
Effects of High Starch and Supplementation of an Olive Extract on the Growth Performance, Hepatic Antioxidant Capacity and Lipid Metabolism of Largemouth Bass (Micropterus salmoides)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Trial and Sample Collection
2.2. Chemical Composition Analysis of Diets
2.3. Plasma and Hepatic Homogenate Parameters
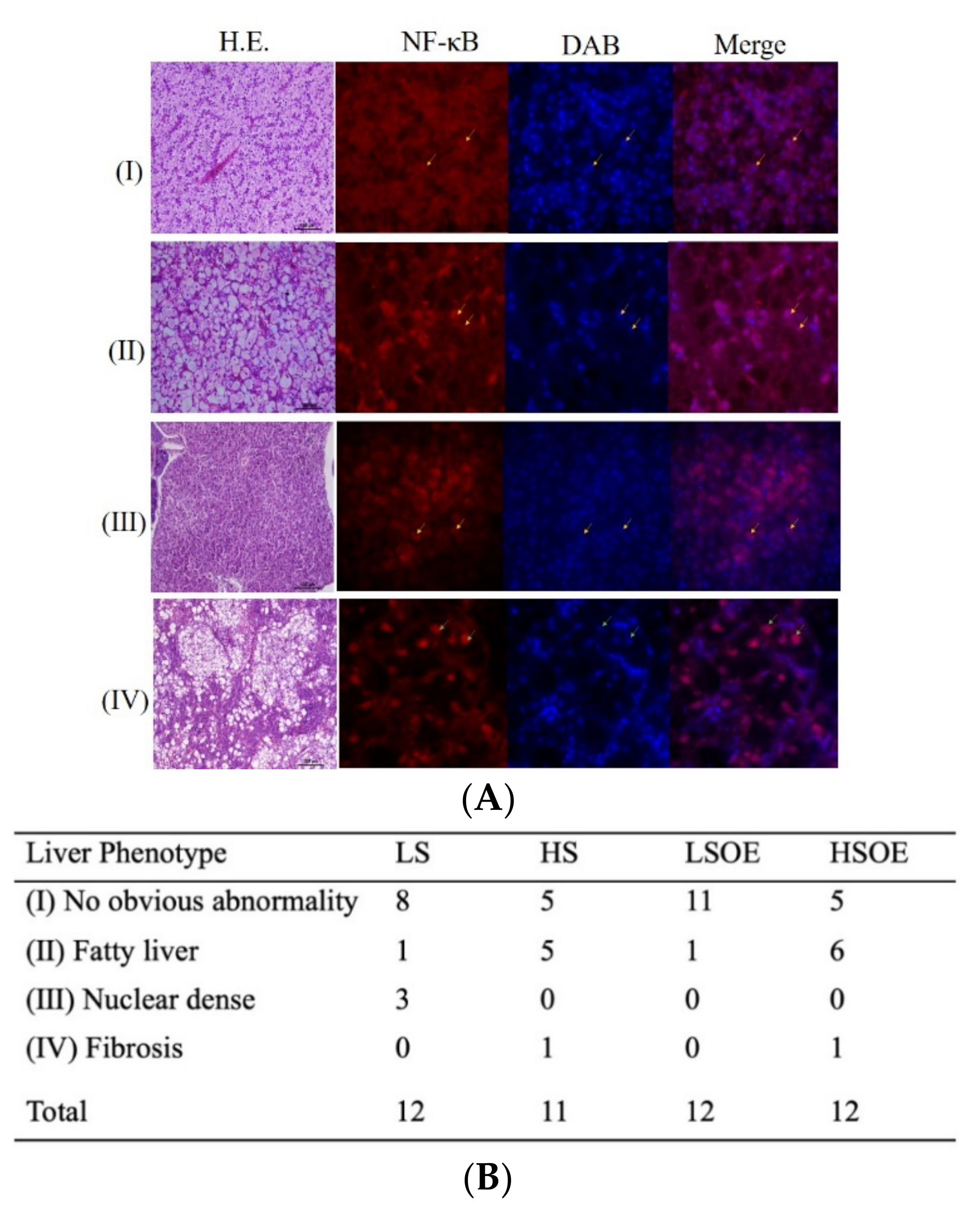
2.4. Hepatic Histopathological and Immunofluorescence Examination
2.5. Quantitative Real-Time PCR
2.6. Western Blot
2.7. Statistical Analysis
3. Results
3.1. Growth Performance and Morphometric Parameters
3.2. Hematological and Liver Functions Parameters
3.3. Hepatic Antioxidant Responses
3.4. Hepatic Pathological Examination—Histology
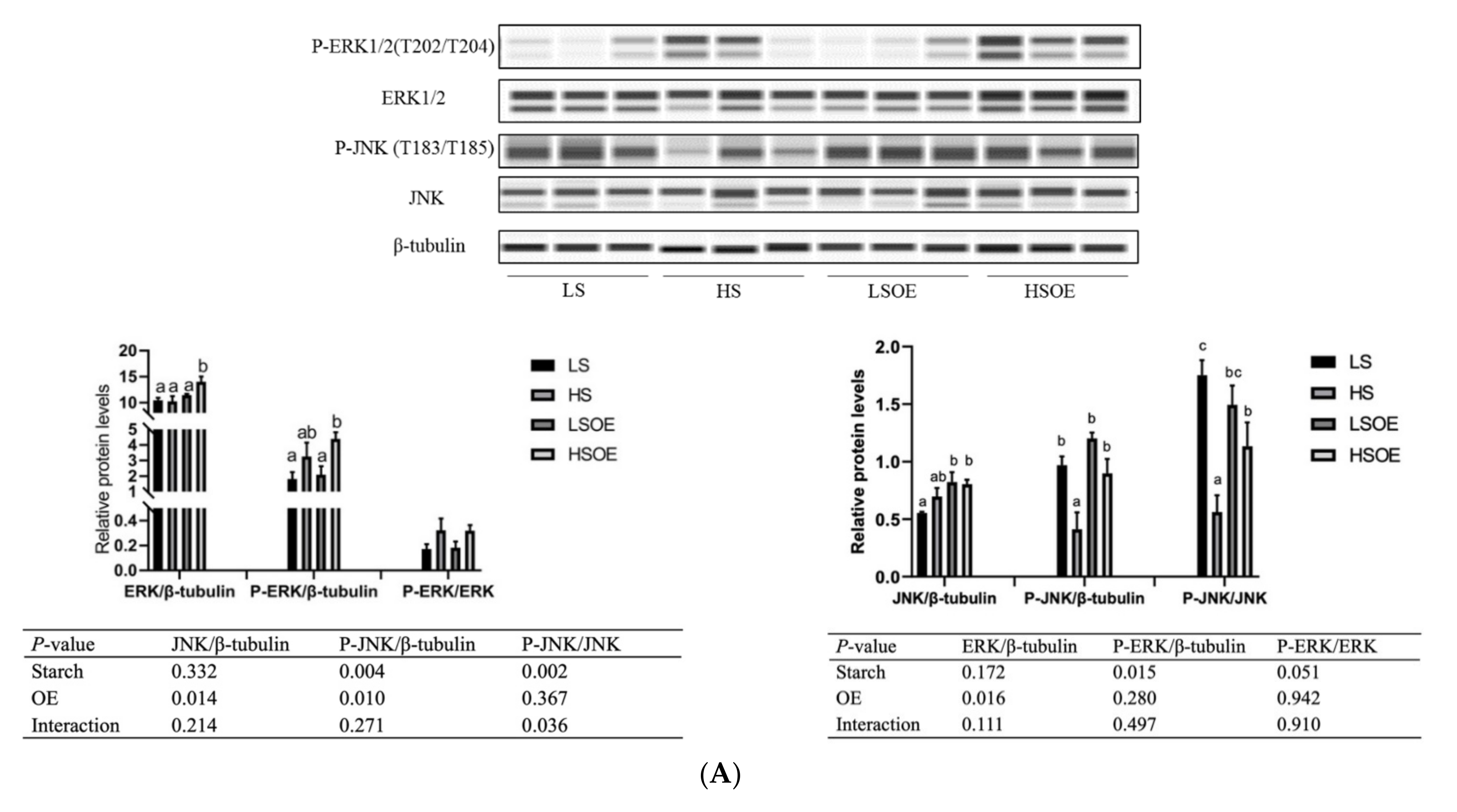
3.5. Hepatic Proliferation and Inflammation Responses
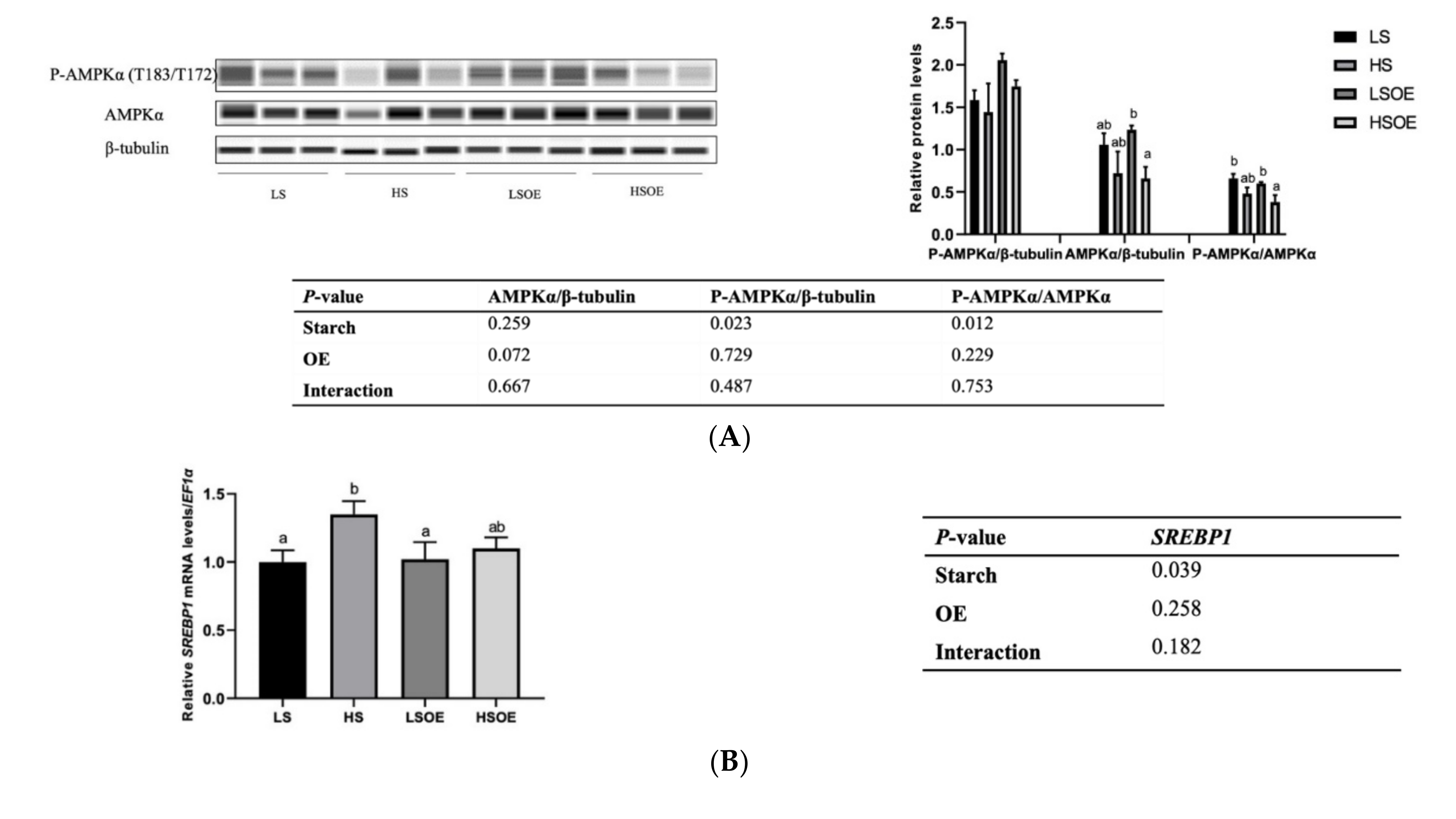
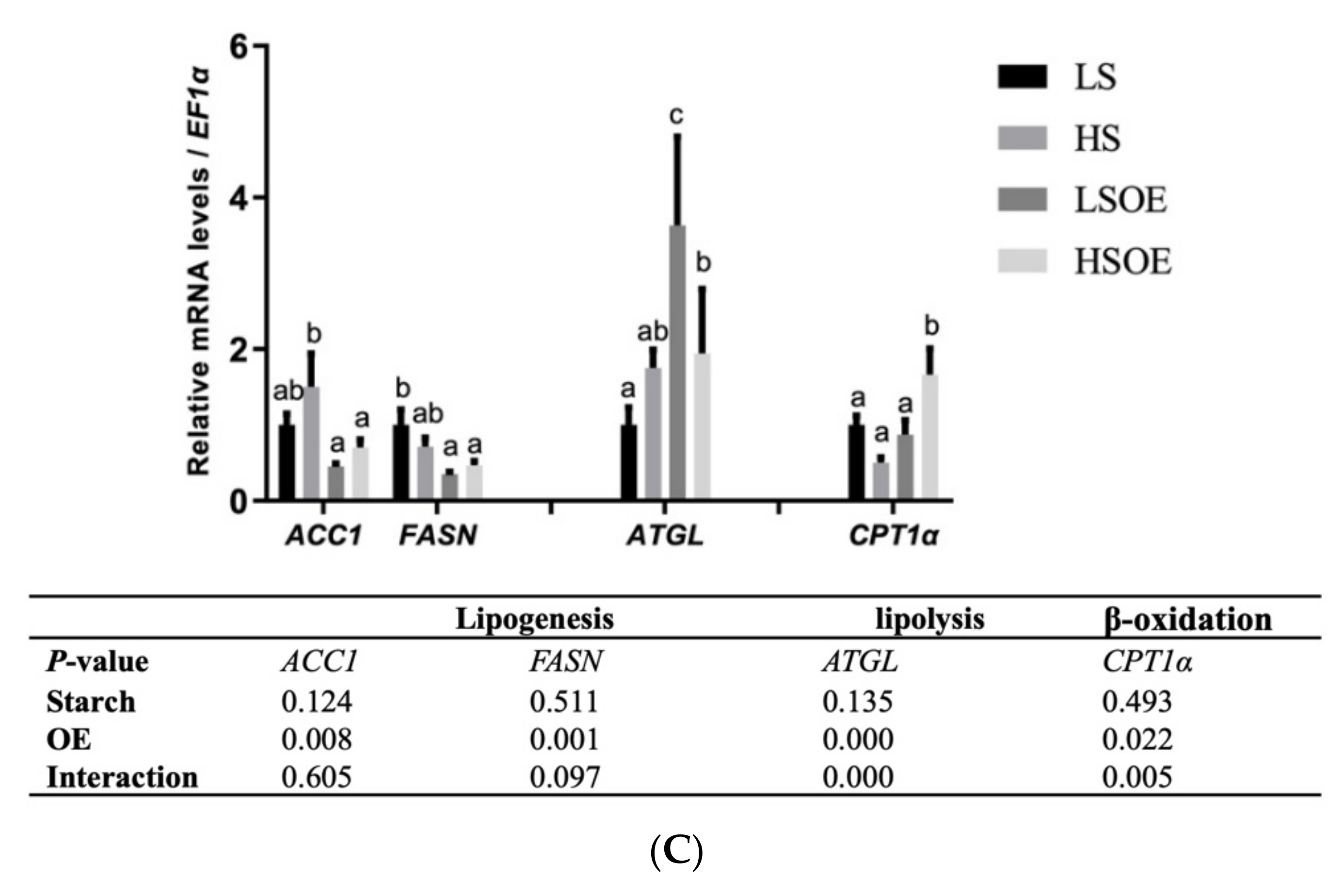
3.6. Hepatic Lipid Metabolism
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sørensen, M.; Morken, T.; Kosanovic, M.; Øverland, M. Pea and wheat starch possess different processing characteristics and affect physical quality and viscosity of extruded feed for Atlantic salmon. Aquac. Nutr. 2011, 17, e326–e336. [Google Scholar] [CrossRef]
- Lin, S.-M.; Shi, C.-M.; Mu, M.-M.; Chen, Y.-J.; Luo, L. Effect of high dietary starch levels on growth, hepatic glucose metabolism, oxidative status and immune response of Juvenile largemouth bass, Micropterus salmoides. Fish Shellfish Immunol. 2018, 78, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Liang, X.; Chen, P.; Wu, X.; Zheng, Y.; Luo, L.; Qin, Y.; Long, X.; Xue, M. Dietary supplementation of Grobiotic®-A increases short-term inflammatory responses and improves long-term growth performance and liver health in largemouth bass (Micropterus salmoides). Aquaculture 2018, 500, 327–337. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, L.; Chen, P.; Liang, X.; Cao, A.; Han, J.; Wu, X.; Zheng, Y.; Qin, Y.; Xue, M. Dietary Bile Acids Enhance Growth, and Alleviate Hepatic Fibrosis Induced by a High Starch Diet via AKT/FOXO1 and cAMP/AMPK/SREBP1 Pathway in Micropterus salmoides. Front. Physiol. 2019, 10, 1430. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Espinosa, C.D.; Abelilla, J.J.; Casas, G.A.; Lagos, L.V.; Lee, S.A.; Kwon, W.B.; Mathai, J.K.; Navarro, D.M.D.L.; Jaworski, N.W.; et al. Non-antibiotic feed additives in diets for pigs: A review. Anim. Nutr. 2018, 4, 113–125. [Google Scholar] [CrossRef]
- Bulfon, C.; Volpatti, D.; Galeotti, M. Current research on the use of plant-derived products in farmed fish. Aquac. Res. 2013, 46, 513–551. [Google Scholar] [CrossRef]
- Rong, J.; Han, Y.; Zha, S.; Tang, Y.; Shi, W.; Guan, X.; Du, X.; He, M.; Liu, G. Triterpene-enriched olive extract as an im-munopotentiator in Black Sea Bream (Acanthopagrus schlegelii). J. Ocean U. China 2020, 19, 428–438. [Google Scholar] [CrossRef]
- Sudjana, A.N.; D Orazio, C.; Ryan, V.; Rasool, N.; Ng, J.; Islam, N.; Riley, T.V.; Hammer, K.A. Antimicrobial activity of commercial Olea europaea (olive) leaf extract. Int. J. Antimicrob. Ag. 2009, 33, 461–463. [Google Scholar] [CrossRef]
- Baba, E.; Acar, Ü.; Yılmaz, S.; Zemheri, F.; Ergün, S. Dietary olive leaf (Olea europea L.) extract alters some immune gene expression levels and disease resistance to Yersinia ruckeri infection in rainbow trout Oncorhynchus mykiss. Fish Shellfish Immunol. 2018, 79, 28–33. [Google Scholar] [CrossRef]
- Zemheri-Navruz, F.; Acar, Ü.; Yılmaz, S. Dietary supplementation of olive leaf extract enhances growth performance, digestive enzyme activity and growth-related genes expression in common carp Cyprinus carpio. Gen. Comp. Endocrinol. 2020, 296, 113541. [Google Scholar] [CrossRef]
- Sánchez-Quesada, C.; López-Biedma, A.; Warleta, F.; Campos, M.; Beltrán, G.; Gaforio, J.J. Bioactive Properties of the Main Triterpenes Found in Olives, Virgin Olive Oil, and Leaves of Olea europaea. J. Agric. Food Chem. 2013, 61, 12173–12182. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Lehtonen, M.; Suuronen, T.; Kaarniranta, K.; Huuskonen, J. Terpenoids: Natural inhibitors of NF-κB signaling with anti-inflammatory and anticancer potential. Cell Mol. Life Sci. 2008, 65, 2979–2999. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, X.; Lian, C.; Ke, J.; Liu, J. Triterpenes and aromatic meroterpenoids with antioxidant activity and neuroprotective effects from Ganoderma lucidum. Molecules 2019, 24, 4353. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Wang, X.; Su, G.; Yue, J.; Sun, Y.; Cao, J.; Zhang, X.; Zhao, Y. The antioxidant and anti-hepatic fibrosis activities of acorns (Quercus liaotungensis) and their natural galloyl triterpenes. J. Funct. Foods 2018, 46, 567–578. [Google Scholar] [CrossRef]
- Yeh, Y.-T.; Cho, Y.-Y.; Hsieh, S.-C.; Chiang, A.-N. Chinese olive extract ameliorates hepatic lipid accumulation in vitro and in vivo by regulating lipid metabolism. Sci. Rep. 2018, 8, 1057. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, P.; Liang, X.F.; Han, J.; Wu, X.F.; Yang, Y.H.; Xue, M. Metabolic disorder induces fatty liver in Japanese seabass, Lateolabrax japonicas fed a full plant protein diet and regulated by cAMP-JNK/NF-kB caspase signal pathway. Fish Shellfish Immun. 2019, 90, 223–234. [Google Scholar] [CrossRef]
- Enes, P.; Panserat, S.; Kaushik, S.; Oliva-Teles, A. Nutritional regulation of hepatic glucose metabolism in fish. Fish Physiol. Biochem. 2008, 35, 519–539. [Google Scholar] [CrossRef]
- Hemre, G.-I.; Mommsen, T.P.; Krogdahl, A. Carbohydrates in fish nutrition: Effects on growth, glucose metabolism and hepatic enzymes. Aquac. Nutr. 2002, 8, 175–194. [Google Scholar] [CrossRef]
- Ma, H.-J.; Mou, M.-M.; Pu, D.-C.; Lin, S.-M.; Chen, Y.-J.; Luo, L. Effect of dietary starch level on growth, metabolism enzyme and oxidative status of Juvenile largemouth bass, Micropterus salmoides. Aquaculture 2018, 498, 482–487. [Google Scholar] [CrossRef]
- Zhang, Y.; Xie, S.; Wei, H.; Zheng, L.; Liu, Z.; Fang, H.; Xie, J.; Liao, S.; Tian, L.; Liu, Y.; et al. High dietary starch impaired growth performance, liver histology and hepatic glucose metabolism of Juvenile largemouth bass, Micropterus salmoides. Aquac. Nutr. 2020, 26, 1083–1095. [Google Scholar] [CrossRef]
- Matsumoto, T.; Terai, S.; Oishi, T.; Kuwashiro, S.; Fujisawa, K.; Yamamoto, N.; Fujita, Y.; Hamamoto, Y.; Furutani-Seiki, M.; Nishina, H.; et al. Medaka as a model for human nonalcoholic steatohepatitis. Dis. Model. Mech. 2010, 3, 431–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawood, M.A.; Koshio, S.; Esteban, M.A. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquac. 2017, 10, 950–974. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Alagawany, M.; Patra, A.K.; Kar, I.; Tiwari, R.; Dawood, M.A.; Dhama, K.; Abdel-Latif, H.M. The functionality of probiotics in aquaculture: An overview. Fish Shellfish Immunol. 2021, 117, 36–52. [Google Scholar] [CrossRef]
- Lieke, T.; Meinelt, T.; Hoseinifar, S.H.; Pan, B.; Straus, D.L.; Steinberg, C.E.W. Sustainable aquaculture requires environ-mental-friendly treatment strategies for fish diseases. Rev. Aquacult. 2020, 12, 943–965. [Google Scholar] [CrossRef] [Green Version]
- Reverter, M.; Bontemps, N.; Lecchini, D.; Banaigs, B.; Sasal, P. Use of plant extracts in fish aquaculture as an alternative to chemotherapy: Current status and future perspectives. Aquaculture 2014, 433, 50–61. [Google Scholar] [CrossRef]
- Nya, E.J.; Austin, B. Use of dietary ginger, Zingiber officinale Roscoe, as an immunostimulant to control Aeromonas hydrophila infections in rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish Dis. 2009, 32, 971–977. [Google Scholar] [CrossRef]
- Nya, E.J.; Austin, B. Use of garlic, Allium sativum, to control Aeromonas hydrophila infection in rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish Dis. 2009, 32, 963–970. [Google Scholar] [CrossRef]
- Goda, A. Effect of dietary ginseng herb (Ginsana® G115) supplementation on growth, feed utilization, and hemato-logical indices of Nile tilapia, Oreochromis niloticus (L.), fingerlings. J. World Aquacult. Soc. 2008, 39, 205–214. [Google Scholar] [CrossRef]
- Aanyu, M.; Betancor, M.B.; Monroig, O. Effects of dietary limonene and thymol on the growth and nutritional physiology of Nile tilapia (Oreochromis niloticus). Aquaculture 2018, 488, 217–226. [Google Scholar] [CrossRef]
- Cho, S.H.; Lee, S.-M.; Park, B.H.; Ji, S.-C.; Lee, J.; Bae, J.; Oh, S.-Y. Effect of dietary inclusion of various sources of green tea on growth, body composition and blood chemistry of the juvenile olive flounder, Paralichthys olivaceus. Fish Physiol. Biochem. 2006, 33, 49–57. [Google Scholar] [CrossRef]
- Wang, Y.; Branicky, R.; Noe, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of Malondial-dehyde and 4-Hydroxy-2-Nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Song, M.; Shi, C.; Lin, S.; Chen, Y.; Shen, H.; Luo, L. Effect of starch sources on growth, hepatic glucose metabolism and an-tioxidant capacity in juvenile largemouth bass, Micropterus salmoides. Aquaculture 2018, 490, 355–361. [Google Scholar] [CrossRef]
- Singh, G.B.; Singh, S.; Bani, S.; Gupta, B.D.; Banerjee, S.K. Anti-inflammatory activity of oleanolic acid in rats and mice. J. Pharm. Pharmacol. 1992, 44, 456–458. [Google Scholar] [CrossRef] [PubMed]
- Marquez-Martin, A.; De La Puerta, R.; Fernandez-Arche, A.; Ruiz-Gutierrez, V.; Yaqoob, P. Modulation of cytokine secretion by pentacyclic triterpenes from olive pomace oil in human mononuclear cells. Cytokine 2006, 36, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Gholamhosseini, A.; Kheirandish, M.R.; Shiry, N.; Akhlaghi, M.; Soltanian, S.; Roshanpour, H.; Banaee, M. Use of a methanolic olive leaf extract (Olea europaea) against white spot virus syndrome in Penaeus vannamei: Comparing the biochemical, hematological and immunological changes. Aquaculture 2020, 528. [Google Scholar] [CrossRef]
- Song, C.; Liu, B.; Xu, P.; Xie, J.; Ge, X.; Zhou, Q.; Sun, C.; Zhang, H.; Shan, F.; Yang, Z. Oxidized fish oil injury stress in Meg-alobrama amblycephala: Evaluated by growth, intestinal physiology, and transcriptome-based PI3K-Akt/NF-κB/TCR inflam-matory signaling. Fish Shellfish Immun. 2018, 81, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Guo, Z.; Zhao, Y.; Wang, Q.; Gao, Y.; Yu, T.; Chen, Y.; Chen, X.; Wang, G. L-carnitine regulated Nrf2/Keap1 activation in vitro and in vivo and protected oxidized fish oil-induced inflammation response by inhibiting the NF-κB signaling pathway in Rhynchocypris lagowski Dybowski. Fish Shellfish Immun. 2019, 93, 1100–1110. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Yang, E.-J.; Ku, S.-K.; Song, K.-S.; Bae, J.-S. Anti-inflammatory Effects of Oleanolic Acid on LPS-Induced Inflammation In Vitro and In Vivo. Inflammation 2012, 36, 94–102. [Google Scholar] [CrossRef]
- Yang, E.-J.; Lee, W.; Ku, S.-K.; Song, K.-S.; Bae, J.-S. Anti-inflammatory activities of oleanolic acid on HMGB1 activated HUVECs. Food Chem. Toxicol. 2012, 50, 1288–1294. [Google Scholar] [CrossRef]
- Biswas, D.K.; Shi, Q.; Baily, S.; Strickland, I.; Ghosh, S.; Pardee, A.B.; Iglehart, J.D. NF-κB activation in human breast cancer specimens and its role in cell proliferation and apoptosis. Proc. Natl. Acad. Sci. USA 2004, 101, 10137–10142. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Liu, H.T. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yao, T.; Pini, M.; Zhou, Z.; Fantuzzi, G.; Song, Z. Betaine improved adipose tissue function in mice fed a high-fat diet: A mechanism for hepatoprotective effect of betaine in nonalcoholic fatty liver disease. Am. J. Physiol. Liver Physiol. 2010, 298, G634–G642. [Google Scholar] [CrossRef] [Green Version]
- Das, M.; Garlick, D.S.; Greiner, D.L.; Davis, R.J. The role of JNK in the development of hepatocellular carcinoma. Genes Dev. 2011, 25, 634–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pai, W.Y.; Hsu, C.C.; Lai, C.Y.; Chang, T.Z.; Tsai, Y.L.; Her, G.M. Cannabinoid receptor 1 promotes hepatic lipid accumulation and lipotoxicity through the induction of SREBP-1c expression in zebrafish. Transgenic Res. 2013, 22, 823–838. [Google Scholar] [CrossRef]
- Kohjima, M.; Enjoji, M.; Higuchi, N.; Kato, M.; Kotoh, K.; Yoshimoto, T.; Fujino, T.; Yada, M.; Yada, R.; Harada, N.; et al. Re-evaluation of fatty acid metabolism-related gene expression in nonalcoholic fatty liver disease. Int. J. Mol. Med. 2007, 20, 351–358. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Xu, S.; Mihaylova, M.M.; Zheng, B.; Hou, X.; Jiang, B.; Park, O.; Luo, Z.; Lefai, E.; Shyy, J.Y.-J.; et al. AMPK Phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin-resistant mice. Cell Metab. 2011, 13, 376–388. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Lu, Y.-F.; Wu, Q.; Xu, S.-F.; Shi, F.-G.; Klaassen, C.D. Oleanolic acid reprograms the liver to protect against hepatotoxicants, but is hepatotoxic at high doses. Liver Int. 2018, 39, 427–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | LS | HS | LSOE | HSOE |
|---|---|---|---|---|
| Fish meal a | 30.0 | 30.0 | 30.0 | 30.0 |
| Cottonseed protein concentrate a | 23.5 | 22.6 | 23.4 | 22.5 |
| Microbial protein a | 4.0 | 4.0 | 4.0 | 4.0 |
| Tapioca starch | 5.0 | 5.0 | 5.0 | 5.0 |
| Wheat flour | 9.0 | 18.0 | 9.0 | 18.0 |
| Wheat gluten meal | 4.0 | 4.0 | 4.0 | 4.0 |
| Soybean meal a | 2.0 | - | 2.0 | - |
| Spay-dried blood cell powder | 4.0 | 4.0 | 4.0 | 4.0 |
| α-cellulose | 4.6 | - | 4.6 | - |
| Ca(H2PO4)2 | 1.7 | 1.7 | 1.7 | 1.7 |
| Lecithin oil | 2.0 | 2.0 | 2.0 | 2.0 |
| Fish oil | 3.5 | 3.5 | 3.5 | 3.5 |
| Soybean oil | 3.5 | 3.5 | 3.5 | 3.5 |
| Vitamin and mineral premix b | 1.4 | 1.4 | 1.4 | 1.4 |
| Kelp powder | 1.5 | 0 | 1.5 | 0 |
| L-Thr | 0.1 | 0.1 | 0.1 | 0.1 |
| DL-Met | 0.2 | 0.2 | 0.2 | 0.2 |
| Olive extract | 0 | 0 | 0.125 | 0.125 |
| Total | 100 | 100 | 100 | 100 |
| Analyzed chemical composition (dry matter basis %) | ||||
| Moisture | 6.10 | 7.43 | 7.25 | 7.34 |
| Crude protein | 50.83 | 51.15 | 51.17 | 51.11 |
| Crude lipid | 12.36 | 12.33 | 12.44 | 12.41 |
| Crude ash | 10.08 | 10.04 | 9.84 | 9.82 |
| Starch c | 9.00 | 14.40 | 9.00 | 14.40 |
| Gross energy (MJ/Kg) | 20.45 | 20.15 | 20.49 | 20.27 |
| Genes | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Tm (°C) | E-Values (%) | Accession Number |
|---|---|---|---|---|---|
| EF1α | TGCTGCTGGTGTTGGTGAGTT | TTCTGGCTGTAAGGGGGCTC | 60.4 | 102.8 | 119901934 |
| ACC1 | ATCCCTCTTTGCCACTGTTG | GAGGTGATGTTGCTCGCATA | 57.5 | 102.2 | 119896388 |
| FASN | TGTGGTGCTGAACTCTCTGG | CATGCCTAGTGGGGAGTTGT | 57.5 | 102.1 | 119915567 |
| ATGL | CCATGATGCTCCCCTACACT | GGCAGATACACTTCGGGAAA | 58 | 99.1 | 119893301 |
| CPT1α | CATGGAAAGCCAGCCTTTAG | GAGCACCAGACACGCTAACA | 60.0 | 98.8 | 119893292 |
| TNFα | CTTCGTCTACAGCCAGGCATCG | TTTGGCACACCGACCTCACC | 63 | 105.7 | 119906688 |
| IL1β | CGTGACTGACAGCAAAAAGAG | GATGCCCAGAGCCACAGTTC | 59.4 | 101.3 | 119914255 |
| TGFβ1 | GCTCAAAGAGAGCGAGGATG | TCCTCTACCATTCGCAATCC | 59 | 95.6 | 119882881 |
| IL10 | CGGCACAGAAATCCCAGAGC | CAGCAGGCTCACAAAATAAACA | 62.1 | 113.6 | 119885912 |
| SREBP1 | AGTCTGAGCTACAGCGACAAGG | TCATCACCAACAGGAGGTCACA | 61 | 98.1 | 119888831 |
| OE (%) | Level of Dietary Starch (%) | OE Level | P Values | ||||
|---|---|---|---|---|---|---|---|
| 9.0 | 14.4 | OE Level | Starch Level | Interaction | |||
| IBW 1 | - | 35.98 ± 0.21 | - | - | - | - | |
| FBW 2 | 0 | 99.98 ± 1.68 | 97.66 ± 1.69 | 98.82 ± 1.18 | 0.223 | 0.002 | 0.008 |
| 0.125 | 109.59 ± 1.77 | 93.34 ± 2.68 | 101.46 ± 3.91 | ||||
| starch level | 104.78 ± 2.41 B | 95.50 ± 1.72 A | |||||
| SR 3 | 0 | 96.67 ± 1.67 | 96.67 ± 1.67 | 96.67 ± 1.05 | 1.000 | 1.000 | 1.000 |
| 0.125 | 96.67 ± 1.67 | 96.67 ± 1.67 | 96.67 ± 1.05 | ||||
| starch level | 96.67 ± 1.05 | 96.67 ± 1.05 | |||||
| SGR 4 | 0 | 1.94 ± 0.03 | 1.91 ± 0.03 | 1.93 ± 0.02 | 0.272 | 0.002 | 0.008 |
| 0.125 | 2.12 ± 0.03 | 1.82 ± 0.06 | 1.97 ± 0.07 | ||||
| starch level | 2.03 ± 0.44 B | 1.86 ± 0.04 A | |||||
| FCR 5 | 0 | 1.01 ± 0.01 | 0.96 ± 0.01 | 0.98 ± 0.01 b | 0.002 | 0.005 | 0.327 |
| 0.125 | 0.95 ± 0.01 | 0.93 ± 0.01 | 0.94 ± 0.01 a | ||||
| starch level | 0.98 ± 0.01B | 0.94 ± 0.01 A | |||||
| FR 6 | 0 | 0.44 ± 0.01 | 0.42 ± 0.01 | 0.43 ± 0.01 | 0.108 | 0.000 | 0.010 |
| 0.125 | 0.46 ± 0.01 | 0.39 ± 0.01 | 0.42 ± 0.02 | ||||
| starch level | 0.45 ± 0.00 B | 0.40 ± 0.01 A | |||||
| OE (%) | Level of Dietary Starch (%) | OE Level | P Values | ||||
|---|---|---|---|---|---|---|---|
| 9.0 | 14.4 | OE Level | Starch Level | Interaction | |||
| CF 1 | 0 | 1.96 ± 0.10 | 1.75 ± 0.03 | 1.85 ± 0.06 b | 0.004 | 0.066 | 0.054 |
| 0.125 | 1.68 ± 0.02 | 1.68 ± 0.03 | 1.68 ± 0.02 a | ||||
| starch level | 1.82 ± 0.06 | 1.72 ± 0.02 | |||||
| VSI 2 | 0 | 7.14 ± 0.10 | 7.60 ± 0.23 | 7.37 ± 0.13 | 0.142 | 0.001 | 0.110 |
| 0.125 | 7.01 ± 0.22 | 8.30 ± 0.29 | 7.66 ± 0.22 | ||||
| starch level | 7.07 ± 0.12 A | 7.95 ± 0.20 B | |||||
| HIS 3 | 0 | 1.72 ± 0.15 | 2.54 ± 0.14 | 2.13 ± 0.13 a | 0.007 | 0.000 | 0.842 |
| 0.125 | 2.05 ± 0.12 | 3.04 ± 0.22 | 2.57 ± 0.17 b | ||||
| starch level | 1.88 ± 0.10 A | 2.79 ± 0.14 B | |||||
| VAI 4 | 0 | 1.46 ± 0.07 | 1.74 ± 0.20 | 1.60 ± 0.11 | 0.115 | 0.348 | 0.489 |
| 0.125 | 1.85 ± 0.17 | 1.90 ± 0.21 | 1.88 ± 0.13 | ||||
| starch level | 1.66 ± 0.10 | 1.82 ± 0.14 | |||||
| OE (%) | Level of Dietary Starch (%) | OE Level | P Values | ||||
|---|---|---|---|---|---|---|---|
| 9.0 | 14.4 | OE Level | Starch Level | Interaction | |||
| Moisture | 0 | 73.39 ± 1.14 | 70.60 ± 0.18 | 72.00 ± 0.81 b | 0.004 | 0.066 | 0.054 |
| 0.125 | 69.87 ± 0.49 | 69.87 ± 0.49 | 69.74 ± 0.26 a | ||||
| starch level | 71.63 ± 0.96 | 70.11 ± 0.27 | |||||
| Crude protein | 0 | 16.58 ± 0.15 | 16.94 ± 0.06 | 16.76 ± 0.11 | 0.142 | 0.001 | 0.110 |
| 0.125 | 16.70 ± 0.15 | 16.13 ± 0.05 | 16.42 ± 0.15 | ||||
| starch level | 16.64 ± 0.10 | 16.54 ± 0.19 | |||||
| Crude lipid | 0 | 5.37 ± 0.84 | 7.93 ± 0.22 | 6.65 ± 0.69 a | 0.007 | 0.000 | 0.842 |
| 0.125 | 9.24 ± 0.36 | 9.16 ± 0.23 | 9.20 ± 0.19 b | ||||
| starch level | 7.31 ± 0.96 | 8.54 ± 0.31 | |||||
| OE(%) | Level of Dietary Starch (%) | OE Level | P Values | ||||
|---|---|---|---|---|---|---|---|
| 9.0 | 14.4 | OE Level | Starch Level | Interaction | |||
| Glucose (mM/L) | 0 | 4.82 ± 0.40 | 4.98 ± 0.70 | 4.90 ± 0.30 | 0.097 | 0.053 | 0.043 |
| 0.125 | 5.05 ± 0.37 | 3.19 ± 0.31 | 4.12 ± 0.33 | ||||
| Starch level | 4.93 ± 0.26 | 4.08 ± 0.36 | |||||
| TG (mM/L) | 0 | 4.31 ± 0.43 | 6.59 ± 0.98 | 5.45 ± 0.60 | 0.895 | 0.598 | 0.038 |
| 0.125 | 5.82 ± 0.77 | 4.68 ± 0.54 | 5.25 ± 0.48 | ||||
| starch level | 5.07 ± 0.47 | 5.64 ± 0.59 | |||||
| TC (mM/L) | 0 | 6.34 ± 0.29 | 7.13 ± 0.48 | 6.74 ± 0.29a | 0.000 | 0.587 | 0.167 |
| 0.125 | 8.73 ± 0.57 | 8.33 ± 0.40 | 8.53 ± 0.34b | ||||
| starch level | 7.53 ± 0.44 | 7.73 ± 0.34 | |||||
| HDL-C (mM/L) | 0 | 1.47 ± 0.23 | 1.20 ± 0.21 | 1.33 ± 0.15a | 0.013 | 0.742 | 0.511 |
| 0.125 | 1.86 ± 0.29 | 2.05 ± 0.25 | 1.95 ± 0.18b | ||||
| starch level | 1.66 ± 0.18 | 1.59 ± 0.19 | |||||
| HDL-C/TC | 0 | 0.23 ± 0.03 | 0.17 ± 0.03 | 0.20 ± 0.02 | 0.363 | 0.522 | 0.190 |
| 0.125 | 0.21 ± 0.03 | 0.22 ± 0.03 | 0.21 ± 0.02 | ||||
| starch level | 0.22 ± 0.02 | 0.19 ± 0.02 | |||||
| LDL-C (mM/L) | 0 | 1.77 ± 0.22 | 2.56 ± 0.21 | 2.16 ± 0.18 | 0.591 | 0.059 | 0.100 |
| 0.125 | 2.22 ± 0.20 | 2.26 ± 0.18 | 2.24 ± 0.13 | ||||
| starch level | 2.00 ± 0.16 | 2.41 ± 0.14 | |||||
| LDL-C/TC | 0 | 0.28 ± 0.03 | 0.37 ± 0.03 | 0.32 ± 0.03b | 0.021 | 0.154 | 0.109 |
| 0.125 | 0.26 ± 0.02 | 0.25 ± 0.02 | 0.25 ± 0.01a | ||||
| starch level | 0.27 ± 0.02 | 0.31 ± 0.02 | |||||
| OE (%) | Level of Dietary Starch (%) | OE Level | P Values | ||||
|---|---|---|---|---|---|---|---|
| 9.0 | 14.4 | OE Level | Starch Level | Interaction | |||
| AKP (U/L) | 0 | 41.15 ± 3.51 | 92.49 ± 8.46 | 66.82 ± 7.97 | 0.551 | 0.000 | 0.001 |
| 0.125 | 69.61 ± 9.19 | 70.34 ± 5.91 | 69.98 ± 9.86 | ||||
| starch level | 55.38 ± 6.01 A | 81.42 ± 5.75 B | |||||
| AST (U/L) | 0 | 4.96 ± 0.58 | 14.58 ± 2.38 | 9.86 ± 2.30 | 0.317 | 0.000 | 0.087 |
| 0.125 | 6.22 ± 0.85 | 10.37 ± 1.11 | 8.87 ± 1.17 | ||||
| starch level | 5.33 ± 1.13 A | 12.89 ± 1.77 B | |||||
| ALT (U/L) | 0 | 4.55 ± 0.51 | 15.86 ± 1.65 | 9.99 ± 1.85 b | 0.022 | 0.000 | 0.000 |
| 0.125 | 6.98 ± 0.91 | 8.50 ± 0.74 | 8.14 ± 1.00 a | ||||
| starch level | 5.73 ± 1.02 A | 12.25 ± 1.45 B | |||||
| OE (%) | Level of Dietary Starch (%) | OE Level | P Values | ||||
|---|---|---|---|---|---|---|---|
| 9.0 | 14.4 | OE Level | Starch Level | Interaction | |||
| ROS (U/mg prot) | 0 | 64.42 ± 4.61 | 86.41 ± 4.42 | 75.41 ± 4.20 | 0.105 | 0.000 | 0.101 |
| 0.125 | 64.33 ± 3.73 | 102.84 ± 6.37 | 83.59 ± 6.12 | ||||
| starch level | 64.37 ± 2.86 A | 94.62 ± 4.30 B | |||||
| T-AOC (µM/g prot) | 0 | 69.12 ± 5.80 | 95.70 ± 8.69 | 82.41 ± 6.10 | 0.749 | 0.185 | 0.009 |
| 0.125 | 91.01 ± 7.21 | 81.60 ± 6.37 | 86.31 ± 4.80 | ||||
| starch level | 80.07 ± 5.29 | 88.65 ± 5.51 | |||||
| CAT (U/mg prot) | 0 | 49.61 ± 3.02 | 47.81 ± 4.39 | 48.71 ± 2.58 b | 0.000 | 0.000 | 0.000 |
| 0.125 | 49.83 ± 3.70 | 8.17 ± 1.48 | 29.00 ± 5.71 a | ||||
| starch level | 49.72 ± 2.31 B | 27.99 ± 5.59A | |||||
| GSH-Px (U/ug prot) | 0 | 4.14 ± 0.70 | 4.57 ± 0.74 | 4.35 ± 0.49 | 0.137 | 0.815 | 0.642 |
| 0.125 | 4.87 ± 0.54 | 5.24 ± 0.42 | 5.06 ± 0.33 | ||||
| starch level | 4.50 ± 0.44 | 4.90 ± 0.42 | |||||
| SOD (U/mg prot) | 0 | 195.90 ± 11.13 | 192.83 ± 11.57 | 194.37 ± 7.76 b | 0.004 | 0.038 | 0.104 |
| 0.125 | 185.02 ± 9.71 | 144.04 ± 7.72 | 164.53 ± 7.99 a | ||||
| starch level | 190.46 ± 7.27 B | 168.44 ± 9.21A | |||||
| MDA (nM/mg prot) | 0 | 4.48 ± 0.49 | 2.70 ± 0.85 | 3.53 ± 0.54 b | |||
| 0.125 | 1.22 ± 0.16 | 2.17 ± 0.45 | 1.70 ± 0.26 a | 0.002 | 0.452 | 0.020 | |
| starch level | 2.75 ± 0.49 | 2.43 ± 0.47 | |||||
| SOD/MDA | 0 | 47.5 ± 6.08 | 103.8 ± 21.79 | 77.52 ± 13.74a | 0.004 | 0.819 | 0.002 |
| 0.125 | 166.93 ± 19.51 | 98 ± 23.14 | 132.47 ± 17.12b | ||||
| starch level | 111.2 ± 19.00 | 100.90 ± 15.37 | |||||
| OE (%) | Level of Dietary Starch (%) | OE Level | P Values | ||||
|---|---|---|---|---|---|---|---|
| 9.0 | 14.4 | OE Level | Starch Level | Interaction | |||
| TG (mM/g prot) | 0 | 0.39 ± 0.06 | 0.32 ± 0.05 | 0.36 ± 0.05 b | 0.010 | 0.838 | 0.197 |
| 0.125 | 0.20 ± 0.03 | 0.25 ± 0.05 | 0.22 ± 0.03 a | ||||
| starch level | 0.30 ± 0.04 | 0.29 ± 0.05 | |||||
| TC (mM/g prot) | 0 | 0.16 ± 0.02 | 0.15 ± 0.01 | 0.16 ± 0.01 | 0.426 | 0.286 | 0.041 |
| 0.125 | 0.12 ± 0.01 | 0.17 ± 0.01 | 0.15 ± 0.01 | ||||
| starch level | 0.14 ± 0.01 | 0.16 ± 0.01 | |||||
| LDL-C (µM/g prot) | 0 | 38.03 ± 3.01 | 42.67 ± 3.04 | 40.35 ± 2.15 b | 0.010 | 0.056 | 0.426 |
| 0.125 | 26.06 ± 2.29 | 35.20 ± 3.30 | 30.63 ± 2.27 a | ||||
| starch level | 32.04 ± 2.39 | 38.94 ± 2.37 | |||||
| LDL-C/TC | 0 | 0.24 ± 0.02 | 0.21 ± 0.02 | 0.26 ± 0.17 | 0.050 | 0.504 | 0.273 |
| 0.125 | 0.22 ± 0.02 | 0.21 ± 0.22 | 0.22 ± 0.13 | ||||
| starch level | 0.23 ± 0.02 | 0.25 ± 0.02 | |||||
| TBA (µM/g prot) | 0 | 3.14 ± 0.21 | 3.47 ± 0.91 | 3.29 ± 0.42 b | 0.001 | 0.917 | 0.551 |
| 0.125 | 1.66 ± 0.14 | 1.43 ± 0.14 | 1.54 ± 0.10 a | ||||
| starch level | 2.45 ± 0.24 | 2.45 ± 0.52 | |||||
| Liver lipid (%) | 0 | 1.99 ± 0.01 | 2.01 ± 0.16 | 2.03 ± 0.06 | 0.606 | 0.007 | 0.006 |
| 0.125 | 1.66 ± 0.10 | 2.27 ± 0.04 | 1.95 ± 0.15 | ||||
| starch level | 1.77 ± 0.10A | 2.16 ± 0.07B | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, X.; Chen, P.; Wu, X.; Xing, S.; Morais, S.; He, M.; Gu, X.; Xue, M. Effects of High Starch and Supplementation of an Olive Extract on the Growth Performance, Hepatic Antioxidant Capacity and Lipid Metabolism of Largemouth Bass (Micropterus salmoides). Antioxidants 2022, 11, 577. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11030577
Liang X, Chen P, Wu X, Xing S, Morais S, He M, Gu X, Xue M. Effects of High Starch and Supplementation of an Olive Extract on the Growth Performance, Hepatic Antioxidant Capacity and Lipid Metabolism of Largemouth Bass (Micropterus salmoides). Antioxidants. 2022; 11(3):577. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11030577
Chicago/Turabian StyleLiang, Xiaofang, Pei Chen, Xiaoliang Wu, Shujuan Xing, Sofia Morais, Maolong He, Xu Gu, and Min Xue. 2022. "Effects of High Starch and Supplementation of an Olive Extract on the Growth Performance, Hepatic Antioxidant Capacity and Lipid Metabolism of Largemouth Bass (Micropterus salmoides)" Antioxidants 11, no. 3: 577. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11030577