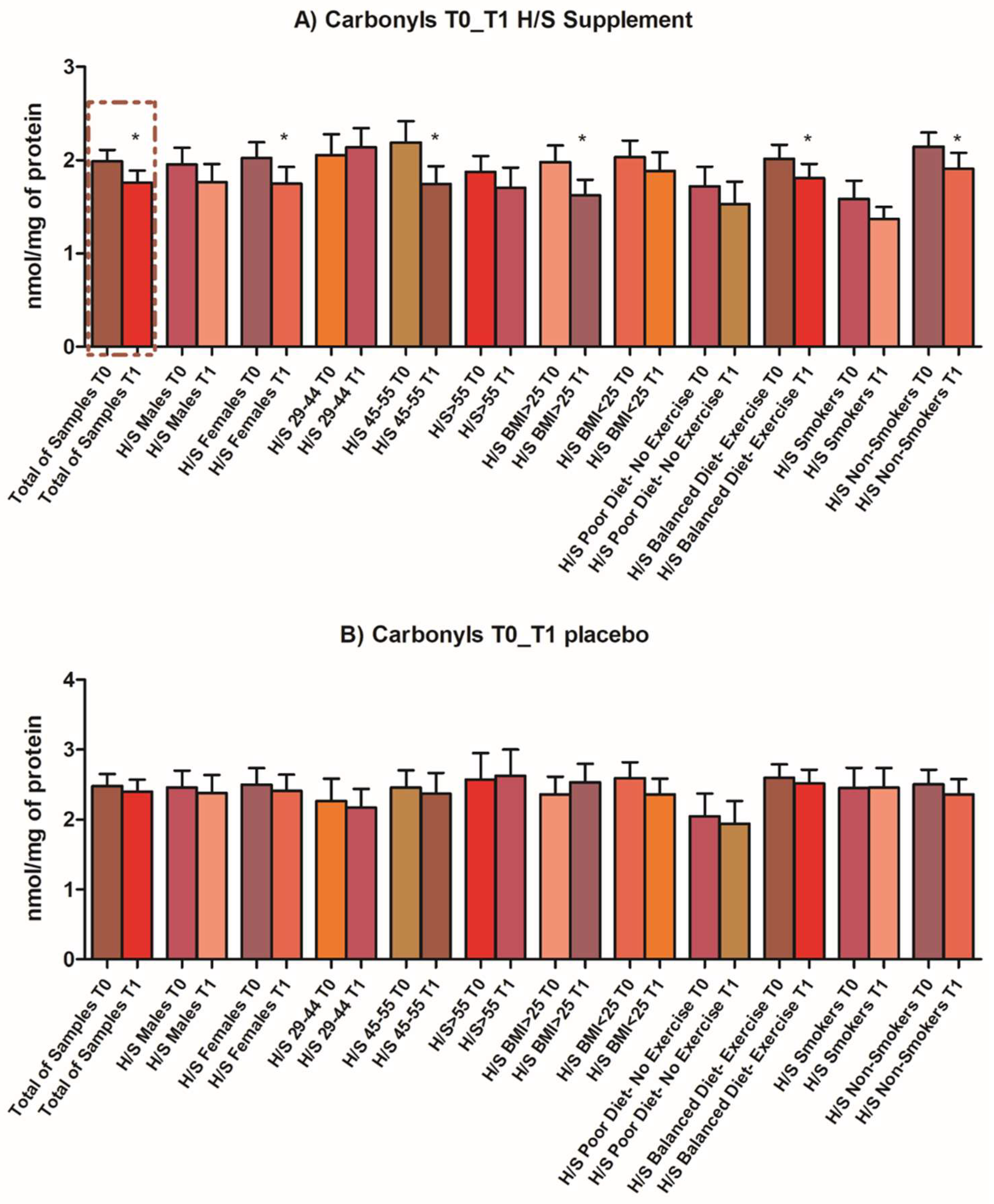
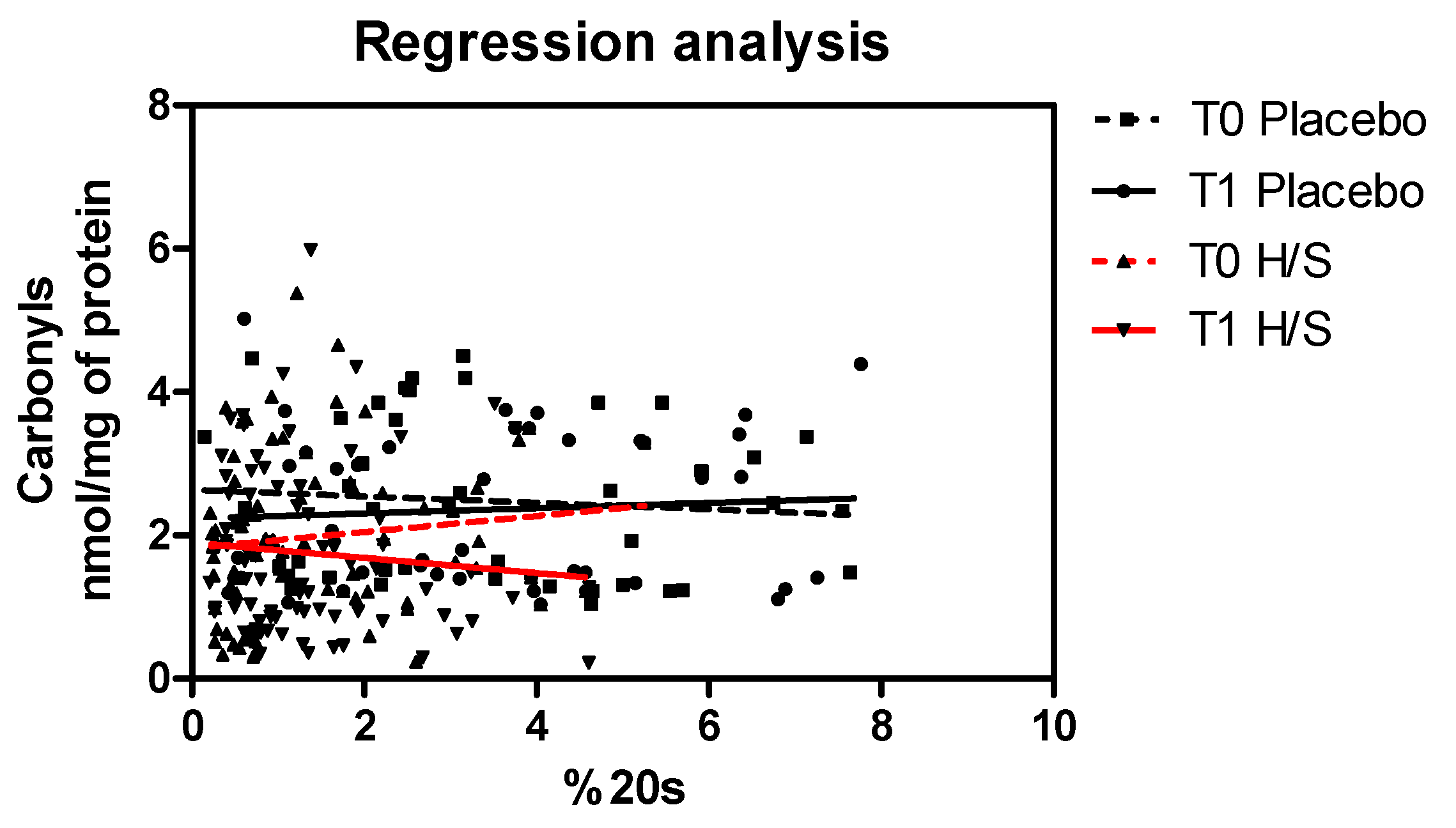
3.3. Antiaging, Antioxidant and Well-Being Properties Assessed In Vivo
4. Discussion
5. Conclusions
6. Patent
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontana, L.; Partridge, L. Promoting health and longevity through diet: From model organisms to humans. Cell 2015, 161, 106–118. [Google Scholar] [CrossRef] [Green Version]
- Chondrogianni, N.; Voutetakis, K.; Kapetanou, M.; Delitsikou, V.; Papaevgeniou, N.; Sakellari, M.; Lefaki, M.; Filippopoulou, K.; Gonos, E.S. Proteasome activation: An innovative promising approach for delaying aging and retarding age-related diseases. Ageing Res. Rev. 2015, 23, 37–55. [Google Scholar] [CrossRef] [PubMed]
- Marshall, R.S.; Vierstra, R.D. Dynamic Regulation of the 26S Proteasome: From Synthesis to Degradation. Front. Mol. Biosci. 2019, 6, 40. [Google Scholar] [CrossRef]
- Voutetakis, K.; Delitsikou, V.; Magouritsas, M.G.; Gonos, E.S. Anti-ageing properties of Khelma Longevity?: Treatment of human fibroblasts increases proteasome levels and decreases the levels of oxidized proteins. N. Biotechnol. 2017, 38, 36–39. [Google Scholar] [CrossRef]
- Athanasopoulou, S.; Chondrogianni, N.; Santoro, A.; Asimaki, K.; Delitsikou, V.; Voutetakis, K.; Fabbri, C.; Pietruszka, B.; Kaluza, J.; Franceschi, C.; et al. Beneficial Effects of Elderly Tailored Mediterranean Diet on the Proteasomal Proteolysis. Front. Physiol. 2018, 9, 457. [Google Scholar] [CrossRef] [Green Version]
- Chondrogianni, N.; Petropoulos, I.; Franceschi, C.; Friguet, B.; Gonos, E.S. Fibroblast cultures from healthy centenarians have an active proteasome. Exp. Gerontol. 2000, 35, 721–728. [Google Scholar] [CrossRef]
- Chondrogianni, N.; de CM Simoes, D.; Franceschi, C.; Gonos, E.S. Cloning of differentially expressed genes in skin fibroblasts from centenarians. Biogerontology 2004, 5, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Chondrogianni, N.; Kapeta, S.; Chinou, I.; Vassilatou, K.; Papassideri, I.; Gonos, E.S. Anti-ageing and rejuvenating effects of quercetin. Exp. Gerontol. 2010, 45, 763–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapetanou, M.; Chondrogianni, N.; Petrakis, S.; Koliakos, G.; Gonos, E.S. Proteasome activation enhances stemness and lifespan of human mesenchymal stem cells. Free Radic. Biol. Med. 2017, 103, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Papaevgeniou, N.; Sakellari, M.; Jha, S.; Tavernarakis, N.; Holmberg, C.I.; Gonos, E.S.; Chondrogianni, N. 18alpha-Glycyrrhetinic Acid Proteasome Activator Decelerates Aging and Alzheimer’s Disease Progression in Caenorhabditis elegans and Neuronal Cultures. Antioxid. Redox Signal. 2016, 25, 855–869. [Google Scholar] [CrossRef] [Green Version]
- Mladenovic Djordjevic, A.N.; Kapetanou, M.; Loncarevic-Vasiljkovic, N.; Todorovic, S.; Athanasopoulou, S.; Jovic, M.; Prvulovic, M.; Taoufik, E.; Matsas, R.; Kanazir, S.; et al. Pharmacological intervention in a transgenic mouse model improves Alzheimer’s-associated pathological phenotype: Involvement of proteasome activation. Free Radic. Biol. Med. 2021, 162, 88–103. [Google Scholar] [CrossRef] [PubMed]
- Kapetanou, M.; Nespital, T.; Tain, L.S.; Pahl, A.; Partridge, L.; Gonos, E.S. FoxO1 is a Novel Regulator of 20S Proteasome Subunits Expression and Activity. Front. Cell Dev. Biol. 2021, 9, 625715. [Google Scholar] [CrossRef] [PubMed]
- Der, G.; Batty, G.D.; Benzeval, M.; Deary, I.J.; Green, M.J.; McGlynn, L.; McIntyre, A.; Robertson, T.; Shiels, P.G. Is telomere length a biomarker for aging: Cross-sectional evidence from the west of Scotland? PLoS ONE 2012, 7, e45166. [Google Scholar] [CrossRef] [PubMed]
- Ellaway, A.; Dundas, R.; Robertson, T.; Shiels, P.G. More miles on the clock: Neighbourhood stressors are associated with telomere length in a longitudinal study. PLoS ONE 2019, 14, e0214380. [Google Scholar] [CrossRef] [PubMed]
- Athanasopoulou, S.; Simos, D.; Charalampopoulou, M.; Tentolouris, N.; Kokkinos, A.; Bacopoulou, F.; Aggelopoulou, E.; Zigkiri, E.; Chrousos, G.P.; Darviri, C.; et al. Significant improvement of stress and aging biomarkers using a novel stress management program with the cognitive restructuring method “Pythagorean Self-Awareness Intervention” in patients with type 2 diabetes mellitus and healthy adults. Mech. Ageing Dev. 2021, 198, 111538. [Google Scholar] [CrossRef] [PubMed]
- Araujo Carvalho, A.C.; Tavares Mendes, M.L.; da Silva Reis, M.C.; Santos, V.S.; Tanajura, D.M.; Martins-Filho, P.R.S. Telomere length and frailty in older adults-A systematic review and meta-analysis. Ageing Res. Rev. 2019, 54, 100914. [Google Scholar] [CrossRef]
- Mensa, E.; Latini, S.; Ramini, D.; Storci, G.; Bonafe, M.; Olivieri, F. The telomere world and aging: Analytical challenges and future perspectives. Ageing Res. Rev. 2019, 50, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Khalangot, M.; Krasnienkov, D.; Vaiserman, A.; Avilov, I.; Kovtun, V.; Okhrimenko, N.; Koliada, A.; Kravchenko, V. Leukocyte telomere length is inversely associated with post-load but not with fasting plasma glucose levels. Exp. Biol. Med. 2017, 242, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Grunnet, L.G.; Pilgaard, K.; Alibegovic, A.; Jensen, C.B.; Hjort, L.; Ozanne, S.E.; Bennett, M.; Vaag, A.; Brøns, C. Leukocyte telomere length is associated with elevated plasma glucose and HbA1c in young healthy men independent of birth weight. Sci. Rep. 2019, 9, 7639. [Google Scholar] [CrossRef]
- Epel, E.S.; Blackburn, E.H.; Lin, J.; Dhabhar, F.S.; Adler, N.E.; Morrow, J.D.; Cawthon, R.M. Accelerated telomere shortening in response to life stress. Proc. Natl. Acad. Sci. USA 2004, 101, 17312–17315. [Google Scholar] [CrossRef] [Green Version]
- Boeck, C.; Salinas-Manrique, J.; Calzia, E.; Radermacher, P.; von Arnim, C.A.F.; Dietrich, D.E.; Kolassa, I.-T.; Karabatsiakis, A. Targeting the association between telomere length and immuno-cellular bioenergetics in female patients with Major Depressive Disorder. Sci. Rep. 2018, 8, 9419. [Google Scholar] [CrossRef] [PubMed]
- Baltzis, D.; Meimeti, E.; Grammatikopoulou, M.G.; Roustit, M.; Mavrogonatou, E.; Kletsas, D.; Efraimidou, S.; Manes, C.; Nikolouzakis, T.K.; Tsiaoussis, J.; et al. Assessment of telomerase activity in leukocytes of type 2 diabetes mellitus patients having or not foot ulcer: Possible correlation with other clinical parameters. Exp. Ther. Med. 2018, 15, 3420–3424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Dong, X.; Cao, L.; Sun, Y.; Qiu, Y.; Zhang, Y.; Cao, R.; Covasa, M.; Zhong, L. Association between telomere length and diabetes mellitus: A meta-analysis. J. Int. Med. Res. 2016, 44, 1156–1173. [Google Scholar] [CrossRef] [PubMed]
- Palacios, J.A.; Herranz, D.; De Bonis, M.L.; Velasco, S.; Serrano, M.; Blasco, M.A. SIRT1 contributes to telomere maintenance and augments global homologous recombination. J. Cell Biol. 2010, 191, 1299–1313. [Google Scholar] [CrossRef] [Green Version]
- Puhlmann, L.M.C.; Valk, S.L.; Engert, V.; Bernhardt, B.C.; Lin, J.; Epel, E.S.; Vrticka, P.; Singer, T. Association of Short-term Change in Leukocyte Telomere Length With Cortical Thickness and Outcomes of Mental Training Among Healthy Adults: A Randomized Clinical Trial. JAMA Netw. Open 2019, 2, e199687. [Google Scholar] [CrossRef]
- Ornish, D.; Lin, J.; Chan, J.M.; Epel, E.; Kemp, C.; Weidner, G.; Marlin, R.; Frenda, S.J.; Magbanua, M.J.M.; Daubenmier, J.; et al. Effect of comprehensive lifestyle changes on telomerase activity and telomere length in men with biopsy-proven low-risk prostate cancer: 5-year follow-up of a descriptive pilot study. Lancet Oncol. 2013, 14, 1112–1120. [Google Scholar] [CrossRef]
- Booth, L.N.; Brunet, A. The Aging Epigenome. Mol. Cell 2016, 62, 728–744. [Google Scholar] [CrossRef] [Green Version]
- Hahn, O.; Grönke, S.; Stubbs, T.M.; Ficz, G.; Hendrich, O.; Krueger, F.; Andrews, S.; Zhang, Q.; Wakelam, M.J.; Beyer, A.; et al. Dietary restriction protects from age-associated DNA methylation and induces epigenetic reprogramming of lipid metabolism. Genome Biol. 2017, 18, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benayoun, B.A.; Pollina, E.A.; Brunet, A. Epigenetic regulation of ageing: Linking environmental inputs to genomic stability. Nat. Rev. Mol. Cell Biol. 2015, 16, 593–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, S.; Raj, K. DNA methylation-based biomarkers and the epigenetic clock theory of ageing. Nat. Rev. Genet. 2018, 19, 371–384. [Google Scholar] [CrossRef]
- Cawthon, R.M. Telomere length measurement by a novel monochrome multiplex quantitative PCR method. Nucleic Acids Res. 2009, 37, e21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgila, K.; Voutetakis, K.; Delitsikou, V.; Chondrogianni, N.; Gonos, E.S. Optimization of in vitro measurement of proteasome activity in mammalian cells using fluorogenic substrates. Free Radic. Biol. Med. 2014, 75 (Suppl. 1), S31. [Google Scholar] [CrossRef] [PubMed]
- Beekman, M.; Blanche, H.; Perola, M.; Hervonen, A.; Bezrukov, V.; Sikora, E.; Flachsbart, F.; Christiansen, L.; De Craen, A.J.M.; Kirkwood, T.B.L.; et al. Genome-wide linkage analysis for human longevity: Genetics of Healthy Aging Study. Aging Cell 2013, 12, 184–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoro, A.; Pini, E.; Scurti, M.; Palmas, G.; Berendsen, A.; Brzozowska, A.; Pietruszka, B.; Szczecinska, A.; Cano, N.; Meunier, N.; et al. Combating inflammaging through a Mediterranean whole diet approach: The NU-AGE project’s conceptual framework and design. Mech. Ageing Dev. 2014, 136–137, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.; Stuetz, W.; Toussaint, O.; Debacq-Chainiaux, F.; Dolle, M.E.T.; Jansen, E.; Gonos, E.S.; Franceschi, C.; Sikora, E.; Hervonen, A.; et al. Associations between Specific Redox Biomarkers and Age in a Large European Cohort: The MARK-AGE Project. Oxid. Med. Cell. Longev. 2017, 2017, 1401452. [Google Scholar] [CrossRef] [PubMed]
- Drouet, S.; Leclerc, E.A.; Garros, L.; Tungmunnithum, D.; Kabra, A.; Abbasi, B.H.; Lainé, É.; Hano, C. A Green Ultrasound-Assisted Extraction Optimization of the Natural Antioxidant and Anti-Aging Flavonolignans from Milk Thistle Silybum marianum (L.) Gaertn. Fruits for Cosmetic Applications. Antioxidants 2019, 8, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filippopoulou, K.; Papaevgeniou, N.; Lefaki, M.; Paraskevopoulou, A.; Biedermann, D.; Křen, V.; Chondrogianni, N. 2,3-Dehydrosilybin A/B as a pro-longevity and anti-aggregation compound. Free Radic. Biol. Med. 2017, 103, 256–267. [Google Scholar] [CrossRef]
- Esposito, T.; Sansone, F.; Russo, P.; Picerno, P.; Aquino, R.P.; Gasparri, F.; Mencherini, T. A Water-Soluble Microencapsulated Milk Thistle Extract as Active Ingredient for Dermal Formulations. Molecules 2019, 24, 1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, J.; Peng, S.; Xu, J.; Fang, J. Reversing ROS-mediated neurotoxicity by chlorogenic acid involves its direct antioxidant activity and activation of Nrf2-ARE signaling pathway. Biofactors 2019, 45, 616–626. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.W.; Boo, Y.C. Siegesbeckiae Herba Extract and Chlorogenic Acid Ameliorate the Death of HaCaT Keratinocytes Exposed to Airborne Particulate Matter by Mitigating Oxidative Stress. Antioxidants 2021, 10, 1762. [Google Scholar] [CrossRef]
- Xanthis, V.; Fitsiou, E.; Voulgaridou, G.-P.; Bogadakis, A.; Chlichlia, K.; Galanis, A.; Pappa, A. Antioxidant and Cytoprotective Potential of the Essential Oil Pistacia lentiscus var. chia and Its Major Components Myrcene and α-Pinene. Antioxidants 2021, 10, 127. [Google Scholar] [CrossRef] [PubMed]
- Vlastos, D.; Mademtzoglou, D.; Drosopoulou, E.; Efthimiou, I.; Chartomatsidou, T.; Pandelidou, C.; Astyrakaki, M.; Chalatsi, E.; Mavragani-Tsipidou, P. Evaluation of the genotoxic and antigenotoxic effects of Chios mastic water by the in vitro micronucleus test on human lymphocytes and the in vivo wing somatic test on Drosophila. PLoS ONE 2013, 8, e69494. [Google Scholar] [CrossRef] [PubMed]
- Drosopoulou, E.; Vlastos, D.; Efthimiou, I.; Kyrizaki, P.; Tsamadou, S.; Anagnostopoulou, M.; Kofidou, D.; Gavriilidis, M.; Mademtzoglou, D.; Mavragani-Tsipidou, P. In vitro and in vivo evaluation of the genotoxic and antigenotoxic potential of the major Chios mastic water constituents. Sci. Rep. 2018, 8, 12200. [Google Scholar] [CrossRef]
- Webb, A.E.; Brunet, A. FOXO transcription factors: Key regulators of cellular quality control. Trends Biochem. Sci. 2014, 39, 159–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabowska, W.; Sikora, E.; Bielak-Zmijewska, A. Sirtuins, a promising target in slowing down the ageing process. Biogerontology 2017, 18, 447–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, G.; Reggiani, F.; Angelini, C.; Finazzi, S.; Astori, E.; Garavaglia, M.L.; Landoni, L.; Portinaro, N.M.; Giustarini, D.; Rossi, R.; et al. Plasma Protein Carbonyls as Biomarkers of Oxidative Stress in Chronic Kidney Disease, Dialysis, and Transplantation. Oxid. Med. Cell. Longev. 2020, 2020, 2975256. [Google Scholar] [CrossRef] [PubMed]
- Frijhoff, J.; Winyard, P.G.; Zarkovic, N.; Davies, S.S.; Stocker, R.; Cheng, D.; Knight, A.R.; Taylor, E.L.; Oettrich, J.; Ruskovska, T.; et al. Clinical Relevance of Biomarkers of Oxidative Stress. Antioxid. Redox Signal. 2015, 23, 1144–1170. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Höhn, A.; Grune, T. The proteasome and the degradation of oxidized proteins: Part II—protein oxidation and proteasomal degradation. Redox Biol. 2014, 2, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Kapetanou, M.; Athanasopoulou, S.; Gonos, E.S. Transcriptional regulatory networks of the proteasome in mammalian systems. IUBMB Life 2022, 74, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Vaiserman, A.; Krasnienkov, D. Telomere Length as a Marker of Biological Age: State-of-the-Art, Open Issues, and Future Perspectives. Front. Genet. 2021, 11, 1816. [Google Scholar] [CrossRef] [PubMed]
- Hudgins, A.D.; Tazearslan, C.; Tare, A.; Zhu, Y.; Huffman, D.; Suh, Y. Age- and Tissue-Specific Expression of Senescence Biomarkers in Mice. Front. Genet. 2018, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Guarasci, F.; D’Aquila, P.; Mandalà, M.; Garasto, S.; Lattanzio, F.; Corsonello, A.; Passarino, G.; Bellizzi, D. Aging and nutrition induce tissue-specific changes on global DNA methylation status in rats. Mech. Ageing Dev. 2018, 174, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; He, X.; Ma, Y.; Zhao, X.; Hou, X.; Hao, E.; Deng, J.; Bai, G. Chlorogenic Acid Targeting of the AKT PH Domain Activates AKT/GSK3β/FOXO1 Signaling and Improves Glucose Metabolism. Nutrients 2018, 10, 1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusku-Kiraz, Z.; Mehmetcik, G.; Dogru-Abbasoglu, S.; Uysal, M. Artichoke leaf extract reduces oxidative stress and lipoprotein dyshomeostasis in rats fed on high cholesterol diet. Phytother. Res. 2010, 24, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Alkushi, A.G. Biological Effect of Cynara cardunculus on Kidney Status of Hypercholesterolemic Rats. Pharmacogn. Mag. 2017, 13, S430–S436. [Google Scholar] [CrossRef]
- Wang, S.; Meckling, K.A.; Marcone, M.F.; Kakuda, Y.; Tsao, R. Synergistic, additive, and antagonistic effects of food mixtures on total antioxidant capacities. J. Agric. Food Chem. 2011, 59, 960–968. [Google Scholar] [CrossRef]
- Caesar, L.K.; Cech, N.B. Synergy and antagonism in natural product extracts: When 1 + 1 does not equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawidowicz, A.L.; Olszowy-Tomczyk, M.; Typek, R. Synergistic and antagonistic antioxidant effects in the binary cannabinoids mixtures. Fitoterapia 2021, 153, 104992. [Google Scholar] [CrossRef] [PubMed]
- Sotler, R.; Poljšak, B.; Dahmane, R.; Jukić, T.; Pavan Jukić, D.; Rotim, C.; Trebše, P.; Starc, A. Prooxidant activities of antioxidants and their impact on health. Acta Clin. Croat. 2019, 58, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Qu, Q.; Kakuda, Y.; Xue, S.; Jiang, Y.; Koide, S.; Shim, Y. Investigation of the antioxidant and synergistic activity of lycopene and other natural antioxidants using LAME and AMVN model systems. J. Food Compos. Anal. 2007, 20, 603–608. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | mg/cap | %/cap | Active Ingredient (mg/cap) |
|---|---|---|---|
| Linum usitatissimum (Seed extract standardized to 40% SDG) | 100 | 21.98 | 40 |
| Silybum marianum (Fruit extract standardized to 50% silymarin) | 50 | 10.99 | 25 |
| Cynara scolymus(Leaf extract standardized to 5% chlorogenic acids) | 100 | 21.98 | 5 |
| Pistacia lentiscus (resinous exudation) standardized to 12% isomasticadienonic acid) | 100 | 21.98 | 12 |
| Magnesium stearate | 10 | 2.20 | |
| Vegetarian Capsule No 0 (hydroxypropyl methylcellulose) | 95 | 20.88 | |
| Content weight | 360 | ||
| Capsule total | 455 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Athanasopoulou, S.; Kapetanou, M.; Magouritsas, M.G.; Mougkolia, N.; Taouxidou, P.; Papacharalambous, M.; Sakellaridis, F.; Gonos, E. Antioxidant and Antiaging Properties of a Novel Synergistic Nutraceutical Complex: Readouts from an In Cellulo Study and an In Vivo Prospective, Randomized Trial. Antioxidants 2022, 11, 468. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11030468
Athanasopoulou S, Kapetanou M, Magouritsas MG, Mougkolia N, Taouxidou P, Papacharalambous M, Sakellaridis F, Gonos E. Antioxidant and Antiaging Properties of a Novel Synergistic Nutraceutical Complex: Readouts from an In Cellulo Study and an In Vivo Prospective, Randomized Trial. Antioxidants. 2022; 11(3):468. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11030468
Chicago/Turabian StyleAthanasopoulou, Sophia, Marianna Kapetanou, Michel Georges Magouritsas, Nikoletta Mougkolia, Polykseni Taouxidou, Michael Papacharalambous, Fotios Sakellaridis, and Efstathios Gonos. 2022. "Antioxidant and Antiaging Properties of a Novel Synergistic Nutraceutical Complex: Readouts from an In Cellulo Study and an In Vivo Prospective, Randomized Trial" Antioxidants 11, no. 3: 468. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11030468